
ABSTRACT
A long history of geological research on the island of Gotland, Sweden, has resulted in a detailed biostratigraphy based on conodonts for the Gotland sedimentary succession, but the relation between the Hamra and Sundre formations, the youngest strata on southern Gotland, has remained poorly resolved. These formations have also remained relatively poorly described in terms of vertebrates compared to other parts of the succession. A survey of museum collections and newly sampled material reveal that the taxonomical compositions and richness of vertebrate faunas remain similar compared to the underlying Burgsvik Sandstone and Oolite members. However, the relative abundance of the respective groups changes: Paralogania ludlowiensis and rare osteostracan remains of Tahulaspis sp. only occur in samples from the lower Hamra Formation, while Thelodus sculptilis becomes more common in samples from Sundre Formation. Conodont and isotope data give support to previous suggestions that the Hamra and Sundre formations may be largely isochronous, and it is possible that the observed differences in vertebrate faunas reflect changes in depositional setting. This interval on Gotland has been suggested to represent a hiatus in the East Baltic sections, where younger strata show an increased importance of acanthodians in the vertebrate faunas. Gotland could therefore give insights into the early stages of this diversification of gnathostomes during late Silurian times. However, this has to be done in combination with data from other areas, as well as with a review and revision of the scale-based taxonomy of Silurian acanthodians from the Baltic Basin.
Introduction
The Silurian strata of Gotland, Sweden, display rich vertebrate assemblages throughout most of the sequence (e.g., Fredholm Citation1988a, Citation1988b, Citation1989, Citation1990; Eriksson et al. Citation2009; Jarochowska et al. Citation2016a; Bremer Citation2017). The most extensive works on Gotland vertebrates to date are those by Fredholm (Citation1988a, Citation1988b, Citation1989, Citation1990), but the only report on vertebrates from the youngest parts of the Gotland stratigraphy was made by Fredholm (Citation1989). Blom et al. (Citation2002) reviewed and revised some earlier reports of anaspids from Gotland and reported scales of Septentrionia mucronata Blom et al. Citation2002, Liivilepis curvata Blom et al. Citation2002, and Hoburgilepis papillata Blom et al. Citation2002 from the “Burgsvik/Hamra beds” at Hoburgen 2 and “Hamra beds” at Hoburgen 3 localities. Later, Eriksson et al. (Citation2009) described two samples from Hamra Formation that contained remains of thelodonts, acanthodians, and osteostracans. Besides these works, reports on the vertebrate assemblages of the youngest strata of Gotland have been scarce and the faunas have remained relatively poorly understood.
It has previously been demonstrated that gnathostomes, in particular acanthodians, become an increasingly important component among the fossils recovered from rocks approaching end-Ludlow in age on both Gotland (Fredholm Citation1988a, Citation1988b) and in the East Baltic (Kaljo & Märss Citation1991). The interval of the Hamra and Sundre formations on Gotland is most likely represented by hiatuses in the well-studied sections of the East Baltic (Märss & Männik Citation2013), making this interval important for increasing our understanding of the faunal changes during this time. However, the scale-based taxonomy of Silurian acanthodians from the Baltic Basin, which was developed in a series of works by Vergoossen (Citation1997, Citation1999a, Citation1999b, Citation1999c, Citation2000, Citation2002a, Citation2002b, Citation2002c, Citation2003a, Citation2003b, Citation2004) and Valiukevičius (Citation1998, Citation2003a, Citation2003b, Citation2004a, Citation2004b), is still in need of review and revision.
In this study, we present previously unpublished material stored in the Palaeontological collections of the Museum of Evolution at Uppsala University and Naturhistoriska Riksmuseet in Stockholm, Sweden, as well as the Department of Geology at Tallinn University of Technology, Estonia. We also present newly sampled material from the youngest strata on Gotland with the aim of getting a better picture of vertebrate diversity in the latest Ludlow of the Baltic Basin. The scale-based taxonomy of acanthodians from the Silurian of the Baltic region is also discussed briefly. These new samples, in conjunction with access to the extensive collection of Lennart Jeppsson (1940–2015) at the NRM, will also enable us to figure and give thorough accounts of conodont distribution in this part of the Gotland sequence. The comparison of the distribution of both conodonts and vertebrates, especially their co-occurrence, allows reconstruction of more reliable biostratigraphical schemes.
Institutional abbreviations Swedish Museum of Natural History, Palaeozoological collections, Stockholm (Sweden) – NRM-PZ; Palaeontological collections, Museum of Evolution, Uppsala University, Uppsala (Sweden) – PMU; Tallinn University of Technology, Tallinn (Estonia) – TalTech.
Geological setting
The youngest part of the Gotland stratigraphy () is divided into the Hamra and Sundre formations (Erlström et al. Citation2009). There is an unconformity surface between the Hamra Formation and the underlying Burgsvik Oolite Member, which can be more or less planar as in the Hoburgen area, or locally eroded down into the oolite as in the area around Husryggen (Eriksson & Calner Citation2008). The Hamra Formation has historically been subdivided into the informal units a, b and c (see Laufeld Citation1974) that are represented by oncoidal limestone (Jeppsson et al. Citation1994; Erlström et al. Citation2009), argillaceous limestone interlayered by thin layers of marl (Jeppsson et al. Citation1994; Eriksson & Calner Citation2008; Erlström et al. Citation2009), and bioherms associated with coarse crinoidal limestone (Jeppsson et al. Citation1994; Samtleben et al. Citation1996) respectively. These units generally pass into micritic and argillaceous limestones toward the north-east (Samtleben et al. Citation1996; Erlström et al. Citation2009), but they can display large variation in facies along their strike (Eriksson & Calner Citation2008).
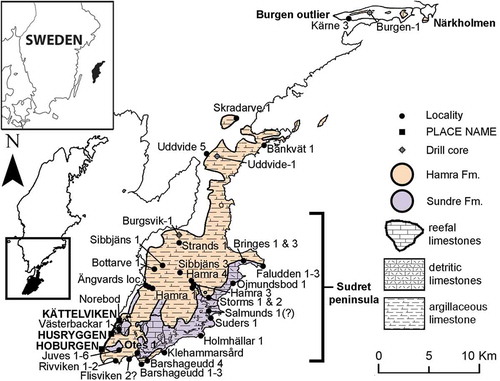
Figure 1. Map of Sudret peninsula in the south of Gotland with the Hamra Formation (peach) and Sundre Formation (purple), as well as the geographical areas (capital letters), localities, and positions of drill-cores discussed in the text. The geographical extent of the formations and their lithologies are based on Eriksson and Calner (Citation2005) and data from the Geological Survey of Sweden (SGU).
The Sundre Formation is mainly composed of large, stromatoporoid-rich bioherms and biostromes that are often continuous with bioherms of the Hamra Formation (Erlström et al. Citation2009). These are associated with coarse grained, crinoidal limestones that are more dominant to the northeast where back-reef sediments also occur (Samtleben et al. Citation1996; Erlström et al. Citation2009). In some southern areas, the reefs develop into atoll-like semi-circles, termed “faros reefs” by Samtleben et al. (Citation2000). These reefs alternate with crinoidal limestones and coarse, sometimes conglomeratic, grainstones with angular stromatopore-bioclasts (Samtleben et al. Citation2000). The reader is referred to the references above and Jeppsson (Citation2005b, p. 280) for detailed lithological descriptions.
Despite the previous subdivisions of the Hamra and Sundre formations based on lithology, only informal biostratigraphic subdivisions have been done (Jeppsson Citation1983, Citation2005b; Jeppsson et al. Citation2006). An early subdivision (Jeppsson Citation1983) followed the informal lithostratigraphic units of Hede (Citation1921, Citation1925, Citation1960). In this subdivision, Hamra a was distinguished by Hindeodella steinhornensis Ziegler Citation1956 and H. st. scanica Jeppsson Citation1974 (here, “Ozarkodina eosteinhornensis”), Ligonodina excavata novoexcavata Jeppsson Citation1972 (here, Oulodus novoexcavatus), and Hindeodella (here, Ozarkodina) confluens Branson and Mehl Citation1933. Hamra b was characterized by Jeppsson (Citation1983) based on conodonts from Bankvät 1 and Strands 1, which contained H. steinhornensis, H. wimani Jeppsson Citation1974 (here, Ozarkodina wimani), “H. snajdri crispa” and Li. (Oulodus) elegans Walliser Citation1964. Additionally, Belodella spp., Pseudooneotodus spp., Dapsilodus? spp., and Li. (Oulodus) excavata? Jeppsson Citation1972 were found. Panderodus equicostatus Rhodes Citation1953 and H. (Wurmiella) excavata Branson and Mehl Citation1933 were found at Kättelviken 5 less than 5 m above the base of the Hamra Formation. Based on the sections at Juves, no substantial differences in conodont composition were found in the Hamra c and Sundre units by Jeppsson (Citation1983). Moreover, Jeppsson (Citation2005b) suggested that the lithostratigraphic division of Hamra a, b, c, and the Sundre Formation does not reflect age differences. However, the Hamra Formation was assigned by Jeppsson et al. (Citation2006) to the upper part of the Oz. snajdri Biozone and the lower part of the Sundre Formation to the Oz. crispa Biozone. Furthermore. Jeppsson et al. (Citation2006) introduced informal divisions of the snajdri Biozone and according to them Oz. remscheidensis (here, Zieglerodina remscheidensis) is absent in the lower part of the Hamra Formation, while its early form appears in its upper part. Even more detailed informal divisions are to be found in Jeppsson’s collection and are reconstructed in the present work based on his annotations on the samples.
The contact to the underlying Burgsvik Oolite Member reflects erosion, but subsequent flooding is evident by the overlying marginal marine sediments. Eriksson and Calner (Citation2008) interpreted the sediments of the lower Hamra Formation as representing a beach barrier system deposited in a sheltered lagoon with localized channel flow, effectively reflecting different depositional environments within a very shallow subtidal setting. The overlying unit b suggests a relative deepening below storm wave base, and the large reef bodies of unit c and the following Sundre Formation could reflect the establishment of a large barrier reef in the area (Samtleben et al. Citation1996; Eriksson & Calner Citation2008). The Hamra and Sundre formations have collectively been interpreted as a highstand system tract (Eriksson & Calner Citation2008), or as transgression followed by stillstand and subsequently a continued rise in sea-level (Kozłowski & Munnecke Citation2010).
Materials and methods
The extensive collection of Lennart Jeppsson stored at the NRM has many samples from the Hamra and Sundre formations that mainly contain conodont material (see next section). These samples have been documented in the Paleobiology Database (www.paleobiodb.org) and can be accessed and downloaded under the reference number 62041. Original identifications by Jeppsson have been preserved and are given together with revised names. Some samples are also associated with undescribed vertebrate dermal remains that will be presented as well.
Samples collected by Helmut Alberti (1932–1984) come from the localities Holmhällar (A880SF, A881SF, A883SF. A888SF, A889SF), Juves Cliff (A890SF), Kättelviken (A910SF, A914SF, A915SF, A921SF), Hoburgen (A898SF), and Hoburgen Lighthouse (A834SF, A912SF) (), and are divided into a series of numbered microslides. In a stratigraphical chart made by Alberti, the level of samples from Holmhällar is labelled “(Hamra)/Sundre”, the age of samples from Kättelviken and Juves cliff are described as “Hamra/Sundre”, and the two from Hoburgen Lighthouse are labelled as “Sundre/(Hamra)”. The sample from Hoburgen (A 898 SF) is not included in the chart. Both Jeppsson’s and Alberti’s samples are housed at the NRM-PZ.
One sample (sample ID115822) collected by Dimitri Kaljo comes from the Västerbackar 1 (Sundre Formation) locality (), and is stored at the Department of Geology, TalTech. More information about the sample can be found at the Geoscience collections of Estonia website (http://geocollections.info/).
Anders Martinsson’s collection and the newly collected samples are housed at PMU. Martinsson’s samples come from Hoburgen 1 (AM1) and Juves localities (AM2). They were collected between 1956–1959 and are labelled “Sundre”, presumably indicating their stratigraphical level. The newly collected material was sampled in September 2014 and comes from the southern part of Gotland (): G14-18OB weighed 27 kg and comes from Barshageudd 2 at GPS (WGS 84: 56°54ʹ20.0”N 18°11ʹ16.0”E); G14-19OB comes from Barshageudd 3 (5.4 kg, 56°54ʹ22.2”N 18°11ʹ39.1”E); G14-20OB from Storms 2 (8 kg, 56°57ʹ59.5”N 18°18ʹ17.7”E); G14-21OB from Hamra 3 (8.9 kg, 56°58ʹ46.0”N 18°16ʹ55.0”E); G14-22OB from Hamra 4 (13.2 kg, 56°58ʹ59.8”N 18°16ʹ42.6”E); G14-23OB from Sibbjäns 3 (5.7 kg, 56°59ʹ28.6”N 18°15ʹ39.5”E). The rock samples were dissolved at the Department of Geology at Lund University, following the technique of Jeppsson and Anehus (Citation1995) and Jeppsson (Citation2005a). The phosphatic remains were separated using the heavy liquid separation technique described by Schiøler (Citation1989). Lithological samples from Barshageudd 2, Hamra 3, Storms 2, and Sibbjäns 3 were analysed for δ13C and δ18O values at the isotope facility of GeoZentrum Nordbayern (see e.g., Jarochowska et al. Citation2016a for details of the analytical procedure).
Scanning electron microscopy (SEM) of microremains coated in a gold-palladium alloy was performed at the Evolutionary Biology Centre, Uppsala University with a Zeiss Supra 35VP. Selected conodonts were coated with gold and photographed using a Vega\\xmu SEM in Erlangen. The majority were photographed in several focus planes and are figured using stacked images created in the program CombineZP (Hadley Citation2005). Conodonts were immersed in ethanol and photographed as stacked images using a Zeiss Axio Zoom V16 with an Axiocam 506 camera at GeoZentrum Nordbayern, University of Erlangen-Nuremberg.
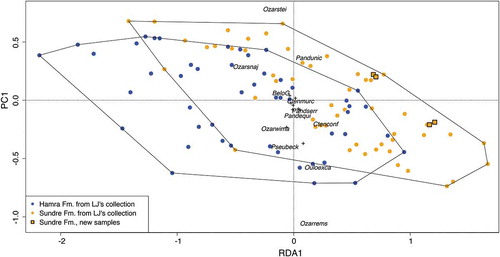
To evaluate the compositional differences in conodont faunas between the Hamra and Sundre formations, presence-absence data from Jeppsson’s collection, as well as from the newly collected samples, was analysed using redundancy analysis (RDA, Ter Braak Citation1986). Only species-level records were used and uncertain identifications were omitted. Four new samples and 120 samples from Jeppsson’s collection satisfied these requirements. A community composition matrix consisting of presence-absence records of 22 conodont species was subjected to RDA with Formation as a constraining factor. RDA implemented in the package “vegan” (version 2.5–4, Oksanen et al. Citation2019) for R Software (version 3.5.1, Team Citation2018).
Conodont biostratigraphy and isotope chemostratigraphy
Globally, two conodont zones are distinguished in the upper Ludfordian: Ozarkodina snajdri Interval Zone and Ozarkodina crispa Zone (Melchin et al. Citation2012). The Oz. snajdri Zone is bounded by the LAD of Pedavis latialata and the FAD of Oz. crispa. The Oz. crispa Zone corresponds to the total range of this species. The boundary with the Přídolí Stage is placed at the base of the “Oz.” eosteinhornensis sensu lato Zone. Regionally, the zonation is often different and/or more detailed, e.g., in the Baltic area (Märss & Männik Citation2013), Prague Basin (Slavík & Carls Citation2012), or in the Carnic Alps (Corradini et al. Citation2015). On Gotland, an informal sub-zonal stratigraphic scheme was proposed by Jeppsson, but never fully documented. These units, in inferred chronological order, were: “without Zieglerodina (‘Ozarkodina’) remscheidensis”, “with early Z. remscheidensis and Oz. wimani”, “with Oz. crispa and Ctenognathodus”, “without Oz. crispa/snajdri”, ‘post Oz. crispa, and “with ‘Rhipidognathus’ but without Oz. crispa”. Most samples, but not all, have also been labelled to indicate their position in the traditional lithostratigraphic scheme of Hede that was mentioned before. Conodont levels did not correspond exactly to the lithostratigraphic division, for example samples with early Z. remscheidensis and Oz. wimani were found both in the Hamra (e.g., at Strands 1) and the Sundre formations (at Klehammarsård). Furthermore, even though the zonation of Jeppsson et al. (Citation2006) referred the base of the Sundre Formation to the base of the Oz. crispa Biozone, which is placed at the FAD of Oz. crispa (Melchin et al. Citation2012), samples with Oz. crispa Walliser Citation1964 identified by Jeppsson were present in samples labelled in his collection as derived from the Hamra Formation. However, some of these samples (e.g., Barshageudd 2, 3 and Sibbjäns 3) have been placed in the Sundre Formation in (Jeppsson et al. Citation2006), but note their geographical positions in .
The Hamra formation
Only samples from Uddvide 5, Skradarve 1 and Kättelviken were explicitly assigned to the “Hamra a” unit or the lowermost Hamra “Beds”. In addition, one sample from Hoburgen 2 was assigned to this unit tentatively. They contained only non-diagnostic taxa: Belodella sp., Oulodus excavatus (Branson & Mehl Citation1933) and Ou. novoexcavatus, Oz. confluens, “Oz. eosteinhornensis”, “Oz. eosteinhornensis scanica”, Oz. wimani, Panderodus equicostatus, Pseudooneotodus beckmanni Bischoff and Sannemann Citation1958, and Wurmiella excavata. Ozarkodina crispa? was also found there, but the identification was based on elements other than P1, therefore it cannot be certain and is not used here to infer on the age.
Among samples assigned to “Hamra b”, those from Husryggen 3 have not been assigned to any informal conodont level. In addition to the taxa reported from “Hamra a”, they contained Belodella sp. G, Be. resima Philip, Citation1965, Ctenognathodus confluens Jeppsson Citation1972, Oulodus elegans, Oz. snajdri, Z. remscheidensis, and Panderodus unicostatus Branson and Mehl Citation1933. They did not contain Oz. crispa, and may therefore be placed in the “with early Z. remscheidensis and Oz. wimani” level. In the “Hamra b” unit, this level was represented by samples from Bankvät 1, as well as Kättelviken 3 and 5. Several samples from these localities contained Z. remscheidensis (G82-30 CB, G81-39, G83-17 CB), as well as Oz. crispa (G82-30 CB, G81-39), Oz. roopaensis Viira Citation1994, Ou. elegans and Belodella sp. L, allowing us to refer them to the “with Oz. crispa and Ctenognathodus” level. Samples from “Hamra b” assigned to the level “with early Z. remscheidensis and Oz. wimani” came from Strands 1, and Bottarve 2. They all contained Oz. wimani, most of them also contained Z. remscheidensis, and sample G75-14LJ contained also Oz. crispa.
The “Hamra c” unit was represented by samples from Barshageudd 1, assigned to the “with Oz. crispa and Ctenognathodus” (although lacking either of these species), Hoburgen 3 and 4, Juves 1, 2, 3, 5, and Kärne 3. Apart from single occurrences of Ctenognathodus murchisoni Pander Citation1856, Icriodus sp. and Erika? divarica Murphy and Matti Citation1982, no difference in terms of conodont species composition could be found compared to other units of the Hamra Formation. Additionally, a number of samples could not be assigned to a unit within the Hamra Formation. These, however, did not contain any taxa beyond those listed above, except for one of Martinsson’s sample, labelled “Hoburgen I/Storburg I”, which contained Coryssognathus? dubius Rhodes Citation1953 and Oz. modesta? Drygant Citation1984.
The Sundre formation
There is no formal lithostratigraphic subdivision of the Sundre Formation. Nonetheless, some of Jeppsson’s samples are marked “lower”, “middle”, and “upper” positions within this formation. The lower Sundre Formation is represented by samples from Faludden 3, Hamra 1, 3, 4, Sibbjäns 3 (all assigned to the “with Oz. crispa and Ctenognathodus” level), Bringes 1 and 3, Hoburgen 4, and Juves 3, 4 and 6. All those localities were characterized by similar conodonts, containing Ctenognathodus confluens (–), Oulodus elegans elegans (–), Ou. excavatus (–), Ou. novoexcavatus, Oz. confluens (–), Oz. crispa (–, ), “Oz. eosteinhornensis” (–), Oz. roopaensis, Oz. snajdri (, ), Oz. wimani (), Pa. equicostatus, Pa. unicostatus, Ps. beckmanni, and W. excavata (). The middle part of the Formation was sampled at Otes 1 and Suders 1. At Suders 1, Oz. modesta? was found in addition to the typical taxa of the formation listed earlier. The Västerbackar 1 locality was assigned to the middle to upper part of the Sundre Formation, but did not differ in terms of conodont fossils. Among samples without more precise lithostratigraphic position than the Sundre Formation, one (GLL75-8) from Klehammarsård was assigned to the “with early Z. remscheidensis and Oz. wimani” level, perhaps erroneously, as it did not contain Oz. wimani, and the presence of Z. remscheidensis with non-diagnostic taxa did not warrant a precise assignment. The remaining samples from Klehammarsård were assigned to the “without Oz. crispa/snajdri” level and contained Z. remscheidensis, Belodella sp. G, Pa. serratus, and otherwise only non-diagnostic, long-ranging taxa. This level included also two samples from Holmhällar. Samples without Oz. crispa were also placed in the “post Oz. crispa” level (Barshageudd 3, , , ; Ojmundsbod 1). Another level was tentatively named “with ‘Rhipidognathus’ but without Oz. crispa”. Rhipidognathus is an Ordovician genus. Based on very limited material present in samples in this level, it was not possible to confirm this identification, therefore the name is used here informally until a full revision can be made. This level “Rhipidognathus” was represented in samples from Faludden 1 and 2, Holmhällar, Norebod, and Storms 1 and 2.
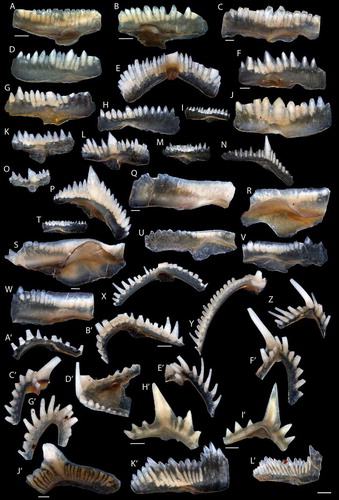
Figure 2. Conodonts from L. Jeppsson’s collection from the Hamra and Sundre formations. A–C. “Ozarkodina” eosteinhornensis Ziegler Citation1956; A. NRM-PZ Co85 from Holmhällar 1, Sundre Fm. (G87-414 LJ); B. NRM-PZ Co144 from Rivviken 1, Hamra or Sundre Fm. (G04-740 LJ); C. NRM-PZ Co87 from Holmhällar 1, Sundre Fm. (G73-78 LJ). D–E. “Oz.” eosteinhornensis?; D. NRM-PZ Co92 from Holmhällar 1 (G94-48 LJ); E. NRM-PZ Co142 from Flisviken 2, Sundre Fm. (G04-741 LJ). F–G. Zieglerodina remscheidensis? Ziegler Citation1960; F. NRM-PZ Co145 from Rivviken 1, Hamra or Sundre Fm. (G04-740 LJ); G. NRM-PZ Co84 from Holmhällar 1, Sundre Fm. (G87-414 LJ). H–I. Ozarkodina sp.; H. NRM-PZ Co134 from Flisviken 2, Sundre Fm.(G04-741 LJ); I. NRM-PZ Co81 from Västerbackar 1, Sundre Fm. (G71-185). J–N. Zieglerodina remscheidensis Ziegler Citation1960; J. NRM-PZ Co88 from Holmhällar 1, Sundre Fm. (G73-78 LJ); K. NRM-PZ Co99 from Ängvards 7, Sundre Fm. (G00-26 LJ); L. NRM-PZ Co118 from Ängvards 9, Hamra Fm. (G00-28 LJ); M. NRM-PZ Co95 from Sibbjans 1, Sundre Fm. (G94-48 LJ); N. NRM-PZ Co135 from Flisviken 2, Sundre Fm. (G04-741 LJ). O. Ozarkodina wimani Jeppsson Citation1974, NRM-PZ Co100 from Ängvards 7 (G00-26 LJ). P, T–V. Ozarkodina snajdri Walliser Citation1964; P. NRM-PZ Co149 from Rivviken 2 (G04-739 LJ); T. NRM-PZ Co80 from Västerbackar 1, Sundre Fm. (G71-185); U. NRM-PZ Co98 from Ängvards 7Sundre Fm. (G00-26 LJ); V. NRM-PZ Co137 from Flisviken 2, Sundre Fm. (G04-741 LJ). Q–S, W. Ozarkodina crispa Walliser Citation1964; Q. NRM-PZ Co172 from Faludden 3Sundre Fm. (G02-131 LJ); R. NRM-PZ Co83 from Holmhällar 1, Sundre Fm. (G87-414 LJ); S. NRM-PZ Co150 from Rivviken 2, Hamra or Sundre Fm. (G04-739 LJ); W. NRM-PZ Co141 from Flisviken 2, Sundre Fm. (G04-741 LJ). X. Wurmiella excavata Branson & Mehl Citation1933, NRM-PZ Co147 from Rivviken 1, Hamra or Sundre Fm. (G04-740 LJ). Y–Z. Oulodus excavatus Jeppsson Citation1972; Y. NRM-PZ Co146 from Rivviken 1Hamra or Sundre Fm. (G04-740 LJ); Z. NRM-PZ Co163 from Barshageudd 3, Hamra Fm. (G03-345 LJ). A’–B’. Ou. excavatus?; A’. NRM-PZ Co161 from Barshageudd 3, Hamra Fm. (G03-345 LJ); B’. NRM-PZ Co153 from Rivviken 2, Hamra or Sundre Fm. (G04-739 LJ). C’–G’. Oulodus elegans Walliser Citation1964; C’. NRM-PZ Co159 from Salmunds 1, Sundre Fm.? (G00-2 LJ); D’. NRM-PZ Co170 from Barshageudd 1, Hamra Fm. (G03-343 LJ); E’. NRM-PZ Co160 from Salmunds 1, Sundre Fm.? (G00-2 LJ); F’. NRM-PZ Co156 from Storms 2, Sundre Fm. (G94-42 LJ); G’. NRM-PZ Co154 from Rivviken 2, Hamra or Sundre Fm. (G04-739 LJ). H’–I’. Ctenognathodus confluens Jeppsson Citation1972, NRM-PZ Co138-139 from, Flisviken 2, Sundre Fm. (G04-741 LJ). J’–L’. Ozarkodina confluens Branson & Mehl Citation1933: J’. NRM-PZ Co173 from Faludden 3, Sundre Fm. (G02-131 LJ); K’. NRM-PZ Co86 from Holmhällar 1, Sundre Fm. (G87-414 LJ); L’. NRM-PZ Co155 from Storms 2, Sundre Fm. (G94-42 LJ). Elements with individual scale bars equal 200 µm in A–B, B’, H’–J’ and 100 µm in C, F, Q, S. The scale bar in bottom right applies to remaining elements and equals 100 µm.
Summary of current stratigraphic data
Samtleben et al. (Citation2000) documented a steady decrease in carbon isotope values across what they called “Hamra/Sundre Beds”, i.e., samples were not differentiated between these two units. The decrease ranged from δ13Ccarb of 7.6‰ at the base of the Hamra Formation at Uddvide 2 to 0.5 at Klehammarsård 3. This decreasing carbon isotope trend is consistent with data from the Uddvide-1 core (Younes et al. Citation2016) and reflects a global isotope trend following the end of the Mid-Ludfordian Carbon Isotope Excursion (MLCIE) or Lau isotope excursion (e.g., Munnecke et al. Citation2010). The trend allows us to arrange samples in a stratigraphic order more precisely than it would be possible based only on conodonts. Based on data from Samtleben et al. (Citation2000), the approximate boundary between the Hamra and Sundre formations in this curve can be placed between the values of 5.4‰ at Rivet 2 (Hamra Formation) and 3.2‰ at Rivviken 2 (Sundre Formation). Globally, the Lau excursion returns to background values (close to 0‰) within either the O. snajdri (e.g., in the Prague Basin, Lehnert et al. Citation2007) or the O. crispa biozones (e.g., Cramer et al. Citation2011), i.e., at variable positions with respect to the conodont zonation. This probably reflects local differences in the carbon cycle, conodont frequencies, sampling intensity, and habitat tracking of individual species. As a result, the δ13Ccarb curve does not afford the precision needed to distinguish between the snajdri and crispa zones.
Another region where investigations of the Silurian vertebrate and conodont distribution, together with carbon isotope studies, have concurrently been carried out is the Canadian Arctic islands (Märss et al. Citation1998, Citation2006). Those authors concluded that the vertebrate taxa in that region and the Baltic region may differ, but all four positive carbon isotope peaks known from the Wenlock–Přídolí and Silurian–Devonian transitions of the Baltic, Australia, and the Central Urals are well represented also in the Canadian Arctic. A carbon isotope peak in the bohemicus tenuis-kozlowskii GZ corresponds to, or is close to, the Lau Event level (middle Ludfordian) in the Baltic.
Conodont and carbon isotope-based stratigraphic position of new samples
The study by Samtleben et al. (Citation2000) did not discern between the Hamra and Sundre formations in terms of their characteristic δ13Ccarb values, but recorded a steady decrease from 7.6‰ to near 0‰ across both units. The conodont content and carbon isotope values of samples newly collected for the study are reported in . They can be tentatively arranged in a stratigraphical order based on a comparable decrease in their δ13Ccarb values, from 6.2‰ att Sibbjäns 3 to 1.7‰ at Storms 2.
Table 1. Conodont counts and carbon isotope values for samples from Sundre formation newly collected for this study. X marks samples with no measured isotope values.
Conodont abundance was lowest in samples with elevated δ13Ccarb values: samples from Sibbjäns 3 and Hamra 3 were effectively barren, whereas at Barshageudd 2 and 3 the yields were 83 and 127 elements per kg, respectively. At Hamra 4 and Storms 2, 14 elements per kg were obtained on average. Most samples were dominated by typical shallow-water fauna characterised by the presence of Ozarkodina confluens, Ctenognathodus confluens (), Oulodus excavatus (), Ou. elegans (–) and, in the stratigraphically youngest samples from Hamra 4 and Storms 2 (Sundre Formation), Ctenognathodus sp. C Strömberg Citation1997. The shallow water affinity of these species has been observed by multiple authors (Aldridge & Jeppsson Citation1984; Strömberg Citation1997; Viira & Einasto Citation2003; Jarochowska et al. Citation2016b, Citation2017).
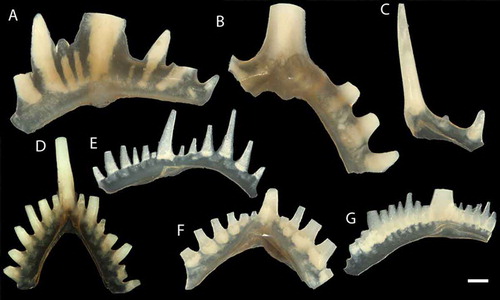
Figure 3. Conodonts from the Sundre Formation at Barshageudd 2 (G14-18OB). A. Erika divarica Murphy & Matti Citation1982. B. Ctenognathodus murchisoni Pander Citation1856. C. Ctenognathodus confluens Jeppsson Citation1972. D–E. Oulodus elegans (Walliser Citation1964). F. Oulodus excavatus (Branson & Mehl Citation1933). G. Wurmiella excavata (Branson & Mehl Citation1933). Scale bar equals 100 µm.
The most diverse conodont fauna was found at Barshageudd and contained Belodella sp. G (Jeppsson Citation1989), Belodella resima (–), Ctenognathodus murchisoni (), Decoriconus fragilis (Branson & Mehl Citation1933; –), four species of Panderodus (P. equicostatus, P. panderi, P. serratus and P. unicostatus), Pseudooneotodus beckmanni (–), Wurmiella excavata (), Erika divarica (), and Zieglerodina remscheidensis. Belodella resima is known from latest Ludfordian through at least Givetian, whereas Belodella sp. G has been described by Jeppsson (Citation1989) based on two specimens from the Silurian/Devonian boundary section at Klonk, and its stratigraphic range has not been documented. Erika divarica has been originally described from the Lower Devonian by Murphy and Matti (Citation1982) and very few new occurrences have been reported worldwide since then. The specimens discovered in the present work are very similar to those found in Jeppsson’s collection and identified as E. divarica by himself (indicated by his labels), and this identification is followed here. Oulodus excavatus and Ou. siluricus? have a long Wenlock–Ludlow stratigraphic range and do not contribute to the age constraint. The range of Ctenognathodus species is poorly known but C. confluens occurs in both units. Jeppsson assigned samples from Barshageudd in his collection to the “post Oz. crispa” level, presumably the youngest strata on Gotland, but the 2.5‰ δ13Ccarb value of sample G14-18OB argues in favour of an older age in either the snajdri or the crispa Zone.
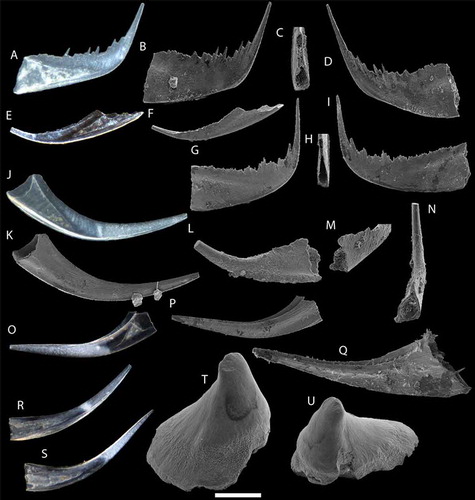
Figure 4. Coniform conodonts from the Sundre Formation at Barshageudd 2 and 3. A–N. Belodella resima Philip, Citation1965; A–I. PMU 34758–34766 from sample G14-18OB; J–N. PMU 34766–34771 from sample G14-19OB. O–S. Decoriconus fragilis Branson & Mehl Citation1933; O–Q. PMU 34772–34774 from sample G14-19OB; R–S. PMU 34775–34776 from sample G14-18OB. T–U. Pseudooneotodus beckmanni Bischoff & Sannemann Citation1958; PMU 34777–34778 from sample G14-19OB. Scale bar equals 100 µm.
The index species Oz. crispa was found only in sample G14-22OB from Hamra 4, where it co-occurred with Oz. confluens, Ctenognathodus confluens, De. fragilis and a P2 element of E. divarica?. The presence of Oz. crispa allows placing the sample in the Oz. crispa Zone of the uppermost Ludfordian, in agreement with Jeppsson’s assignment of these strata to his “with Oz. crispa and Ctenognathodus” level. Sample G14-20OB from the Sundre Formation at Storms 2 contained a low-diversity assemblage formed by Oz. confluens and Ctenognathodus sp. C, Ct. murchisoni and Ct. confluens, and lacked any age-diagnostic taxa. This composition suggests very shallow, restricted environment. Geological superposition suggests that strata at this locality are younger than those at Hamra 4, but an explicit assignment to the Oz. crispa Zone, or an even younger interval, is not warranted by the conodont assemblage.
Composition of conodont communities in the Hamra and Sundre formations
Constrained redundancy analysis of species-level conodont composition in Jeppsson’s and the newly collected samples revealed very high variability in both Hamra and Sundre formations; only 2.5% of variance could be explained by the assignment to the Formation, the remaining 97.5% was unconstrained variation (). A slight overall shift between the two units could be detected in the composition of the communities, with Ozarkodina remscheidensis, Oulodus excavatus, and Pseudooneotodus beckmanni being most characteristic for the Hamra Formation, and Ozarkodina steinhornensis and Panderodus unicostatus for the Sundre Formation.
Figure 5. Redundancy analysis of species-level conodont faunas in samples from Lennart Jeppsson’s collections from the Hamra and Sundre formations (blue and orange circles, respectively), as well as of the newly collected samples in this study (squares). The analysis was constrained by formations. For clarity, only the most abundant species are shown (shortened taxonomic names).
Systematic palaeontology
Agnathans
Subclass Thelodonti Jaekel Citation1911
Order Thelodontiformes Kiær, 1932
Family Coelolepididae PANDER, Citation1856
Genus Thelodus Agassiz, Citation1838
Thelodus parvidens Agassiz, Citation1838(–)
Figure 6. A–G: Thelodus parvidens; A. GIT 791–1 from sample ID115822; B. NRM-PZ C6016 from G00-26LJ; C–D. NRM-PZ C6017–6018 from G03-345LJ; E. NRM-PZ C6019 from G94-42LJ; F–G. NRM-PZ C6020–6021 from G71-184BLJ. H–J. Thelodus trilobatus; H. NRM-PZ C6022 from A912SF; I–J. NRM-PZ C6023–6024 from G04-74LJ. K–Q. Thelodus sculptilis; K. NRM-PZ C6025 from G87-414LJ; L. NRM-PZ C6026 from G00-21LJ; M. NRM-PZ C6027 from G87-414LJ; N. NRM-PZ C6028 from G00-21LJ; O–P. NRM-PZ C6030–6031 from G87-414LJ. All in external view with forward facing left, except F and J (lateral view). Scale bar equals 200 μm.

Material
More than a thousand scales.
Description
Head scales of Thelodus parvidens have crenulated margins all around the circumference of the crown, but the central area of the crown is smooth and flat (). The neck in these scales is low and the base is usually quite large and rounded. Transitional scales are crenulated along the anterior and lateral crown rims, and (similar to the head scales) the central part of the crown is smooth and flat (). The neck is usually low in the transitional scales, but can sometimes be rather high as well. Trunk scales have smooth, drop-shaped and flat crowns ending in a posterior point (), or the crown can be more angular and rhomboidal in outline (). In some trunk scales, the base is of similar dimensions as the crown, while others have a restricted base and a large, posteriorly overhanging crown. Depending on the age of the scale, the base can be shallow or deep and the centrally positioned pulp opening can be large or very small. The neck is prominent and often high with vertical ribs on the antero- and posterolateral sides. Some scales with typical trunk scale morphologies have very low necks and often widely open pulp cavities only surrounded by a thin rim. The transition between the base and neck is often smooth, but can be developed as a quite prominent edge in some scales in the samples at hand.
The bicostatiform scales have two parallel and medially running ribs on the crown that are divided by a median furrow (). There may be an additional set of ribs and troughs on the lateral sides. These scales are quite different compared to the regular trunk scales, but they are included in the T. parvidens scale-set because they occur together with regular scales on articulated specimens (see Remarks).
A few traquairiform scales () have been found in these samples. They have a moderately deep base, limited neck, and a strongly inclined crown with an almost central apex. Ridges that converge toward this point have a knee-like bend where they meet the neck. There are often scales that acquire crown morphologies in between traquairiform scales and normal transitional scales, where the posterior part is smooth and pointed, but the anterior part host poorly developed ridges and furrows ().
Remarks
Thelodus parvidens was originally a scale-based taxon of fairly limited definition, but the discovery of shared histological features among morphologically distinct scales within assemblages of microremains led to the inclusion of costatiform, pugniform, bicostatiform, and trilobatiform scales (see Vergoossen Citation2002a for discussion). In parallel to this, the re-examination of a semi-articulated specimen of Thelodus macintoshi Stetson, 1928 suggested that macintoshi was a younger synonym to T. parvidens, and led to the discovery of bicostatiform and trilobatiform scales in association with regular T. parvidens scales (Turner Citation1986). However, this treatment of T. macintoshi was not agreed upon by all (e.g., Märss & Miller Citation2004). Thelodus traquairi Gross Citation1967 has been presented as traquairiform scales within the squamation of T. parvidens, although some kept it as a separate species (see Märss et al. Citation2007). A body zonation scheme has been implemented when studying assemblages of disarticulated thelodont scales for a long time (Märss et al. Citation2007), and Vergoossen (Citation2002a) presented several different morphotypes for T. parvidens, but also flagged some difficulties in assigning scales to different taxa using body zonations in mixed microvertebrate assemblages.
Besides being a relatively broad taxon in terms of scale morphologies, T. parvidens was also long-ranging and seems to have lived in a wide range of environments (Kaljo et al. Citation2015), since their remains are found in many different lithologies from upper Ludlow, Ludfordian, to late Přídolí (excluding its uppermost part) (Märss Citation1986; Märss & Miller Citation2004; Märss & Männik Citation2013). On Gotland, it has been reported from the Ethelhem Formation of middle Ludlow age to the topmost part of the Gotland stratigraphy in rock-types ranging from marls to reefal limestone, sandstones, and shallow water oolites (Fredholm Citation1988a, Citation1989; Eriksson et al. Citation2009).
Occurrence on Gotland
Ethelhem Formation (Fredholm Citation1988a), När Formation (Fredholm Citation1988a; Eriksson et al. Citation2009), Eke Formation (Spjeldnaes Citation1950; Fredholm Citation1989; Eriksson et al. Citation2009), Burgsvik and Hamra formations (Fredholm Citation1989; Eriksson et al. Citation2009). Lower Hamra Formation: Kättelviken (A910SF, A921AF), Skradarve 1 (G02-139LJ). Middle Hamra Formation: Bottarve 2 (G83-5LL). Upper Hamra Formation: Rivviken 1 (G04-740LJ), Juves (AM2), Juves kliff (A890SF). Sundre Formation (Fredholm Citation1989). Lower Sundre Formation: Hamra 4 (G14-22OB), Rivviken 2 (G04-739LJ), Barshageudd 1 (G03-343LJ), Barshageudd 4? (G04-736LJ), Faludden 3 (G02-131LJ). Upper Sundre Formation: Barshageudd 2 (G14-18OB), Barshageudd 3 (G14-19OB), Holmhällar 1 (G73-78BLJ), Holmhällar (A880SF, A881SF, A883SF, A888SF, A889SF), Västerbackar 1 (ID115822, G71-184BLJ), Barshageudd 3 (G03-345LJ), Storms 2 (G94-42LJ, G14-20OB). Unclear stratigraphical placement: Hoburgen (A898SF), Hoburgen Lighthouse (A834SF and A912SF), Hoburgen 1 (AM1).
Thelodus trilobatus Hoppe, Citation1931(–)
Material
33 scales.
Description
Thelodus trilobatus displays a wide range of scale morphologies (Märss et al. Citation2007). The scales recovered in the samples treated here can generally be divided into three scale types. The first type () has a crown with three posterior points. There is a median trough alongside two rims on the crown that end in one point, and rather large and smooth lateral extensions on either side that also end in points that overhang the base. In some of these scales, there are additional longitudinal ridges on the lateral extensions of the crown. The central posterior point of the crown has pronounced keel along the ventral side. The base is large and anteriorly offset, and it often has a substantial anterior process. The pulp opening is small and placed posteriorly on the base. The second type () has a flattened crown morphology that ends in several points posteriorly. There is a shallow median furrow and two medial ribs at the anterior end of the crown. The base is similar to the first type, but the anterior process is usually not as large. The crown in both these scale types is horizontal, but turns gently downwards toward the anterior. The neck is low but quite distinct. The third scale type () has a large, smooth and flat crown with a drop-like morphology that ends in one posterior point. The neck in these scales is high, but the base is of similar morphology as the other two types.
Remarks
Several thelodont taxa appear to have had trilobatiform scales as part of their squamation, for example Thelodus, Turinia, and Lanarkia, the latter two found in articulated specimens (Märss et al. Citation2007). As discussed above, trilobatiform scales were discovered in association to regular parvidens-type scales on a semi-articulated specimen. However, Vergoossen (Citation2002a) emphasized that only a single trilobatiform scale was present on the specimen and he speculated that it could even be allochthonous. Furthermore, a huge difference in the abundance of trilobatiform scales have been observed in largely coeval samples from different environmental settings in the Holy Cross Mountains of Poland. More than a hundred scales were recovered from one sample but only five scales were found in the other one, while the number of typical parvidens-type scales remained fairly constant in both (Bremer et al. Citation2017). This is surprising if they indeed come from the same species.
Occurrence on Gotland
Equated with T. parvidens in previous works on Gotland, see distribution for T. parvidens. Trilobatiform scales confirmed in: Lower Hamra Formation: Kättelviken (A921SF). Middle Hamra Formation: Bottarve 2 (G83-5LL). Upper Hamra Formation: Rivviken 1 (G04-740LJ), Ängvards 7 over 8 under 3 (G00-26LJ). Lower Sundre Formation: Hamra 4 (G14-22OB), Rivviken 2 (G04-739LJ), Barshageudd 1 (G03-343LJ), Flisviken 2 (G04-741LJ). Upper Sundre Formation: Storms 2 (G94-42LJ), Västerbackar 1 (G71-184BLJ), Salmunds 1 (G00-2LJ).
Thelodus sculptilis Gross, Citation1967(–)
Material
45 scales.
Description
In general, the majority of Thelodus sculptilis scale crown surfaces are rather heavily sculptured, while those of T. parvidens are generally smooth. Nevertheless, T. sculptilis may display a similar range of scales as T. parvidens and some of the special scales are hard to distinguish morphologically between the two. Head scales are crenulated all along the circumference, but the grooves and ridges generally extend radially much further onto the crown (), and the crowns are generally not as smooth and flat as those in T. parvidens. The same is generally true for other, more anterior and transitional scales (–). Some anterior scales may have a smoother crown (), but the lateral grooves are usually deeper and the anteromedian extension is more pronounced than those in scales of T. parvidens. Trunk scales of T. sculptilis are easily distinguished as their crowns have two pronounced medial furrows that converge posteriorly, and a central bulge on the crown anteriorly that sometimes hosts a shallow furrow as well (–). The outline of the crown is rhomboidal to drop-like and can end in several posterior points (), or a single posterior point (). The base is usually rhomboidal to roundish, quite low, and hosts a large, circular pulp cavity. The transition between base and neck is usually distinct, and the neck itself can be low or rather high. In some trunk scales, the neck hosts vertical ribs mainly on the posterolateral sides.
Remarks
Fredholm (Citation1989) described scales of T. sculptilis from Burgsvik Formation at the Glasskär 1 locality (Burgen outlier), but no scales were figured and no counts were given because the samples had not been fully picked through at that time. One scale identified as T. sculptilis in sample G02-134LJ from Kättelviken 1 locality in the Burgsvik Oolite Member of the Burgsvik Formation were figured in Nilsson (Citation2005: fig. 13P). Eriksson et al. (Citation2009) referred reports of T. sculptilis by Fredholm (Citation1989) in sample G82-321DF as possibly coming from the Burgsvik Formation, but this residue came from a loose slab that had most likely been transported away from its original location. One scale identified as T. sculptilis from Skradarve 1 (sample G02-139LJ) of the Hamra Formation was figured in Eriksson et al. (Citation2009: ). A review of the material in these previous studies needs to be done before any definite comments can be made on these records. However, scales identified as Loganellia cuneata Gross Citation1947 from the remaining residue of sample G02-134LJ (OB pers. obs. 2019) have similar features on its crown as the scales mentioned above.
Occurrence on Gotland
Burgsvik Formation (Fredholm Citation1989; Eriksson et al. Citation2009), Hamra Formation (Eriksson et al. Citation2009). Sundre Formation (Fredholm Citation1989). Lower Sundre Formation: Hamra 4 (G14-22OB). Upper Sundre Formation: Barshageudd 3 (G14-19OB), Salmunds 1 (G00-2LJ), Holmhällar 1 (G87-414ALJ).
Order Loganelliiformes Turner,1991
Family Loganelliidae Märss et al., 2002
Genus Loganellia Turner,1991
Loganellia cuneata Gross, Citation1947(–)
Figure 7. A–D: Loganellia cuneata; A. specimen GIT 791–2 from sample ID115822; B–D. NRM-PZ C6032–6034 from G02-139LJ. E–K. Paralogania ludlowiensis; NRM-PZ C6035–6041 from G02-139LJ. L–O. palmatilobate scales; NRM-PZ C6042–6045 from G02-139LJ. All in external view with forward facing left, except A (lateral view). Scale bar equals 200 μm.

Material
9 scales.
Description
The few scales identified as Loganellia cuneata in the samples presented here generally display two morphologies. One of the scales () has a large, tumid base with a short anterior spur and a small pulp cavity opening on the posterior end. The neck is marked but low. The slightly inclined and curved crown has a midline and two lateral troughs divided by pronounced ridges that converge toward the single posterior point. The remaining scales are all low and elongated narrow, but host similar troughs and ridges on the crown (–), although the median part in the scale in is rather smooth. The base is often only developed as a narrow rim around a widely open pulp cavity, which extends as a large pulp canal through the crown ().
Remarks
Scales of L. cuneata are very rare in the samples at hand, and some of them may represent fin scale variants, making their affinities more uncertain. Fredholm (Citation1988a) reported “Logania” cuneata from several localities, but all of these previous reports were revised as other taxa (mainly as Lanarkia horrida Traquair Citation1898) by Nilsson (Citation2005). Eriksson et al. (Citation2009) reported additional scales of Lan. horrida, but the material of previous studies needs to be reviewed before any further comments can be made.
Occurrence on Gotland
När Formation (Fredholm Citation1988a, but see remark above). Lower Hamra Formation: Skradarve 1 (G02-139LJ). Upper Sundre Formation: Västerbackar 1 (ID115822).
Order Shieliiformes Märss, Wilson & Thorsteinsson, 2002
Family Shieliidae Märss, Wilson & Thorsteinsson, 2002
Genus Paralogania Karatajūte-Talimaa, 1997
Paralogania ludlowiensis Gross, Citation1967 (–)
Material
98 scales in total.
Description
Anterior/transitional scales (–) have a large base that extends outwards anteriorly and laterally. The transition to the crown is smooth, but the edge to the top of the flat crown is steep. The crown is rounded and smooth anteriorly, but the posterolateral side has several extensions. Another transitional (or special) scale has a large, more rhomboidal base and an outstretched crown with wide, delicate lateral extensions (). Trunk scales are large and have elongated oval and often slightly asymmetrical crowns with sharp anterior and posterior points (–). The crowns are flat with a steep angle to the lateral sides. The crown surface sometimes has a shallow, median trough as well as an anterior notch (). Prominent lateral ridges originate roughly halfway along the crown at the base and crown interface. These rise obliquely posteriorly and meet just below the posterior point. Beneath these ridges are five or more postero-lateral extensions (spines), although only their attachments generally preserve. In exquisitely preserved scales (e.g., Miller & Märss Citation1999) the delicate extensions are long and leaf-like. The base is widened anteriorly but narrows posteriorly and has a mostly small pulp opening positioned close to the posterior corner of the base. There is often a furrow expanding anteriorly from the pulp opening on the base. Most of the trunk scale crowns are quite broad, but some are narrow and outstretched with small lateral ridges and extensions restricted to the posterior-most part of the crown (). There are also smaller scales with a narrow, longitudinally outstretched crown morphologies () that approach the morphology of fin scales (Miller & Märss Citation1999: pl. 2, fig. 25).
Remarks
Scales of P. ludlowiensis are found in samples from areas preserving the lower parts of Hamra Formation (Hoburgen, Skradarve, Ängvards 4?). The taxon is extremely abundant in the underlying Burgsvik Sandstone and Burgsvik Oolite members (OB pers. obs. 2019), and it has been proposed that its distribution is fairly limited to similar lithologies (Kaljo et al. Citation2015; Bremer et al. Citation2017). A possible exception to this is sample A912SF, which also contains several typical scales of P. ludlowiensis, but is labelled as coming from the Hoburgen Lighthouse and was listed as “Sundre/Hamra-kalk” in Alberti’s stratigraphical chart. There is a lighthouse situated on top of a hill of massive limestones close to the southern tip of Gotland, but no such locality has been found in any lists of Gotland localities (e.g., Laufeld Citation1974). Hence, the location, lithology, and stratigraphical position of this sample is uncertain, and there are oolites present in the general area.
Occurrence on Gotland
Eke and Burgsvik formations (Fredholm Citation1989; Eriksson et al. Citation2009), Hamra Formation (Eriksson et al. Citation2009). Lower Hamra Formation: Skradarve 1 (G02-139LJ), Ängvards 4? (G90-125LJ). Unclear stratigraphical position: Hoburgen Lighthouse (A912SF).
Palmatilobate scales of Paralogania? sp. Märss, Citation2006 (–)
Material
9 scales.
Description
The base is moderately thick on the posterior and lateral sides, and often hosts a substantial anterior spur. The transition to the neck is usually developed as a well-marked edge and the neck is usually low, but can sometimes be quite pronounced (). The crown usually has a more pronounced median area that can either have a furrow (, ) or be almost flat (), and which ends in one posterior point. The posterior point has thinner lateral extensions that originate at a keel on the ventral side. The crown has two additional areas on each side with medial troughs that also end in free points posteriorly and have thinner lateral extensions originating on the underside as well.
Remarks
Relatively short and multi-ridged palmatilobate (or palmatiform) scales were first described by Märss (Citation2006) in several drill cores from the Kuressaare Stage of Estonia. They were similar to some Paralogania species and Nethertonodus laadjalaensis Märss Citation2006 in morphology, sculpture, and ultrasculpture. Palmatilobate scales did not have the upturned spines on the anterior parts of the ridges that often occur on N. laadjalaensis, and the anterior spur was shorter and often directed downwards. Palmatilobate scales were more common in samples than N. laadjalaensis and often occurred together with Paralogania perensae Märss Citation2006 and other upper Ludlow–Přídolí taxa such as L. cuneata (Märss Citation2006). In older sedimentary rocks, very rare but similar types of scale morphologies have been found together with P. martinssoni scales. At present we conclude that palmatilobate scales could belong to the species of genera Nethertonodus (N. laadjalaensis), and Paralogania (P. martinssoni, P. ludlowiensis?, P. perensae) in a similar way as more simple trilobatiform scales belong to the species of genus Thelodus (T. parvidens, T. laevis, T. carinatus). Comparable types of scales occur also in species of the genus Shielia (see S. taiti in Märss & Ritchie Citation1998: figs 27, 29–31E; S. parca and S. gibba in; Märss et al. Citation2006: text-figs. 20 and 21, respectively).
Occurrence on Gotland
När Formation (Fredholm Citation1988a). Lower Hamra Formation: Skradarve 1 (G02-139LJ). Upper Hamra Formation; Ängvards 7 över 8 under 3 (G00-26LJ). Lower Sundre Formation: Hamra 4 (G14-22OB).
Indeterminable Thelodonti (–)
Figure 8. A. possible head scale of Thelodus parvidens; specimen NRM-PZ C6046 from G03-345LJ. B–C. pinnal scales probably of Loganellia; NRM-PZ C6047–6048 from G02-139LJ. D–I. traquairiform scales; D–E. NRM-PZ C6049–6050 from G04-738LJ; F. NRM-PZ C6051 from A889SF; G. GIT 791–3 from ID115822; H. NRM-PZ C6052 from G00-86LJ; I. NRM-PZ 6053 from G03-345LJ. J. Thelodontida gen. et sp. indet.; NRM-PZ C6054 from G00-26LJ. All in external view with forward facing left. Scale bar equals 200 μm.

There are also a number of indeterminable thelodont scale types in the samples. The first one is a costatiform scale () from the Barshageudd 3 (G03-345LJ) sample, which has prominent ridges and grooves radiating from a central point on the crown. The free tips on the periphery of these ridges are slightly up-turned. The neck is pronounced and restricted, and the base is low and round with a round pulp opening. The ornamentation suggests that it could be a head-scale within the squamation of T. parvidens as suggested by Gross (Citation1967). Two scales have shallow bases and extremely elongated crowns hosting several ridges and troughs converging toward a single posterior point (–). These are most likely pinnal scales of a Loganellia, but their exact affinities are unclear. Another scale () could be a special scale within a Thelodus squamation, but its affinity is uncertain. The base in this scale is deep, especially anteriorly and the neck is constricted and moderately high. The crown is triangular but wider than long and the two lateral corners have pronounced ridges that turn downwards toward the base. The crown has a median ridge anteriorly that extends as a faint rib to the posterior point. There is also one more ridge in between the median one and both lateral corners. Similar scales were identified as Thelodus traquairi by Vergoossen (Citation2002a: pl. 2 fig. 25), but its affinity is actually uncertain. Samples A889, G00-2LJ, G00-26LJ, G00-86LJ, G03-345LJ, G04-739, G14-22OB, and ID115822 contain rare scales (–) that show similarities to other scales described as T. traquairi by Vergoossen (Citation2002a: pl. 3, figs. 42–43). However, it is unclear whether these scales represent its own taxon, or if they are specialized scales within the squamation of T. parvidens (see Märss et al. Citation2007). It is not included in T. parvidens here because of the presence of T. sculptilis, which shows a somewhat similar morphological range of head scales as T. parvidens. Another scale has a gently inclined, triangular crown with irregular small ridges running in an anteroposterior direction (). Vergoossen (Citation2002a: pl. 1, figs. 16–17) described similar scales as “monocusped trilobatus form with (bi)costatus features” of T. parvidens. We do not agree with this because the scale is strongly elongate, which is not common in T. parvidens.
Subclass Osteostraci LANKESTER, Citation1868
Order incertae sedis
Family incertae sedis
Genus Tahulaspis Märss et al., Citation2014
Tahulaspis sp. (–)
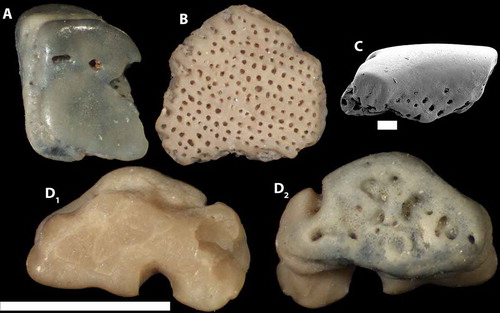
Figure 9. A-B. Tahulaspis sp.; NRM-PZ C6055–6056 from sample A912SF. C. Osteostraci gen. et sp. indet.; NRM-PZ C6057 from sample A912SF. D. Osteostraci gen. et sp. indet.; NRM-PZ C6058 from A912SF. A–B and D1 in external view, D2 in basal view, and C in lateral view. Scale bar represents 1 mm in A–B and D, and 100 μm in C.
Material
A total of eight fragmentary remains.
Description
These osteostracan remains are small and fragmentary, but most show possible affinities to Tahulapsis ordinata Märss et al. Citation2014 or Tahulaspis praevia Märss et al. Citation2014. Two fragments have large, smooth and flat-topped ridges divided by round pores (e.g., ). The largest piece present in the sample () is entirely covered by irregular rows of small and round pores in between flat ridges. The irregularity of the sculpture makes it reminiscent of Tah. praevia, but more material is needed for a definite assignment.
Remarks
Only a few osteostracan remains are present in the samples presented here, most of them coming from the Hoburgen area and the Skradarve 1 localities. The two Tahulapsis taxa discussed above have similar ranges in Estonia (Märss et al. Citation2014), so it is possible that both are present.
Occurrence on Gotland
Lower Hamra Formation: Skradarve 1 (G02-139LJ). Unclear stratigraphical placement: Hoburgen Lighthouse (A912SF).
Gen. et sp. indet. (–)
Material
Four fragments
Description
The A912SF sample contained a small piece consisting of a smooth ridge with a distinct notch at the anterior end, a single posterior point, and several small pores along the bottom (). There are two small and fragmentary remains of osteostracan ridges similar to also in the sample from Skradarve 1. Another fragment in the A912SF sample is composed of a flat-topped rib with an irregular outline (). The transition to the vertical sides is smooth, as is the transition to the massive base. The base has several irregular vascular canal openings of differing size (). The affinity of these fragments remains uncertain
Occurrence on Gotland
Lower Hamra Formation: Skradarve 1 (G02-139LJ). Unclear stratigraphical position: Hoburgen Lighthouse (A912SF).
5.2. Gnathostomes
Class Acanthodii Owen, 1846
Order Climatiiformes BERG, Citation1940
Family Climatiidae BERG, Citation1940
Genus Nostolepis Pander, Citation1856
Nostolepis striata Pander, Citation1856 (–)
Figure 10. A–J. Acanthodian scales referred to Nostolepis striata; A–B. NRM-PZ P16351–16352 from sample G71-185LJ; C. PMU 23100 from AM1; D–H. NRM-PZ P16353–16357 from G71-185LJ; I–J. PMU 23101–23102 from AM1. K–N. Gomphonchus sandelensis; K. NRM-PZ P16358 from G71-185LJ; L. NRM-PZ P16359 from G87-414LJ; M. NRM-PZ P16360 from G00-26LJ; N. NRM-PZ P16361 from G83-5LL. O–Q. “pinaceus variant”; NRM-PZ P16362–16364 from A912SF. R. “forma bifurcata”; NRM-PZ P16365 from G02-139LJ. Scale bar equals 200 μm.

Material
148 scales.
Diagnosis based on Gross (Citation1947). Moderately tumid base (although flatter bases do occur) with centrally or anteriorly placed swelling and rhomboidal outline. The neck is usually restricted anteriorly, but higher on the posterior side of the crown. The crown is mostly elongated rhomboidal with a drawn out posterior point. Head scales have a high, concave neck around a rounded crown. The crown is usually inclined, but sometimes nearly horizontal. Crown sculpture is extremely variable with ribs of varying sizes and number radiating from the anterior end toward the posterior point. These can extend far onto the uneven crown, or be restricted to the anterior, creating an otherwise smooth crown. Some crowns have several riblets on the lateral sides, as well as riblets that are more pronounced at the anterior end. These riblets can sometimes converge anteriorly to create anterior points. Crowns may be largely symmetrical, or asymmetrical where one side of the crown is more prominent. The crown also has a median furrow, mainly developed at the anterior end. Outside the perimeter of the crown is an additional ridge on each side that rise obliquely toward the posterior point.
Description.
The “nostolepid” scales from the samples presented here display a wide array of scale morphologies, but the most numerous are typical Nostolepis striata scales as figured by Gross (Citation1947). These have almost symmetrically rhomboidal and tumid bases, and an inclined, triangular crown ending in one posterior point (–). The crown hosts a number of riblets along the anterior end, two of which develop into prominent lateral rims. On the outside of these are oblique ridges on either side of the crown that converge toward the posterior point. The median area of the crown can be smooth () or have a shallow to deep furrow (–). As a result, some scales have a seemingly two-pointed crown posteriorly (–), although this may potentially be the result of two individual scales having grown together in some cases (). One of the scales with a typical N. striata crown morphology has a much steeper and pointed crown, which is much smaller relative to the size of the base (). A special variant is only represented by one scale in the material and has features typical for N. striata as described above, but the crown is laterally flattened and the neck is restricted all around ().
Some of the N. striata scales have large and more elongated bases, hosting elongated and almost flat crowns (–). In a few of these scales, the anterior riblets converge toward the anterior (), while other have a more open and obliquely slanting side composed of several ribs (). The lateral oblique ridges are generally more constricted and originate roughly halfway along the crown.
Remarks
Gross (Citation1947, Citation1971) included a wide array of scale morphologies and established taxa from middle Silurian to lower Devonian rocks into N. striata. The definition of N. striata was kept fairly broad also by Vergoossen (Citation1999a, Citation2002a, Citation2002b, Citation2002c, Citation2003a, Citation2004), although he did consider Gross’ inclusion of taxa from such a wide geographical extent and long temporal range as problematic. Valiukevičius (Citation2003a, Citation2004a) viewed the morphological definition of N. striata as too broad and erected several taxa from it, a few of which are similar to the scales presented above. The scales in – are reminiscent of scales figured as transitional scales of Nostolepis amplifica Valiukevičius Citation2003a or Nostolepis elegans Brotzen Citation1934 in Valiukevičius (Citation2005: –). Vergoossen (Citation2002c: pl. 16, figs.94–95) considered scales similar to – as part of the N. striata squamation, but recognized them as a distinguishable “elegans form group”, although scales figured by him usually have long posterior overhangs, and scales with similar crowns to that in were considered as N. striata trunk scales (Vergoossen Citation2004: pl. 5, fig. 59). Valiukevičius and Burrow (Citation2005) emphasized the importance of histology in the taxonomy of Nosolepis-type acanthodian scales (see Section Discussion). The comparisons above therefore remain tentative, because no histological studies have been performed on the scales presented in this work.
Occurrence on Gotland
När Formation (Fredholm Citation1988a; Eriksson et al. Citation2009), Eke Formation (Spjeldnaes Citation1950), Burgsvik and Hamra formations (Fredholm Citation1989; Eriksson et al. Citation2009). Middle Hamra Formation: Bottarve 2 (G83-5LL). Upper Hamra Formation: Ängvards 7 över 8 under 3 (G00-26LJ). Sundre Formation: in Fredholm (Citation1989), Hoburgen 1 (AM). Lower Sundre Formation: Hamra 4 (G14-22OB). Upper Sundre Formation: Holmhällar (A883SF, A889SF), Holmhällar 1 (G87-414ALJ), Västerbackar 1 (G71-184BLJ, G71-185LJ, ID115822), Storms 2 (G94-42LJ, G14-20OB), Barshageudd 3 (G03-345LJ). Unclear stratigraphical position: Hoburgen (A898SF), Hoburgen Lighthouse (A 834 SF).
Regional occurrence
Considering the taxonomical revisions discussed above and in the following sections, the regional distribution of N. striata is difficult to delimit. Vergoossen (Citation2002b) point out that scales have been referred to the Gross (Citation1947, Citation1971) definition of N. striata from strata world-wide ranging in age from Wenlock to Emsian.
Order Ischnacanthiformes BERG, Citation1940
Family Ischnacanthidae WOODWARD, Citation1891
Genus Gomphonchus Gross, Citation1971
Gomphonchus sandelensis Pander, Citation1856 (–)
Material
343 scales.
Diagnosis based on Vergoossen (Citation1999c). Large adult flank scales (up to 1.4 mm) with a smooth crown that is rhombic or has a rounded anterior crown margin. The crown extends beyond the base posteriorly and all around. Anterior crown may host faint and short radial riblets. The neck is high and has ribbing. Convex base with antero-central swelling.
Description
The base is either symmetrically or asymmetrically rhomboidal with an anteriorly placed swelling. The neck is low to high, but always distinct all around with a sharp edge to the crown. The crown is also rhomboidal with sharp lateral corners, but a smooth anterior outline (). In some scales, however, the anterior crown hosts small riblets and troughs (), while some scales can have quite marked anterior furrows (). Most scales have crowns that are as large as, or larger, than the base, and there is always a posterior overhang. However, in some scales the crown is smaller than the base on both the lateral and anterior sides.
There are also scales that acquire an outstretched rhomboidal morphology of both base and crown (). These are quite unlike the other “gomphonchid” scales, but they are still considered as such because the base has an offset swelling and the crown has sharp edges to a distinct neck all around the crown.
Remarks
All the “gomphonchid” scales are here tentatively referred to as G. sandelensis, but there are some taxonomical uncertainties regarding this taxon. Gross (Citation1947) included scales described as Gomphonchus volborthi Rohon, 1893 in G. sandelensis, but Vergoossen (Citation1999a, Citation1999b) considered G. volborthi a valid taxon distinguishable by scales having low necks, pronounced anterior furrows and ridges, as well as crown outlines smaller than the base. Furthermore, he considered G. sandelensis as younger (late Přídolí) than G. volborthi, subsequently referring older reports by Fredholm (Citation1988a) to G. volborthi. However, Valiukevičius (Citation2003b) did not agree with this designation and viewed the specimens figured by Fredholm (Citation1988a: –) as G. sandelensis. Furthermore, he viewed Rohon’s (Citation1893)G. volborthi as potentially being related to his Nostolepis paravolborthi Valiukevičius Citation2003b, and he considered the specimens referred to G. volborthi by Lehman (Citation1937) as G. sandelensis, while the specimens described by Vergoossen (Citation1999b) as possibly being related to N. paravolborthi.
Occurrence on Gotland
När Formation (Fredholm Citation1988a; Eriksson et al. Citation2009), Eke Formation (Spjeldnaes Citation1950; Eriksson et al. Citation2009), Burgsvik and Hamra formations (Fredholm Citation1989; Eriksson et al. Citation2009). Lower Hamra Formation: Skradarve 1 (G02-139LJ), Kättelviken (Husryggen?) oberhalb (A915SF), Ängvards 8 (G00-27LJ), Ängvards 4? (G90-125LJ), Ängvards 6 over 5 under 4 (G00-25LJ). Middle Hamra Formation: Sibbjäns 1 (G94-48LJ), Bottarve 2 (G83-5LL), Ängvards 9 över 4 (G00-28LJ). Upper Hamra Formation: Ängvards 7 over 8 under 3 (G00-26LJ). Sundre Formation (Fredholm Citation1989). Lower Sundre Formation: Hamra 4 (G14-22OB). Upper Sundre Formation: Holmhällar (A880SF, A889SF), Holmhällar 1 (G87-414ALJ), Västerbackar 1 (ID115822, G71-184BLJ, G71-185LJ), Salmunds 1 (G00-2LJ), Storms 2 (G94-42LJ, G14-20OB), Barshageudd 3 (G03-345LJ). Unclear stratigraphical position: A 898 SF (Hoburgen), Hoburgen Lighthouse (A912SF).
Regional occurrence
The reader is referred to Vergoossen (Citation1999b) and Vergoossen (Citation2003b: ch. 5) for distribution of G. sandelensis and G. volborthi (including Gotland material), as well as the taxonomical comments by Valiukevičius (Citation2003b) mentioned above.
Family Poracanthodidae VERGOOSSEN, Citation1997
Genus Radioporacanthodes Vergoossen, Citation1999c
Diagnosis based on Vergoossen (Citation1999c). Poracanthodids having porosiform pore-canal system, which is less dense than punctatiforms. Scales may have large pores on the crown surface arranged per growth zone over radial canals. Rare arcade canals connecting few of the radial canals and sending up pore canals to the surface. Arcade canals not interconnected. Chains of pore canals may replace radial canals. New rows of pores may be inserted between existing ones. Flank scales grew superpositionally or combined superpositional and appositional growth. The growth zones generally form a diamond-shaped or, posteriorly, a zigzag pattern.
Radioporacanthodes spp. (–)
Material
Two “forma bifurcata” scales in sample G02-139LJ and four “pinaceus” scales in sample A912SF.
Description
Poracanthodids are only represented by two generally poorly preserved morphotypes in the material presented. The first morphotype is similar to the “pinaceus” variant described by Vergoossen (Citation1999b). These have a rhomboidal base, which is broader than long and anteriorly bulging. The neck is high and concave all around with vertical slits on the posterior side, and sometimes pores along the anterior side (). The crown is flat, rounded, and broader than long, and has small riblets along the anterior edge (). The crown has a zigzag pattern of slits producing several crown layers with several sharp posterior points (). The second morphotype is similar to the “forma bifurcata” of Vergoossen (Citation1999b) and is represented by two scales, but the extremely poor preservation of both has obliterated many of the crown features. However, the typical deep radial grooves on the crown are still evident, and the unevenly arcade slits can also be distinguished (). The base of these scales is similar to that in the “pinaceus” variant.
Remarks
Vergoossen (Citation1997, Citation1999b, Citation2000) separated several scale variants from Radioporacanthodes porosus Brotzen Citation1934, including the “forma bifurcata” and the “pinaceus” variant, in conjunction with his revision of the genus. However, no new taxa were formally erected for them and it remains unclear whether they truly represent separate taxa or merely variations within the squamation of a poracanthodid acanthodian (Burrow et al. Citation1999). Nevertheless, an articulated specimen of Radioporacanthodes menneri (Valiukevičius Citation1992) displays a strikingly uniform squamation, giving support to the separation of different scale forms.
Occurrence of Poracanthodids on Gotland
Burgsvik Formation (Fredholm Citation1989; Eriksson et al. Citation2009), Hamra Formation (Fredholm Citation1989; Eriksson et al. Citation2009). Lower Hamra Formation: Skradarve 1 (G02-139LJ). Unclear stratigraphic position: Hoburgen Lighthouse (A912SF).
Regional occurrence. The reader is referred to Vergoossen (Citation1999b) for the distribution of the different form groups separated from (Radio)poracanthodes porosus s.l.
Other acanthodian remains (–)
Figure 11. A–C. Indeterminable acanthodian scales; A. NRM-PZ P16366 from sample A989SF; B. NRM-PZ P16367 from G00-26LJ; C. NRM-PZ P16368 from A912SF. D. Acanthodian tessera; PMU 23103 from AM1. A1, B1, C1, and D in external view, A2, B2 and C2 in basal view with anterior facing upward. Scale bar equals 1 mm.

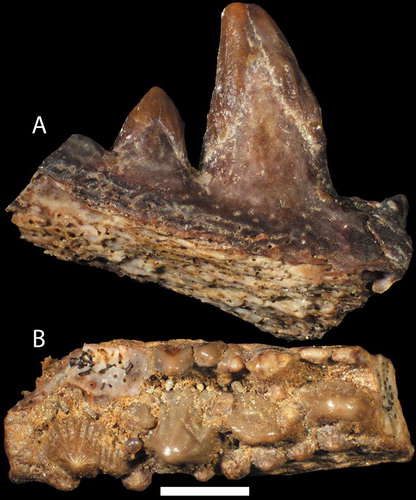
Figure 12. A–B. Two large ischnacanthiform jaw fragments; A. NRM-PZ P16369 in lingual view; B. NRM-PZ P16370 viewed from the top with anterior facing left. Both elements come from sample G00-2LJ. Scale bar represents 1 mm.
In total, five special acanthodian scales have been found in samples A 898 SF (Hoburgen), A 912 SF (Hoburgen Lighthouse), G00-26LJ (Ängvards 7). Three of them look like the one in , while two of them are slightly different (). They are similar to the umbellate and eigenartige scales that Gross (Citation1971: pl. 5, figs.14–25) described for N. striata and G. sandelensis respectively. Another large scale of uncertain affinity () has a wide and smoothly rounded crown and a restricted neck with vertical slits on the posterior side. The slightly tumid base is also wider than long, being outstretched rhomboidal. There are also a few tesserae of climatiid affinity in the material ().
Ischnacanthiforms are also represented by larger fragments such as pieces of teeth, jaws, and fin spines. The two largest, and best preserved, jaw fragments were found in the G00-2LJ sample from upper Sundre Formation at the Salmunds 1 locality. Specimen NRM-PZ P16369 preserves part of the jawbone with one large and one small tooth (). Both teeth are capped by a shiny tissue, presumably composed of orthodentine (Denison Citation1979). The second jaw fragment, specimen NRM-PZ P16370 (), has one main tooth row with individual teeth getting larger anteriorly. The lateral edge of the jawbone has a straight row of unevenly sized teeth, and there is a slanting row of peg-like teeth on the lingual side as well.
Discussion
Age of the Hamra and Sundre formations and their correlations with Estonia
Taxonomic concepts employed by Jeppsson (Citation2005b) and Jeppsson et al. (Citation2006) in their conodont zonation on Gotland has been revised and expanded (Miller & Aldridge Citation1997; Viira & Aldridge Citation1998; Slavík & Carls Citation2012). The original material of Jeppsson remains to be revised in the light of the recent taxonomic refinements, since he did not describe or illustrate the key taxa.
In the correlation between Gotland and Estonia by Jeppsson et al. (Citation1994), the majority of Uduvere Beds in Estonia was correlated with the previously used, informal Hemse unit d of the Hemse Group, and the Tahula Beds with Hamra unit b. The first correlation was based on the absence of Oz. snajdri in both units, and the presence of Coryssognathus dubius (there “Dentacodina dubia”), which has a truncated range on Gotland compared to other areas. It was also based on the assumption that Andreolepis hedei was absent from the Uduvere Beds, which has been shown to be wrong (Märss Citation1986: fig. 41; Märss & Männik Citation2013). Later works placed the Uduvere Beds in the Andreolepis hedei Vertebrate Zone (VZ), in which the När Formation (upper part of old unit d and unit e) on Gotland is also placed (Märss & Männik Citation2013). The introduction of carbon isotope stratigraphy, based on detailed biostratigraphy, permitted a more precise and accurate subdivision of beds. In this framework, the Hamra and Sundre formations correspond to the lowering limb of the Lau Event, or Mid-Ludfordian Carbon Isotope Excursion (MLCIE), and the Torgu Beds (now in the rank of Formation) is placed above it and at the level of a smaller, late Ludfordian excursion (Kaljo et al. Citation2015). However, carbon isotope data from Estonia in this interval is derived from the Ohesaare core, which represents a platform slope setting (Kaljo & Martma Citation2006), and therefore the corresponding interval in the core is much thinner than in the epiplatform sections, from which most of the conodont data is derived (e.g., Jeppsson et al. Citation1994; Viira & Aldridge Citation1998). As a result, the δ13Ccarb development within the Uduvere Beds and their correlation with Gotland remains somewhat undefined. The most recent revision of conodont stratigraphy in Estonia placed the Uduvere Beds in the Oz. snajdri Zone and the Tahula Beds in the Ozarkodina remscheidensis baccata/Ozarkodina snajdri parasnajdri Zone, which spans into the Kudjape Beds of Kuressaare Stage, upper Ludfordian, and is only regionally distinguished in Estonia (Männik Citation2014). As noted earlier, Oz. snajdri has a long stratigraphic range on Gotland and in Estonia (Jeppsson Citation1983; Jeppsson et al. Citation1994; Märss & Männik Citation2013). In the Ventspils-D3 core from Latvia, O. snajdri s.l. (Walliser Citation1964) occurs in Ventspils and lower part of Minija formations (Märss & Männik Citation2013: ). Also for Oz. crispa, Kaljo et al. (Citation2015) noted its occurrence stratigraphically much lower in Estonia than elsewhere, i.e., in the Sauvere and Himmiste beds, which are correlated with the Hemse Group on Gotland, as well as in the Uduvere Beds. These authors also proposed that Oz. crispa expanded its range into deeper water facies, corresponding to the Kuressaare Formation in Estonia and to the Ventspils Fm. in the Ventspils core from Latvia. Shifts in facies may then explain its higher FAD in this core compared to T. sculptilis, which displays uniform FADs across cores according to Kaljo et al. (Citation2015). The LAD of Oz. crispa is recorded at the base of the Minija Fm. in the Ventspils core, which is correlated with lowermost Přídolí. Consequently, resolving the positions of the Uduvere and Tahula beds with respect to the MLCIE would require isotope data tied to conodont occurrences from periplatform settings in Estonia and perhaps a numerical analysis of vertebrate and isotope occurrences, e.g., through constrained optimization (Sadler Citation2012). Also on Gotland, the Hamra and Sundre formations are not readily distinguished based on conodont composition and the index taxon Oz. crispa is found in both, although much more commonly in the latter. The slight shift in the overall conodont communities () is not due to the presence or absence of any particular species, but rather a gradual shift that may represent either gradual evolutionary replacement of taxa through time or shifting along an environmental gradient. The fact that localities assigned to the youngest strata show higher δ13Ccarb values than expected in this study supports the notion that the boundary between the two formations may be diachronous, and that they represent two nearly coeval types of lithologies and depositional environments.
The Hamra and Sundre formations also show differences in the composition of their vertebrate faunas. The seemingly environmentally restricted thelodont P. ludlowiensis (see above) occurs together with osteostracan fragments cf. Tahulapsis in the lower Hamra Formation. However, these taxa are not found in samples from the reef-associated limestones of the upper Hamra and Sundre formations, while scales identified as T. sculptilis are more commonly recovered from these strata. This agrees with studies on the ecology of thelodonts, where P. ludlowiensis was reported as occurring in restricted marine, or even freshwater environments, while T. sculptilis mainly lived in more open marine environments (Turner Citation1999; Ferrón et al. Citation2018), although ranging from lagoonal to the slope belt (Märss Citation1986). Furthermore, the latter taxon has been suggested as having a demersal lifestyle on hard substrates based on functional analyses of its scales (Ferron & Botella Citation2017). This further supports the notion that the faunal shift in this interval is the result of changing environments, especially if the reports of T. sculptilis from the older Burgsvik Formation is taken into account (Fredholm Citation1989; Eriksson et al. Citation2009).
Comparisons to Silurian vertebrate faunas outside of Gotland
The early vertebrate remains in samples from two different sections of the upper Słupianka Member (Winnica Formation) in the Holy Cross Mountains, Poland, were described in Bremer et al. (Citation2017). These strata have been suggested as largely coeval with the Hamra and Sundre formations on Gotland based on chemo- and sequence stratigraphy, but they were deposited on the opposite side of the Baltic Basin (Kozłowski & Munnecke Citation2010). A sample from the Winnica site contained thelodonts Thelodus parvidens, T. trilobatus, and anaspid fragments similar to Liivilepis. The range of “nostolepid” and “gomphonchid” acanthodian scales were similar to those described here, but the poracanthodid scale-variants were more abundant (Bremer et al. Citation2017). The other sample came from an oolitic layer of potentially slightly younger age than the Sundre Formation at the top of the Rzepin section, and displayed a more similar fauna to Gotland regarding abundance of acanthodians, as well as a few fragments of an osteostracan similar to Tahulapsis ordinata (Bremer et al. Citation2017). Unlike Hamra and Sundre, however, this sample was rich in scales of Paralogania ludlowiensis, possibly as a result of the depositional setting of the sampled layer (see Bremer et al. Citation2017). Polish vertebrate remains were also listed from 40 samples of drill cores in northern and eastern Poland by Märss (Citation1997). The listed Silurian taxa were probably younger than the ones described from Gotland and the Winnica Formation, since they contained a fauna typical of the upper Kaugatuma and lower Ohesaare stages in East Baltic (Märss Citation1997). The lower part of the Miastko-1 drill core contained scales of T. sculptilis, which is typical for older faunas, but these were suggested to have been re-deposited (ibid.).
The vertebrate biostratigraphy of the East Baltic is much better investigated and described, but there is a hiatus between the Uduvere Beds of the Paadla Regional Stage (RS) and the Tahula Beds of lower Kuressaare RS in Estonia and Latvia, largely corresponding to the O. crispa conodont Zone sensu Walliser (Citation1964) according to Märss and Männik (Citation2013). The Hamra and Sundre formations may therefore represent part of this hiatus. The Uduvere Beds include Thelodus parvidens, T. carinatus, T. laevis, Paralogania martinssoni, Phlebolepis elegans, osteostracan Dartmuthia procera, acanthodians Nostolepis striata and Gomphonchus sandelensis, heterostracan Archegonaspis? sp., as well as Andreolepis hedei (Märss Citation1986, Citation1992; Märss et al. Citation2007, Citation2014), which agrees well with the taxonomic composition of vertebrate faunas (besides T. laevis and D. procera) from the När Formation on Gotland (Fredholm Citation1988a, Citation1988b). Both the Uduvere Beds and the När Formation are placed in the Andreolepis hedei VZ (Märss & Männik Citation2013), which is in the rising limb of the MLCIE (Lau Event) on Gotland (Eriksson & Calner Citation2008). However, the Uduvere Beds are placed in the O. snajdri CZ, which on Gotland includes the Burgsvik and Hamra formations (Jeppsson et al. Citation2006), i.e., in the declining limb of the MLCIE (Eriksson & Calner Citation2008). On Gotland, A. hedei ranges from slightly below the Pol. siluricus CZ (Fredholm Citation1988b) to the lower part of O. snajdri CZ in the Burgsvik Sandstone Member (Märss Citation1992, Citation2001; Märss & Männik Citation2013), but the associated faunal list for the Burgsvik Formation is quite different compared to the list for the typical A. hedei VZ described in Märss and Männik (Citation2013). However, Märss (Citation1992, Citation2001) described two different faunal associations for A. hedei: one older with Phl. elegans and one younger with T. sculptilis or P. ludlowiensis. Vergoossen (Citation1999a) also described A. hedei together with T. sculptilis from strata probably of comparable age in southern Sweden. Therefore, it is possible that both these associations are present on Gotland, with the lower one agreeing better with the fossil fauna of the Uduvere Beds.
According to Kaljo and Martma (Citation2006) and Kaljo et al. (Citation2015), the Sundre Formation probably only reaches a level equivalent to the lowermost Tahula Beds of the Kuressaare RS. The Tahula Beds contain vertebrates also present at the top of the Gotland sequence (T. parvidens, T. sculptilis, extremely rare P. ludlowiensis, Loganellia cuneata, Tahulalepis elongituberculata, Tah. ordinata, N. striata, G. sandelensis, and poracanthodid acanthodians), but also several additional taxa not seen on Gotland (Märss Citation1986, Citation1992; Blom et al. Citation2002; Märss Citation2006; Märss et al. Citation2007, Citation2014).
The Pagėgiai Formation in Lithuania is placed within the O. crispa CZ and displays thelodonts containing T. parvidens, T. trilobatus, T. sculptilis, and P. ludlowiensis, but also the younger Thelodus admirabilis (Märss Citation1982) as well as heterostracans (Karatajūtė-Talimaa & Brazauskas Citation1994). There are also a number of mainly “nostolepid” and “gomphonchid” acanthodian taxa (see Valiukevičius Citation2006). Overall, this fauna is similar, but precise comparisons are difficult based on literature.
On Gotland, numerous acanthodian remains first appear in the När Formation, but they are only represented by “nostolepids” and “gomphonchids” (Fredholm Citation1988a, Citation1988b). These are joined by poracanthodid acanthodians in the younger Burgsvik Formation (Fredholm Citation1989; Eriksson et al. Citation2009). In the East Baltic, “gomphonchid” remains have been reported from the much older, upper Llandovery strata of Adavere RS, and rare “gomphonchid” and “nostelpid” remains are scattered throughout the Wenlock in the Jaani, Jaagarahu, and Rootsiküla stages (Märss Citation1986), but they remain uncommon until the Paadla RS (Märss Citation1986; Kaljo & Märss Citation1991). Valiukevičius (Citation2006) reported the oldest appearance of acanthodians in the lower Wenlock of Lithuania, as well as a diversification in the mid-Ludfordian of the Pagėgiai Formation (dominated by “nostolepids”). Poracanthodids are first found in the Tahula Beds of the lower Kaugatuma RS in the East Baltic (Märss Citation1986) and in the post-Ludlow Minja Formation in Lithuania (Valiukevičius Citation2006). The acanthodian diversity eventually becomes a dominant part of the Silurian vertebrate faunas in the lower Přídolí Kaugatuma RS (Märss & Einasto Citation1978; Kaljo & Märss Citation1991; Valiukevičius Citation2006), which Märss (Citation1992) called the “acanthodian event”. The appearance of poracanthodid acanthodians in the Burgsvik Formation presented in previous studies, and rare occurrences in the Hamra and Sundre formations presented here, may then reflect the early stages of this acanthodian diversification. The rarity of poracanthodids also in the youngest beds on Gotland may be related to facies, since poracanthodid remains have been suggested as a deeper water equivalent to several vertebrate biozones by Märss and Männik (Citation2013).
The taxonomic state of Silurian acanthodians from the Baltic Basin
Our understanding of the interrelationships of early jawed vertebrates has changed during the last decade (see Brazeau and Friedman (Citation2015) and Burrow et al. (Citation2016) for more thorough discussions). Brazeau (Citation2009) and Davis et al. (Citation2012) showed a para- or polyphyletic “Acanthodii” with representatives ending up on the chondrichthyan, osteichthyan, and gnathostome stems, while Zhu et al. (Citation2013) and Long et al. (Citation2015) resolved all acanthodians on the chondrichthyan stem. Dupret et al. (Citation2014), Giles et al. (Citation2015) and Burrow et al. (Citation2016) also placed the acanthodians within the chondrichthyan total group, but with varying relationships within the clade. With these changing views in mind, the term “acanthodians” are herein used in its broadest sense, mainly for practical reasons.
The works of Vergoossen (Citation1997, Citation1999a, Citation1999b, Citation1999c, Citation2000, Citation2002a, Citation2002b, Citation2002c, Citation2003a, Citation2003b, Citation2004) and Valiukevičius (Citation1998, Citation2003a, Citation2003b, Citation2004a, Citation2004b) scrutinised the taxonomy and systematics of acanthodian scale-taxa from the western and eastern parts of the Baltic Sea region respectively. Both researchers emphasised the problem that scale-based definitions for several acanthodian taxa had become too broad, since the morphological range of scales assigned to a single taxon had expanded as more material was described and referred to it (see Vergoossen Citation2002b; Valiukevičius Citation2003a). As a result, one taxon would eventually display exaggerated temporal and geographical ranges, as well as incorporate more than one biological species and vice versa. Despite these previous efforts, our understanding of acanthodian scale taxonomy is still in need of review and revision, but this is beyond the scope of this study.
The uncertainties discussed above have implications for this work, because the samples treated here seem to reflect an early diversification of acanthodians in the Silurian Baltic Basin, as previously demonstrated by Kaljo and Märss (Citation1991) and Valiukevičius (Citation2006). The acanthodian scales presented here are placed in the generalized “nostolepid”, “gomphonchid”, and poracanthodid groups. Scales of Nostolepis-type are referred to climatiid acanthodians (Denison Citation1979; Valiukevičius & Burrow Citation2005), and “gomphonchids” and poracanthodids are respectively placed in the families Ischnacanthidae Woodward Citation1891 and Poracanthodidae Vergoossen Citation1997, within the order Ischnacanthiformes Berg Citation1940 (see Burrow Citation2013). Vergoossen (Citation1997) saw it necessary to revise the genus Poracanthodes Brotzen Citation1934 when it was discovered that it incorporated scales of two main histological types: punctatiform and porosiform histology. Vergoossen (Citation1999c) divided the genus into the genera Poracanthodes, Radioporacanthodes Vergoossen Citation1999c, Zemlyacanthus Vergoossen Citation1997, and Gomphonchoporus Vergoossen Citation1999c. The latter was erected when scales combining poracanthodid- and Gomphonchus-type histology were discovered (see Vergoossen Citation1999c and references therein). Vergoossen (Citation1999c) therefore separated Gomphonchus hoppei from the family Ischnacanthidae Woodward Citation1891 and instead placed it in the new genus Gomphonchoporus within the family Poracanthodidae Vergoossen Citation1997. This also led to a change in the definition of Poracanthodidae (Vergoossen Citation1999c), which was later seemingly supported by Valiukevičius (Citation2004a) and Burrow (Citation2013). The separation of punctatiform from non-punctatiform acanthodians also led to the recognition that many of the scale-based definitions within these taxa were too broad. Vergoossen (Citation2002a) reinstated and separated R. biblicus from R. porosus and revised several previous assignments of the latter to R. biblicus or other forms (see Vergoossen Citation1999c). Some of these re-assignments were later supported by Burrow (Citation2003). The scales reported as “forma bifurcata” and “pinaceus” variant in this work were viewed as belonging to a porosiform poracanthodid scale group by Vergoossen (Citation1997), and according to Burrow et al. (Citation1999) they probably represent sets of specialized scales or possibly different species, but their exact affinities remain unclear.
Three families of ischnacanthiforms were recognised by Burrow (Citation2013): Ischnacanthidae, Poracanthodidae, and Acritolepidae. Burrow (Citation2013) did, however, stress that only a few of these are represented by articulated specimens that are all of Lochkovian age. The genus Zemlyacanthus was erected by Vergoossen (Citation1997) for the articulated porosiform poracanthodid of early Devonian age, previously described as “Poracanthodes” menneri by Valiukevičius (Citation1992). Vergoossen (Citation1997) did this because all scales from this specimen only show appositional growth, while all other porosiforms have superpositional growth or a combination of both (Burrow Citation2013). However, Burrow (Citation2013) still postulated that the species should be referred to Radioporacanthodes based on overall morphological and histological similarities, thereby rendering Zemlyacanthus invalid.
Valiukevičius (Citation2003a, Citation2003b, Citation2004a, Citation2004b) largely focused on the “nostolepid” type of scales in his research, and erected many new scale-based taxa by extracting scale types previously assigned to N. striata, while stressing the un-realistic longevity and excessively diverse scale forms of this taxon. This was done in conjunction with investigations of their potential biostratigraphical importance (Valiukevičius Citation1998, Citation2005), which led to several stratigraphically constrained taxa. Valiukevičius (Citation2003a, Citation2003b, Citation2004a, Citation2004b) also put emphasis on differences in scale histology, and Valiukevičius and Burrow (Citation2005) even considered histology of primary importance for taxonomic designations of isolated scales.
Zidek (Citation1985) demonstrated that differences in the squamation of Acanthodes resulted from the scale-cover spreading from posterior to anterior along the body and from the lateral lines. This was also presented by Brazeau (Citation2012) as a possible explanation for differences in the morphology and histology of scales covering the body of the early gnathostome Ptomachanthus anglicus Miles Citation1973. By analysing 188 complete specimens of Triazeugacanthus Miles Citation1966, Chevrinais et al. (Citation2017) investigated the ontogeny of the squamation cover in this acanthodian. They discovered an initiation-area below the dorsal fin spine, from which the scale-cover spread anterior and posterior through ontogeny until it covered almost the entire animal in adult specimens. Another initiation-area was observed along the dorsal midline of the body, where scales also fused in later stages (Chevrinais et al. Citation2017). It was clear that scale morphology differed between young and adult individuals, as well as between old and new scales within the same individual, as a result of this spreading scale-cover (Chevrinais et al. Citation2017). Valiukevičius and Burrow (Citation2005) mentioned that the growth lamellae do not reflect the developmental stage of the animal, but Chevrinais et al. (Citation2017) supported the suggestion of Zidek (Citation1985) and Karatajūtė-Talimaa (Citation1998) that growth of individual scales is correlated with species ontogeny.
When examined in more detail, many articulated acanthodian-type taxa and other early gnathostomes display a wide range of scale morphologies covering the body (Trinajstic Citation2001; Brazeau Citation2012; Burrow Citation2013; Burrow et al. Citation2013, Citation2015, Citation2016), but apparently to differing degrees (e.g., R. menneri in Valiukevičius Citation1992). If the morphology of different scale-types from different parts of the body is reflected in the way they grew, then morphologically similar scales (with a presumed similar function) but widely different histology may support the importance of histology in scale-based taxonomy as pointed out by Valiukevičius and Burrow (Citation2005). Trinajstic (Citation2001) demonstrated that isolated scales can be identified taxonomically, and even be determined to certain regions of the body, when compared to articulated specimens. Unfortunately, few articulated Silurian specimens have been found and our understanding of their squamation is therefore limited (Burrow Citation2013).
Reviewing assemblages of acanthodian scales (both morphologically and histologically) from collections of different geographical areas in a stratigraphically constrained framework, as well as comparing them to squamation patterns in articulated specimens, may help to revise the taxonomy of Silurian acanthodians from the Baltic Basin. This could potentially bridge some of the discrepancies between the previous works as described above. Similar efforts have proven extremely fruitful for other early vertebrates, such as thelodonts (see Märss et al. Citation2007), anaspids (Blom et al. Citation2002), and osteostracans (Märss et al. Citation2014).
Conclusions
The conodont and isotope data presented here give some support to the view that the traditional Hamra and Sundre formations may represent two largely isochronous units, which was suggested by Jeppsson (Citation2005b). However, the composition of additional samples in Jeppsson’s collection at the NRM needs to be investigated before anything conclusive can be said. Furthermore, additional sampling for isotopes may also help elucidate the relationship between the two formations.
Correlation of the youngest strata on Gotland to other areas in the Baltic Basin is problematic because many of the conodont taxa used in previous correlations have been shown to have longer ranges than previously thought. The placement of the East Baltic Uduvere Beds as largely coeval to the När Formation on Gotland is supported by isotope data and similarities in vertebrate faunas. It is, however, in conflict with the placement of Uduvere Beds in the O. snajdri CZ compared to Gotland. However, as previously mentioned, this conodont has a longer range both on Gotland and in the East Baltic, possibly in relation to environmental conditions. On the other hand, the differences in isotope values may also result from depositional variations between sections, and the vertebrate index taxon Andreolepis hedei also occur in younger strata on Gotland. The relation of Hamra and Sundre formations to the Tahula Beds in East Baltic is also problematic, but it seems likely that the latter are slightly younger.
Regardless of the issues outlined above, the younger parts of the Gotland stratigraphy see an increase in acanthodian diversity, which is in line with previously observed patterns in similarly aged strata in other parts of the Baltic Basin and beyond. The precise mapping of these diversity patterns is partly hindered by current issues with the scale-based taxonomy of Silurian acanthodians from the Baltic Sea region. However, an increased understanding of the squamation of early gnathostomes may enable comparisons of distinct scale-types to articulated specimens, and develop a scheme based on scale-sets similar to those for thelodonts, anaspids, and osteostracans. This, combined with detailed morphological and histological investigations in a geographically and stratigraphically constrained framework, may be the way forward when dealing with the taxonomy of isolated acanthodian scales.
Acknowledgments
The authors would like to thank Henning Blom for providing feedback during this work and for the processing of samples under his project funding. We would also like to thank Thomas Mörs for giving us access to the collections at the NRM and for lending us material, Jan Ove Ebbestad for giving us access to the collections and the photography setup at PMU, as well as Michael Streng (Uppsala University) for aiding us with the SEM. Last, but not least, we want to thank Linda Wickström (SGU) for giving access to map material and data for producing the maps in this work, and the reviewers for their helpful comments. The fieldwork and collection of new samples for this study was supported by Helge Ax:son Johnsons stiftelse. This is Paleobiology Database publication 345.
Disclosure statement
No potential conflict of interest was reported by the authors.
References
- Agassiz, L., 1838: On the fishes of the ludlow rocks, or upper beds of the Silurian system. In R.I. Murchisson (ed.): Report of the British association for the advancement of science, 7th transactions, 91. Liverpool
- Aldridge, R.J. & Jeppsson, L., 1984: Ecological specialists among Silurian conodonts. Special Papers in Palaeontology 32, 141–149
- Berg, L.S., 1940: Classification of fishes, both recent and fossil. Trudy Zoologicheskogo Instituta, Akademiia Nauk SSSR 5, 346–517
- Bischoff, G. & Sannemann, D., 1958: Unterdevonische Conodonten aus dem Frankenwald. Notizblatt des Hessischen Landesamtes für Bodenforschung zu Wiesbaden 86, 87–110
- Blom, H., Märss, T. & Miller, C.G., 2002: Silurian and earliest Devonian birkeniid anaspids from the Northern Hemisphere. Earth and Environmental Science Transactions of the Royal Society of Edinburgh 92 (03), 263–323. doi:10.1017/S0263593300000250
- Branson, E.B. & Mehl, M.G., 1933: Conodonts from the Bainbridge (Silurian) of Missouri. University of Missouri Studies 8, 39–52
- Brazeau, M.D., 2009: The braincase and jaws of a Devonian ‘acanthodian’ and modern gnathostome origins. Nature 457 (7227), 305–308. doi:10.1038/nature07436
- Brazeau, M.D., 2012: A revision of the anatomy of the early Devonian jawed vertebrate Ptomacanthus anglicus miles. Palaeontology 55 (2), 355–367. doi:10.1111/j.1475-4983.2012.01130.x
- Brazeau, M.D. & Friedman, M., 2015: The origin and early phylogenetic history of jawed vertebrates. Nature 520 (7548), 490–497. doi:10.1038/nature14438
- Bremer, O., 2017: Silurian vertebrates of Gotland (Sweden) and the Baltic Basin, 61. Acta Universitatis Upsaliensis, Uppsala University, Uppsala
- Bremer, O., Niedźwiedzki, G., Blom, H., Dec, M. & Kozłowski, W., 2017: Vertebrate microremains from the upper Silurian winnica formation of the Holy Cross Mountains, Poland. Geological Magazine 155 (7), 1523–1541. doi:10.1017/S0016756817000681
- Brotzen, F., 1934: Erster Nachweis von Unterdevon im Ostseegebiete durch Konglomeratgeschiebe mit Fischresten. II Teil (Paläontologie). Zeitschrift für Geschiebeforschung 10, 1–65
- Burrow, C.J., 2003: Poracanthodid acanthodian from the Upper Silurian (Pridoli) of Nevada. Journal of Vertebrate Paleontology 23 (3), 489–493. doi:10.1671/1888
- Burrow, C.J., 2013: Reassessment of Ischnacanthus? scheii Spjeldnaes (Acanthodii, Ischnacanthiformes) from the latest Silurian or earliest Devonian of Ellesmere Island, arctic Canada 1. Canadian Journal of Earth Sciences 50 (9), 945–954. doi:10.1139/cjes-2013-0068
- Burrow, C.J., Davidson, R.G., Den Blaauwen, J.L. & Newman, M.J., 2015: Revision of Climatius reticulatus Agassiz, 1844 (Acanthodii, Climatiidae), from the Lower Devonian of Scotland, based on new histological and morphological data. Journal of Vertebrate Paleontology 35 (3), 1–15. doi:10.1080/02724634.2014.913421
- Burrow, C.J., Den Blaauwen, J., Newman, M. & Davidson, R., 2016: The diplacanthid fishes (Acanthodii, Diplacanthiformes, Diplacanthidae) from the Middle Devonian of Scotland. Palaeontologia Electronica 19 (1), 1–83
- Burrow, C.J., Newman, M.J., Davidson, R.G. & Den Blaauwen, J.L., 2013: Redescription of Parexus recurvus, an early Devonian acanthodian from the Midland Valley of Scotland. Alcheringa: an Australasian Journal of Palaeontology 37 (3), 392–414. doi:10.1080/03115518.2013.765656
- Burrow, C.J., Vergoossen, J., Turner, S., Uyeno, T.T. & Thorsteinsson, R., 1999: Microvertebrate assemblages from the Upper Silurian of Cornwallis Island, Arctic Canada. Canadian Journal of Earth Sciences 36 (3), 349–361. doi:10.1139/e98-098
- Chevrinais, M., Sire, J.-Y. & Cloutier, R., 2017: From body scale ontogeny to species ontogeny: Histological and morphological assessment of the late Devonian acanthodian Triazeugacanthus affinis from Miguasha, Canada. PLoS One 12 (4), 1–31. doi:10.1371/journal.pone.0174655
- Corradini, C., Corriga, M.G., Männik, P. & Schönlaub, H.P., 2015: Revised conodont stratigraphy of the Cellon section (Silurian, Carnic Alps). Lethaia 48 (1), 56–71. doi:10.1111/let.12087
- Cramer, B.D., Brett, C.E., Melchin, M.J., Männik, P., Kleffner, M.A., Mclaughlin, P.I., Loydell, D.K., Munnecke, A., Jeppsson, L., Corradini, C., Brunton, F.R. & Saltzman, M.R., 2011: Revised correlation of Silurian provincial series of North America with global and regional chronostratigraphic units and δ13Ccarb chemostratigraphy. Lethaia 44 (2), 185–202. doi:10.1111/let.2011.44.issue-2
- Davis, S.P., Finarelli, J.A. & Coates, M.I., 2012: Acanthodes and shark-like conditions in the last common ancestor of modern gnathostomes. Nature 486 (7402), 247–250. doi:10.1038/nature11080
- Denison, R.H., 1979: Acanthodii. Pt. 5, 62. Gustav Fischer Verlag, Stuttgart
- Drygant, D., 1984: Correlation and conodonts of the Silurian – lower Devonian deposits of Volyno-Podolia, Naukova Dumka, Kiev
- Dupret, V., Sanchez, S., Goujet, D., Tafforeau, P. & Ahlberg, P.E., 2014: A primitive placoderm sheds light on the origin of the jawed vertebrate face. Nature 507 (7493), 500–503. doi:10.1038/nj7493-523a
- Eriksson, M.E. & Calner, M., 2005. The dynamic Silurian earth. Sveriges geologiska undersökning Report 99.
- Eriksson, M.E. & Calner, M., 2008: A sequence stratigraphical model for the Late Ludfordian (Silurian) of Gotland, Sweden: implications for timing between changes in sea level, palaeoecology, and the global carbon cycle. Facies 54 (2), 253–276. doi:10.1007/s10347-007-0128-y
- Eriksson, M.E., Nilsson, E.K. & Jeppsson, L., 2009: Vertebrate extinctions and reorganizations during the late Silurian Lau event. Geology 37 (8), 739–742. doi:10.1130/G25709A.1
- Erlström, M., Persson, L., Sivhed, U. & Wickström, L., 2009: Beskrivning till regional berggrundskarta över Gotlands län. Sveriges geologiska undersökning K 221.
- Ferron, H.G. & Botella, H., 2017: Squamation and ecology of thelodonts. PLoS One 12 (2), e0172781. doi:10.1371/journal.pone.0172781
- Ferrón, H.G., Martínez‐Pérez, C., Turner, S., Manzanares, E. & Botella, H., 2018: Patterns of ecological diversification in thelodonts. Palaeontology 61 (2), 303–315. doi:10.1111/pala.12347
- Fredholm, D., 1988a: Vertebrates in the Ludlovian Hemse Beds of Gotland, Sweden. GFF 110 (2), 157–179
- Fredholm, D., 1988b: Vertebrate biostratigraphy of the Ludlovian Hemse Beds of Gotland, Sweden. GFF 110 (3), 237–253
- Fredholm, D., 1989: Silurian vertebrates of Gotland, Sweden. Ph. D. Lund University, Lund, Sweden. 47 pp.
- Fredholm, D., 1990: Agnathan vertebrates in the lower Silurian of Gotland, Sweden. GFF 112 (1), 61–80
- Giles, S., Friedman, M. & Brazeau, M.D., 2015: Osteichthyan-like cranial conditions in an early Devonian stem gnathostome. Nature 520 (7545), 82–85. doi:10.1038/nature14065
- Gross, W., 1947: Die Agnathen und Acanthodier des obersilurischen Beyrichienkalks. Palaeontographica Abteilung A, 96, 91–158
- Gross, W., 1967: Über Thelodontier-Schuppen. Palaeontographica Abteilung A, 127, 1–67
- Gross, W., 1971: Downtonische und dittonische acanthodier-reste des ostseegebietes. Palaeontographica Abteilung A, 136,1–82
- Hadley, A., 2005: CombineZ version 5.1. URL
- Hede, J. E., 1925: Beskrivning av Gotlands silurlager. In H. W. Munthe, J. E. Hede & L. von Prost (eds.): Gotlands geologi. En översikt, 13–30. Sveriges Geologiska Undersökning C331, Stockholm, Sweden.
- Hede, J. E., 1960: The Silurian of Gotland. In G. Regnéll & J. E. Hede (eds.): The Lower Palaeozoic of Scania. The Silurian of Gotland. Guide to Excursions Nos A 22 and C 17, 44–87. Sveriges Geologiska Undersökning, Stockholm, Sweden.
- Hede, J.E., 1921: Gottlands Silurstratigrafi. Sveriges Geololgiska Undersökning C305, 1–100
- Hoppe, K.-H., 1931: Die Coelolepiden und Acanthodier des Obersilurs der Insel Ösel. Ihre Paläobiologie und Paläontologie. Palaeontographica (1846-1933), 76, 35–94
- Jaekel, O., 1911: Die Wirbeltiere; eine Übersicht über die fossilen und lebenden Formen, 252. Gebrüder Borntraeger, Berlin
- Jarochowska, E., Bremer, O., Heidlas, D., Pröpster, S., Vandenbroucke, T.R.A. & Munnecke, A., 2016a: End-wenlock terminal Mulde carbon isotope excursion in Gotland, Sweden: integration of stratigraphy and taphonomy for correlations across restricted facies and specialized faunas. Palaeogeography, Palaeoclimatology, Palaeoecology 457, 304–322
- Jarochowska, E., Munnecke, A., Frisch, K., Ray, D.C. & Castagner, A., 2016b: Faunal and facies changes through the mid Homerian (late Wenlock, Silurian) positive carbon isotope excursion in Podolia, western Ukraine. Lethaia 49 (2), 170–198. doi:10.1111/let.12137
- Jarochowska, E., Viira, V., Einasto, R., Nawrot, R., Bremer, O., Männik, P. & Munnecke, A., 2017: Conodonts in Silurian hypersaline environments: Specialized and unexpectedly diverse. Geology 45 (1), 3–6. doi:10.1130/G38492.1
- Jeppsson, L., 1972: Some Silurian conodont apparatuses and possible conodont dimorphism. Geologica et Palaeontologica 6, 51–69.
- Jeppsson, L., 1974: Aspects of late Silurian conodonts. Fossils and Strata 6, 1–54.
- Jeppsson, L., 1983: Silurian conodont faunas from Gotland. Fossils and Strata 15, 121–144.
- Jeppsson, L., 1989: Latest Silurian conodonts from Klonk, Czechoslovakia. Geologica et Palaeontologica 23, 21–37.
- Jeppsson, L., 2005a: Biases in the recovery and interpretation of micropalaeontological data. In Symposium on bias and completeness in the conodont fossil record held at the 8th International Conodont Symposium, 57–71. Toulouse, France.
- Jeppsson, L., 2005b: Conodont-based revisions of the Late Ludfordian on Gotland, Sweden. GFF 127 (4), 273–282. doi:10.1080/11035890501274273
- Jeppsson, L. & Anehus, R., 1995: A buffered formic acid technique for conodont extraction. Journal of Paleontology 69 (4), 790–794. doi:10.1017/S0022336000035319
- Jeppsson, L., Eriksson, M.E. & Calner, M., 2006: A latest Llandovery to latest Ludlow high-resolution biostratigraphy based on the Silurian of Gotland—a summary. GFF 128 (2), 109–114. doi:10.1080/11035890601282109
- Jeppsson, L., Viira, V. & Männik, P., 1994: Silurian conodont-based correlations between Gotland (Sweden) and Saaremaa (Estonia). Geological Magazine 131 (2), 201–218. doi:10.1017/S0016756800010736
- Kaljo, D., Einasto, R., Martma, T., Märss, T., Nestor, V. & Viira, V., 2015: A bio-chemostratigraphical test of the synchroneity of biozones in the upper Silurian of Estonia and Latvia with some implications for practical stratigraphy. Estonian Journal of Earth Sciences 64 (4), 267–283. doi:10.3176/earth.2015.33
- Kaljo, D. & Märss, T., 1991: Pattern of some Silurian bioevents. Historical Biology 5 (2–4), 145–152. doi:10.1080/10292389109380397
- Kaljo, D. & Martma, T., 2006: Application of carbon isotope stratigraphy to dating the Baltic Silurian rocks. GFF 128 (2), 123–129. doi:10.1080/11035890601282123
- Karatajūtė-Talimaa, V., 1998: Determination methods for the exoskeletal remains of early vertebrates. Mitteilungen ausdemMuseum für Naturkunde in Berlin, Geowissenschaftliche Reihe 1, 21–52.
- Karatajūtė-Talimaa, V. & Brazauskas, A., 1994: Distribution of vertebrates in the Silurian of Lithuania. Geologija 17, 106–114
- Kozłowski, W. & Munnecke, A., 2010: Stable carbon isotope development and sea-level changes during the Late Ludlow (Silurian) of the Łysogóry region (Rzepin section, Holy Cross Mountains, Poland). Facies 56 (4), 615–633. doi:10.1007/s10347-010-0220-6
- Lankester, E.R., 1868: A monograph of the fishes of the old red sandstone of Britain. Part I. The Cephalaspidae, 62. Monograph of the Palaeontographical Society, London
- Laufeld, S., 1974: Reference localities for palaeontology and geology in the Silurian of Gotland, 172. Sveriges Geologiska Undersökning, Stockholm
- Lehman, J.P., 1937: Les Poissons du Downtonian de la Scanie (Suède). Mémoire Faculté des Sciences de l’Université de Paris, 98.
- Lehnert, O., Frýda, J., Buggisch, W., Munnecke, A., Nützel, A., Křiž, J. & Manda, S., 2007: δ13C records across the late Silurian Lau event: New data from middle palaeo-latitudes of northern peri-Gondwana (Prague Basin, Czech Republic). Palaeogeography, Palaeoclimatology, Palaeoecology 245 (1–2), 227–244. doi:10.1016/j.palaeo.2006.02.022
- Long, J.A., Mark-Kurik, E., Johanson, Z., Lee, M.S., Young, G.C., Min, Z., Ahlberg, P.E., Newman, M., Jones, R., Den Blaauwen, J., Choo, B. & Trinajstic, K., 2015: Copulation in antiarch placoderms and the origin of gnathostome internal fertilization. Nature 517 (7533), 196–199. doi:10.1038/nature13825
- Männik, P., 2014: The Silurian system in Estonia. In H. Bauert, O. Hints, T. Meidla & P. Männik (eds.): 4th annual meeting of IGCP 591 The early to Middle Paleozoic revolution, Estonia 10-19 June 2014. Abstracts and field guide, 123–128. University of Tartu, Estonia.
- Märss, T., 1982: Thelodus admirabilis n. sp. (Agnatha) from the Upper Silurian of the East Baltic. Eesti NSV Teaduste Akadeemia Toimetised, 31. Geoloogia 3, 112–116
- Märss, T., 1986: Silurian vertebrates of Estonia and west Latvia, 104. Academy of Sciences of the Estonian SSR, Institute of Geology, Tallinn
- Märss, T., 1992: Vertebrate history in the Late Silurian. Proceedings of the Estonian Academy of Sciences, Geology 41 (4), 204
- Märss, T., 1997: Vertebrates of the Pridoli and Silurian-Devonian boundary beds in Europe. Modern Geology 21 (1), 17–42
- Märss, T., 2001: Andreolepis (Actinopterygii) in the upper Silurian of northern Eurasia. Proceedings of the Estonian Academy of Sciences, Geology 50 (3), 174–189
- Märss, T., 2006: Thelodonts (Agnatha) from the basal beds of the Kuressaare Stage, Ludlow, Upper Silurian of Estonia. Proceedings of the Estonian Academy of Sciences, Geology 55 (1), 43–66
- Märss, T., Afanassieva, O. & Blom, H., 2014: Biodiversity of the Silurian osteostracans of the East Baltic. Earth and Environmental Science Transactions of the Royal Society of Edinburgh 105 (2), 73–148. doi:10.1017/S1755691014000218
- Märss, T., Caldwell, M., Gagnier, P., Goujet, D., Männik, P., Martma, T. & Wilson, M., 1998: Distribution of silurian and lower devonian vertebrate microremains and conodonts in the baillie-hamilton and cornwallis island sections, Canadian Arctic. Proceedings of the Estonian Academy of Sciences, Geology 47 (2), 51–76.
- Märss, T. & Einasto, R., 1978: Distribution of vertebrates in deposits of various facies in the North Baltic Silurian. Eesti NSV Teaduste Akadeemia Toimetised Geoloogia 27, 16–22
- Märss, T. & Männik, P., 2013: Revision of Silurian vertebrate biozones and their correlation with the conodont succession. Estonian Journal of Earth Sciences 62 (4), 181–204. doi:10.3176/earth.2013.15
- Märss, T. & Miller, C.G., 2004: Thelodonts and distribution of associated conodonts from the Llandovery–lowermost Lochkovian of the Welsh Borderland. Palaeontology 47 (5), 1211–1265. doi:10.1111/j.0031-0239.2004.00409.x
- Märss, T. & Ritchie, A., 1998: Articulated thelodonts (Agnatha) of Scotland. Earth and Environmental Science Transactions of the Royal Society of Edinburgh 88 (3), 143–195. doi:10.1017/S026359330000691X
- Märss, T., Turner, S. & Karatajūtė-Talimaa, V., 2007: Agnatha II-Thelodonti, 143. Verlag Dr Friedrich Pfeil, Münich
- Märss, T., Wilson, M.V.H. & Thorsteinsson, R., 2006: Silurian and Lower Devonian thelodonts and putative chondrichthyans from the Canadian Arctic Archipelago, 144. The Palaeontological Association, London
- Melchin, M.J., Sadler, P.M. & Cramer, B.D., 2012: The Silurian Period. In F.M. Gradstein, J.G. Ogg, M. Schmitz & G. Ogg (eds.): The geologic time scale 2012, 525–558. Elsevier, Amsterdam
- Miles, R. S., 1973: Articulated acanthodian fishes from the old red sandstone of england, with a review of the structure and evolution of the acanthodian shoulder-girdle. Bulletin Of The British Museum (Natural History) 24, 111–213.
- Miles, R.S., 1966: The acanthodian fishes of the Devonian Plattenkalk of the paffrath trough in the Rhineland. Arkiv für Zoologi 18, 147–194
- Miller, C.G. & Aldridge, R.J., 1997: Ozarkodina rernscheidensis plexus conodonts from the upper Ludlow (Silurian) of the Welsh Borderland and Wales. Journal of Micropalaeontology 16, 41–49. doi:10.1144/jm.16.1.41
- Miller, C.G. & Märss, T., 1999: A conodont, thelodont and acanthodian fauna from the lower Přidoli (Silurian) of the Much Wenlock area, Shropshire. Palaeontology 42 (4), 691–714. doi:10.1111/1475-4983.00093
- Munnecke, A., Calner, M., Harper, D.A.T. & Servais, T., 2010: Ordovician and Silurian sea–water chemistry, sea level, and climate: A synopsis. Palaeogeography, Palaeoclimatology, Palaeoecology 296 (3–4), 389–413. doi:10.1016/j.palaeo.2010.08.001
- Murphy, M.A. & Matti, J.C., 1982: Lower Devonian Conodonts (hesperius-kindlei Zones), Central Nevada. University of California Publications Geological Sciences 123, 1–83
- Nilsson, E.K., 2005: Extinctions and faunal turnovers of early vertebrates during the Late Silurian Lau Event, Gotland, Sweden. Master’s thesis. University of Lund, Lund, Sweden. 32 pp.
- Oksanen, J., Blanchet, F.G., Friendly, M., Kindt, R., Legendre, P., Mcglinn, D., Minchin, P.R., O’hara, R.B., Simpson, G.L., Solymos, P., Stevens, M.H.M., Szoecs, E. & Wagner, H., 2019: vegan: Community Ecology Package. R package version 2.5-4. URL
- Pander, C.H., 1856: Monographie der fossilen Fische des silurischen Systems des Russisch-Baltischen Gouvernements, 91. Buchdruckerei der Kaiserlichen Akademie der Wissenschaften, St. Petersburg
- Philip, G.M., 1965: Lower Devonian conodonts from the Tyers area, Gippsland, Victoria. Proceedings of the Royal Society of Victoria 79 (1), 95–118
- Rhodes, F.H.T., 1953: Some British lower Palaeozoic conodont faunas. Philosophical Transactions of the Royal Society of London. Series B, Biological Sciences 237 (647), 261–334. doi:10.1098/rstb.1953.0005
- Rohon, J. V., 1893: Die obersilurischen fische von oesel. mémoires de l'académie imperiale des sciences de st. pétersbourg, Tome. 41(5),1–113.
- Sadler, P.M., 2012: Integrating carbon isotope excursions into automated stratigraphic correlation: an example from the Silurian of Baltica. Bulletin of Geosciences 87 (4), 681–694. doi:10.3140/bull.geosci.1307
- Samtleben, C., Munnecke, A. & Bickert, T., 2000: Development of facies and C/O-isotopes in transects through the Ludlow of Gotland: evidence for global and local influences on a shallow-marine environment. Facies 43 (1), 1–38. doi:10.1007/BF02536983
- Samtleben, C., Munnecke, A., Bickert, T. & Pätzold, J., 1996: The Silurian of Gotland (Sweden): facies interpretation based on stable isotopes in brachiopod shells. Geologische Rundschau 85 (2), 278–292. doi:10.1007/BF02422234
- Schiøler, P., 1989: Non-toxic low-cost heavy liquid separation in the Geological Survey of Greenland. Rapport Grønlands Geologiske Undersøgelse 145, 11–13
- Slavík, L. & Carls, P., 2012: Post-Lau Event (late Ludfordian, Silurian) recovery of conodont faunas of Bohemia. Bulletin of Geosciences 87 (4), 815–832. doi:10.3140/bull.geosci.1368
- Spjeldnaes, N., 1950: On some vertebrate fossils from Gotland with some comments on the stratigraphy. Arkiv För Mineralogi Och Geologi 8 (1), 211–219
- Strömberg, C., 1997: The conodont Ctenognathodus in the Silurian of Gotland, Sweden. Master’s thesis. Lund University, Lund, Sweden. 56 pp.
- Team, R. C., 2018: R: A language and environment for statistical computing. R Foundation for statistical computing, Vienna, Austria. URL https://www.R-project.org/
- Ter Braak, C.J., 1986: Canonical correspondence analysis: a new eigenvector technique for multivariate direct gradient analysis. Ecology 67 (5), 1167–1179. doi:10.2307/1938672
- Traquair, R.H., 1898: Report on fossil fishes. Summary of Progress of the Geological Survey of the United Kingdom for 1897, 72–76.
- Trinajstic, K., 2001: Acanthodian microremains from the Frasnian Gneudna formation, Western Australia. Records of the Western Australian Museum 20 (2), 187–198
- Turner, S., 1986: Thelodus macintoshi Stetson 1928: The largest known Thelodont (Agnatha: Thelodonti). Breviora 486, 1–18
- Turner, S., 1999: Early Silurian to Early Devonian thelodont assemblages and their possible ecological significance. In A.J. Boucot & J.D. Lawson (eds.): Paleocommunities: a case study from the Silurian and Lower Devonian, 42–78. Cambridge University Press, Cambridge
- Valiukevičius, J., 1992: First articulated Poracanthodes from the lower Devonian of Severnaya Zemlya. In E. Mark-Kurik (ed.): Fossil fishes as living animals, 193–214. Academy of Sciences of Estonia, Tallinn
- Valiukevičius, J., 1998: Acanthodians and zonal stratigraphy of lower and Middle Devonian in East Baltic and Byelorussia. Palaeontographica Abteilung A, 1–53.
- Valiukevičius, J., 2003a: New Silurian nostolepids (Acanthodii, Pisces) of Lithuania. Geologija 42, 51–68
- Valiukevičius, J., 2003b: New late Silurian to Middle Devonian acanthodians of the Timan-Pechora region. Acta Geologica Polonica 53 (3), 209–245
- Valiukevičius, J., 2004a: Silurian acanthodian succession of the Lužni-4 borehole (Latvia). Acta Universitatis Latviensis 679, 120–147
- Valiukevičius, J., 2004b: New Wenlock-Pridoli (Silurian) acanthodian fishes from Lithuania. Acta Palaeontologica Polonica 49 (1), 147–160
- Valiukevičius, J., 2005: Silurian acanthodian biostratigraphy of Lithuania. Geodiversitas 27 (3), 349–380
- Valiukevičius, J., 2006: Event pattern in the development of Siluro-Devonian acanthodians of Lithuania. Neues Jahrbuch fur Geologie und Palaontologie-Monatshefte (6), 321–343. doi:10.1127/njgpm/2006/2006/321
- Valiukevičius, J. & Burrow, C.J., 2005: Diversity of tissues in acanthodians with Nostolepis-type histological structure. Acta Palaeontologica Polonica 50 (3), 635–649
- Vergoossen, J., 1997: Revision of poracanthodid acanthodians. In A. Ivanov, M.V.H. Wilson & A. Zhuravalov (eds.): Palaeozoic strata and fossils of the Eurasian Arctic, 23–26. St. Petersburg
- Vergoossen, J., 1999a: Late Silurian fish microfossils from Helvetesgraven, Skåne (southern Sweden)(I). Geologie En Mijnbouw 78, 267–280. doi:10.1023/A:1003866312419
- Vergoossen, J., 1999b: Siluro-Devonian microfossils of Acanthodii and Chondrichthyes (Pisces) from the Welsh Borderland, South Wales. Modern Geology 24 (1), 23–90
- Vergoossen, J., 1999c: Late Silurian fish microfossils from an East Baltic-derived erratic from Oosterhaule, with a description of new acanthodian taxa. Geologie En Mijnbouw 78 (2), 231–251. doi:10.1023/A:1003803814177
- Vergoossen, J., 2000: Acanthodian and chondrichthyan microremains in the Siluro-Devonian of the Welsh Borderland, Great Britain, and their biostratigraphical potential. Courier Forschungsinstitut Senckenberg, 175–200.
- Vergoossen, J., 2002a: Late Silurian fish microfossils from Ramsåsa, locality H, Scania, south Sweden, with some remarks on the body zonation scheme used in thelodont studies. Scripta Geologica 123, 41–69
- Vergoossen, J., 2002b: Late Silurian fish microfossils from Klinta and Rinnebäcks Bro (Scania, south Sweden), with remarks on the morphology of Nostolepis striata trunk scales. Scripta Geologica 123, 71–92
- Vergoossen, J., 2002c: Late Silurian fish microfossils from Ramsåsa (sites D and ‘south of church’), Skåne, south Sweden. Scripta Geologica 123, 93–158
- Vergoossen, J., 2003a: First record of fish microfossils from Ramsåsa, site C, Skåne, southern Sweden. Scripta Geologica 126, 1–78
- Vergoossen, J., 2003b: Fish microfossils from the upper Silurian Öved Sandstone Formation, Skåne, Southern Sweden – A paleontological appraisal of biostratigraphical tools. Ph. D thesis. Rijksuniversiteit Groningen, Groningen, Netherlands. 328 pp.
- Vergoossen, J., 2004: Fish microfossils from Ramsåsa, site E, Scania, southern Sweden (mid Palaeozoic). Scripta Geologica 127, 1–70
- Viira, V., 1994: A new upper Silurian conodont species from Estonia. Proceedings of the Estonian Academy of Sciences 43 (1), 32–37
- Viira, V. & Aldridge, R.J., 1998: Upper Wenlock to Lower Přídolí (Silurian) conodont biostratigraphy of Saaremaa, Estonia, and a correlation with Britain. Journal of Micropalaeontology 17 (1), 33–50. doi:10.1144/jm.17.1.33
- Viira, V. & Einasto, R., 2003: Wenlock-Ludlow boundary beds and conodonts of Saaremaa Island, Estonia. Proceedings of the Estonian Academy of Sciences 52 (4), 213–238
- Walliser, O.H., 1964: Conodonten des Silurs. Abhandlungen des Hessischen Landesamtes für Bodenforschung zu Wiesbaden 41, 1–106.
- Woodward, A.S., 1891: Catalogue of the fossil fishes in the British Museum (Natural History), Part II: XLIV, 567. Trustees of the British Museum (Natural History), London
- Younes, H., Calner, M. & Lehnert, O., 2016: The first continuous δ13C record across the Late Silurian Lau Event on Gotland, Sweden. GFF 139 (1), 63–69. doi:10.1080/11035897.2016.1227362
- Zhu, M., Yu, X., Ahlberg, P.E., Choo, B., Lu, J., Qiao, T., Qu, Q., Zhao, W., Jia, L., Blom, H. & Zhu, Y., 2013: A Silurian placoderm with osteichthyan-like marginal jaw bones. Nature 502 (7470), 188–193. doi:10.1038/nature12617
- Zidek, J., 1985: Über das Wachstum von Acanthodes (Acanthodii: Pisces) Meßwerte und ihre Bedeutung. Paläontologische Zeitschrift 59 (1–2), 147–166. doi:10.1007/BF02986006
- Ziegler, W., 1956: Unterdevonische Conodonten, insbesondere aus dem Schönauer und dem Zorgensis-Kalk. Notizblatt des Hessischen Landesamtes für Bodenforschung 84, 93–106.
- Ziegler, W., 1960: Conodonten aus dem Rheinischen Unterdevon (Gedinnium) des Remscheider Sattels (Rheinisches Schiefergebirge). Paläontologische Zeitschrift 34 (2), 169–201. doi:10.1007/BF02987050