
ABSTRACT
New fossil discoveries are reported from the Grammajukku Formation at Luobákte south of Lake Torneträsk in northern Swedish Lapland, including a fauna of Small Shelly Fossils (SSF) from a limestone bed in the uppermost part of the formation and new occurrences of brachiopods and trilobites in siltstones of the lower part of the formation. The moderately diverse SSF fauna is the first of its kind reported from the Swedish Caledonides and includes the first record of the tommotiid Lapworthella schodackensis and the bradoriid spine Mongolitubulus spinosus from Baltica, together with fragmentary specimens of Bradoria sp. and remains of one additional bradoriid arthropod, a protoconodont and a helcionelloid mollusc. In addition, the limestone bed yields abundant specimens of the brachiopods Botsfordia cf. caelata and Eoobolus cf. priscus and an unidentified ellipsocephalid trilobite. Lower down in the Grammajukku Formation, specimens of both brachiopod taxa, orthothecid hyoliths, the trilobite Ellipsocephalus cf. gripi and an unidentified holmiid trilobite were found at several levels in a siltstone, previously regarded as unfossiliferous. These discoveries markedly increase the known diversity of the palaeobiota from the Grammajukku Formation in northern Lapland and provide new insights into the biostratigraphy and palaeoenvironment of the lower Cambrian in Scandinavia and the palaeobiogeography of Cambrian faunas in general.
Introduction
After the break-up of Rodinia in the late Proterozoic, the palaeocontinent Baltica was situated in the Southern Hemisphere and was moving northwards. During the first half of the Cambrian, Baltica was situated at middle to high southern latitudes (c. 35–60°) with modern Scandinavia along its northern margin (Meert Citation2013). The climate during the Cambrian is poorly resolved but is generally interpreted to have been relatively warm with tropical to sub-tropical temperatures even at high latitudes (Hearing et al. Citation2018; Wotte et al. Citation2019). The shallow Cambrian seas hosted the first major radiation of metazoan life – the so called “Cambrian Explosion” when all the major extant phyla of animals evolved during a 20–30 million year period from the terminal Ediacaran through the first four stages of the Cambrian. Rich fossil faunas of early Cambrian (Terreneuvian and Series 2) are known from most Cambrian palaeocontinents, but the fossil record of Baltica remains comparatively poorly understood and described faunas are largely restricted to trilobites. Consequently, our understanding of the Cambrian marine ecosystems of Baltica remains incomplete until other components of the fauna are documented.
The so-called Small Shelly Fossils (SSFs) include a great diversity of typically very small, skeletal fossils representing a wide range of invertebrates that are commonly extracted from carbonates using weak acids (Bengtson Citation2004). Although such fossil assemblages occur throughout the Phanerozoic (Dzik Citation1994; Freeman et al. Citation2019), the term SSF, as commonly defined, is restricted to assemblages from the Cambrian where they include examples of the oldest known representatives of many fossil and extant animal clades (Matthews & Missarzhevsky Citation1975; Bengtson Citation2004). This informal group incorporates early members of well-known animal clades, such as Mollusca, Brachiopoda and Porifera (Bengtson et al. Citation1990; Skovsted Citation2006), and also many problematic fossils that, historically, have been difficult to place in modern animal groups, although they now appear to belong the stem groups of modern lineages (Szaniawski Citation2002; Vinther & Nielsen Citation2005; Skovsted et al. Citation2008, Citation2014; Demidenko & Parkhaev Citation2020). The oldest SSF assemblages stem from the terminal Ediacaran (Grant Citation1990), but the Cambrian witnessed a remarkable, successive increase in taxonomic diversity and morphological disparity of SSFs from the Terreneuvian through to the, as yet undefined, Cambrian Stage 4 (Kouchinsky et al. Citation2012). Thus, the fossil record of SSFs in the Ediacaran–Cambrian reflects the dramatic changes in the Earth’s biota around the beginning of the Phanerozoic – the famous “Cambrian Explosion”.
Historically, SSF assemblages have been studied extensively from early Cambrian carbonate successions in Siberia (Rozanov et al. Citation1969), South China (Qian & Bengtson Citation1989), Australia (Bengtson et al. Citation1990), Mongolia (Esakova & Zhegallo Citation1996) and Laurentia (Skovsted Citation2006). From the palaeocontinent of Baltica, early works on brachiopods and some other fossils that fit under the SSF umbrella were published during the late nineteenth and early twentieth centuries (Moberg Citation1892; Wiman Citation1903; Kautsky Citation1945). However, in recent times, only a few studies targeting individual fossil taxa have been published (Poulsen Citation1967; Bengtson Citation1968, Citation1970; Dies Álvarez et al. Citation2008; Streng et al. Citation2008a). Instead, almost all attention has been devoted to coeval trilobite faunas (Bergström & Ahlberg Citation1981; Ahlberg et al. Citation1986; Ebbestad et al. Citation2003; Axheimer et al. Citation2007; Cederström et al. Citation2011; Høyberget et al. Citation2019 and references therein). This is particularly true for the autochthonous Cambrian sequences that crop out along the eastern margins of the Caledonian mountains in northern Scandinavia.
The stratigraphic section at Luobákte (alternative spelling: Luopakte; northern slope of Mt. Luovarri), south of Lake Torneträsk (), represents one of the most completely exposed sections of lower Cambrian autochthonous strata in the Caledonian mountains (). The section was described by Moberg (Citation1908) and Kulling (Citation1960) and proposed by Thelander (Citation1982) as the type section for the Torneträsk Formation (Dividalen Group) although the uppermost unit of the formation was later assigned to the Grammajukku Formation by Nielsen & Schovsbo (Citation2011). The presence of Cambrian trace fossils in the lower parts of the Torneträsk Formation has been known since the work of Moberg (Citation1908), and trilobites have been described from the uppermost part of the overlying Grammajukku Formation (Ahlberg Citation1980a, Citation1980b; Axheimer et al. Citation2007). Here we report a new fauna of Small Shelly Fossils (SSF) and brachiopods from the uppermost bioclastic limestone bed of the Grammajukku Formation and new outcrops containing brachiopods and trilobites from the lower part of the formation, in an interval, which hitherto has been considered unfossiliferous (Kulling Citation1960; Axheimer et al. Citation2007). The biostratigraphic and palaeobiogeographic implications of the fossil fauna are discussed.
Figure 1. Geological map. A. Map of Sweden with the study area indicated. B. Simplified geological map of the Torneträsk area with the fossil locality of Luobákte indicated. Brown colour represents outcrop areas of the autochthonous Cambrian strata along the margin of the Caledonian thrust sheets. C. Field photo of the Luobákte locality, taken from a low ridge north of Mt. Luovarri. The extent of the main stratigraphic units and approximate boundaries are indicated
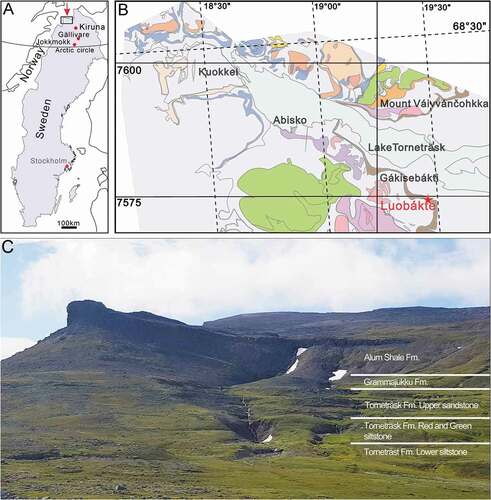
Geological setting
As in other parts of Scandinavia, Cambrian rocks were deposited in northern Sweden on the denuded Proterozoic craton of Baltica after an initial transgression some time in the early Cambrian (Nielsen & Schovsbo Citation2011). In northern Sweden, the Cambrian succession consists of the Dividalen Group, mainly siliciclastic sandstones and siltstones deposited during the first half of the Cambrian, followed by deposition of black alum shale during the later half of the period (Thelander Citation1982). This was presumably followed by deposition of limestone during the Ordovician before the onset of the Caledonian Orogeny, which led to the displacement onto the craton of a series of nappes or thrust sheets derived from the west (Roberts & Gee Citation1985). Owing to its structural weakness, the mid- to upper Cambrian alum shale generally served as a glide plane for the lowermost thrust sheet, obliterating any younger rocks but preserving the lower part of the Cambrian succession relatively intact (Roberts & Gee Citation1985). Exposures of the Cambrian sequence are generally poor, being largely constrained to the easternmost margin of the preserved thrust sheets where these autochthonous (or para-autochthonous) rocks crop out sporadically. However, parts of the succession may also be included in the lowermost thrust sheet itself (Ahlberg Citation1980a).
In the Torneträsk area (), the Cambrian Dividalen Group consists of a series of sandstones and siltstones with intercalated conglomerate, breccia and minor limestone, and the overlying alum shale (Moberg Citation1908; Kulling Citation1960; Thelander Citation1982). Thelander (Citation1982) designated the lower Cambrian succession as the Torneträsk Formation and divided it into five informal members. These are in ascending order: the “lower sandstone member”, the “lower siltstone member”, the “red and green siltstone member”, the “upper sandstone member” and the “upper siltstone member”. North of Lake Torneträsk, a sixth member, the Vakkejokk Breccia Member, was recognized between the “lower siltstone member” and the “red and green siltstone member” (Thelander Citation1982). The Vakkejokk Breccia Member has since been recognized as an impactite (Nielsen & Schovsbo Citation2011; Ormö et al. Citation2017). The uppermost unit of the Torneträsk Formation, the “upper siltstone member”, was excluded from the Torneträsk Formation by Nielsen & Schovsbo (Citation2011) based on a general sequence-stratigraphic analysis and instead considered to be a northern extension of the Grammajukku Formation, which otherwise crops out in southern and central Lapland (Willdén Citation1980).
The Torneträsk Formation at Luobákte is about 97 m thick and was described in detail by Moberg (Citation1908), Kulling (Citation1960), Thelander (Citation1982) and is also discussed by McLoughlin et al. (Citation2021 [This volume]). The overlying Grammajukku Formation at Luobákte is about 18 m thick (; Thelander Citation1982). The contact with the underlying “upper sandstone member” of the Torneträsk Formation is unconformable above a thin conglomeratic quartz sandstone bed and may represent a significant time gap (Nielsen & Schovsbo Citation2011). According to Kulling (Citation1960) and Vogt (Citation1967), the basal 0.5–1 m of the Grammajukku Formation consists of a conglomeratic dolomitic limestone containing phosphorite pebbles. However, only loose blocks of this unit were detected by Thelander (Citation1982), and no comparable rock types were found at this level during our investigation. The lower Grammajukku Formation consists of a homogeneous greenish-grey siltstone, about 15.5 m thick (Thelander Citation1982). An oblique cleavage pattern is developed in the siltstone, and tectonic disturbances are evident as slickensides along oblique fault planes, although no noticeable displacement could be detected in the field. The top 1.7 m of the formation consists of a dark brown to black thick-bedded calcareous siltstone, capped by a 15-cm-thick limestone layer with abundant fossil fragments and pyrite aggregates (Thelander Citation1982) that is equivalent to the “bioclastic limestone” of Axheimer et al. (Citation2007). This bed is succeeded by the black Alum Shale Formation, which is strongly deformed by the thrusting of the overlying nappes and is devoid of discernible bedding or other sedimentary structures.
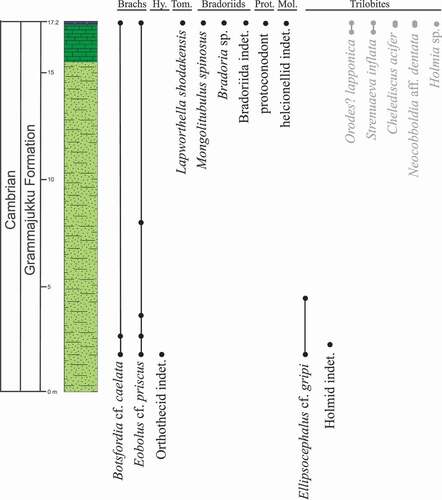
Figure 2. Lithostratigraphic section through the Grammajukku Formation, Torneträsk Formation, Luobákte, Lapland, including ranges of fossil taxa. Biostratigraphic ranges of taxa in light grey have been taken from Axheimer et al. (Citation2007), all other taxa (in regular black text) are documented from this study. Abbreviations: Brachs-Brachiopods, Hy.-Hyoliths, Prot.-Protoconodonts and Mol.-Molluscs
Material and methods
The autochthonous Cambrian stratigraphic section at Luobákte was investigated along the sides of a small creek on the northern slope of Mt. Louvarri during fieldwork in August 2020 (). A sample of “bioclastic limestone” (c. 3 kg), collected from the top bed of the Grammajukku Formation was treated with 5% acetic acid using standard protocols (Jeppsson et al. Citation1999). The resulting acid-resistant residue was wet sieved using 2 mm, 500 µm and 63 µm sieves and scanned manually for fossils using a stereomicroscope. Selected specimens were gold-coated and documented using a Quanta 650 FEG SEM at an operating voltage of 15Kv. Samples of calcareous siltstone collected at 0.5, 1.25 and 1.45 m below the top of the formation were processed in the same manner but did not dissolve and yielded no fossils.
Macroscopic body fossils from the lower part of the Grammajukku Formation were collected from a measured section () spanning the lowermost 8 m of the siltstone unit on the northwest facing slope adjacent to the creek. The horizontally bedded siltstone was exposed continuously through this interval, although exposures were somewhat degraded by weathering. Selected specimens were photographed using a Canon EOS 5D Mark III camera. All recovered specimens are deposited in the collections of the Department of Palaeobiology, Swedish Museum of Natural History (acronym NRM).
Several important type specimens from the Grammatjukku Formation at Aistjakk, originally described by Kautsky (Citation1945) are currently missing from the collections of the Department of Palaeobiology of the NRM. This material includes the type specimens of several species relevant for the current study.
“Upper siltstone member” of the Torneträsk Formation or Grammajukku Formation?
Based on a sequence-stratigraphic analysis of the lower Cambrian (Terreneuvian and Cambrian Series 2) rocks of Scandinavia, Nielsen & Schovsbo (Citation2011) re-evaluated the stratigraphy of the autochtonous Cambrian strata of the Swedish and Norwegian Caledonides, including the Torneträsk area. They proposed to exclude the “upper siltstone member” sensu Thelander (Citation1982) from the Torneträsk Formation and referred this fine-grained interval to the Grammajukku Formation, which is best known from the central Lapland region around Laisvall (Kulling Citation1942; Willdén Citation1980). The authors noted that the Torneträsk Formation thins progressively southwards from Luobákte and that an east-west-trending palaeohigh, the Parnajåkkå–St. Sjöfallet basement High, appears to have existed south of Kiruna. In the Stora Sjöfallet region, siltstones recognized as the “upper siltstone member” by Thelander (Citation1982) rest directly on Proterozoic basement. South of this region, the lower Cambrian strata are divided into the basal sandstone-dominated Laisberg Formation and overlying siltstone-dominated Grammajukku Formation (Kulling Citation1942; Willdén Citation1980).
As discussed by Nielsen and Schovsbo (Citation2011), the “upper siltstone member” at Luobákte is lithologically similar to the Grammajukku Formation at Laisvall (Willdén Citation1980), although it is thinner and contains less sand. At both localities, the succession begins with thin conglomerates containing phosphorite pebbles and terminates with thin fossiliferous limestone units. The fauna of the Grammajukku Formation at Laisvall was described by Kautsky (Citation1945) and Ahlberg (Citation1984), and is referred to the Vergalian–Rausvian H. kjerulfi–O.’ linnarssoni trilobite Zone (Nielsen & Schovsbo Citation2011), the same zone represented by the fauna known from the bioclastic limestone at the top of the “upper siltstone member” at Luobákte (Ahlberg & Bergström Citation1978; Axheimer et al. Citation2007).
The new fossil fauna from the Grammajukku Formation at Loubákte reported here includes trilobites (Ellipsocephalus cf. gripi) and brachiopods (Botsfordia cf. caelata) with a strong affinity to the fauna of the uppermost Grammajukku Formation at Aistjakk, close to Laisvall (Kautsky Citation1945). This observation strengthens the interpretation that the “upper siltstone member” at Luobákte is indeed the lateral equivalent of the Grammajukku Formation, and we adhere to the lithostratigraphic subdivision proposed by Nielsen & Schovsbo (Citation2011).
Fauna of the Grammajukku Formation
Previously, the only fossils documented from the autochthonous succession at Luobákte were trace fossils from various levels in the Torneträsk Formation (Moberg Citation1908; Kulling Citation1960; Thelander Citation1982; Jensen & Grant Citation1998), the problematic Platysolenites and Sabellidites from the “lower siltstone member” (Moberg Citation1908; Jensen & Grant Citation1998) and a trilobite fauna from the uppermost 2 m of the overlying Grammajukku Formation (Moberg Citation1908; Ahlberg Citation1980b, Citation1983; Axheimer et al. Citation2007). The fauna described previously from the uppermost Grammajukku Formation is restricted to five species of trilobites (Chelediscus acifer, Holmia sp., Neocobboldia aff. dentata, Orodes? lapponica (Ahlberg Citation1980b) and Strenuaeva inflata (Ahlberg & Bergström Citation1978); see discussion by (Axheimer et al. Citation2007). However, Axheimer et al. (Citation2007) also mentioned various non-trilobite fossils including the brachiopods Glyptias cf. favosa (Linnarsson Citation1869) and Lingulella? sp., the bradoriid Indiana sp. together with unidentified helcionelloid molluscs, but these taxa have never been illustrated. In addition, Ahlberg (Citation1980a) described the trilobites Orodes? lapponica, Ellipsocephalus cf. gripi (Kautsky Citation1945) and Strenuaeva sp. from a limestone bed in the overlying thrusted rocks of the lowermost nappe, considered to represent the same stratigraphic level.
Our investigations of the siltstones in the lower part of the Grammajukku Formation reveal several new fossiliferous beds yielding brachiopods (Botsfordia cf. caelata and Eoobolus cf. priscus), orthothecid hyoliths and trilobites (Ellipsocephalus cf. gripi and an unidentified holmiid), in an interval hitherto considered unfossiliferous (). Our investigation of the limestone layers at the top of the Grammajukku Formation resulted in a new SSF fauna from the uppermost bioclastic limestone bed. The fauna is moderately diverse with nine species in total, although most specimens are small or fragmentary. However, taxa with originally phosphatic shells (linguliform brachiopods Botsfordia cf. caelata () and Eoobolus cf. priscus (), the tommotiid Lapworthella schodackensis (), the bradoriids Bradoria sp., Mongolitubulus spinosus and indeterminable bradoriid remains () are reasonably well preserved compared to an indeterminable protoconodont element and calcareous taxa that are restricted to poorly resolved internal moulds (trilobites, helcionelloid mollusc: ).
Figure 3. Fossils from the lower part of Grammajukku Formation, Torneträsk Formation, Luobákte, Lapland. A–D. Ellipsocephaus cf. gripi. A. NRM Ar72661, cephalon in dorsal view. B. NRM Ar72662, cephalon in dorsal view. C. NRM 72663, cephalon in dorsal view. D. NRM Ar72664, cephalon in dorsal view. E. Holmia? sp. NRM, Ar72665, possible genal spine with prominent terrace lines. F. Ind. Orthothecid hyolith NRM Mo193048. G, J. Eoobolus cf. priscus (Poulsen Citation1932); G. NRM Br151128, ventral valve interior. J. NRM Br151129, dorsal valve exterior. H, I. Botsfordia cf. caelata (Hall Citation1847), NRM Br151130. H. Partly disarticulated complete shell showing dorsal valve exterior (lower right) and partial ventral valve interior (upper left). I. Dorsal valve exterior showing pustulose external shell ornament

Figure 4. Botsfordia cf. caelata (Hall, Citation1847) from the bioclastic limestone, top of the Grammajukku Formation, Luobákte, Lapland. A, B. NRM Br151131. A. Posterior part of broken ventral valve exterior. B. Detail of posterior margin in oblique view showing outline of brephic shell and open pedicle groove. C, D. NRM Br151132. C. External view of broken ventral valve. D. Detail of shell ornament showing postulose ornament and superimposed radial costellae. E, F. NRM Br151133. E. Ventral valve interior. F. Oblique view showing imprints of muscle scars, pedicle nerves and vascular system. G. NRM Br151134, ventral valve interior showing muscle scars and imprints of pedicle nerves. H. NRM Br151135, Dorsal valve interior showing imprints of muscle scars and short median septum. I, J. NRM Br151136. I. Dorsal valve interior in oblique lateral view showing muscle scars and median septum. J. Oblique posterior view showing profile of brephic shell with two pustules. K, L. NRM Br151137. K. External view of shell fragment. L. Detail of shell surface showing pustulose ornament. Scale bars equal 200 µm in A–C, E–K; 100 µm in D, L

Figure 5. Eoobolus cf. priscus (Poulsen Citation1932) from the bioclastic limestone, top of Grammajukku Formation, Torneträsk Formation, Luobákte, Lapland. A, B. NRM Br151138. A. Fragmentary ventral valve in plan view. B. Oblique view of apex showing outline of brephic shell and pedicle notch. C. NRM Br151139, internal view of fragmentary ventral valve. D. NRM Br1515140, ventral valve interior showing pseudointerarea and median tongue. E. NRM Br151141, ventral valve interior showing pseudointerarea. F, G. NRM Br151142. F. Dorsal valve exterior. G. Detail of pustulose surface sculpture, oblique view. H, I. NRM Br151143. H. Dorsal valve interior showing pseudointerarea and visceral area. I. Oblique view of visceral area showing bisected median tongue and reticulate ornament (indicated by arrows). J. NRM Br 151144, complete dorsal valve in exterior view. K, L. NRM Br151145. K. External view of fragmentary dorsal valve. L. Oblique view of broken shell margin showing columnar shell structure. Scale bars equal 200 µm in A–F, H, J, K; 100 µm in G, I; 20 µm in L

Figure 6. Lapworthella shodackensis (Lochman Citation1956) from the bioclastic limestone, top of Grammajukku Formation, Luobákte, Lapland. A, K. NRM X10299. A. View of concave lateral side of conical sclerite with circular cross-section. K. Detail of shell ornament in oblique apertural view showing denticles on rib crests and growth lines on adapertural slope of ribs. B. NRM X10300, lateral view of spine-shaped sclerite. C. NRM X10301, lateral view of spine-shaped sclerite. D. NRM X10302, lateral view of narrow conical sclerite. E. NRM X10303, sub-symmetrical sclerite with rectangular cross-section viewed from convex surface. F. NRM X10304, asymmetrical sclerite with rectangular cross-section viewed from concave side. G, H. NRM X10305. G. Asymmetrical conical sclerite in apical view. H. Lateral view. I, J. NRM X10306. I. Fragment of sclerite with rectangular cross-section. J. Detail of shell surface in oblique view showing rectangular ornament on adapical slope of ribs. Scale bars equal 50 µm in A–I; 20 µm in J, K
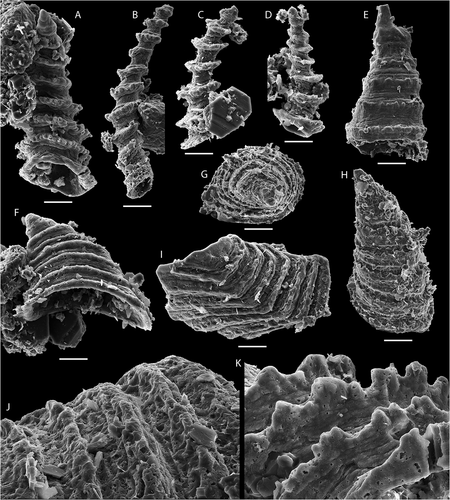
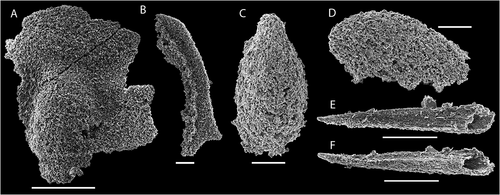
Figure 7. Bradoriid arthropods from the bioclastic limestone, top of Grammajukku Formation, Luobákte, Lapland. A–F. Bradoria sp. A, B. NRM Ar72666. A. Plan view of broken valve fragment. B. Oblique lateral view. C–F. NRM Ar72667. C. Broken valve fragment in oblique view from ventral margin. D. Detail of shell ornament. E. Detail of D showing two types of shell perforations. F. Detail of E with enlargement of open perforation with internal spikes. G–J. Mongolitubulus spinosus (Hinz Citation1987). G. NRM Ar72668, spine tip in lateral view. H. NRM Ar72669, spine with flaring base in lateral view. I, J. NRM Ar72670. I. Spine fragment in lateral view. J. Detail of reticulate ornament with serrated edges. K. NRM Ar72671, valve fragment of bradoriid with spinose margins in lateral view. Scale bars equal 500 µm in A–C; 100 µm in G–I, K; 40 µm in D, J; 10 µm in E; 4 µm in F
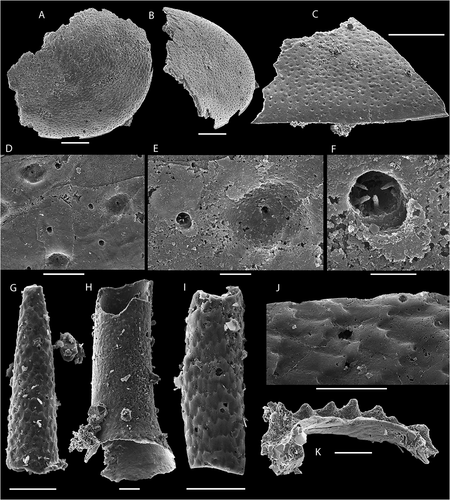
Figure 8. Fossils from the bioclastic limestone, top of Grammajukku Formation, Luobákte, Lapland. A, B. Fragments of ellipsocephalid trilobites. A. NRM Ar72672, cranidium in dorsal view. B. NRM Ar72673, free cheek with small spine. C, D. Laterally compressed helcionelloid mollusc NRM Mo193049. C. Shell in dorsal view. D. Oblique lateral view showing overhanging apex. E, F. Broken protoconodont element NRM X10307. E. Plan view. F. Oblique lateral view. Scale bars equal 1 mm in A; 200 µm in B–F
In the following detailed taxonomic description of the faunal components, we combine the material from both the siltstone layers and the bioclastic limestone, as the two brachiopod taxa are shared between these assemblages.
Trilobites
Five species of trilobites were reported from the uppermost limestone unit of the Grammajukku Formation at Luobákte by Axheimer et al. (Citation2007). Our investigations revealed poorly preserved trilobite remains from the acid-resistant residue of the bioclastic limestone and various trilobite specimens from several beds in the lower part of the siltstone unit. The specimens are flattened and, in some cases, disturbed by minor tectonic deformation connected to small scale movement in the sediment.
Most of the recovered trilobite specimens represent smooth cephala of ellipsocephalids with a tapering bullet-shaped glabella and wide anterior border with a rounded triangular margin (). In outline and relative proportions of the glabella and anterior border, the specimens are similar to Ellipsocephalus gripi (Kautsky Citation1945), originally described from Aistjakk in central Lapland. However, the flattened nature of the specimens from Luobákte make detailed comparison to the highly convex cephalon of E. gripi difficult and leaves the identification uncertain. The acid resistant residue of the bioclastic limestone also includes rare internal moulds of cranidia and free cheeks of ellipsocephalid trilobites () similar to Strenuaeva inflata (Ahlberg & Bergström Citation1978), which was reported from the same level by Axheimer et al. (Citation2007), but the acid-liberated specimens are too poorly preserved to identify with any certainty. A single large isolated genal spine of a trilobite with a pronounced ornament of terrace lines was also found in the siltstone-hosted material from the lower Grammajukku Formation (). This specimen may represent a large holmiid trilobite but further identification is not possible at this time.
Brachiopods
Two distinct brachiopod species were recovered from both the acid-resistant residue of the bioclastic limestone and from the underlying siltstones of the Grammajukku Formation. The first species is characterized by a sub-circular shell and a distinctive coarsely pustulose adult external ornament typical of acrothelids, whereas the second species is a linguloid with a sub-triangular oval shell and a more subdued ornament.
Botsfordia cf. caelata (Hall, Citation1847)
Two brachiopod specimens with a subcircular shell morphology were found in siltstones from the Grammajukku Formation. The best-preserved specimen has a complete dorsal valve c. 5 mm wide in external view with a partial mould of the slightly displaced ventral valve exposed to one side (). The shell surface of the dorsal valve reveals concentric growth lines with a superimposed pattern of pustules in a roughly hexagonal arrangement, increasing in size towards the anterior margin (). Specimens in the acid-resistant residues of the bioclastic limestone have a similar morphology and adult ornament, and are considered to be conspecific with the siltstone-hosted specimens. Despite the specimens in acid residues being fragmentary, they preserve several fine details of the external and internal morphology. The ventral valve has a marginal apex with circular brephic shell overhanging a wide triangular open delthyrium () and a narrow catacline pseudointerarea with a triangular pedicle groove (). Two rounded umbonal muscle scars are situated just anterior of the ventral propareas and a W-shaped impression of middle and outside lateral muscle scars are found closer to the valve center (). Indistinct V-shaped impressions of the pedicle nerves () and vascular lateralia () are preserved in some cases. The gently convex dorsal valve has a marginal apex with two tubercles on the brephic shell (). The dorsal pseudointerarea is very narrow with a broadly triangular pedicle groove raised above the valve floor (). The dorsal visceral area is divided by a short and low median septum (). Dorsal lateral muscle scars converging with the margins of the pedicle groove are preserved in some specimens (). The external shell surfaces of the acid-isolated specimens are commonly exfoliated, but an adult ornament of rounded pustules (12–20 µm in diameter) may be preserved, at least on some portions of the shell (), locally in combination with fine radial capillae (). No micro-ornament was evident on the brephic shell.
The sub-circular shell morphology, open ventral delthyrium with pseudointerarea and the pustulose adult shell ornament clearly indicate that the new material represents a species of the acrothelid Botsfordia (Matthew Citation1891), despite the lack of pitted micro-ornament on the brephic shell. Botsfordia has an almost global distribution in Cambrian Series 2 and the early Miaolingian (Ushatinskaya & Korovnikov Citation2016). Strand (Citation1929) reported the widespread species B. caelata (Hall Citation1847) from the Oslo region of Norway, and Kautsky (Citation1945) described B. thorslundi from the Grammatjukku Formation of the Laisvall area in central Lapland. Although only the external characteristics of the Norwegian material of Botsfordia are known, the better-illustrated B. thorslundi has been largely ignored by later researchers. Kautsky (Citation1945) distinguished B. thorslundi from B. caelata based on the more regular arrangement of pustules in oblique intersecting rows rather than along growth lines. The material described from siltstones at Luobákte appears to have a regular ornament of external pustules more similar to that described from B. thorslundi. However, we note that the ornament of B. caelata is variable and may approach that illustrated for B. thorslundi (Skovsted et al. Citation2017) and, based on the otherwise remarkable morphological similarity, the two taxa may prove to be synonyms. The figured specimens of Botsfordia thorslundi (Kautsky Citation1945, pl. 9, , pl. 10, ) are held in the Swedish Museum of Natural History, although some of the best-preserved specimens, unfortunately, are among the type specimens currently held by Andor Nyers (Örebro) and could not be reinvestigated.
Glyptias cf. favosa (Linnarsson Citation1869) was reported but not illustrated from the bioclastic limestone at Luobákte by Axheimer et al. (Citation2007). The material described here differs from Glyptias by the pustulose shell ornament, sub-circular shell shape and lack of elevated posterolateral muscle fields in dorsal valves.
Eoobolus cf. priscus (Poulsen Citation1932).
The second brachiopod species present in the Cambrian succession at Luobákte is a lingulid with a gently biconvex oval shell and V-shaped posterior ventral margin. Specimens in siltstone are up to 6 mm long and preserve either the internal () or external surface (), although neither in great detail. Ventral valve length is approximately 133% of valve width (), whereas the dorsal valve length is approximately 118% of valve width (). Acid-isolated specimens from the bioclastic limestone are smaller and almost invariably fragmentary, but preserve details of shell ornament and internal morphology (). The posterior margin of the ventral valve forms an angle of about 78–92° (). The ventral brephic shell is oval with an arcuate posterior notch reflecting the pedicle groove (). The ventral pseudointerarea is well developed with a deeply incised pedicle groove between raised propareas bisected by pronounced flexure lines (). The pedicle groove is V-shaped, forming an angle of 10–18° and lies roughly at the same level as the visceral area, which is raised slightly above the valve floor and extended anteriorly by a straight-sided median tongue (). The dorsal valve is oval with a crescentic brephic shell (). The dorsal pseudointerarea is raised slightly over the valve floor and has a very wide median groove and narrow propareas with indistinct flexure lines (). The dorsal visceral area is thickened with a narrow, slightly bisected median tongue and a superimposed reticulate surface pattern (). The external surface of both valves is poorly preserved but, in some cases, has a fine ornament of uniformly small (average diameter c. 10 µm) randomly arranged pustules (). No micro-ornament was detectable on the brephic shell. Along broken valve margins or where outer shell layers have been removed, an acrotretid-like columnar shell structure is locally preserved with regularly spaced orthogonal, hollow columns (diameter of 6–8 µm; ).
The general morphology, sharp angle of the ventral posterior margin and the pustulose adult shell ornament are typical of the globally distributed Eoobolus (Matthew Citation1902). However, the pitted larval shell ornament, which is characteristic of Eoobolus, was not observed, although this might be a consequence of poor surface preservation. Most specimens from Luobákte appear to be partly exfoliated or covered by sediment grains (). The pits of the larval shell in Eoobolus are typically less than 1 µm in diameter (Skovsted & Holmer Citation2005; Ushatinskaya & Korovnikov Citation2014) and the much larger pustules of the adult shell ornament are only visible in a few specimens from Luobákte. Similarly, no pustulose larval shell ornament is evident in co-occurring shells of Botsfordia although this is a characteristic feature of all known acrothelid species (Claybourn et al. Citation2020). In shell shape, apical angle of the ventral valve and in the morphology of both ventral and dorsal pseudointerareas, the shells from Luobákte are very similar to Eoobolus priscus (Poulsen Citation1932), originally described from Cambrian Stage 4 of northeast Greenland (Skovsted & Holmer Citation2005). Morphologically similar brachiopods have been described from Cambrian Stage 3 to Miaolingian strata worldwide, in some cases referred to E. priscus, but the high variability of these brachiopods and the unreliable preservation of key characters, such as larval shell ornament, means that it is currently difficult to confidently identify species of Eoobolus (Balthasar et al. Citation2009). Consequently, the material from Luobákte is here referred to Eoobolus cf. priscus. The brachiopod Lingulella? sp., reported from Luobákte by Axheimer et al. (Citation2007) may represent the same taxon.
Streng et al. (Citation2008b) described Eoobolus? sp. aff. E. priscus from the Forsemölla Limestone (Miaolingian) at Gislövshammar in Skåne, Sweden. The affinity of this taxon to Eoobolus in general and E. priscus in particular was questioned by Streng et al. (Citation2008b), primarily based on the presence of a columnar shell structure in the specimens from Gislövshammar. However, a similar columnar shell structure has since been described from specimens of E. priscus from Greenland and Newfoundland (Peel et al. Citation2016; Skovsted et al. Citation2017) and the use of shell structure for taxonomic determination has also been criticized (Ushatinskaya Citation2012; Ushatinskaya & Korovnikov Citation2014). The specimens from Luobákte differ from Eoobolus? sp. aff. E. priscus by the V-shaped pedicle groove and larger propareas in ventral valves and by the less well developed dorsal median tongue and, consequently, they probably represent a different species. Kautsky (Citation1945) described two lingulid species, Obolus (Westonia) lapponicus (Kautsky Citation1945) and Lingulella (Lingulepis) westergårdi (Kautsky Citation1945) from Aistjakk in central Lapland. Reinvestigation of this material (CBS, personal observation 2020) suggests that these specimens belong to a single species and may be conspecific with the specimens from Luobákte and, hence, to Eoobolus cf. priscus. In fact, Regnell (Citation1948) noted the similarity between the specimens from Aistjakk and the type material of E. priscus from Greenland. In older literature, numerous vaguely similar lingulid species have been reported from lower Cambrian strata and glacial erratics of Scandinavia (see for example Wiman Citation1903 and summary by Walcott Citation1912, p. 146). Some of these may be equivalent to Eoobolus cf. priscus from Luobákte but this is difficult to evaluate without detailed reinvestigation of relevant type material.
Tommotiids
Lapworthella schodackensis (Lochman Citation1956)
Cone-shaped tommotiid sclerites with prominent co-marginal ribs are conspicuous components of the fauna of the bioclastic limestone (). The sclerites are highly variable in cross-section and rate of expansion and conform to Lapworthella (Cobbold Citation1921). They are, however, generally very small compared to published records of lapworthellid sclerites, with a maximum height of <500 µm. Some specimens have a rectangular cross-section and a relatively high rate of expansion (), but the majority of sclerites are very slender cones () or spines () with rounded or oval cross-section. The co-marginal growth sets are 25–50 µm wide (40–20 ribs/mm) and the ribs are crowned by rows of densely spaced, slightly irregular denticles (c. 50–140 denticles/mm; ). The apertural slope of the ribs is ornamented by fine growth lines (), whereas the apical inter-rib groove locally bears a fine reticulate ornament ().
Two species of Lapworthella have been reported from Scandinavia to date: L. cornu (Wiman Citation1903), originally based on two sclerites from a glacial erratic collected at Billuden in Uppland, Sweden (Bengtson Citation1980), but also occurring in Avalonia (Hinz Citation1987; Landing et al. Citation2008); and L. bornholmiensis (Poulsen Citation1942) from the Miaolingian of Bornholm, Skåne and Bohuslän in southern Scandinavia (Devaere & Skovsted Citation2021 [this volume]). The high morphological variability of Lapworthella sclerites has often been emphasized and, traditionally, species have been defined based on differences in ornament, particularly the presence/absence and density of denticles on the co-marginal ribs (Landing Citation1984; Bengtson et al. Citation1990), but the ornaments on the apical inter-rib groove and apertural slope of the ribs are also useful because they reflect differences in the growth regimen of the sclerites (Devaere & Skovsted Citation2017). The dense rows of denticles along the co-marginal ribs of the sclerites from Luobákte differentiates the material from species lacking denticles, such as L. bornholmiensis, but also from denticulate species with fewer denticles/mm, such as L. cornu (12–25) (Landing et al. Citation2008; Devaere & Skovsted Citation2017). The high density of denticles in the Luobákte sclerites conforms with L. schodackensis (Lochman Citation1956) from Cambrian Stage 4 strata of eastern Laurentia and Siberia, although the density in this species appears to vary widely (35–144 in material from New York State [Landing Citation1984]; 28–43 in specimens from Siberia [Rozanov et al. Citation2010]; 25–77 in specimens from Greenland [Devaere & Skovsted Citation2017]). More importantly, the reticulate ornament on the inter-rib groove and fine growth lines on the apertural slope of the ribs also conform with the pattern in L. schodackensis (Devaere & Skovsted Citation2017), supporting assignment to this species. The lack of any larger specimens in the Luobákte material is unique to this collection, although a dominance of small specimens was reported by Landing (Citation1984) for L. schodackensis of New York State.
Recently, morphologically distinct sclerite types have been recognized for Lapworthella schodackensis and L. bornholmiensis (Devaere & Skovsted Citation2017, Citation2021 [this volume]). The collection from Luobákte includes rare bilaterally symmetrical sclerites with a rectangular cross-section and a possible double apex (), which roughly corresponds to the A-type sclerites; rapidly expanding asymmetrical sclerites with elongate rectangular aperture () similar to B-type sclerites; asymmetrical, cone-shaped sclerites with oval cross-section (); and slender spine-shaped sclerites with circular cross-section () that may correspond to D-type sclerites as defined by Devaere and Skovsted (Citation2017, Citation2021). However, a more detailed study of a larger collection would be required to elucidate the scleritome structure of L. schodackensis from Luobákte.
Bradoriids
Bradoriid arthropod remains are among the most common elements in the SSF fauna from the bioclastic limestone (). These include three potential taxa: fragments of large, highly convex punctate bradoriid valves (), ornamented spines (), and rare fragments of a species with small spines along the valve margin ().
Bradoria sp.
Most of the preserved valve fragments bear a characteristic punctate ornament and presumably belong to the same taxon (). As no complete specimens were found, the morphology of this species remains uncertain but the largest valve fragments reveal a uniform, highly convex valve with a seemingly oval or circular outline (). Along some portions of the margin, a narrow admarginal ridge is separated from the valve proper by a sharp furrow (). The smooth valve surface is interrupted by numerous shallow pits or cup-shaped depressions with a diameter of c. 25 µm. A small circular pore with a diameter of c. 2 µm is situated in the centre of each pit. In addition, circular pores of a slightly larger diameter (c. 5 µm) open directly at the valve surface between the pits (). Some of these latter pores preserve a series of 1–5 radially arranged spikes (about 500 nm in diameter) converging close to the centre of the pore opening (). Both pits and larger perforations seem to be distributed randomly at a similar density over the valve surface.
The large, highly convex valve morphology with a very narrow or only partly developed admarginal ridge in combination with a pitted valve surface is reminiscent of Bradoria (Matthew Citation1899). This genus is widespread in Cambrian Stage 3–4 rocks of Avalonia and has also been reported from Laurentia, Australia and South China (Siveter & Williams Citation1997; Williams & Siveter Citation1998; Zhang Citation2007). Various bradoriid taxa have been described from the lower Cambrian of Sweden (see review by Streng et al. Citation2008a, p. 12) and these include several taxa that are at least superficially similar to the bradoriid shell fragments from Luobákte. Two species have been described from the Grammatjukku Formation in central and southern Lapland: Indianites ovoides (Kautsky Citation1945) (Kautsky Citation1945) from Aistjakk near Laisvall in central Lapland; and Walcottella? aff. apicalis (Ulrich & Bassler Citation1931) (Streng et al. Citation2008a) from Bergmyrhobben close to Storuman in southern Lapland. The better-known species, Walcottella? aff. apicalis has a pitted shell surface, but the average diameter of individual pits is about 100 µm, compared to only 25 µm in the Luobákte material. The presence of a large anterodorsal node and a wide, flat border is also different from the new specimens described here. Indianites ovoides is only known from a single specimen (Kautsky Citation1945, pl. 27, ), which has a large ovoid valve with a narrow border, reminiscent of the specimens described here, but with a smooth or finely wrinkled surface ornament (Kautsky Citation1945, p. 149). The type and only specimen belongs to the Swedish Museum of Natural History (NRM Ar9056) but is, unfortunately, among the specimens currently held by Andor Nyers (Örebro) and could not be re-examined.
Several other bradoriid species with a broadly similar valve morphology were described by Wiman (Citation1903) from glacial erratics supposedly derived from Cambrian strata on the subsurface of the Bothnian Sea (see review by Streng et al. Citation2008a). One of these species, originally described as Bradorona nitida (Wiman Citation1903), is a large, inflated bradoriid with a small anterodorsal node, a very narrow admarginal ridge and a pitted valve surface (Wiman Citation1903, p. 48, pl. 1, figs 34, 36–37), which is very similar to the new specimens from Luobákte. Although considered to represent a species of Liangshanella (Huo Citation1956) by Streng et al. (Citation2008a), the morphological characteristics of this species, in particular the fine pitting and the very narrow admarginal ridge, are more characteristic of Bradoria.
As indicated above, the general morphology of the material from Luobákte suggests an affinity to Bradoria, although the fragmentary nature of the specimens precludes detailed comparison to any particular species. Indiana sp., reported from Luobákte by Axheimer et al. (Citation2007) probably represents the same taxon. It may be worth noting that the material described here preserves various details of the surface ornament that lack equivalency in any described bradoriid species (two types of perforations, internal spikes within open pores). However, we note that few species of Bradoria have been studied using SEM, and it is possible that equivalent details will be revealed in the future, allowing a better comparison of the Luobákte material to established species.
Mongolitbulus spinosus (Hinz Citation1987)
In addition to the punctate bradoriid valve fragments, hollow, thin-walled ornamented tubes interpreted as broken spines of a spinose bradoriid occur in the studied material (). Some of the smallest specimens (diameter c. 100 µm) appear to taper to a point (), whereas some larger specimens (diameter up to c. 300 µm) have a flared base with uneven, broken margins (diameter up to 500 µm; ). These tubular specimens bear a fine cancellate ornament composed of longitudinally extended polygons with scalloped or serrated edges ().
Ornamented, hollow spines are common constituents of SSF faunas in the Cambrian and are commonly referred to Mongolitubulus (Missarzhevsky Citation1977), although as presently defined, this is probably a form taxon including fossils derived from various pan-arthropods (Skovsted & Peel Citation2001; Dzik Citation2003; Li et al. Citation2012; Caron et al. Citation2013). However, species with an originally phosphatic shell and a well-developed flared base, are generally considered to represent bradoriid arthropods (Skovsted et al. Citation2006; Topper et al. Citation2007, Citation2013; Betts et al. Citation2014; Kouchinsky et al. Citation2015). Individual species of Mongolitubulus are defined by a combination of spine shape and ornament. The cancellate ornament and straight, evenly tapered spine morphology of the specimens from Luobákte are very similar to Mongolitubulus spinosus (Hinz Citation1987), which is known from the lower Cambrian of England (under the name Rushtonites Hinz Citation1987) and Siberia (Kouchinsky et al. Citation2015), and the material is referred to this species here.
Bradoriida indet
Finally, a few fragmentary specimens of a presumed bradoriid with a ventral margin ornamented by a distinct row of regularly spaced cone-shaped spines are found in the assemblage (). These specimens are unlikely to belong to either of the bradoriid species discussed above and probably represent a separate taxon. However, too few morphological details are preserved to identify this further and the specimens are referred to Bradoriida indet.
Other fossils
Straight, tubular fossils, partly filled with yellow-weathering pyrite were found in the siltstone layers (), and although not well enough preserved for detailed identification, these tubes are interpreted to represent orthothecid hyoliths. In the acid-resistant residue of the bioclastic limestone, a few further taxa are identified in addition to brachiopods, bradoriids and lapworthellid sclerites. A single internal mould of a laterally compressed helcionelloid mollusc () and a single partial protoconodont element () were also recovered, but are also too poorly preserved for further identification and are left in open nomenclature.
Biostratigraphic and palaeobiogeographic significance
The biostratigraphy of the lower Cambrian (Series 1 and 2) of Baltica is poorly constrained because fossils are rare at many localities and stratigraphic levels (Nielsen & Schovsbo Citation2007, Citation2011) and few absolute age controls are available for early Cambrian biozones, globally. In Cambrian Series 2, biostratigraphic resolution is generally based on trilobites; non-trilobite biostratigraphy remains in its infancy. The trilobites reported from the uppermost Grammajukku Formation at Luobákte are not strictly age diagnostic, but Axheimer et al. (Citation2007) suggested that they belong to the Ornamentaspis? linnarssoni Assemblage Zone based on the generic composition and the relative stratigraphic position of the fossiliferous outcrops at the top of the “lower Cambrian”. The discovery of the new fauna from the same level at Loubákte complements the known trilobite fossil record and suggests that the taxonomic diversity of the Cambrian faunas of Baltica was higher than previously considered.
The SSF fauna reported herein is the first of its type ever reported from the autochthonous sequence of the Caledonian mountains in Sweden, although some SSF taxa have been reported by Rosén (Citation1919) and Kautsky (Citation1945). The scarcity of comparable faunas described previously reduces the biostratigraphic utility of the material. However, a growing body of evidence suggests that brachiopods and other SSF taxa may be more widespread and diverse in strata of Cambrian Series 2 in Baltica than previously considered and, with renewed study, these taxa may help improve biostratigraphic and palaeogeographic resolution. In this way, the new fauna from Luobákte may provide important baseline data for future studies.
In general, many non-trilobite taxa from the Cambrian Series 1 and 2 appear to have relatively long stratigraphic ranges. However, detailed investigation of multiple sections spanning similar stratigraphic intervals throughout a region may provide enough data for meaningful biostratigraphic resolution among SSF taxa. The early Cambrian succession of South Australia provides an example of an area where SSF taxa, primarily tommotiids and bradoriids, have been successfully employed to define small shelly fossil biozones (Betts et al. Citation2016, Citation2017, Citation2018). A growing body of evidence, including this report, suggests that tommotiids, particularly species of Lapworthella, are an important component of early Cambrian faunas also in Baltica. Devaere and Skovsted (Citation2021 [this volume]) revised Lapworthella bornholmiensis from Bornholm and southern Sweden and noted that the range of this taxon spans uppermost Series 2 and Miaolingian strata. So far, L. schodackensis has been found only in Baltica at Luobákte, but this species is common in medial Cambrian Stage 4 rocks of eastern Laurentia (Lochman Citation1956; Landing Citation1984; Skovsted Citation2006; Devaere & Skovsted Citation2017) and in roughly coeval strata of Siberia (Rozanov et al. Citation2010). The third lapworthellid species found in Baltica, L. cornu, is currently known only from glacial erratics, but this species is better known from Britain where it occurs in the early Stage 4 Strenuella sabulosa trilobite Zone. Thus, the stratigraphic ranges of lapworthellid tommotiids in Baltica may provide the embryo of a new small shelly fossil biostratigraphy, although this would need to be developed in much greater detail.
The discovery of new fossil layers yielding brachiopods and the trilobite Ellipsocephalus cf. gripi in the lower part of the Grammajukku Formation may be of at least regional biostratigraphic significance because, previously, Ellipsocephalus gripi has been found only in the Grammajukku Formation at Laisvall in southern Lapland. At Laisvall, Ellipsocephalus gripi occurs in the uppermost Grammajukku Formation (Kautsky Citation1945; Willdén Citation1980; Ahlberg Citation1983). At Luobákte, we document a possible occurrence of the species in the lower part of this formation, in addition to a record from the conglomerate at the base of the Grammajukku Formation at Vaivantjåkka, north of Torneträsk (Ahlberg Citation1980b). Furthermore, the discovery of the new fossiliferous beds in the lower Grammajukku Formation suggests that future concerted collecting efforts through this interval might yield palaeontological data of significance for biostratigraphy and/or the evolution of faunas in Cambrian Stage 4.
The discovery of the new fossils from Luobákte extends the known geographic ranges of several taxa, primarily Lapworthella schodackensis and Mongolitubulus spinosus, with the first report of these taxa from the palaeocontinent Baltica. The occurrence of L. schodackensis provides a link to Laurentian faunas of Cambrian Stage 4 (Lochman Citation1956; Landing Citation1984; Devaere & Skovsted Citation2017) and Siberia (Rozanov et al. Citation2010). However, although M. spinosus is also known from Siberia (Kouchinsky et al. Citation2015) and potentially as a Small Carbonaceous Fossil (SCF) from Laurentia (Butterfield & Nicholas Citation1996; Caron et al. Citation2013), it is best known from Avalonian Great Britain (Hinz Citation1987). The possible link with Avalonia mirrors the distribution of the eodiscid trilobites Neocobboldia aff. dentata and Chelediscus acifer reported from Luobákte by Axheimer et al. (Citation2007).
This study provides a glimpse into the early Cambrian marine life of Baltica, introducing a small portion of the life that existed in the shallow seas around the continent. We envisage an active bottom-dwelling community grazing and burrowing in the soft sediments on the inner continental shelf, within the photic zone of a temperate sea. Although the recovered fauna of the upper Grammajukku Formation is restricted to taxa with biomineralized hard parts and likely represents only a fraction of the total ecosystem, the fauna consists of taxa with diverse feeding modes. These include taxa with a presumed epibenthic grazing life mode, such as small helcionelloid molluscs and the tommotiid Lapworthella, and benthic suspension feeders in the form of brachiopods. Benthic deposit feeders are represented by orthothecid hyoliths, eodiscid and ellipsocephalid trilobites, together with the bradoriids Bradoria sp. and Mongolitubulus spinosus. Associated protoconodonts were probably planktic micropredators and the rare holmiid trilobites represent macroscopic benthic predators at the top of the food chain. Thus, the fauna includes every trophic level from primary consumers (grazers and suspension feeders) through detritus feeders to predators.
The presence of carbonates, a relatively diverse fauna and the absence of glaciogene sedimentary features implies that Baltica was not glaciated during the early Cambrian, supporting previous studies invoking a warm climate for the early Cambrian (Hearing et al. Citation2018; Wotte et al. Citation2019). Further studies involving detailed and large-scale sampling of the Luobákte fauna will likely provide new insights into one of the earliest multicellular faunas on Earth from a mid-palaeolatitude setting.
Acknowledgments
We thank the staff at the Abisko Scientific Research Station for valuable help and advice during fieldwork in August 2020. Steffen Kiel (Stockholm) is thanked for help with translation from German to English. This work was supported by funding from the Swedish Museum of Natural History and the Swedish Research Council (VR grant number 2016-04610 to C.B.S; VR grant number 2018-04527 to S.M., VR grant number 2017-05183 to T.T. and VR grant number 2019-4061 to V.V).
Disclosure statement
No potential conflicts of interest were reported by the authors.
Additional information
Funding
References
- Ahlberg, P., 1980a: Early Cambrian trilobites from Mount Luopakte, northern Sweden. Sveriges geologiska undersökning C 765, 1–12.
- Ahlberg, P., 1980b: Early Cambrian trilobites from northern Scandinavia. Norsk Geologisk Tidsskrift 60, 153–159.
- Ahlberg, P., 1983: A lower Cambrian trilobite fauna from Jämtland, central Scandinavian Caledonides. Geologiska Föreningens i Stockholm Förhandlingar 105, 349–361. doi:https://doi.org/10.1080/11035898309454573.
- Ahlberg, P., 1984: Lower Cambrian trilobites and biostratigraphy of Scandinavia. Lund Publications in Geology 22, 1–37.
- Ahlberg, P. & Bergström, J., 1978: Lower Cambrian ptychopariid trilobites from Scandinavia. Sveriges geologiska undersökning Ca 49, 1–41 pp.
- Ahlberg, P., Bergström, J., & Johansson, J., 1986: Lower Cambrian olenellid trilobites from the Baltic faunal province. Geologiska Föreningens i Stockholm Förhandlingar 108, 39–56. doi:https://doi.org/10.1080/11035898609453745.
- Axheimer, N., Ahlberg, P., & Cederstrom, P., 2007: A new lower Cambrian eodiscoid trilobite fauna from Swedish Lapland and its implications for intercontinental correlation. Geological Magazine 144, 953–961. doi:https://doi.org/10.1017/S0016756807003597.
- Balthasar, U., 2009: The brachiopod Eoobolus from the early Cambrian mural formation (Canadian Rocky Mountains). Paläontologische Zeitschrift 83, 407–418. doi:https://doi.org/10.1007/s12542-009-0026-4.
- Balthasar, U., Skovsted, C.B., Holmer, L.E., & Brock, G.A., 2009: Homologous skeletal secretion in tommotiids and brachiopods. Geology 37, 1143–1146. doi:https://doi.org/10.1130/G30323A.1.
- Bengtson, S., 1968: The problematic genus Mobergella from the lower Cambrian of the Baltic area. Lethaia 1, 325–351. doi:https://doi.org/10.1111/j.1502-3931.1968.tb01625.x.
- Bengtson, S., 1970: The lower Cambrian fossil Tommotia. Lethaia 3, 363–392. doi:https://doi.org/10.1111/j.1502-3931.1970.tb00829.x.
- Bengtson, S., 1980: Redescription of the Lower Cambrian Lapworthella cornu. Geologiska Föreningen i Stockholm Förhandlingar, 102, 53–55.
- Bengtson, S., 2004: Early skeletal fossils. The Paleontological Society Papers 10, 67–78. doi:https://doi.org/10.1017/S1089332600002345.
- Bengtson, S., Morris, S.C., Cooper, B., Jell, P.A., & Runnegar, B., 1990: Early Cambrian fossils from South Australia. Memoir of the Association of Australasian Palaeontologists 9, 1–364.
- Bergström, J. & Ahlberg, P., 1981: Uppermost lower Cambrian biostratigraphy in Scania, Sweden. Geologiska Föreningen I Stockholm Förhandlingar 103, 193–214. doi:https://doi.org/10.1080/11035898109454518.
- Betts, M.J., Paterson, J.R., Jacquet, S.M., Andrew, A.S., Hall, P.A., Jago, J.B., Jagodzinski, E.A., Preiss, W.V., Crowley, J.L., Brougham, T., Mathewson, C.P., García-Bellido, D.C., Topper, T.P., Skovsted, C.B., & Brock, G.A., 2018: Early Cambrian chronostratigraphy and geochronology of South Australia. Earth-Science Reviews 185, 498–543. doi:https://doi.org/10.1016/j.earscirev.2018.06.005.
- Betts, M.J., Paterson, J.R., Jago, J.B., Jacquet, S.M., Skovsted, C.B., Topper, T.P., & Brock, G.A., 2016: A new lower Cambrian shelly fossil biostratigraphy for South Australia. Gondwana Research 36, 176–208. doi:https://doi.org/10.1016/j.gr.2016.05.005.
- Betts, M.J., Paterson, J.R., Jago, J.B., Jacquet, S.M., Skovsted, C.B., Topper, T.P., & Brock, G.A., 2017: Global correlation of the early Cambrian of South Australia: shelly fauna of the Dailyatia odyssei zone. Gondwana Research 46, 240–279. doi:https://doi.org/10.1016/j.gr.2016.05.005.
- Betts, M.J., Topper, T.P., Valentine, J.L., Skovsted, C.B., Paterson, J.R., & Brock, G.A., 2014: A new early Cambrian bradoriid (Arthropoda) assemblage from the northern flinders ranges, South Australia. Gondwana Research 25, 420–437. doi:https://doi.org/10.1016/j.gr.2013.05.007.
- Butterfield, N.J. & Nicholas, C.J., 1996: Burgess shale-type preservation of both non-mineralizing and ‘shelly’ Cambrian organisms from the Mackenzie mountains, northwestern Canada. Journal of Paleontology 70, 893–899. https://www.jstor.org/stable/1306492
- Caron, J.B., Smith, M.R., & Harvey, T.H.P., 2013: Beyond the burgess shale: Cambrian microfossils track the rise and fall of hallucigeniid lobopodians. Proceedings of the Royal Society B 280, 20131613. doi:https://doi.org/10.1098/rspb.2013.1613.
- Cederström, P., Ahlberg, P., Nilsson, C.H., Ahlgren, J., & Eriksson, M.E., 2011: Moulting, ontogeny and sexual dimorphism in the Cambrian ptychopariid trilobite Strenuaeva inflata from the northern Swedish Caledonides. Palaeontology 54, 685–703. doi:https://doi.org/10.1111/j.1475-4983.2010.01021.x.
- Claybourn, T.M., Skovsted, C.B., Holmer, L.E., Pan, B., Myrow, P.M., Topper, T.P., & Brock, G.A., 2020: Brachiopods from the Byrd group (Cambrian Series 2, Stage 4) central transantarctic mountains, East Antarctica: biostratigraphy, phylogeny and systematics. Papers in Palaeontology 6, 349–383. doi:https://doi.org/10.1002/spp2.1295.
- Cobbold, E.S., 1921: The Cambrian horizons of Comley (Shropshire) and their brachiopoda. Pteropoda. Gasteropoda, etc. Quarterly Journal of the Geological Society 76, 325–386. doi:https://doi.org/10.1144/GSL.JGS.1920.076.01-04.10.
- Demidenko, Y.E. & Parkhaev, P.Y., 2020: Morphology, paleobiology and systematics of the mobergellids, a group of Cambrian problematic fossils. Paleontological Journal 54, 441–458. doi:https://doi.org/10.1134/S0031030120050044.
- Devaere, L. & Skovsted, C.B., 2017: New early Cambrian sclerites of Lapworthella schodackensis from NE Greenland: advancements in knowledge of lapworthellid taxonomy, sclerite growth and scleritome organization. Geological Magazine 154, 1061–1072. doi:https://doi.org/10.1017/S0016756816000698.
- Devaere, L. & Skovsted, C.B., 2021 [this volume]: The last of the tommotiids: lapworthella bornholmiensis (Poulsen, 1942) from Cambrian Stage 4 to Guzhangian (Miaolingian) strata of Bornholm and southern Sweden. GFF.
- Dies Álvarez, M.E., Gozalo, R., Cederström, P., & Ahlberg, P., 2008: Bradoriid arthropods from the lower-middle Cambrian of Scania, Sweden. Acta Palaeontologica Polonica 53, 647–656. doi:https://doi.org/10.4202/app.2008.0409.
- Dzik, J., 1994: Evolution of ‘small shelly fossils’ assemblages of the Early Paleozoic. Acta Palaeontologica Polonica 39, 247–313.
- Dzik, J., 2003: Early Cambrian lobopodian sclerites and associated fossils from Kazakhstan. Palaeontology 46, 93–112. doi:https://doi.org/10.1111/1475-4983.00289.
- Ebbestad, J.O.R., Ahlberg, P., & Høyberget, M., 2003: Redescription of Holmia inusitata (Trilobita) from the lower Cambrian of Scandinavia. Palaeontology 46, 1039–1054. doi:https://doi.org/10.1111/1475-4983.00332.
- Esakova, N.V. & Zhegallo, E.A., 1996: Biostratigraphy and fauna of the lower Cambrian of Mongolia. Trudy, Sovmestnaya Rossiysko-Mongol’skaya Paleontologicheskaya Ekspeditsiya 46, 214.
- Freeman, R.L., Dattilo, B.F., & Brett, C.E., 2019: An integrated stratinomic model for the genesis and concentration of “small shelly fossil”-style phosphatic microsteinkerns in not-so-exceptional conditions. Palaeogeography, Palaeoclimatology, Palaeoecology 535, 109344. doi:https://doi.org/10.1016/j.palaeo.2019.109344.
- Grant, S.W., 1990: Shell structure and distribution of Cloudina, a potential index fossil for the terminal Proterozoic. American Journal of Science 290, 261.
- Hall, J.A., 1847: Paleontology of New York 1. C, 338. van Benthuysen, Albany.
- Hearing, T.W., Harvey, T.H.P., Williams, M., Leng, M.J., Lamb, A.L., Wilby, P.R., Gabbott, S.E., Pohl, A., & Donnadieu, Y., 2018: An early Cambrian greenhouse climate. Science Advances 4, eaar5690. doi:https://doi.org/10.1126/sciadv.aar5690.
- Hinz, I., 1987: The Lower Cambrian microfauna of Comley and Rushton, Shropshire, England. Palaeontographica. Abteilung A, Paläozoologie, Stratigraphie 198, 41–100.
- Høyberget, M., Ebbestad, J.O.R., & Funke, B., 2019: Re-evaluation of the stratigraphically important olenellid trilobite Holmia cf. mobergi from the Cambrian Series 2, Stage 3 and its implications for the lower Cambrian stratigraphy in the Mjøsa area, Norway. Norwegian Journal of Geology 99, 1–30. doi:https://doi.org/10.17850/njg99-1-04.
- Huo, S.C., 1956: Brief notes on lower Cambrian Archaeostraca from Shensi and Yunnan. Acta Palaeontologica Sinica 4, 425–445. (in Chinese with English abstract).
- Jensen, S. & Grant, S.W., 1998: Trace fossils from the dividalen group, northern Sweden: implications for early Cambrian biostratigraphy of Baltica. Norsk Geologisk Tidsskrift 78, 305–317.
- Jeppsson, L., Anehus, R., & Fredholm, D., 1999: The optimal acetate buffered acetic acid technique for extracting phosphatic fossils. Journal of Paleontology 73, 964–972. doi:https://doi.org/10.1017/S0022336000040798.
- Kautsky, F., 1945: Die Unterkambrische Fauna vom Aistjakk in Lapland. Geologiska Föreningen I Stockholm Förhandlingar 67, 129–211. doi:https://doi.org/10.1080/11035894509446095.
- Kouchinsky, A., Bengtson, S., Clausen, S., & Vendrasco, M.J., 2015: An early Cambrian fauna of skeletal fossils from the Emyaksin formation, northern Siberia. Acta Palaeontologica Polonica 60, 421–512. doi:https://doi.org/10.4202/app.2012.0004.
- Kouchinsky, A., Bengtson, S., Runnegar, B., Skovsted, C., Steiner, M., & Vendrasco, M., 2012: Chronology of early Cambrian biomineralization. Geological Magazine 149, 221–251. doi:https://doi.org/10.1017/S0016756811000720.
- Kulling, O., 1942: Grunddragen av fjällkedjerandens bergbyggnad inom Västerbottens län. Sveriges geologiska undersökning C 445, 1–319 pp.
- Kulling, O., 1960: On the caledonides of Swedish Lapland. Sveriges geologiska undersökning Ba 16, 150–177 pp. In: Description to accompany the map of the Pre-Quaternary rocks of Sweden.
- Landing, E., 1984: Skeleton of lapworthellids and the suprageneric classification of tommotiids (Early and Middle Cambrian phosphatic problematica). Journal of Paleontology 58, 1380–1398. http://www.jstor.org/stable/1304888
- Landing, E., Johnson, S.C., & Geyer, G., 2008: Faunas and Cambrian volcanism on the Avalonian marginal platform, southern New Brunswick. Journal of Paleontology 82, 884–905. doi:https://doi.org/10.1666/07-007.1.
- Li, G.X., Steiner, M., Zhu, M.Y., & Zhao, X., 2012: Early Cambrian eodiscoid trilobite Hupeidiscus orientalis from South China: ontogeny and implications for affinities of Mongolitubulus-like sclerites. Bulletin of Geosciences 87, 159–169. doi:https://doi.org/10.3140/BULL.GEOSCI.1224.
- Linnarsson, J.G.O., 1869: On some fossils found in the Eophyton Sandstone, at Lugnås, in Sweden. Geological Magazine 6, 393–406. doi:https://doi.org/10.1017/S0016756800162910.
- Lochman, C., 1956: Stratigraphy, paleontology, and paleogeography of the Elliptocephala asaphoides strata in Cambridge and Hoosick Quadrangles, New York. Geological Society of America Bulletin 67, 1331–1396. doi:https://doi.org/10.1130/0016-7606(1956)67[1331:SPAPOT]2.0.CO;2.
- Matthew, G.F., 1891: Illustrations of the fauna of the St. John Group No. 5. Royal Society of Canada, Proceedings and Transactions (Series 1, Section 4) 8, 123–166. doi:https://doi.org/10.5962/bhl.title.41116.
- Matthew, G.F., 1899: Preliminary notice of the Etcheminian Fauna of Cape Breton. Bulletin of the Natural History Society of New Brunswick 4, 198–208.
- Matthew, G.F., 1902: Notes on Cambrian faunas. Transactions of the Royal Society of Canada (Series 2, Section 4) 18, 93–112. doi:https://doi.org/10.5962/bhl.title.63589.
- Matthews, S.C. & Missarzhevsky, V.V., 1975: Small shelly fossils of late Precambrian and early Cambrian age: a review of recent work. Journal of the Geological Society 131, 289–304. doi:https://doi.org/10.1144/gsjgs.131.3.0289.
- McLoughlin, S., Vajda, V., Topper, T.P., Liu, F., Johansson, O., & Skovsted, C.B., 2021 [this volume]: New trace fossils and algal remains from the lower Cambrian Torneträsk Formation, northern Sweden. GFF.
- Meert, J.G., 2013: Ediacaran–Early Ordovician paleomagnetism of Baltica: a review. Gondwana Research 25, 159–169. doi:https://doi.org/10.1016/j.gr.2013.02.003.
- Missarzhevsky, V.V., 1977: Konodonty (?) i fosfatnye problematiki kembriya Mongolii i Sibiri. Bespozvonochnye Paleozoya Mongolii, Trudy Sovmestaya Sovetsko-Mongolskaya Paleontologicheskaya Ekspeditsiya 10, 19.
- Moberg, J.C., 1892: Om en nyupptäckt fauna i block af kambrisk sandsten, insamlade av Dr. N.O. Holst. Geologiska Föreningens i Stockholm Förhandlingar 14, 103–120. doi:https://doi.org/10.1080/11035899209444297.
- Moberg, J.C., 1908: Bidrag till kännedomen om de kambriska lagren vid Torneträsk. Sveriges geologiska undersökning C 212 2, 1–30 pp.
- Nielsen, A.T. & Schovsbo, N.H., 2007: Cambrian to basal Ordovician lithostratigraphy in southern Scandinavia. Bulletin of the Geological Society of Denmark 53, 47–92. doi:https://doi.org/10.37570/bgsd-2006-53-04.
- Nielsen, A.T. & Schovsbo, N.H., 2011: The lower Cambrian of Scandinavia: depositional environment, sequence stratigraphy and palaeogeography. Earth-Science Reviews 107, 207–310. doi:https://doi.org/10.1016/j.earscirev.2010.12.004.
- Ormö, J., Nielsen, A.T., & Alwmark, C., 2017: The Vakkejokk Breccia: an early Cambrian proximal impact ejecta layer in the North‐Swedish Caledonides. Meteoritics & Planetary Science 52, 623–645. doi:https://doi.org/10.1111/maps.12816.
- Peel, J.S., Streng, M., Geyer, G., Kouchinsky, A., & Skovsted, C.B., 2016: ‘Ovatoryctocara granulata’ assemblage (Cambrian series 2–series 3 boundary) of Löndal, North Greenland. Australasian Palaeontological Memoirs 49, 1–241.
- Poulsen, C., 1932: The Lower Cambrian faunas of east Greenland. Meddelelser Om Grønland 87, 1–66.
- Poulsen, C., 1942: Nogle hidtil ukendte fossiler fra Bornholms Exsulanskalk. Meddelelser Fra Dansk Geologisk Förening 10, 212–235.
- Poulsen, C., 1967: Fossils from the lower Cambrian of Bornholm. Danske Videnskabernes Selskab, Matematisk-Fysiske Meddelelser 36, 1–48.
- Qian, Y. & Bengtson, S., 1989: Palaeontology and biostratigraphy of the early Cambrian Meishucunian stage in Yunnan province, South China. Fossils and Strata 24, 1–156.
- Regnell, G., 1948: An outline of the succession and migration of non-crinoid pelmatozoan faunas in the lower Paleozoic of Scandinavia. Arkiv För Kemi, Mineralogi Och Geologi 26A, 1–55.
- Roberts, D. & Gee, D.G., 1985: An introduction to the structure of the Scandinavian Caledonides. In: D.G. Gee & B.A. Sturt (eds.): The caledonide orogen – Scandinavia and related areas, 55–68. John Wiley & Sons Ltd, Chichester.
- Rosén, S., 1919: Über einige neue Problematica in einem Fossilfuhrenden kalkstein aus dem nordschwedischen Hochgebirge. Bulletin of the Geological Institute of the University of Uppsala 16, 159–168.
- Rozanov, A.Y., Missarzhevsky, V.V., Volkova, N.A., Voronova, L.G., Krylov, I.N., Keller, B.M., Korolyuk, I.K., Lendzion, K., Michniak, R., Pyhova, N.G., & Sidorov, A.D., 1969: The tommotian stage and the Cambrian lower boundary problem. Transactions of the Academy of Sciences of the USSR Nauka 206, 1–380.
- Rozanov, A.Y., Parkhaev, P.Y., Demidenko, Y.E., Karlova, G.A., Korovnikov, I.V., Shabanov, Y.Y., Ivancov, A.Y., Luchinina, V.A., Malakhovskaya, A.E., Melnikova, L.M., Naimark, E.B., Ponomarenko, A.G., Skorlotova, N.A., Sundukov, V.M., Tokarev, D.A., Ushatinskaya, G.T., & Kipriyanova, L.D., 2010: Fossils from the lower Cambrian stage stratotypes, 225. PIN RAN, Moscow.
- Siveter, D.J. & Williams, M., 1997: Cambrian bradoriid and phosphatocopid arthropods of North America. Special Papers in Palaeontology 57, 1–69.
- Skovsted, C.B., 2006: Small Shelly fauna from the upper lower Cambrian Bastion and Ella Island formations, north-east Greenland. Journal of Paleontology 80, 1087–1112. doi:https://doi.org/10.1666/0022-3360(2006)80[1087:SSFFTU]2.0.CO;2.
- Skovsted, C.B., Brock, G.A., & Paterson, J.R., 2006: Bivalved arthropods from the lower Cambrian mernmerna formation, Arrowie Basin, South Australia and their implications for identification of Cambrian “small shelly fossils”. Memoirs of the Association of Australasian Palaeontologists 32, 7–41.
- Skovsted, C.B., Brock, G.A., Paterson, J.R., Holmer, L.E., & Budd, G.E., 2008: The scleritome of Eccentrotheca from the lower Cambrian of South Australia: lophophorate affinities and implications for tommotiid phylogeny. Geology 36, 171–174. doi:https://doi.org/10.1130/G24385A.1.
- Skovsted, C.B., Clausen, S., Álvaro, J.J., & Ponleve, D., 2014: Tommotiids from the early Cambrian (Series 2, Stage 3) of Morocco and the evolution of the tannuolinid scleritome and setigerous shell structures in stem group brachiopods. Palaeontology 57, 171–192. doi:https://doi.org/10.1111/pala.12060.
- Skovsted, C.B. & Holmer, L.E., 2005: Early Cambrian brachiopods from north‐east Greenland. Palaeontology 48, 325–345. doi:https://doi.org/10.1111/j.1475-4983.2005.00450.x.
- Skovsted, C.B., Knight, I., Balthasar, K.U., & Boyce, W.D., 2017: Depth related brachiopod faunas from the lower Cambrian forteau formation of southern Labrador and western Newfoundland, Canada. Palaeontologia Electronica 20, 54A, 1–52. doi:https://doi.org/10.26879/775.
- Skovsted, C.B. & Peel, J.S., 2001: The problematic fossil Mongolitubulus from the Lower Cambrian of Greenland. Bulletin of the Geological Society of Denmark 48, 135–147.
- Strand, T., 1929: The Cambrian beds of the Mjøsen district in Norway. Norsk Geologisk Tidsskrift 10, 308–365.
- Streng, M., Ebbestad, J.O.R., & Moczydłowska, M., 2008a: A Walcottella -like bradoriid (Arthropoda) from the lower Cambrian of Sweden. GFF 130, 11–19. doi:https://doi.org/10.1080/11035890801301011.
- Streng, M., Holmer, L.E., Popov, L.E., & Budd, G.E., 2008b: Columnar shell structures in early linguloid brachiopods–new data from the middle Cambrian of Sweden. Transactions of the Royal Society of Edinburgh 98, 221–232. doi:https://doi.org/10.1017/S1755691007079820.
- Szaniawski, H., 2002: New evidence for the protoconodont origin of chaetognaths. Acta Palaeontologica Polonica 47, 405–419. http://www.paleo.pan.pl/acta/acta47/app47−405.pdf
- Thelander, T., 1982: The Torneträsk formation of the dividal group, northern Swedish Caledonides. Sveriges geologiska unders#ökning C 789, 1–41 pp.
- Topper, T.P., Skovsted, C.B., Brock, G.A., & Paterson, J.R., 2007: New bradoriids from the lower Cambrian Mernmerna formation, South Australia: systematics, biostratigraphy and biogeography. Memoirs of the Association of Australasian Palaeontologists 33, 67–100.
- Topper, T.P., Skovsted, C.B., Harper, D.A.T., & Ahlberg, P., 2013: A bradoriid and brachiopod dominated shelly fauna from the Furongian (Cambrian) of Västergötland, Sweden. Journal of Paleontology 87, 69–83. doi:https://doi.org/10.1666/12-047R.1.
- Ulrich, E.O. & Bassler, R.S., 1931: Cambrian bivalved Crustacea of the order Conchostraca. Proceedings of the US National Museum 78, 1–130. doi:https://doi.org/10.5479/si.00963801.78-2847.1.
- Ushatinskaya, G.T., 2012: The oldest lingulids of the Siberian platform: microornamentation and shell structure. Paleontological Journal 46, 1298–1308. doi:https://doi.org/10.1134/S003103011212009X.
- Ushatinskaya, G.T. & Korovnikov, I.V., 2014: Revision of the early-middle Cambrian Lingulida (Brachiopoda) from the Siberian platform. Paleontological Journal 48, 26–40. doi:https://doi.org/10.1134/S0031030114010158.
- Ushatinskaya, G.T. & Korovnikov, I.V., 2016: Revision of the Superfamily Acrotheloidea (Brachiopoda, Class Linguliformea, Order Lingulida) from the lower and middle Cambrian of the Siberian platform. Paleontological Journal 50, 450–462. doi:https://doi.org/10.1134/S0031030116050130.
- Vinther, J. & Nielsen, C., 2005: The early Cambrian Halkieria is a mollusc. Zoologica scripta 34, 81–89. doi:https://doi.org/10.1111/j.1463-6409.2005.00177.x.
- Vogt, T., 1967: Fjellkjedestudier i den østlige del av Troms. Norges Geologiske Undersøkelse 248, 1–60. https://hdl.handle.net/11250/2674874
- Walcott, C.D., 1912: Cambrian Brachiopods. U.S. Geological Survey Monograph 60, 1–235.
- Willdén, M.Y., 1980: Paleoenvironment of the autochthonous sedimentary rock sequence at Laisvall, Swedish Caledonides. Stockholm Contributions in Geology 33, 1–100.
- Williams, M. & Siveter, D.J., 1998: British Cambrian and Tremadoc bradoriid and phosphatocopid arthropods. Palaeontographical Society Monograph 152, 1–49.
- Wiman, C., 1903: Studien über das Nordbaltischen Silurgebiet. I. Olenellussandstein, Obolussandstein und Ceratopygescheifer. Bulletin of the Geological Institution of the University of Uppsala 6, 12–76.
- Wotte, T., Skovsted, C.B., Whitehouse, M.J., & Kouchinsky, A., 2019: Isotopic evidence for temperate oceans during the Cambrian explosion. Scientific Reports 9, 6330. doi:https://doi.org/10.1038/s41598-019-42719-4.
- Zhang, X.G., 2007: Phosphatized bradoriids (Arthropoda) from the Cambrian of China. Palaeontographica Abteilung A 281, 93–173. doi:https://doi.org/10.1127/pala/281/2007/93.