
ABSTRACT
A new rare Upper Ordovician mound dwelling palaeoloricate chiton is described as Crassaplax collicola gen. et sp. nov. from three large isolated intermediate valves from the Katian carbonate mud mounds of the Boda Limestone in the Siljan district of central Sweden. Together with the previously known Spicuchelodes? sp., also from the Boda Limestone, these are the only known Late Ordovician mound dwellers so far among the palaeoloricates, adding to Ordovician environments with reported chiton remains. Crassaplax collicola is distinguished from other Ordovician palaeoloricate chitons by their thick and large valves that can be 2 cm in length, and especially the clear differentiation between the lateral and central valve areas.
Introduction
A stoutly built, new basal chiton (Mollusca) is described as Crassaplax collicola gen. et sp. nov. from isolated valves found in the Upper Ordovician (Katian) carbonate mud mounds of the Boda Limestone in the Siljan district of central Sweden (). It is a rare addition to the unusually diverse fauna of the Boda Limestone, and represents one of only two chiton taxa known from the Ordovician of Baltoscandia (Högström et al. Citation2009). Molluscs (e.g., cephalopods, bivalves, gastropods and rostroconchs) are otherwise the third most diverse and abundant group of the Boda Limestone (see Ebbestad & Högström Citation2007a for references).
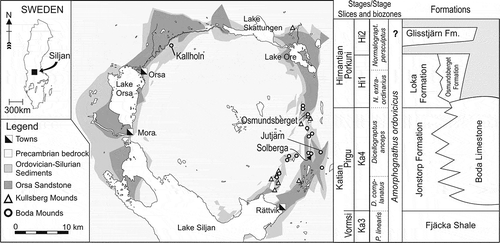
Figure 1. Map of the Siljan ring structure with the location of the older Kullsberg and the younger Boda carbonate mounds and Jutjärn quarry (WGS 84 60°58ʹ57.7”N 15°15ʹ3.3”E). The stratigraphical column to the right shows the global standard for stages, graptolite, and conodont biozones, in addition to the regional stratigraphy of the Siljan district, Stage Slices based on Bergström et al. (Citation2009). Stratigraphy based on Ebbestad et al. (Citation2015) and Kröger et al. (Citation2016)
Fossil polyplacophorans are generally placed in the Palaeoloricata of Bergenhayn (Citation1955), characterised by the absence of the articulamentum layer with exposed apophyses typical of neoloricates. Vendrasco & Runnegar (Citation2004) and Pojeta et al. (Citation2010) regarded the Palaeoloricata as a convenient morphological grouping that most likely is paraphyletic, containing stem members of the clade Aculifera that encompass both Polyplacophora and Aplacophora (Sigwart & Sutton Citation2007; Smith et al. Citation2011; Vinther et al. Citation2011, Citation2012; Sutton & Sigwart Citation2012 and references in these). The aculiferan divergence may have happened in the Ordovician where palaeoloricates with a narrow morphology potentially represent stem-aplacophorans, in that they do not possess a foot, e.g. the typical creeping sole of modern chitons. Rather these stem forms have a spiculate girdle surrounding the dorsal valves and a vermiform body plan (Vinther et al. Citation2011; Sutton & Sigwart Citation2012). Two such forms from the Late Ordovician (Katian) of Scotland, Septemchiton grayiae (in reality S. vermiformis) and Phthipodochiton thraivensis (see ), were discussed as possible stem-aplacophorans (Vinther et al. Citation2011; Sutton & Sigwart Citation2012), but assessing whether other Ordovician palaeoloricate taxa are stem-aplacophorans or stem-polyplacophorans is difficult without special preservation or articulated specimens (see discussion in Sutton & Sigwart Citation2012). Thus, the position of Crassaplax remains unclear, although the large size (uncertain character) and thick valves (compared to the thin valves of the before mentioned forms) may suggest a polyplacophoran affinity. Support for the clade Aculifera today comes from both molecular and fossil data, for example Giribet (Citation2014), Vinther et al. (Citation2017) and Kocot et al. (Citation2020).
Table 1. Upper Ordovician – lower Silurian (Sandbian–Telychian) palaeoloricate taxa. Ordovician Stage Slices follow those defined by Bergström et al. (Citation2009)
Lower Palaeozoic palaeoloricates are rare fossils, showing diversity peaks in the Lower Ordovician and middle Silurian (Hoare & Pojeta Citation2006; Puchalski et al. Citation2008). The scant record may not signify a biological signal but rather seems to be a factor of a geographically and stratigraphically thinly spread and sporadic sampling effort (Puchalski et al. Citation2008). This was amply demonstrated by the study of Hoare & Pojeta (Citation2006) from U.S.A. which alone increased the number of chitons known from the Ordovician by 42% and specifically for the Middle Ordovician by over 150%. In the case of the Boda Limestone, the sampling effort has been high since commercial quarrying started in the 1930’s. In particular, Elsa Warburg and Orvar Isberg contributed during the 1920’s and 1930’s to the extant collections, but only one valve was found among this early material. It is significant that the new finds all are from Jutjärn quarry, which today is the only active quarry in the Boda Limestone. It has provided access to the deeper core material and fossil pockets in these, which in other quarries are long since gone. This may suggest that chiton material may be more common but is not recovered due to limited outcrop that exist for a short time only. However, the Boda Limestone mounds represent a high energy environment, where remains of organisms likely are fragmented, destroyed, and often eroded, thus becoming increasingly difficult to recognise for the non-specialist. Cherns & Wright (Citation2000), also pointed out that chiton valves have lower preservation potential due to disarticulation and aragonitic dissolution. Additionally, a recent study by Peebles & Smith (Citation2019) suggested that dissolution before burial can be enough to remove chiton remains from the fossil record over time. In some instances, chitons are very common in localized facies, such as Solenocaris solenoides Young & Young, Citation1868 in the Ordovician of Girvan, Scotland (Stewart & Owen Citation2008).
Due to the sparse records and a few geographically specific monographic studies, the geographical distribution of Ordovician chitons is biased towards northern and marginal Laurentia (e.g. U.S.A., Ireland, Scotland) and for the Silurian towards Baltica (e.g. Gotland) (see for example Bergenhayn Citation1955; Candela et al. Citation2014; Cherns Citation1998a, Citation1998b, Citation2004; Hoare & Pojeta Citation2006; Sigwart Citation2007; Sutton & Sigwart Citation2012; Vendrasco & Runnegar Citation2004). While Palaeozoic chitons are known from a range of depositional settings, Crassaplax and Spicuchelodes? sp. from the Boda Limestone are the only known carbonate mound dwellers ().
The inferred large size of Crassaplax individuals reflects the distribution of other large benthos in the Boda Limestone. Kröger (Citation2013) and Kröger & Ebbestad (Citation2014) showed that the diverse cephalopod fauna with 61 species known represent the top predators in the trophic structure of the Boda mounds. Large species of trilobites, bivalves and some brachiopods are among the dominating elements of the benthos, to which the large Crassaplax can be added. These were probably important prey for the abundant cephalopods.
Geological setting and material
The Boda Limestone of the Siljan district, central Sweden, comprises large carbonate mud mounds showing no organic frame work. The structures may reach a diameter of up to 1000 m with a thickness of 100–140 m, and are well exposed in a number of quarries (Ebbestad & Högström Citation2007b). The core consists of dense micrites replaced laterally by pelmatozoan flank facies. These two are Katian in age, while a terminal bioclastic facies, the Osmundsberget Formation, is entirely Hirnantian (Suzuki et al. Citation2009; Rasmussen et al. Citation2010; Kröger et al. Citation2016). The depositional environment seems to represent cool, nutrient rich water in the deeper neritic zone with a depth of 100–140 m (Kröger & Ebbestad Citation2014).
Synsedimentary and early postdepositional vertical fissures of variable size and development are characteristic for the core facies (referred to as pockets by Suzuki & Bergström Citation1999). The fissures are arranged radially or circumferential along the mounds and display complex morphologies and depositional history (Kröger et al. Citation2015, Citation2016). Fissures are often filled with a coquina of one of nine common trilobites interspersed with other fossil groups as a minor component. Typically, the faunal elements trapped in the fissures would live on the top surface of the mounds, being mostly transported into their resting place (Suzuki & Bergström Citation1999). Macrofossils are uncommon in the micritic facies. A plethora of trilobites, brachiopods, echinoderms, molluscs and corals are described from the Boda Limestone (see references in Ebbestad & Högström Citation2007a; Kröger Citation2013), but many of the molluscan groups are still in great need of descriptive work and revision.
Three intermediate valves of the herein described Crassaplax are known, all collected from the Jutjärn quarry in Dalarna. They are stored at the Museum of Evolution, Uppsala University, Uppsala, Sweden (PMU). Specimen PMU 25141 was found in a fissure filling dominated by Stenopareia oviformis (Warburg, Citation1925) excavated from the deepest part of the quarry in 2005. Other associated taxa are the trilobites Eobronteus laticauda (Wahlenberg, Citation1818) and Sphaerexochus calvus M’Coy, Citation1846, as well as the gastropod Cataschisma microsoma (Koken in Koken & Perner, Citation1925). The other two valves, PMU 26954 and PMU 26955, were collected 2009 and the former is also associated with Stenopareia oviformis. It is therefore likely that the fossils are from contemporaneous deposits. Only the intermediate valves are known.
Terminology of polyplacophorans and parameters for standard measurements follow that of Cherns (Citation1998a). The term valve has been used for polyplacophorans to a larger extent than sclerite as used by Högström et al. (Citation2009); here we use the term valve.
Systematic palaeontology
Phylum Mollusca Linnaeus, Citation1758
Palaeoloricata Bergenhayn, Citation1955
Remarks
Following the discussion in especially Sutton and Sigwart (Citation2012) we see it fit to regard Palaeoloricata as an unresolved clade containing genera that may belong to the stem group Aculifera, the Polyplacophora and/or the Aplacophora.
Genus
Crassaplax gen. nov
Type species
Crassaplax collicola sp. nov from the Katian Boda Limestone of the Siljan district, Dalarna, Sweden.
Diagnosis
As for type species.
Etymology
From Latin Crassus for thick or stout and plax from the Greek word for plate. The gender is feminine.
Remarks
Specimen PMU 25141 was described as Chelodes sp. A by Högström et al. (Citation2009). However, Chelodes as understood today is morphologically widely defined, as well as a long ranging genus (for example, Lower Ordovician – upper Silurian/Lower Devonian stated by Cherns Citation1998a, and late Cambrian – Early Devonian stated by; Pojeta et al. Citation2003), with a global distribution. It does seem as the current understanding of Chelodes requires a thorough examination and redefinition but this is outside our scope here. Crassaplax distinguishes itself from Chelodes primarily in the highly arched, roughly squarish valves compared to the often substantially elongated Chelodes ones, and the sharp distinction between central and lateral valve areas. Comparing median length/length ratios () for Crassaplax with those of different Chelodes taxa described in Cherns (Citation1998a) give similar results, although the ratio may be somewhat lower in Crassaplax compared to that of Chelodes. Although not always present in Chelodes, granulated ornamentation as described by Cherns (Citation1998a) is seemingly missing in the Boda Limestone taxon. Additionally, Crassaplax exhibit central and lateral valve areas that are very distinctly separated in a manner not seen in most Chelodes taxa, Cherns (Citation1998a) indicated that in this genus the valve areas are lacking or weakly expressed, although a few cases are more well-defined. Preacanthochiton Bergenhayn, Citation1960 share similarities with Crassaplax in the overall shape of valves but differ primarily in the weaker differentiation of medial and lateral valve areas, as Chelodes does.
Table 2. Measurements for the three described plates of Crassaplax collicola gen. et sp. nov., terminology and measurements after Cherns (Citation1998a). The apical angle in specimen PMU 26954 is an estimate, as the posterior-lateral margins are not well preserved
Crassaplax collicola sp. nov. ,
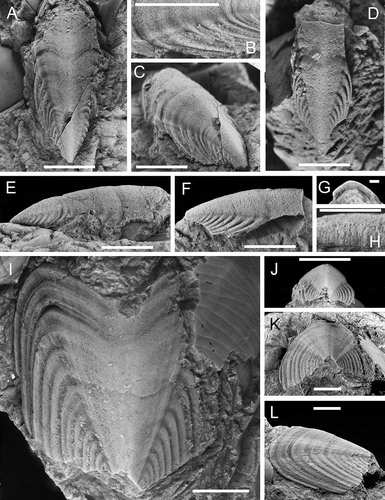
Figure 2. A–L. Crassaplax collicola n. gen, n. sp. A–C, E. PMU 26954, dorsal, detail of lateral valve area, dorsal oblique, and lateral views respectively. The lateral margins are largely missing D, F–H, J. PMU 26955, dorsal, lateral, anterior and cross section of anterior margin respectively. The lateral margins are largely missing I, K–L. PMU 25141, Holotype and most complete specimen. All specimens from Jutjärn quarry. PMU 25141, coll., J.O.R. Ebbestad, 2005; PMU 26954, 26955, coll. J. Suzuki, 2009. Scale bar in A–F, I–L = 0.5 cm; scale bar in G, H = 0.1 cm
2009 Chelodes sp. A. Högström, Ebbestad & Suzuki, fig. 5A–F.
Material
Three isolated intermediate valves from Jutjärn quarry (), Upper Ordovician (Katian) Boda Limestone, the Siljan district, Dalarna, Sweden. Measurements are given in .
Holotype
PMU 25141, .
Etymology
The species epithet collicola means Mound Dweller. From Latin Collis meaning hill or mound, and New Latin Colus meaning dweller.
Diagnosis
Intermediate valves thick and can reach a large size. Shell with distinct embayment, and comarginal rugae on lateral valve area, Ridges absent on highly arched central valve area. Jugal angle around 100°. Slightly beaked but sharp apex with a general squarish appearance of valves in dorsal view.
Description
Large (2 cm in length), robust valves, ventral surface not exposed. Central valve area large and essentially without ornamentation except for faint traces of the comarginal rugae near the anterior margin, rugae running nearly straight (antero-posteriorly) on the lateral valve areas. Conspicuous anterior embayment clearly visible in specimens PMU 26954 and PMU 25141, highly arched valve, and sharply marked line between central and lateral valve areas. PMU 25141 roughly square in dorsal view. PMU 26954 () has broken lateral margins, partially preserved anterior margin with distinct embayment, apical region nearly intact showing pointed slightly beaked apex, the valve is cracked. PMU 26955 () has no preserved complete margin but apical region nearly complete and very similar to PMU 26954, these two also of equal size. PMU 25141 () exposes a nearly complete anterior margin with prominent embayment and large lateral valve areas whereas the apical region is not preserved. See also for a schematic illustration of valve arrangement, including inferred pointed apices.
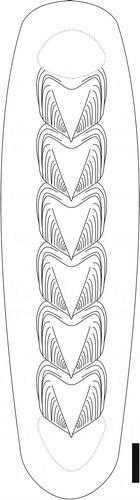
Figure 3. Schematic illustration of Crassaplax collicola, head and tail valves are unknown, and in this illustration represented by stippled markings. The outline of the body is conjectural, and the presence/absence of a foot is unknown. Scale bar = 1 cm
Remarks
There is a clear difference between the two listed apical angle values for specimens PMU 25141 and 26954, likely depending on differential preservation of the posterolateral portions of the lateral margins. The two smaller specimens therefore do not preserve the squarish outline seen in the more complete specimen. The two smaller valves have no complete margins and exact measurements of the apical angle is therefore difficult, but there is a good match in general appearance between the three specimens. The median length/length ratio (see ), is a rough estimate of the extent of the embayment in this taxon and the correspondence of the three values is a clear indication of their similar morphology.
Crassaplax collicola is a large taxon where a complete specimen may have reached a length of 15–20 cm (). The intermediate valves of Crassaplax are comparable in size to those of Preacanthochiton baueri Hoare and Pojeta (Citation2006), an Ordovician taxon from U.S.A. with valves of broadly similar shape, faint ridges on the lateral valve areas and an embayment of similar extent compared to the Boda Limestone taxon, but consistently described as possessing thinner valves. Some of the larger individuals of Chelodes from the Silurian of Gotland may have reached a total length of around 10 cm (Cherns Citation1998a), but far from all Chelodes were that large.
Acknowledgments
We are grateful to Peter Andersson (SMA Mineral, Rättvik) for access to various quarries owned or operated by SMA Mineral. Michael Vendrasco (Pasadena) provided insightful reviews that greatly improved the manuscript, as well as Lesley Cherns (Cardiff) on an early version of the paper.
Disclosure statement
No potential conflict of interest was reported by the authors.
References
- Bergenhayn, J.R.M., 1955: Die fossilen Schwedischen Loricaten nebst einer vorläufigen Revision des Systems der ganzen Klasse Loricata. Lunds Universitets Årsskrift, Nya Förhandlingar, Afdelningen 2 51, 1–46.
- Bergenhayn, J.R.M., 1960: Cambrian and Ordovician loricates from North America. Journal of Paleontology 34, 168–178.
- Bergström, S.M., Chen, X., Gutíerrez Marco, J.C. & Dronov, A., 2009: The new chronostratigraphic classification of the Ordovician System and its relations to major regional series and stages and δ13 Chemostratigraphy. Lethaia 42, 97–107.
- Candela, Y., Cherns, L. & Troalen, L., 2014: First record of a polyplacophoran from the Southern Uplands of Scotland. Scottish Journal of Geology 50, 87–91.
- Cherns, L., 1998a: Chelodes and closely related Polyplacophora (Mollusca) from the Silurian of Gotland, Sweden. Palaeontology 41, 545–573.
- Cherns, L., 1998b: Silurian polyplacophoran molluscs from Gotland, Sweden. Palaeontology 41, 939–974.
- Cherns, L., 2004: Early Palaeozoic diversification of chitons (Polyplacophora, Mollusca) based on new data from the Silurian of Gotland, Sweden. Lethaia 37, 445–456.
- Cherns, L. & Wright, V.P., 2000: Missing molluscs as evidence for large scale, early skeletal aragonitic dissolution in a Silurian sea. Geology 28, 791–794.
- Dzik, J., 1994: Machaeridians, chitons, and conchiferan molluscs of the Mójcza Limestone. In J. Dzik, E. Olempska & A. Pisera (eds.): Ordovician Carbonate Platform Ecosystem of the Holy Cross Mountains. Palaeontologia Polonica 53, 213–252.
- Ebbestad, J.O.R. & Högström, A.E.S., 2007a: Ordovician of the Siljan District, Sweden. In J.O.R. Ebbestad, L.M. Wickström & A.E.S. Högström (eds.): WOGOGOB 2007. Field guide and Abstracts. Sveriges geologiska undersökning, Rapporter och meddelanden 128, 7–26.
- Ebbestad, J.O.R. & Högström, A.E.S., 2007b: Locality descriptions, the Siljan District. In J.O.R. Ebbestad, L.M. Wickström & A.E.S. Högström (eds.): WOGOGOB 2007. Field guide and Abstracts. Sveriges geologiska undersökning, Rapporter och meddelanden 128, 52–58.
- Ebbestad, J.O.R., Högström, A.E.S., Frisk, Å.M., Martma, T., Kaljo, D., Kröger, B. & Pärnaste, H., 2015: Terminal Ordovician stratigraphy of the Siljan district, Sweden. GFF 137, 36–56.
- Giribet, G., 2014: On Aculifera: a review of hypotheses in tribute to Christoffer Schander. Journal of Natural History 48, 2739–2749.
- Hoare, R.D. & Pojeta, J., Jr., 2006: Ordovician Polyplacophora (Mollusca) from North America. Paleontological Society Memoir 64, 1–27.
- Högström, A.E.S., Ebbestad, J.O.R. & Suzuki, Y., 2009: Armoured annelids and molluscs from the Upper Ordovician Boda Mud Mounds, central Sweden. GFF 131, 245–252.
- Kocot, K.M., Poustka, A.J., Stöger, I., Halanych, K.M. & Schrödl, M., 2020: New data from Monoplacophora and a carefully-curated dataset resolve molluscan relationships. Scientific Reports, 10. doi:10.1038/s41598-019-56728-w.
- Koken, E. & Perner, J., 1925: Die Gastropoden des Baltischen Untersilurs. Mémoires de L’ ácadémie des sciences de Russie Serie 8, Classe Physico–Mathématique 37, 1–326.
- Kröger, B., 2013: The cephalopods of the Boda Limestone, Late Ordovician, of Dalarna, Sweden. European Journal of Taxonomy 41, 1–110.
- Kröger, B. & Ebbestad, J.O.R., 2014: Palaeoecology and palaeogeography of Late Ordovician (Katian–Hirnantian) cephalopods of the Boda Limestone, Siljan district, Sweden. Lethaia 47, 15–30.
- Kröger, B., Ebbestad, J.O.R. & Lehnert, O., 2016: Accretionary Mechanisms and Temporal Sequence of Formation of the Boda Limestone Mud-Mounds (Upper Ordovician), Siljan District, Sweden. Journal of Sedimentary Research 86, 363–379.
- Kröger, B., Ebbestad, J.O.R., Lehnert, O., Ullmann, C.V., Corte, C., Frei, R. & Rasmussen, C.M.Ø., 2015: Subaerial speleothems and deep karst in central Sweden linked to Hirnantian glaciations. Journal of the Geological Society 172, 349–356.
- Linnaeus, C.V., 1758: Systema naturae per regna tria naturae, secundum classes, ordines, genera, species, cum characteribus, differentiis, synonymis, locis. Tomus I. Editio decima, reformata. Ii + 824 p. Laurentius Salvius, Holmiae.
- M’Coy, F., 1846: Synopsis of the Silurian fossils of Ireland, 72 p. Dublin University Press, Dublin.
- Peebles, B.A. & Smith, A.M., 2019: Wasting away in the intertidal: the fate of chiton valves in an acidifying ocean. Palaios 34, 281–290.
- Pojeta, J., Jr., Eernisse, D.J., Hoare, R.D. & Henderson, M.D., 2003: Echinochiton dufoei: a new spiny Ordovician chiton. Journal of Paleontology 77, 646–654.
- Pojeta, J., Jr., Vendrasco, M.J. & Darrough, G., 2010: Upper Cambrian chitons (Mollusca, Polyplacophora) from Missouri, USA. Bulletins of American Paleontology 379, 82.
- Puchalski, S.S., Eernisse, D.J. & Johnson, C.C., 2008: The effect of sampling bias on the fossil record of chitons (Mollusca, Polyplacophora). American Malacologicl Bulletin 25, 87–95.
- Rasmussen, C.M.Ø., Ebbestad, J.O.R.& Harper, D.A.T., 2010: Unravelling a Late Ordovician pentameride (Brachiopoda) hotspot from the Boda Limestone, Siljan District, central Sweden. GFF 132, 133–152.
- Reed, F.R.C., 1911: A new fossil from Girvan. (1.– Sedgwick Museum notes). Geological Magazine, Decade 5 8, 337–339.
- Rhoads, D.C., 1962: Microfossils of problematic affinity from the Maquoketa Formation of eastern Iowa and western Illinois. Journal of Paleontology 36, 1334–1340.
- Sigwart, J.D., 2007: The Irish fossil Polyplacophora. Irish Journal of Earth Sciences 25, 27–38.
- Sigwart, J.D. & Sutton, M.D., 2007: Deep molluscan phylogeny: synthesis of palaeontological and neontological data. Proceedings of the Royal Society of London B 274, 3413–3419.
- Smith, S.A., Wilson, N.G., Goetz, F.E., Feehery, C., Andrade, S.C., Rouse, G.W., Giribet, G. & Dunn, C.W., 2011: Resolving the evolutionary relationships of molluscs with phylogenomic tools. Nature 480, 364–367.
- Stewart, S. & Owen, A.W., 2008: Probing the deep shelf: a Lagerstätte from the Upper Ordovician of Girvan, Southwestern Scotland. Lethaia 41, 139–146.
- Sutton, M.D. & Sigwart, J.D., 2012: A chiton without a foot. Palaeontology 55, 401–411.
- Suzuki, Y. & Bergström, J., 1999: Trilobite taphonomy and ecology in Upper Ordovician carbonate buildups in Dalarna, Sweden. Lethaia 32, 159–172.
- Suzuki, Y., Shiino, Y. & Bergström, J., 2009: Stratigraphy, carbonate facies and trilobite associations in the Hirnantian part of the Boda Limestone, Sweden. GFF 131, 299–310.
- Vendrasco, M.J. & Runnegar, B., 2004: Late Cambrian and Early Ordovician stem group chitons (Mollusca: polyplacophora) from Utah and Missouri. Journal of Paleontology 78, 675–689.
- Vinther, J., Jell, P., Kampouris, G., Carney, R., Racicot, R.A. & Briggs, D.E.G., 2012: The origin of multiplacophorans – convergent evolution in aculiferan molluscs. Paleontology 55, 1007–1019.
- Vinther, J., Parry, L., Briggs, D.E.G. & Van Roy, P., 2017: Ancestral morphology of crown-group molluscs by a new Ordovician stem aculiferan. Nature 542, 471–474.
- Vinther, J., Sperling, E.A., Briggs, D.E. & Peterson, K.J., 2011: A molecular palaeobiological hypothesis for the origin of aplacophoran molluscs and their derivation from chiton-like ancestors. Proceedings of the Royal Society B 279, 1259–1268.
- Wahlenberg, G., 1818: Petrificata telluris svecanae. Nova Acta Regiae Societatis Scientiarum Upsaliensis 8, 1–116.
- Warburg, E., 1925: The trilobites of the Leptaena Limestone in Dalarne. Bulletin of the Geological Institution of the University of Uppsala 17, 1–446.
- Woodward, H., 1885: On a new species of Helminthochiton from the Upper Bala (Silurian) of Girvan, Ayrshire. Geological Magazine, New Series, Decade 3 2, 352–358.
- Young, & Young, J., 1868: Solenocaris solenoides. In: on new forms of Crustacea, from Silurian Rocks at Girvan, by J. Young. Proceeding of the Natural History Society of Glasgow 1, 173.