
ABSTRACT
Here we investigate megaspores from 10 Triassic‒Jurassic localities of southern Sweden and Bornholm, Denmark, based on collections housed in the Swedish Museum of Natural History. We identify and describe 19 megaspore taxa belonging to three stratigraphically constrained assemblages, representing the Rhaetian, Hettangian and Pliensbachian, respectively. Megaspores are abundant and diverse (12 taxa) in the Rhaetian assemblage. Diversity markedly decreases across the Triassic–Jurassic boundary (TJB), with species richness reducing from 12 to two taxa. The Hettangian assemblage is well-preserved but depauperate, and is overwhelmingly dominated by Nathorstisporites hopliticus. A subsequent recovery of lycopsid diversity followed, recorded by an increase in richness to six taxa in the Pliensbachian assemblage. The disappearance of the hygrophilous and diverse heterosporous lycophyte communities across the TJB, suggests a shift to drier conditions in the earliest Jurassic. This is supported by lithological changes from coal-forming environments in the Rhaetian to sandstone-dominated fluvial-estuarine facies in the Hettangian. Throughout this study, we analysed the megaspores using fluorescence microscopy, which revealed detailed morphological features on specimens that were otherwise opaque under visible light. This non-destructive technique is particularly useful for examining opaque megaspores embedded in permanent mounting media, such as epoxy resin, and may provide new insights into historical megaspore collections elsewhere.
Introduction
The province of Skåne (Scania), southern Sweden, hosts substantial successions of Triassic and Jurassic strata, deposited mainly in continental to paralic settings, offering opportunities to assess vegetation turnover across the Triassic–Jurassic boundary (TJB). Sweden is part of the Baltic plate and was located around 45°N (Torsvik & Cocks Citation2017) during the Early Jurassic. The climate was warm and humid, and the vegetation was abundant and diverse (Vajda & Wigforss-Lange Citation2009). The Triassic–Jurassic successions of Skåne host coal seams, evidence of the once lush vegetation, which are intercalated with clays that have exceptional ceramic properties. During the 1900s, these clays were quarried and used for pottery, bricks and tiles (Ahlberg et al. Citation2003). The quarrying exposed rich fossil plant assemblages from the coal seams and clays spanning the Triassic–Jurassic transition (see Linné Citation1751; Hermelin Citation1773; Nathorst Citation1875, Citation1876, Citation1878a, Citation1878b, Citation1878c, Citation1879a, Citation1879b, Citation1886, Citation1902, Citation1906a, Citation1906b, Citation1906c, Citation1907, Citation1908a, Citation1908b, Citation1908c, Citation1908d, Citation1909a, Citation1909b, Citation1911, Citation1912; Lundblad Citation1948, Citation1949a, Citation1949b, Citation1950a, Citation1950b, Citation1954, Citation1958; Pott & McLoughlin Citation2009, Citation2011; Slodownik et al. Citation2021). Much of this fossil material is now housed at the Swedish Museum of Natural History (NRM). Stratigraphic studies of the Triassic and Jurassic successions of Skåne have to some extent relied on these “macro” fossil plants, but numerous palynological studies have also been conducted, revealing distinct pollen and spore assemblages that have proven extremely useful for the biostratigraphic zonation of this region (Tralau Citation1975; Lund Citation1977; Guy-Ohlson Citation1981, Citation1982, Citation1986, Citation1990; Guy-Ohlson & Malmquist Citation1985; Guy-Ohlson & Norling Citation1988, Citation1994; Vajda Citation2001; Lindström & Erlström Citation2006; Larsson Citation2009; Petersen et al. Citation2013; Vajda et al. Citation2013, Citation2016; Lindström et al. Citation2017). Megaspores also occur within these successions, but have so far been little studied compared to the macrofossil and miospore assemblages. Megaspores are large spores produced by heterosporous plants, and during the early Mesozoic, lycophytes (Pleuromeiales, Isoetales, Selaginellales) were the only producers of megaspores (Kovach & Batten Citation1989). Megaspore-producing water-ferns appeared during the latest Jurassic and became more abundant throughout the Cretaceous to Cenozoic (Collinson et al. Citation1985, Citation2010, Citation2013; Vajda & McLoughlin Citation2005; Cúneo et al. Citation2013). Heterosporous lycophytes generally lived on floodplains, commonly as pioneer plants following ecosystem disturbance locally, or globally (e.g. Retallack Citation1997; Looy et al. Citation1999; McLoughlin et al. Citation2014; Vajda et al. Citation2020). Megaspores generally preserve well in the fossil record due to their resistant and typically thick sporopollenin walls, and have proven valuable for biostratigraphy (e.g. Marcinkiewicz Citation1971; Kovach & Batten Citation1989; Morris et al. Citation2009; Neri et al. Citation2018; Shevchuk et al. Citation2020; Li et al. Citationin press), particularly throughout episodes of rapid biotic turnover (e.g. Vajda & Raine Citation2003; Vajda & McLoughlin Citation2005).
Prior to this investigation, two taxa, Lycostrobus scottii (=Nathorstisporites hopliticus) and Trileites pinguis (=Banksisporites pinguis), have been formally described from Triassic–Jurassic successions of Sweden. Nathorst (Citation1902) described Androstrobus scotti, from Lower Jurassic deposits, which he interpreted as a male cycad cone. Upon re-examination, Nathorst (Citation1908a) found in situ megaspores, indicating this was a lycophyte strobilus, and so re-named it, Lycostrobus scottii. The megaspores were re-described by Lundblad (Citation1956), together with L. scottii megaspores from various drillcores from Skåne. The other megaspores documented from the Triassic of Skåne were specimens of Trileites pinguis (Lundblad Citation1959). Further taxa were later recorded from coeval deposits from Bornholm, Denmark, documented by Gry (Citation1969) and Koppelhus & Batten (Citation1992). More recently, an unidentified megaspore was documented from a thin section of a permineralized fern rhizome from the Pliensbachian of Korsaröd, Skåne (McLoughlin & Bomfleur Citation2016).
Here we aim to document and describe Triassic and Jurassic megaspores housed in the collections of the NRM. We assess the biostratigraphic relevance of these megaspores and apply UV fluorescence microscopy for morphological analysis of dispersed megaspores.
Geological setting
During the Triassic and Jurassic, basin formation in Scandinavia was controlled by continental rifting along the Sorgenfrei-Tornquist Zone. This is a continuous zone of deformation stretching from southeastern Europe to the central North Sea (Sorgenfrei & Buch Citation1964), and is a part of the Fennoscandian Border Zone, constituting a prominent component of the landscape in Skåne (Bergström Citation1984; Norling & Bergström Citation1987; Larsson et al. Citation2000). Swedish Upper Triassic and Lower Jurassic sedimentary successions are restricted to the southernmost province, Skåne. These successions were deposited in alluvial, deltaic, coastal plain and shallow marine shelf environments in several sedimentary basins along the Fennoscandian Border Zone (; Ahlberg et al. Citation2003; Lindström et al. Citation2017).
Figure 1. Geological map of Skåne (southern Sweden) with studied localities, modified after Kruger et al. (Citation2021, this issue). 1. Åbro BH-207/64 and Billesholm. 2. Hyllinge. 3. Textile Factory, Eslöv. 4. Höllviken II. 5. Stabbarp BH-74. 6. Helsingborg harbour. 7. Svedala I. 8. Kävlinge BH-928
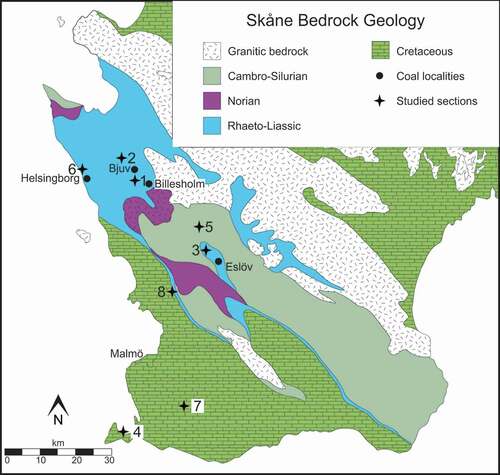
In northwestern Skåne, uppermost Triassic (Rhaetian) to Lower Jurassic successions are represented by the Höganäs and Rya formations (). The Rhaetian–Hettangian Höganäs Formation incorporates three subdivisions: the Vallåkra, Bjuv and Helsingborg members (in stratigraphic order). The TJB lies within the Höganäs Formation, approximately at the transition between the Bjuv and Helsingborg members, and the facies spanning this interval comprise plant-bearing mudstones, siltstones, sandstones and coals, characteristic of continental and deltaic environments. The overlying Rya Formation (Sinemurian‒lower Aalenian) is subdivided into four subunits: the Döshult, Pankarp, Katslösa and Rydebäck members (Troedsson Citation1951; Norling Citation1968; Pieńkowski Citation1991; Vajda & Wigforss-Lange Citation2009; Lindström et al. Citation2017). A major part of the Lower Jurassic succession of NW Skåne (the three upper members of the Rya Formation) is marine in origin (Kruger et al. Citation2021, this issue), represented by sandstones, siltstone and ferruginous oolitic sandstones in the topmost Rydebäck Member. These successions yield diverse marine fossil assemblages, including bivalves, ammonites, ostracods and foraminifera, but also contain rich spore and pollen assemblages (Norling et al. Citation1993 and references therein).
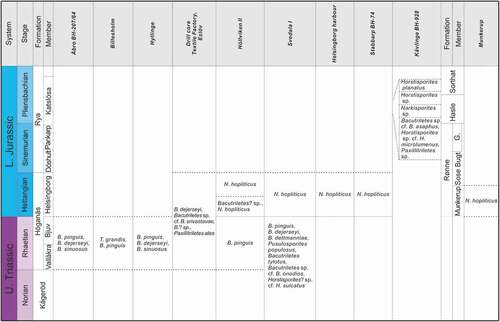
Figure 2. Early Mesozoic megaspore assemblages recorded in this study from Skåne (Sweden) and Bornholm (Denmark). G. Galgeløkke Member
The Triassic to basal Jurassic successions of Bornholm are represented by the Kågeröd (Norian) and Rønne (Hettangian‒Sinemurian) formations, which are separated by an unconformity (Gravesen et al. Citation1982; ). The Rønne Formation is subdivided into the Munkerup, Sose Bugt and Galgeløkken members (Gravesen et al. Citation1982; Michelsen et al. Citation2003). The base of the lowermost unit, the Munkerup Member, marks the base of the Jurassic, and this member comprises dark, lacustrine clays intercalated with fine sandstone beds (Gravesen et al. Citation1982; Koppelhus Citation1991; Vajda & Wigforss-Lange Citation2009).
Material
Megaspores studied here are from historical collections of the Department of Palaeobiology, NRM. The megaspores include specimens mounted in Canada balsam on glass slides, and unmounted specimens, stored in microfossil sorting trays (cardboard slides with glass covers). A portion of the material examined was collected by A.G. Nathorst and B. Lundblad from various localities across Skåne and Bornholm, and some specimens have been published (Nathorst Citation1902, Citation1908a; Lundblad Citation1950a, Citation1950b, Citation1956). Previously unstudied material is also investigated here from the Kävlinge drillcore (BH-928, the Norling foraminifera collection), the Höllviken II drillcore (the Brotzen foraminifera collection), and the Munkerup beds of Bornholm (collected by B. Lundblad). Details of each locality are provided below.
Åbro BH-207/64, 46.39–46.37 m (Rhaetian)
Megaspores from one sample from the drillcore Åbro BH-207/64, located ca. 5 km southeast of Bjuv, Skåne (), were re-studied (slide numbers: S054802-01 to S054802-25). The sample originates from interval 46.39–46.37 m, from a coal seam of Rhaetian age (Lundblad Citation1950a, Citation1950b).
Billesholm (Rhaetian)l
Megaspores were examined from three samples from the coal mine of Billesholm. Megaspores were stored as unmounted (two samples, slide numbers: S054353-03, S054719-02-01), and mounted specimens (one sample, slide number: S054798). Plant macrofossils from the Rhaetian coals of the Billesholm mines, associated with these megaspore collections, were studied previously by Halle (Citation1921) and Lundblad (Citation1950a).
Hyllinge, lower coal bed (Rhaetian)l
Megaspores from Hyllinge (slide numbers: S064827-02, S064850-02, S064865-01), originate from outcrop samples stratigraphically below the lower coal bed (Rhaetian), from almost black, laminated mudstones corresponding to Layer 1 of Nathorst (Citation1878a). The sampled bed has been dated previously as Rhaetian based on the macroflora (Nathorst Citation1878a; Harris Citation1937; Lundblad Citation1950b).
Drillcore “Textile factory”, Eslöv, 209.8 m (Rhaetian)l
Megaspores were re-examined from one sample taken at 209.8 m depth from a drillcore of the “Textile Factory”, north of Eslöv (). Four slides, prepared by B. Lundblad (slide numbers: S063488, S063489, S063492, S063493), were studied here. This sample has been dated to the Rhaetian Lepidopteris ottonis Zone (Harris Citation1937).
Drillcore Höllviken II, 1480.78–1477.85 m (Rhaetian), 1433.81 m, 1428.08–1422.08 m (Jurassic)l
Megaspores were examined from three intervals of the Höllviken II drillcore: 1428.08–1422.08 m (slide numbers: S066554, S066555), 1433.81 m (slide numbers: S066549, S066556–S066558, S066560–S066563, S066565, S066566), and 1480.78–1477.85 m (slide numbers: S066586, S066587). The intervals have been dated as Rhaetian (1480.78–1477.85 m) and Jurassic (1433.81 m, 1428.08–1422.08 m), based on foraminifera, plant cuticle and megaspores (Brotzen Citation1950; Lundblad Citation1956).
Drillcore Svedala I, 1570–1560 m (Rhaetian), 1531 m (Hettangian)l
Svedala I borehole is located at the western margin of the Skurup Platform () and was drilled by the Geological Survey of Sweden (SGU) between 1948 and 1950 (Norling Citation1981; Vajda Citation2001). The core intersects Upper Triassic, Jurassic sediments and Cretaceous poorly consolidated glauconitic sandstones (Ahlberg & Olsson Citation2001; Vajda Citation2001). Megaspores were re-studied from samples from depths of 1570–1560 m (slide numbers: S079701, S079702, S079713–S079716, S079718 to S079728) and 1531 m (slide numbers: S079741 to S079743). Interval 1577.75–1538.37 m has been dated as Rhaetian, based on palynology (Ahlberg et al. Citation2002). Level 1531 m has been dated as Hettangian based on its lithology and fossil content (Lundblad Citation1956, Citation1959).
Helsingborg Harbour (Hettangian)l
Megaspores mounted on glass slides embedded in Canada balsam by A.G. Nathorst (slide numbers: S064343, S064347, S064348, S064744, S064747) were studied. These derive from an outcrop section discovered during an expansion of Helsingborg Harbour in the late 1800s and dated to Hettangian (Lundblad Citation1956).
Drillcore Stabbarp BH-74, 5 m (Hettangian)l
The Stabbarp borehole, No. 74 is situated immediately north of the old coal mine of Stabbarp, near to the town of Eslöv (). The sample containing the studied megaspore assemblage (slide number: S075612) is from a thin coal bed from a depth of ca. 5 m in the drillcore. These strata are also referred to as the “Gamla övre gruvan” (The old upper mine), which comprise the coal beds “John Ericsson” and “Jean Molin” and are Hettangian in age (Lundblad Citation1956).
Drillcore Kävlinge BH-928, 46.91 – 33.75 m (Pliensbachian)l
Megaspores recovered from four levels: 46.91–46.07 m, 42.25–41.15 m, 41.15–40.40 m and 34.75–33.75 m were studied here. The core was recovered west of Kävlinge village () and spans the interval 83.25–28.75 m. The lithology consists of grey-green sand and sandstones, rich in oolitic chamosite (Norling Citation1968). The lower portion, between 83.25 m and 54 m, contains a moderately rich foraminiferal assemblage dated to Liassic γ (lower Pliensbachian), which is devoid of megaspores; upper Liassic γ (upper lower Pliensbachian) continental strata occur above 54 m (Norling Citation1968).
Munkerup, Bornholm, Denmark (Hettangian)l
Megaspores were examined from one sample from the Munkerup Member from coastal outcrops in southwestern Bornholm, Denmark. The material was accessioned into the collections of the NRM in 1961, possibly by B. Lundblad. The clay beds of Munkerup host abundant plant fossils characteristic of the Early Jurassic Thaumatopteris flora (Harris Citation1937), and the succession has been dated as early Hettangian based on these macrofossils and palynology (Koppelhus Citation1991).
Methods
Reflected light microscopy
Unmounted specimens were initially investigated using an Olympus SZX10 reflected light microscope at ×6.3 to ×63 magnifications. Due to the relatively low level of magnification of reflected light microscopy, selected specimens were subsequently examined using transmitted light, UV light and scanning electron microscopy.
Transmitted light microscopyl
Specimens that were mounted on glass slides and embedded in Canada balsam (hampering SEM investigation) were examined using an Olympus BX51 transmitted light microscope, and photographed with an Olympus DP71 camera. Objective lenses of ×5 and ×10 were used to locate specimens, and ×20 was used for imaging. Where possible, slides were inverted and examined from both sides to assess proximal and distal surfaces of the megaspores.
Fluorescence microscopyl
Megaspores that were mounted in Canada balsam were studied and imaged with UV light using an Olympus BX51 microscope and photographed using an Olympus DP71 camera. Various filters were used to examine megaspores under UV light, among which the WIB (blue filter) was optimal for imaging the studied material.
Scanning electron microscopy (SEM)l
Megaspores from the drillcore Kävlinge BH-928, Billesholm and Munkerup (Bornholm) () were examined using scanning electron microscopy. Thirteen specimens were mounted onto SEM stubs (with carbon tape) and gold coated for 20–50 seconds using an Agar high-resolution sputter coater. Specimens were studied using an ESEM FEI Quanta FEG 650 scanning electron microscope at the NRM.
Systematic palaeontology
Nineteen species belonging to eight genera were identified and described. Several taxa are presented in open nomenclature due to insufficient specimens, poor preservation or a lack of SEM observations for specimens sealed in glass slides. We follow the descriptive terminology of Punt et al. (Citation2007).
Genus Trileites Erdtman, Citation1947 ex Potonié, Citation1956
Trileites grandis Fuglewicz, Citation1973()
Description
Trilete megaspore, rounded amb. Laesurae straight, narrow, ca. 25 μm wide, ca. 200 μm long, labra developed, not tapering to the top, ca. 25 μm wide. Curvaturae absent. Spore surface smooth.
Dimensionsl
Equatorial diameter: 725 (738) 750 μm (n = 2).
Occurrencel
Rhaetian; Billesholm (S054353-03, S054719-02-01).
Remarksl
The two specimens studied here are indistinguishable from Trileites grandis described by Fuglewicz (Citation1973). Trileites grandis differs from: T. levis Fuglewicz, Citation1973 (230–400 μm) in its larger size; from T. murrayi (Harris, Citation1961) Marcinkiewicz, Citation1971 in lacking laesurae with marginal grooves; from T. crassitectatus Fuglewicz, Citation1977 in lacking strongly developed laesurae (in the form of thick and rounded bands); from T. validus Fuglewicz, Citation1973 in lacking long (=spore radius) laesurae and slightly raised spore margins; from T. vulgaris Fuglewicz (Citation1973) and T. flexuosus Fuglewicz, Citation1977 in lacking concave contact areas and/or convex spore sides; and from T. polonicus Fuglewicz, Citation1973, T. tenellus Fuglewicz, Citation1973 and T. robustus Fuglewicz, Citation1980 in lacking curvaturae.
Genus Banksisporites Dettmann, Citation1961
Remarksl
Two separate layers of the exine were first recognized as a taxonomically significant character for dispersed fossil megaspores by Høeg et al. (Citation1955), and a new genus, Duosporites was erected. Likewise, Dettmann (Citation1961) established Banksisporites based on the presence of a cavate exine; this consists of an intexine (homogeneous or composed of granules) and an exoexine (smooth or granular), which are fused near the laesurae on the proximal surface. However, Potonié (Citation1966) rejected Banksisporites because the detachment of two exine layers was difficult to observe on the type species originally described by Harris (Citation1935). Fuglewicz (Citation1973, p. 419) also disagreed with the erection of Banksisporites and transferred B. sinuosus to Trileites. Batten and Kovach (Citation1990) recorded the transfer of Fuglewicz (Citation1973), but stated that the type species B. pinguis must remain unless this genus is regarded as a junior synonym of Trileites. Glasspool (Citation2000) considered the presence of an inner body as a character of limited taxonomic importance, and did not incorporate this feature into the emended diagnosis of Singhisporites. Glasspool (Citation2003) later emended the diagnosis of Banksisporites, using surface ornamentation as a taxonomic character, which differs from the original description by Dettmann (Citation1961).
Banksisporites pinguis (Harris, Citation1935) Dettmann, Citation1961()
Descriptionl
Trilete megaspore, always with a rounded-triangular or circular amb. Laesurae straight or slightly undulate, rarely reaching the equator, labra strongly developed, up to 55 μm high, tapering gradually in width. Curvaturae absent. Exine two-layered, cavate, adherent in proximal area around the laesurae (). Surface smooth, rarely folded.
Dimensionsl
Equatorial diameter: 248 (472) 993 μm (n = 230).
Occurrencel
Rhaetian; Åbro BH-207/64 (S054802-01–S054802-25); Billesholm (S054798); Höllviken II, 1480.78–1477.85 m (S066586, S066587); Hyllinge (S064827-02, S064850-02, S064865-01); Svedala I, 1570–1560 m (S079701, S079715, S079716, S079720, S079721, S079728).
Remarksl
The studied specimens are consistent with the type species, except that 35 of the 230 grains examined here, are larger than those described by Dettmann (Citation1961), which range between 210 and 530 μm in equatorial diameter.
Banksisporites dejerseyi Scott et Playford, Citation1985()
Descriptionl
Trilete megaspore, amb generally circular to subcircular, rarely rounded triangular. Laesurae straight to slightly sinuous, almost reaching the equator, labra strongly raised, or in a few cases weakly developed, tapering gradually in width. Curvaturae absent. Exine two-layered, cavate, adherent around the laesurae on the proximal face. Surface smooth as viewed with fluorescence, showing prominent folds mainly peripherally, and around the labra.
Dimensionsl
Equatorial diameter: 290 (468) 662 μm (n = 25).
Occurrencel
Rhaetian; Åbro BH-207/64 (S054802-03–S054802-06, S054802-12, S054802-16, S054802-18–S054802-20); Hyllinge (S064850-02); Drillcore Textile Factory, Eslöv, 209.8 m (S063492); Svedala I, 1570–1560 m (S079701, S079719, S079721–S079723, S079728).
Remarksl
The Skåne specimens are similar to those described by Scott & Playford (Citation1985) in possessing prominent peripheral folds.
Banksisporites dettmanniae Banerji, Kumaran et Maheshwari, Citation1978()
Descriptionl
Trilete megaspore, circular in outline. Laesurae straight, never extending to the equator, labra uniformly high with flat crests. Curvaturae distinct, perfectly developed, delimiting the contact area. Spore surface smooth, strongly folded on the distal face. Exine two-layered, inner layer indistinct on the single specimen examined.
Dimensionsl
Equatorial diameter: 662 μm (n = 1).
Occurrencel
Rhaetian; Svedala I, 1570–1560 m (S079701).
Remarksl
This specimen is similar to the type material of Banksisporites dettmanniae in having perfectly developed curvaturae near the equator, but the inner layer of the studied specimen is less distinct compared to those described by Banerji et al. (Citation1978).
Banksisporites sinuosus Dettmann, Citation1961()
Descriptionl
Trilete megaspore with rounded triangular amb. Laesurae strongly sinuous near the proximal pole, almost reaching the equator, labra up to 30 μm high and 25 μm wide. Curvaturae absent. Exine two-layered, cavate. Surface smooth.
Dimensionsl
Equatorial diameter: 256 (328) 453 μm (n = 17).
Occurrencel
Rhaetian; Åbro BH-207/64 (S054802-01, S054802-08, S054802-12, S054802-15, S054802-17–S054802-20); Hyllinge (S064827-02).
Remarksl
Spores attributed to this species from Skåne have distinctly sinuous laesurae consistent with Banksisporites sinuosus specimens described from the Upper Triassic of Australia by Dettmann (Citation1961). However, the Swedish examples differ in having longer laesurae that almost reach the equator.
Fuglewicz (Citation1973) transferred Banksisporites sinuosus to Trileites, suggesting the “inner body structure” was not considered a valid taxonomic feature for the erection of Banksisporites. However, Fuglewicz (Citation1973) provided only SEM images of the external spore morphology, and transmitted light micrographs that may have revealed the inner body structure were lacking. The Polish specimens described by Fuglewicz (Citation1973) are also markedly older (Early Triassic) than the Australian type material (Dettmann Citation1961). Thus morphological variation regarding the presence or absence of an inner body among these assemblages would not be particularly surprising given that they are separated by considerable time and distance. Here we retain the use of Banksisporites, as the inner body may prove to be a useful distinguishing feature among morphologically similar megaspore taxa in future studies, and may indeed shed light on potential producers.
Genus Pusulosporites Fuglewicz, Citation1973Pusulosporites populosus Fuglewicz, Citation1973
Descriptionl
Trilete megaspore, amb circular to subtriangular. Laesurae straight or slightly undulate, generally spanning half the spore radius, labra developed, ca. 15 μm wide, up to ca. 35 μm high, jagged towards the margins. Curvature absent. Spore surface covered with dense short gemmae, verrucae, coni, spinae, bacula with sharp, rounded or blunt apices. These sculptural elements are in some cases united into rugulae.
Dimensionsl
Equatorial diameter: 319 (533) 618 μm (n = 4).
Occurrencel
Rhaetian; Svedala I, 1570–1560 m (S079716, S079726, S079727).
Remarksl
The ornamentation of our specimens is consistent with examples of Pusulosporites populosus described by Fuglewicz (Citation1973, p. 425, pl. XXXII, ): “numerous, short appendages ended up by sharp or rounded apices” as determined under transmitted light. In some cases, the rugulae united by various sculptural elements are comparable with the type of ornamentation on Hughesisporites australis (McLoughlin et al. Citation2002, fig. 66A–F) and Verrutriletes sp. A (McLoughlin et al. Citation2002, fig. 73C–D); however, the megaspores of this study are dominantly covered with separated sculptural elements (), among which coni and spinae are common, but are absent on Hughesisporites australis and Verrutriletes sp. A.
Genus Bacutriletes van der Hammen, Citation1954 ex Potonié, Citation1956 emend. Banerji, Jana et Maheshwari, Citation1984
Remarksl
Banerji et al. (Citation1984) emended the generic diagnosis to encompass species with rounded bacula, and those with bacula confined to one face. However, Potonié (Citation1956) already included those species with rounded bacula in the generic diagnosis to read: “ … Exine covered on all sides with truncated or rounded bacula … ” (translated from German, Potonié Citation1956, p. 35)
Bacutriletes tylotus (Harris, Citation1935) Potonié, Citation1956
Descriptionl
Trilete megaspore, more or less globular. Laesurae straight and narrow, usually spanning half the radius of the spore, never reaching the equator, labra weakly developed, up to 30 μm high on some specimens. Curvature absent. Entire spore surface covered by cylindrical bacula typically up to 50 μm long, and 30 μm wide at the base. Bacula smaller towards the proximal pole, bearing transversely truncate ends. Bacula straight or in a few cases bent, unbranched, generally occurring in groups of two or three, where they may be fused at their apices. Smaller bacula or grana weakly developed between larger sculptural elements.
Dimensionsl
Equatorial diameter: 442 (568) 682 μm (n = 46).
Occurrencel
Rhaetian; Svedala I, 1570–1560 m (S079701, S079702, S079713–S079715, S079718, S079720, S079723).
Remarksl
Our specimens are similar to those described by Harris (Citation1935) in bearing diagnostic apically truncated and usually clustered bacula. Subordinate sculptural elements between bacula were faintly visible on specimens mounted in Canada balsam (, D, F). The type of clustering and fusion of sculptural elements is similar to the ribbon-like architecture of Singhisporites hystrix (Slater et al. Citation2011, pl. IV); however, the specimens studied here lack the multi-branched and reticulate flared sculptural elements of S. hystrix, which are also relatively uniform compared to the more varied sculptural elements of Bacutriletes tylotus.
Bacutriletes sp. cf. B. asaphus Fuglewicz, Citation1973(–D)
Descriptionl
Trilete megaspore, amb circular. Laesurae straight, labra well developed, narrow, generally tapering to the top. Curvaturae distinct as low ridges, narrow, imperfect. Contact area covered with more or less evenly distributed bacula, usually flattened at their apices and broadened and united basally to form a delicate reticulum.
Dimensionsl
Equatorial diameter: 369 μm (n = 1).
Occurrencel
Pliensbachian; Kävlinge BH-928, 46.91–46.07 m.
Remarksl
The single specimen recorded was heavily corroded so that only one interradial area possessed ornamentation. The more or less reticulate pattern formed by the short baculate appendages is similar to the description of Bacutriltes asaphus by Fuglewicz (Citation1973). Specimens described as Bacutriltes reticuliferus by Bertelsen & Michelsen (Citation1970) differs from Bacutriletes sp. cf. B. asaphus in having shorter laesurae, lacking curvaturae, and bearing densely spaced blunt bacula over the whole spore surface.
Bacutriletes sp. cf. B. srivastavae Banerji , Jana et Maheshwari, Citation1984 ()
Descriptionl
Trilete megaspore, amb circular? (only one specimen preserved in lateral orientation), laesurae inconspicuous. Curvaturae indistinct. Spore surface covered by closely distributed but discrete, cylindrical, not tapering, bacula, rounded apically.
Dimensionsl
Equatorial diameter: 368 μm (n = 1).
Occurrencel
Rhaetian; Drillcore Textile Factory, Eslöv, 209.8 m (S063489).
RemarksThis specimen is comparable with that of Bacutriletes srivastavae in bearing dense bacula that are not confluent at the base; and the configuration of the bacula on the spore outline under transmitted light () is identical to that of B. srivastavae (Banerji et al. Citation1984, pl. 4, ). However, on the studied specimen, bacula are smaller on the proximal face compared to the Indian specimens.
Bacutriletes sp. cf. B. onodios (Harris, Citation1961) Hopkins et Sweet, Citation1976()
Descriptionl
Trilete megaspore, amb circular. Laesurae conspicuous, short, extending to half of the spore radius. Curvaturae absent. Varied coni, gemmae and short clavae present on contact area. Remainder of spore surface densely but irregularly covered with baculate appendages, up to 85 μm long and ca. 25 μm wide, cylindrical or slightly tapering to rounded or blunt apices. Bacula usually straight, or slightly bent near the apices, in some cases fused at the base and bifurcated at the top.
Dimensionsl
Equatorial diameter: 549 μm (n = 1).
Occurrencel
Rhaetian; Svedala I, 1570–1560 m (S079718).
Remarksl
The long bacula in the studied specimen are similar to those of Bacutriletes arnoldii (Miner, Citation1932) Potonié, Citation1956, B. clavatus Marcinkiewicz, Citation1960, B. corynactis (Harris, Citation1961) Marcinkiewicz, Citation1971, B. spicatus (Marcinkiewicz, Citation1962) Marcinkiewicz, Citation1971, B. onodios (Harris, Citation1961) Hopkins et Sweet, Citation1976 and B. corynactiformis Fuglewicz, Citation1977. However, Bacutriletes arnoldii differs from the studied specimen in having vermiculate bacula; B. clavatus has clavae that are narrow at the base but widen at the top; B. corynactis and B. spicatus have baculate appendages that are usually curved, or even recurved; and B. corynactiformis has smooth contact areas and clavate appendages on the distal surface.
The studied specimen is very similar to Bacutriletes onodios with respect to its straight and almost cylindrical bacula, however, the labra on our specimen are less prominent than those described by Harris (Citation1961), which are 40‒70 μm high.
Bacutriletes? sp.()
Descriptionl
Trilete megaspore, amb subtriangular, laesurae distinct, straight or strongly curved near the proximal pole, labra well developed, with small gemmae and bacula. Curvaturae faintly conspicuous or lacking. Spore surface covered with numerous, densely distributed coni, verrucae, bacula, clavae and/or spinae, and a few short fimbriae. Sculpture more prominent on the distal face. Appendages usually curved or bent backwards, rarely fused at the base.
Dimensionsl
Equatorial diameter: 413 (546) 678 μm (n = 2).
Occurrencel
Hettangian; Höllviken II, 1433.81 m (S066549). Rhaetian; Drillcore Textile Factory, Eslöv, 209.8 m (S063493).
Remarksl
These specimens are characterized by their variable and elongate surface ornamentation. We tentatively identify these specimens as Bacutriletes? sp. because the bacula are more developed than other appendages. The contact area is faintly delineated by denser ornament (), suggesting that these specimens could alternatively be assigned to Narkisporites Kannegieser et Kozur, Citation1972; however, the sculptured curvaturae, which are typical of Narkisporites, are absent on one of the two specimens here assigned to Bacutriletes? sp. ().
Genus Narkisporites Kannegieser et Kozur, Citation1972
Narkisporites sp.()
Descriptionl
Trilete megaspores, amb circular. Laesurae distinct. Labra straight and raised, up to 175 μm long and 65 μm high, rounded at the top, extended to curvaturae. Curvaturae conspicuous, bearing various sizes of coni, spinae and blunt bacula. Surface of contact area smooth, slightly swollen in the centre. Distal surface covered with evenly distributed bacula, slightly tapering to the top, blunt at the apices, not forming a reticulum.
Dimensionsl
Equatorial diameter: 338 μm (n = 1).
Occurrencel
Pliensbachian; Kävlinge BH-928, 42.25–41.15 m.
Remarksl
This specimen was attributed to Narkisporites Kannegieser et Kozur, Citation1972 based on the presence of a distinct curvaturae sculptured with coni, spinae and bacula, although sculptures on the curvaturae are heavily corroded, making comparison with other material difficult.
Genus Horstisporites Potonié, Citation1956
Horstisporites planatus (Marcinkiewicz, Citation1960) Marcinkiewicz, Citation1971()
Descriptionl
Trilete megaspore, amb circular. Laesurae straight or slightly curved, narrow, almost extending to the equator, labra tapering to the top. Curvaturae developed perfectly, consisting of low ridges. Spore surface covered with reticula, lumina irregularly polygonal, ca. 5–20 μm, muri low and narrow, developed on both the proximal and distal surfaces. Finer reticula are present within the coarse reticula, representing the construction of the exoexine ().
Dimensionsl
Equatorial diameter: 340 (345) 349 μm (n = 2).
Occurrencel
Pliensbachian; Kävlinge BH-928, 34.75–33.75 m.
Remarksl
The two specimens of Horstisporites planatus recorded here are consistent with those described by Marcinkiewicz (Citation1971, pl. XII, figs. 1–9) from Poland. These specimens all have a thin exine, which allows the observation of reticulum under transmitted light; this feature was not visible under reflected light. The presence of a similar delicate reticulum described by Marcinkiewicz (Citation1960, Citation1971; areolae diameters of 4–30 μm) and clear curvaturae further support our identification.
Figure 3. Stratigraphical occurrences of Triassic and Jurassic megaspores recorded in Skåne and Bornholm
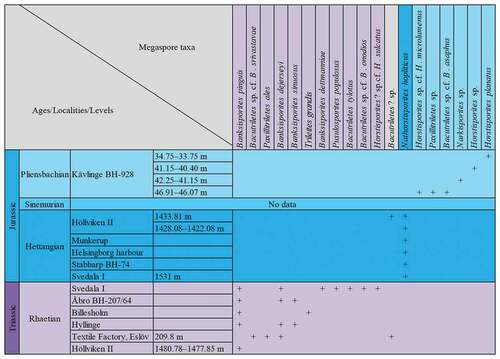
Figure 4. Trileites and Banksisporites. Scale bars = 100 μm. A, B. Trileites grandis Fuglewicz, Citation1973. A. Proximal view, SEM, S054719-02-01. B. Proximal view, SEM, S054353-03. C–F. Banksisporites dettmanniae Banerji, Kumaran et Maheshwari Citation1978. C. Proximal view, transmitted light, S079701, R36. D. Proximal view, transmitted light, overexposure, S079701, R36. E. Proximal view, fluorescence, S079701, R36. F. Distal view, fluorescence, S079701, R36
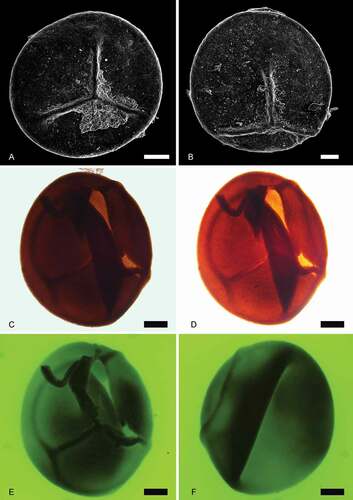
Figure 5. Banksisporites. Scale bars = 100 μm. A–F. Banksisporites pinguis (Harris, Citation1935) Dettmann, Citation1961. A. Proximal view, transmitted light, S054802-01-03, R40-2. B. Proximal view, fluorescence, S054802-01-03, R40-2. C. Megaspore tetrad, transmitted light, S064827-02-01, F31-2. D. Megaspore tetrad, fluorescence, S064827-02-01, F31-2. E. Lateral view, transmitted light, S054802-01-04, P38-3. F. Lateral view, fluorescence, S054802-01-04, P38-3. G–H. Banksisporites sinuosus Dettmann, Citation1961. G. Proximal view, transmitted light, S054802-05-01, R37. H. Proximal view, fluorescence, S054802-05-01, R37. I–L. Banksisporites dejerseyi Scott et Playford, Citation1985. I. Oblique view, transmitted light, S064850-02-01, P29-1. J. Oblique view, fluorescence, S064850-02-01, P29-1. K. Proximal view, transmitted light, S054802-05-01, R37. L. Proximal view, fluorescence, S054802-05-01, R37
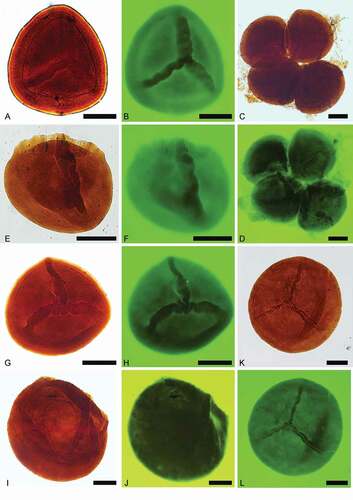
Figure 6. Pusulosporites populosus Fuglewicz, Citation1973. Scale bars = 100 μm. A. Distal view, transmitted light, S079726, O53-2. B. Distal view, fluorescence, S079726, O53-2. C. Proximal view, transmitted light, S079727, M43. D. Proximal view, fluorescence, S079727, M43. E. Proximal view, transmitted light, S079716, L37. F. Proximal view, fluorescence, S079716, L37
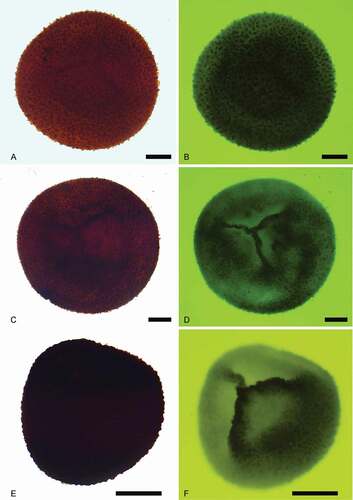
Figure 7. Bacutriletes tylotus (Harris, Citation1935) Potonié, Citation1956. Scale bars = 100 μm. A. Proximal view, transmitted light, S079701, R37-1. B. Proximal view, fluorescence, S079701, R37-1. C. Lateral view, transmitted light, S079701, V38. D. Lateral view, fluorescence, S079701, V38. E. Distal view, transmitted light, S079701, J37-2. F. Distal view, fluorescence, S079701, J37-2
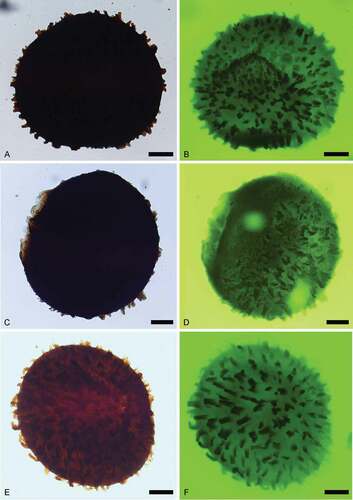
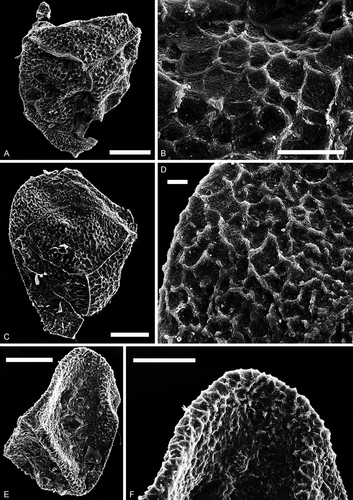
Figure 8. Narkisporites, Bacutriletes and Horstisporites. Scale bars = 100 μm, or indicated otherwise. A, B. Narkisporites sp. A. Lateral view, SEM, Kävlinge BH-928, 42.25–41.15 m. B. Enlargement, SEM, showing blunt and spiny projections outside the contact area, Kävlinge BH-928 42.25–41.15 m. Scale bar = 50 μm. C, D. Bacutriletes sp. cf. B. asaphus Fuglewicz, Citation1973. C. Proximal view, SEM, Kävlinge BH-928, 46.91–46.07 m. D. Enlargement, SEM, Kävlinge BH-928, 46.91–46.07 m. Scale bar = 50 μm. E, F. Horstisporites sp. cf. H. microlumenus Dettmann, Citation1961. E. Proximal view, SEM, Kävlinge BH-928, 46.91–46.07 m. F. Enlargement, SEM, Kävlinge BH-928, 46.91–46.07 m. Scale bar = 50 μm
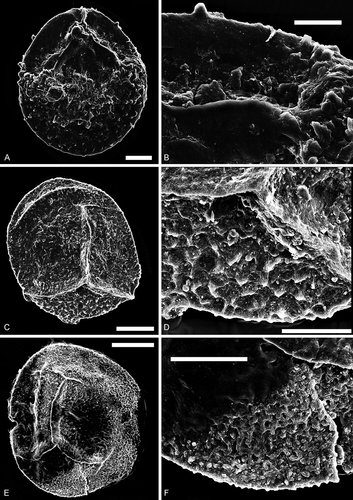
Figure 9. Bacutriletes. Scale bars = 100 μm. A–F, I, L. Bacutriletes? sp. A. Oblique view, transmitted light, S066549, R26. B. Oblique view, transmitted light, over-exposure, S066549, R26. C. Oblique view, fluorescence, S066549, R26. D. Proximal view, transmitted light, S063493-02, N42-4. E. Proximal view, fluorescence, S063493-02, N42-4. F. Enlargement of C, fluorescence, S066549, R26. I. Enlargement of C, fluorescence, S066549, R26. L. Distal view of a broken specimen, fluorescence, S063493-01, N42-4. G, H. Bacutriletes sp. cf. B. srivastavae Banerji, Jana et Maheshwari Citation1984. G. Lateral view, transmitted light, S063489, M44-4. H. Lateral view, fluorescence, S063489, M44-4. J, K. Bacutriletessp. cf. B. onodios (Harris, Citation1961) Hopkins et Sweet, Citation1976. J. Proximal view, fluorescence, S079718-01, P40-3. K. Distal view, fluorescence, S079718-01, P40-3

Horstisporites compositus Li, Batten, Li et Peng (Citationin press) is similar to the H. planatus specimens described here in that both have narrow curvaturae and a composite reticulate sculpture (a fine reticulum occurring within the lumina of a coarse reticulum). Li et al. (Citationin press) note that the fine reticulation within the larger areolae is a manifestation of the open construction of the exoexine, thus its value for morphological classification should be carefully considered. For example, the fine reticulation is also present in H. minutus Schultz et Noll, Citation1987 and H. alveolatus Schultz et Noll, Citation1987 (Schultz & Noll Citation1987, pl. 4, figs. 6, 9); however, H. minutus is smaller (spore diameter is consistently around 225 µm) and the muri of its reticulum are arranged radially; H. alveolatus shows subdivision of the coarse mesh of the reticulum into much finer segments (Schultz & Noll Citation1987, pl. 4, fig. 8) and areolae tend to be smaller towards the distal surface. Reticula on our specimens do not show evidence of radial arrangement, and are more or less evenly developed across the whole surface.
Horstisporites sp. cf. H. microlumenus Dettmann, Citation1961()
Descriptionl
Trilete megaspores, amb circular, laesurae narrow, curved, labra thin and short, ca. 160 μm long from the curvaturae to proximal pole, extending to two-thirds of spore radius. Curvaturae imperfect, narrow, ca. 75 μm long. Spore surface densely covered with irregular ridges, ca. 3 μm wide, tapering towards the rounded top, never sharply spinose at the crests, forming reticulate to slightly rugulate surface patterns. In the contact area, ridges coalesce to form tiny circular or polygonal reticula, lumina ca. 3 μm, but generally less than 5 μm, in diameter. Outside contact areas, ridges are arranged to form rugulae or imperfect reticula. Spore outline includes short projections formed by ridged sculpture.
Dimensionsl
Equatorial diameter: 366 μm (n = 1).
Occurrencel
Pliensbachian; Kävlinge BH-928, 46.91–46.07 m.
Remarksl
This specimen is similar to Horstisporites microlumenus Dettmann, Citation1961 in having very small lumina co-occurring with rugulae. However, the lumina of the studied specimen are even smaller (ca. 3 μm) than those (7–18 μm) described by Dettmann (Citation1961). Furthermore, the curvaturae are variably absent to well developed on the specimens of Dettmann (Citation1961, pl. 2, fig. 9) or perfectly developed (Dettmann Citation1961, pl. 2, fig. 8). The variation in ornamentation of the Swedish specimens, from a proximal reticulum to distal rugulae, is uncertain due to corrosion.
Horstisporites sp.()
Figure 10. Horstisporites. Scale bars = 100 μm, or indicated otherwise. A–D. Horstisporites planatus (Marcinkiewicz, Citation1960) Marcinkiewicz, Citation1971. A. Proximal view, SEM, Kävlinge BH-928, 34.75–33.75 m. B. Enlargement, SEM, showing the micro-reticulation in the surface reticulation, Kävlinge BH-928, 34.75–33.75 m. Scale bar = 20 μm. C. Proximal view, SEM, Kävlinge BH-928, 34.75–33.75 m. D. Enlargement, SEM, showing the micro-reticulation in the surface reticulation, Kävlinge BH-928, 34.75–33.75 m. Scale bar = 10 μm. E, F. Horstisporites sp. E. Oblique view, SEM, Kävlinge BH-928, 41.15–40.40. F. Enlargement, SEM, Kävlinge BH-928, 41.15–40.40. Scale bar = 50 μm
Descriptionl
Trilete megaspore, amb triangular. Laesurae distinct, labra strongly raised, ca. 25 μm wide. Imperfect reticula developed on labra and curvaturae.
Dimensionsl
Equatorial diameter: 300 μm (n = 1).
Occurrencel
Pliensbachian; Kävlinge BH-928, 41.15–40.40 m.
Remarksl
Similarly to Horstisporites sp. described here, Horstisporites bertelsenii Fuglewicz, Citation1977 has distinct, strongly elevated trilete laesurae and lacks reticula in the contact areas. However, corrosion of the contact area of Horstisporites sp. and a lack of information on the distal surface hamper a more resolved identification.
Horstisporites? sp. cf. H. sulcatus Fuglewicz, Citation1973
Descriptionl
Trilete megaspore, amb circular. Proximal surface convex. Laesurae straight, almost extending to the equator, labra well developed, membranous, ca. 20 μm wide, up to 60 μm high. Curvaturae distinct, perfectly developed. Contact area scabrate to weakly rugulate; distal surface covered with robust rugulae, 10–20 μm wide, never extending across curvaturae to the contact area.
Dimensionsl
Equatorial diameter: 426 (521) 706 μm (n = 220).
Occurrencel
Rhaetian; Svedala I, 1570–1560 m (S079701, S079713, S079718, S079723–S079725).
Remarksl
These specimens are similar to Horstisporites sulcatus Fuglewicz, Citation1973; both have well-developed laesurae and relatively high labra on the convex contact area, distinct curvaturae, and rugulate ridges on the distal surface. However, there are differences between our specimens and those described by Fuglewicz (Citation1973); the specimens studied here possess wider (10–20 μm) rugulate ridges on the distal surface, which are narrow on the illustrated specimens (10–12 μm, Fuglewicz Citation1973, pl. XXX, ). The specimens of Fuglewicz (Citation1973) have rugulate ornamentation that is different from the “alveolar to reticulate patterns” of Horstisporites Potonié, Citation1956, thus our specimens, and those described by Fuglewicz (Citation1973), may need to be transferred to another genus in the future following more thorough examination.
Our specimens are similar to Biharisporites scarber Marcinkiewicz, Citation1960, with respect to the occurrence of distinct curvature and “larger clusters of low appendages that covered the entire surface of the spore” (translated from Polish, Marcinkiewicz Citation1960, p. 719), and to illustrations of B. scarber in subsequent publications (e.g. Marcinkiewicz Citation1962, pl. IX, , Citation1971, pl. VIII, , Citation1981, pl. XVIII, ). However, in a later description of B. scarber, Marcinkiewicz (Citation1981) states “the distal surface of the sizable area is decorated with small conical processes, irregularly shaped nodules appear on the contact surfaces” (translated from Polish, Marcinkiewicz Citation1981, p. 83). This agrees with the generic diagnosis of Biharisporites Potonié, Citation1956 emend. Bharadwaj et Tiwari, Citation1970 (“exine ornamented with coni, setae or spinae of various shapes … ”, Bharadwaj & Tiwari Citation1970, p. 38), but is different to the description of Marcinkiewicz (Citation1960) and that provided here, therefore we do not assign the specimens recorded here to Biharisporites.
Genus Paxillitriletes (Hall et Nicolson, Citation1973) Batten et Koppelhus, Citation1993
Paxillitriletes ales (Harris, Citation1935) Batten et Koppelhus, Citation1993()
Descriptionl
Trilete megaspore, elliptical in equatorial view. Laesurae distinct, characterized by raised membraneous labra that are slightly incised, jagged at the crests, up to 100 μm high and usually pierced by small holes. Labra subtended by long hair-like capilli, up to 90 μm long. Other parts of the spore surface are covered with short, sparsely distributed capilli or spinae.
Dimensionsl
Equatorial diameter: 381 μm (n = 1).
Occurrencel
Rhaetian; Drillcore Textile Factory, Eslöv, 209.8 m (S063488).
Remarksl
This specimen is similar to those described as Triletes ales (=Paxillitriletes ales) by Harris (Citation1935). The studied specimen lacks an equatorial zona, and this is consistent with the morphology of the type specimen (Harris Citation1935, p. 163, pl. 25, ). An equatorial zona is also absent on Paxillitriletes mensurus (Harris, Citation1935) Hall et Nicolson, Citation1973 and P. arcticus (Bose, Citation1961) Batten et Kovach, Citation1990 (e.g. Harris Citation1935, pl. 26, ; Bose Citation1961, pl. 2, ). The absence of this feature conflicts with the emended generic diagnosis of Paxillitriletes “ … equator surrounded by a zona which is commonly narrow … ” (Batten & Koppelhus Citation1993, p. 38). The inconsistent occurrence of the equatorial zona may necessitate further emendation of the genus, or splitting the genus in two.
Figure 11. Horstisporites? sp. cf. H. sulcatus Fuglewicz, Citation1973. Scale bars = 100 μm. A. Oblique view, transmitted light, S079713, D31-3. B. Oblique view, fluorescence, S079713, D31-3. C. Lateral view, transmitted light, S079725, K34-3. D. Lateral view, fluorescence, S079725, K34-3. E. Distal view, transmitted light, S079701, L32-2. F. Distal view, fluorescence, S079701, L32-2
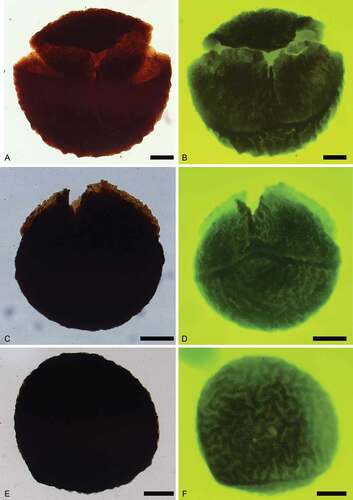
Figure 12. Paxillitriletes. Scale bars = 100 μm, or indicated otherwise. A, B. Paxillitriletes ales (Harris, Citation1935) Batten et Koppelhus, Citation1993. A. Lateral view, transmitted light, S063488, M42-3. B. Lateral view, fluorescence, showing strongly developed capilli near triradiate lamellae, S063488, M42-3. C–F. Paxillitriletes sp. C. Lateral view, SEM, Kävlinge BH-928, 46.91–46.07 m. D. Enlargement, SEM, Kävlinge BH-928, 46.91–46.07 m. Scale bar = 30 μm. E. Enlargement, SEM, Kävlinge BH-928, 46.91–46.07 m. Scale bar = 50 μm. F. Enlargement, SEM, Kävlinge BH-928, 46.91–46.07 m. Scale bar = 50 μm
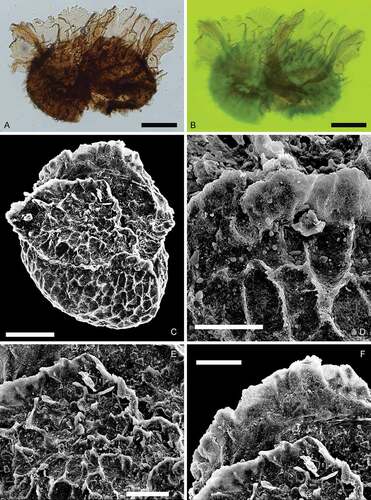
This specimen differs from: Paxillitriletes reticulatus (Madler, Citation1954) Hall et Nicolson, Citation1973, P. phyllicus (Murray, Citation1939) Hall et Nicolson, Citation1973 and P. thorenensis (Mädler, Citation1954) Hall et Nicolson, Citation1973 in lacking reticulation; from P. fairlightensis (Batten, Citation1969) Hall et Nicolson, Citation1973 by its larger size (mean maximum diameter of P. fairlightensis is 224 μm; Batten Citation1969); and from P. granulatus (Mädler, Citation1954) Hall et Nicolson, Citation1973 in lacking fine granules and small warts on the distal surface.
Paxillitriletes sp. ()
Descriptionl
Trilete megaspore, slightly elliptical in lateral view. Laesurae distinct, usually with strongly developed membraneous labra, up to ca. 75 μm high, dissected and slightly convoluted, subtended by elongate and membraneous appendages ca. 30 μm high, spinae occur near appendages, up to ca. 25 μm long. Curvaturae distinct, narrow, raised to form a narrow equatorial zona, merged with labra and membraneous appendages. Both surfaces covered with a course irregular polygonal reticulate ornament.
Dimensionsl
Equatorial diameter: 396 μm (n = 1).
Occurrencel
Pliensbachian; Kävlinge BH-928, 46.91–46.07 m.
Remarksl
This specimen differs from: Paxillitriletes ales (Harris, Citation1935) Batten et Koppelhus, Citation1993 in bearing reticula; from P. fairlightensis (Batten, Citation1969) Hall et Nicolson, Citation1973 in its larger size; and from P. granulatus (Mädler, Citation1954) Hall & Nicolson, Citation1973 in lacking fine granules and small warts on the distal surface.
Similarly to Paxillitriletes reticulatus, P. arcticus, P. phyllicus and P. thorenensis, Paxillitriletes sp. from Kävlinge has reticulate ornamentation. Paxillitriletes arcticus possesses varied meshes, and based on the micrographs of Bose (Citation1961), the reticulum is imperfectly developed. Paxillitriletes phyllicus and P. thorenensis have spinae and warts, respectively, on the muri. Paxillitriletes sp. is most similar to P. reticulatus, the specimen is slightly corroded and the spinae near the labra on the proximal surface are poorly preserved (), making further identification difficult.
Genus Nathorstisporites Jung, Citation1958
Nathorstisporites hopliticus Jung, Citation1958()
Figure 13. Nathorstisporites hopliticus Jung, Citation1958. Scale bars = 100 μm. A. Oblique view, transmitted light, S079741, L41-4. B. Oblique view, fluorescence, S079741, L41-4. C. Enlargement, fluorescence, S079741, L41-4. D. Enlargement, fluorescence, S079741, L41-4. E. Lateral view, transmitted light, S075612, M38-2. F. Lateral view, fluorescence, S075612, M38-2
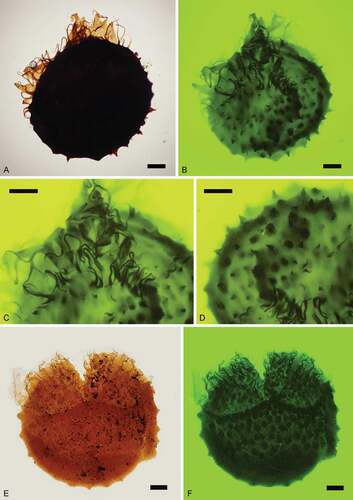
Figure 14. Nathorstisporites hopliticus Jung, Citation1958. Scale bars = 100 μm, or indicated otherwise. A. Proximal view, fluorescence, S066557, P36-2. B. Proximal view, SEM, Munkerup. C. Enlargement, SEM, Munkerup. Scale bar = 50 μm. D. Distal view, SEM, Munkerup. E. Enlargement, SEM, Munkerup. Scale bar = 10 μm. F. Enlargement, SEM, Munkerup. Scale bar = 20 μm
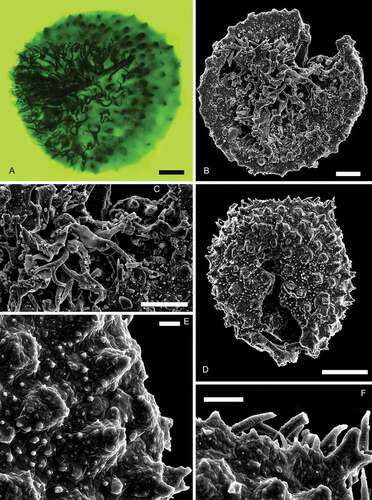
Descriptionl
Trilete megaspore, amb circular. Laesurae distinct, straight or curved, bordered by highly raised membranous labra that are jagged at the crests, partly consisting of strongly sinuous capilli (long hair-like appendages), up to 170 μm long. Capilli grow on numerous rounded verrucae in the contact area and are in some cases bifurcate at the apices, and small coni/tips occur on the top of rounded verrucae on the distal surface. Curvaturae distinct, formed by densely distributed circular verrucae near the equator, merged with the laesurae. Circular verrucae generally ca. 50 μm wide, intervening spaces are ca. 10–50 μm wide. Smaller gemmae, coni and verrucae occur on larger verrucae and over the entire spore surface.
Dimensionsl
Equatorial diameter: 497 (679) 928 μm (n = 107).
Occurrencel
Hettangian; Svedala I, 1531 m (S079741–S079743); Stabbarp BH-74 (S075612); Höllviken II, 1428.08–1422.08 m (S066554, S066555), 1433.81 m (S066549, S066556–S066568, S066560–S066563, S066565, S066566); Helsingborg harbour (S064343, S064347, S064348, S064744, S064747); Munkerup, Bornholm.
Remarksl
The specimens recorded here are larger than those described by Jung (Citation1958; size range: 375–535 μm).
Nathorstisporites hopliticus differs from: N. cornutus Fuglewicz, Citation1980 in lacking horn-like appendages near the laesurae; from N. flagellulatus Dettmann, Citation1961 in lacking thick and large spine-like processes on the distal surface. Nathorstisporites imprimus Reinhardt et Fricke, Citation1969 has bacula and verrucae on the distal surface that differ from the coni of N. hopliticus. Nathorstisporites muricatus Scott et Playford, Citation1985 has complex and long proximal capilli relative to its small size. Nathorstisporites nammalensis Sah et Jain, Citation1968 and N. reticulatus Dettmann, Citation1961 are both distinguished from N. hopliticus by their reticulate surface. Nathorstisporites pulcherrima Helby, Citation1966 has varied spine-like appendages on the distal surface. Nathorstisporites peltasticus has shorter lips, rarely bifurcate proximal capilli and blunt coni.
Discussion
Based on the occurrence of megaspores from the various study sites, we recognize three stratigraphically constrained assemblages, corresponding to the Rhaetian, Hettangian and Pliensbachian stages, respectively ().
The Rhaetian assemblage
This assemblage is characterized by abundant megaspores of relatively high diversity, and comprises Trileites grandis, Banksisporites pinguis, Banksisporites dejerseyi, Banksisporites sinuosus, Banksisporites dettmanniae, Pusulosporites populosus, Bacutriletes tylotus, Bacutriletes sp. cf. B. srivastavae, Bacutriletes sp. cf. B. onodios, Bacutriletes? sp., Horstisporites? sp. cf. H. sulcatus and Paxillitriletes ales. We note that taxa in open nomenclature (except Bacutriletes? sp.) are confined to this assemblage. The occurrences of most taxa in this assemblage, i.e. Banksisporites pinguis, B. dejerseyi, B. sinuosus, B. dettmanniae, Bacutriletes tylotus and Paxillitriletes ales, agree with their previously documented global ranges (Kovach & Batten Citation1989).
The Rhaetian assemblage represents a moderately diverse heterosporous lycopsid flora of Sellaginellales (Trileites?, Banksisporites pinguis; Lundblad Citation1950a, Citation1950b; Kovach Citation1994) or Pleuromeiales (Trileites, Banksisporites; Helby & Martin Citation1965; Balme Citation1995; Taylor et al. Citation2009), and Isoetales (Paxillitriletes; Kovach & Dilcher Citation1985; Kovach Citation1994; Slater et al. Citation2015). This diverse heterosporous lycopsid assemblage recorded from Rhaetian successions that preserve coals, suggests a consistently wet environment for southern Scandinavia during the latest Triassic.
The Hettangian assemblage
This assemblage comprises two taxa, Nathorstisporites hopliticus and Bacutriletes? sp. With the exception of Bacutriletes? sp., all megaspore taxa of the Rhaetian assemblage disappear and are replaced by N. hopliticus, indicating a major loss in diversity across the TJB. Although N. hopliticus is restricted to the Hettangian assemblage, it is recorded from several sites and is generally abundant, but the record of Bacutriletes? sp. is based on a single specimen. Nathorstisporites hopliticus has previously been found in situ within the reproductive structure of the Pleuromeian lycophyte, Lycostrobus scottii (Nathorst Citation1908a; Lundblad Citation1956; Balme Citation1995). The loss of most of the Rhaetian megaspore taxa recorded here, and the dominance of N. hopliticus in the Hettangian indicate a major change in heterosporous lycopsid parent vegetation throughout the Triassic–Jurassic interval in Scandinavia. A decline in megaspore diversity across this interval has also been observed from various localities on a global scale, from Greenland (Harris Citation1937), Sweden (Lundblad Citation1956, Citation1959, this study), Germany (Jung Citation1960), Poland (Marcinkiewicz Citation1971), China (Li et al. Citationin press) and Norway (Morris et al. Citation2009). The disappearance of Sellaginellales and Isoetales from the Rhaetian to the Hettangian assemblage likely represents a shift to drier conditions, which is supported by the lithological change from coal- and mudstone-dominated, to sandstone-dominated facies across the Triassic–Jurassic transition.
The Pliensbachian assemblage
This assemblage was identified from material collected by E. Norling during his work on foraminifera from Kävlinge BH-928 (Norling Citation1968). Six megaspore taxa were recognized from four drillcore samples (): Horstisporites planatus (34.75–33.75 m), Horstisporites sp. (41.15–40.40 m), Narkisporites sp. (42.25–41.15 m), and Bacutriletes sp. cf. B. asaphus, Horstisporites sp. cf. H. microlumenus and Paxillitriletes sp. (46.91–46.07 m). Horstisporites planatus is regarded as an index taxon for the upper Sinemurian to the Toarcian(?) in Poland (Marcinkiewicz Citation1971). The presence of this species in sample 34.75–33.75 m, therefore, suggests an age younger than late Sinemurian; this agrees with dating based on foraminifera that indicates interval 54–28.75 m is Pliensbachian (Norling Citation1968).
The producers of several taxa recorded in the Pliensbachian assemblage remain unresolved, for example, Horstisporites has been described with various wall structures (Kovach Citation1994), and these different forms may have been produced by different parent plants. However, Paxillitriletes sp., probably indicates the presence of isoetalean lycopsids at this time (Kovach Citation1994).
Fluorescence microscopy
A common problem that arises when examining megaspores is that specimens are commonly opaque under transmitted light microscopy (e.g. ). This may inhibit identification of the megaspores and is difficult to overcome if the specimens are embedded in a permanent mounting medium, such as epoxy resin or Canada balsam. Reflected light microscopy and SEM may provide a solution, but only if the specimens have not been embedded in this way. Here we used UV fluorescence microscopy to examine opaque megaspores that were embedded in a fixed mounting medium. Prior to the use of this technique, we were only able to assess the overall shape and size of the opaque specimens (e.g. ). Fluorescence microscopy is used frequently in the analysis of plant cuticle remains (e.g. Bomfleur et al. Citation2014), and to a lesser extent for palynology and palynofacies analysis (e.g. Rodrigues et al. Citation2020). This method provided exceptional results: UV revealed fine-scale details, such as surface folds (), raised trilete labra (), and spore wall architecture () in specimens that were opaque under transmitted light. This technique permits identification of opaque megaspores in standard palynological slides, and assessment of the distal and proximal faces by inverting the slide and/or adjusting the focal plane ().
Conclusions
Based on fossil material housed in the collections of the NRM, we described 19 megaspore taxa from Triassic‒Jurassic successions of Scandinavia. We recognized three distinct megaspore assemblages from the Rhaetian, Hettangian and Pliensbachian stages, and note a decrease in megaspore diversity across the TJB. Our application of fluorescence microscopy to the analysis of opaque megaspores provided exceptional results, suggesting that this non-destructive method may be useful for the re-assessment of historical collections elsewhere.
Acknowledgments
We thank two anonymous reviewers for their detailed and constructive critiques, which improved this paper.
Disclosure statement
No potential conflict of interest was reported by the author(s).
Additional information
Funding
References
- Ahlberg, A., Arndorff, L. & Guy-Ohlson, D., 2002: Onshore climate change during the Late Triassic marine inundation of the Central European Basin. Terra Nova 14, 241–248. doi:https://doi.org/10.1046/j.1365-3121.2002.00416.x.
- Ahlberg, A. & Olsson, I., 2001: Petroleum assessment of the Mesozoic succession in the Höllviken Graben and on the Skurup Platform, southern Sweden. GFF 123, 85–95. doi:https://doi.org/10.1080/11035890101232085.
- Ahlberg, A., Olsson, I. & Šimkevičius, P., 2003: Triassic–Jurassic weathering and clay mineral dispersal in basement areas and sedimentary basins of southern Sweden. Sedimentary Geology 161, 15–29. doi:https://doi.org/10.1016/S0037-0738(02)00381-0.
- Balme, B.E., 1995: Fossil in situ spores and pollen grains: an annotated catalogue. Review of Palaeobotany and Palynology 87, 81–323.
- Banerji, J., Jana, B.N. & Maheshwari, H.K., 1984: The fossil floras of Kachchh, II—Mesozoic megaspores. The Palaeobotanist 33, 190–227.
- Banerji, J., Kumaran, K.P.N. & Maheshwari, H.K., 1978: Upper triassic sporae dispersae from the Tiki formation: megaspores from the Janar Nala Section, South Rewa Gondwana Basin. The Palaeobotanist 25, 1–26.
- Batten, D.J., 1969: Some British Wealden megaspores and their facies distribution. Palaeontology 12, 333–350.
- Batten, D.J. & Koppelhus, E.B., 1993: Morphological reassessment of some zonate and coronate megaspore genera of mainly post-Palaeozoic age. Review of Palaeobotany and Palynology 78, 19‒40. doi:https://doi.org/10.1016/0034-6667(93)90015-M.
- Batten, D.J. & Kovach, W.L., 1990: Catalog of Mesozoic and tertiary megaspores. American Association of Stratigraphic Palynologists, Contributions Series 24, 1–227.
- Bergström, J., 1984: Lateral movements in the Tornquist Zone. GFF 106, 379–380.
- Bertelsen, F. & Michelsen, O., 1970: Megaspores and ostracods from the Rhaeto-Liassic section in the boring Rødby No. 1, southern Denmark. Danmarks Geologiske Undersøgelse, II Række 94, 1–60.
- Bharadwaj, D.C. & Tiwari, R.S., 1970: Lower Gondwana megaspores—a monograph. Palaeontographica B 129, 1‒70.
- Bomfleur, B., McLoughlin, S. & Vajda, V., 2014: Fossilized nuclei and chromosomes reveal 180 million years of genomic stasis in royal ferns. Science 343, 1376‒1377. doi:https://doi.org/10.1126/science.1249884.
- Bose, M.M., 1961: Leaf-cuticle and other plant microfossils from the Mesozoic rocks of Andøya, Norway. The Palaeobotanist 8, 1‒7.
- Brotzen, F., 1950: De geologiska resultaten från borrningarna vid Höllviken. Del. II: undre kritan och trias. Sveriges Geologiska Undersökning C 505, 1–48.
- Collinson, M.E., Barke, J., van der Burgh, J., Van Konijnenburg-van Cittert, J.H.A., Heilmann-Clausen, C., Howard, L.E. & Brinkhuis, H., 2010: Did a single species of Eocene Azolla spread from the Arctic basin to the southern North Sea? Review of Palaeobotany and Palynology 159, 152–165. doi:https://doi.org/10.1016/j.revpalbo.2009.12.001.
- Collinson, M.E., Batten, D.J., Scott, A.C. & Ayonghe, S.N., 1985: Palaeozoic, Mesozoic and contemporaneous megaspores from the Tertiary of southern England: indicators of sedimentary provenance and ancient vegetation. Journal of the Geological Society 142, 375–395. doi:https://doi.org/10.1144/gsjgs.142.2.0375.
- Collinson, M.E., Smith, S.Y., Van Konijnenburg-van Cittert, J.H.A., Batten, D.J., Van Der Burgh, J., Barke, J. & Marone, F., 2013: New observations and synthesis of Paleogene heterosporous water ferns. International Journal of Plant Sciences 174, 350–363. doi:https://doi.org/10.1086/668249.
- Cúneo, N.R., Hermsen, E.J. & Gandolfo, M.A., 2013: Regnellidium (Salviniales, Marsileaceae) macrofossils and associated spores from the Late Cretaceous of South America. International Journal of Plant Sciences 174, 340–349. doi:https://doi.org/10.1086/668811.
- Dettmann, M.E., 1961: Lower Mesozoic megaspores from Tasmania and South Australia. Micropaleontology 7, 71‒86. doi:https://doi.org/10.2307/1484144.
- Erdtman, G., 1947: Suggestions for the classification of fossil and recent pollen grains and spores. Svensk Botanisk Tidskrift 41, 104‒114.
- Fuglewicz, R., 1973: Megaspores of Polish Buntersandstein and their stratigraphical significance. Acta Palaeontologica Polonica 18, 401‒453.
- Fuglewicz, R., 1977: New species of megaspores from the Trias of Poland. Acta Palaeontologica Polonica 22, 405‒431.
- Fuglewicz, R., 1980: Stratigraphy and palaeogeography of lower Triassic in Poland on the basis of megaspores. Acta Geologica Polonica 30, 417‒470.
- Glasspool, I.J., 2000: Megaspores from the Late Permian, Lower Whybrow coal seam, Sydney Basin, Australia. Review of Palaeobotany and Palynology 110, 209‒227. doi:https://doi.org/10.1016/S0034-6667(00)00010-5.
- Glasspool, I.J., 2003: A review of Permian Gondwana megaspores with particular emphasis on material collected from coals of the Witbank Basin of South Africa and the Sydney Basin of Australia. Review of Palaeobotany and Palynology 124, 227‒296. doi:https://doi.org/10.1016/S0034-6667(02)00252-X.
- Gravesen, P., Rolle, F. & Surlyk, F., 1982: Lithostratigraphy and sedimentary evolution of the Triassic, Jurassic and lower Cretaceous of Bornholm, Denmark. Danmarks Geologiske Undersøgelse B 7, 1–51.
- Gry, H., 1969: Megaspores from the Jurassic of the Island of Bornholm, Denmark. Meddelelser Fra Dansk Geologisk Førening 19, 69–89.
- Guy-Ohlson, D., 1981: Rhaeto-Liassic palynostratigraphy of the Valhall bore No. 1 Scania. GFF 103, 233–248.
- Guy-Ohlson, D., 1982: Biostratigraphy of the lower Jurassic–Cretaceous unconformity at Kullemölla southern Sweden. Sveriges Geologiska Undersökning Ca 52, 1–45.
- Guy-Ohlson, D., 1986. Jurassic palynology of the Vilhelmsfält Bore No. 1, Scania, Sweden. Toarcian–Aalenian: section of Palaeobotany, 127. Swedish Museum of Natural History, Stockholm.
- Guy-Ohlson, D., 1990: Pliensbachian palynology of the Karindal bore no. 1, north-west Scania, Sweden. Review of Palaeobotany and Palynology 65, 217–228. doi:https://doi.org/10.1016/0034-6667(90)90072-Q.
- Guy-Ohlson, D. & Malmquist, E., 1985: Lower Jurassic biostratigraphy of the oppegard bore No. 1, NW Scania, Sweden. Sveriges Geologiska Undersökning, Rapporter och Meddelanden 40, 1–11.
- Guy-Ohlson, D. & Norling, E., 1988: Upper Jurassic litho- and biostratigraphy of NW Scania, Sweden. Sveriges Geologiska Undersökning Ca 72, 1–37.
- Guy-Ohlson, D. & Norling, E., 1994: Jurassic sequences in Sweden. Geobios 17, 275–286. doi:https://doi.org/10.1016/S0016-6995(94)80146-0.
- Hall, J.W. & Nicolson, D.H., 1973: Paxillitriletes, a new name for fossil megaspores hitherto invalidly named Thomsonia. Taxon 22, 319‒320. doi:https://doi.org/10.2307/1218173.
- Halle, T.G., 1921: On the sporangia of some Mesozoic ferns. Arkiv För Botanik 17, 1–27.
- Harris, T.M., 1935: The fossil flora of Scoresby Sound, East Greenland. Part 4: ginkgoales, coniferales, lycopodiales and isolated fructifications. Meddelelser Om Grønland 112, 1–176.
- Harris, T.M., 1937: The fossil flora of Scoresby Sound, East Greenland. Part 5: stratigraphic relations of the plant beds. Meddelelser Om Grønland 112, 1–114.
- Harris, T.M., 1961. The Yorkshire Jurassic flora I. Thallophyta‒Pteridophyta, 212. British Museum (Natural History), London.
- Helby, R., 1966: Triassic plant microfossils from a shale within the Wollar Sandstone, N.S.W. Journal and Proceedings, Royal Society of New South Wales 10, 61‒73.
- Helby, R. & Martin, A.R.H., 1965: Cylostrobus gen. nov., cones of Lycopsidean plants from the Narrabeen Group (Triassic) of New South Wales. Australian Journal of Botany 13, 389–404. doi:https://doi.org/10.1071/BT9650389.
- Hermelin, S.G., 1773: Anmärkningar om Boserups stenkols-grufva och de öfrige stenkols-försök uti Skåne. Kungliga Svenska Vetenskapsakademiens Handlingar 34, 236–254.
- Høeg, O.A., Bose, M.N. & Manum, S., 1955: On the double walls in fossil megaspores, with description of Duosporites congoensis n. gen., n. sp. Nytt Magasin for Botanik 4, 101‒106.
- Hopkins, W.S. & Sweet, A.R., 1976: A microflora from a short section of the Paleogene Kishenehn Formation, southeastern British Columbia. Geological Survey of Canada, Paper 76-1B, 307‒309.
- Jung, W., 1958: Zur Biologie und Morphologie einiger disperser Megasporen, vergleichbar mit solchen von Lycostrobus scotti, aus dem Rhät‒Lias Frankens. Geologische Blätter für Nordost-Bayern 8, 114‒130.
- Jung, W., 1960: Die dispersen Megasporen der fränkischen Rhät‒Lias-Grenzschichten. Palaeontographica B 107, 127‒170.
- Kannegieser, V.E. & Kozur, H., 1972: Zur Mikropaläontologie des Schilfsandsteins (Karn). Geologie 21, 185‒215.
- Koppelhus, E.B., 1991: Palynology of the lower Jurassic Rønne formation on Bornholm, eastern Denmark. Bulletin of the Geological Society of Denmark 39, 91–109.
- Koppelhus, E.B. & Batten, D.J., 1992: Megaspore assemblages from the Jurassic and lowermost Cretaceous of Bornholm, Denmark. Danmarks Geologiske Undersøgelse Series A 32, 1–81.
- Kovach, W., 1994. A review of Mesozoic megaspore ultrastructure. In M.H. Kurmann & J.A. Doyle (eds.): Ultrastructure of Fossil Spores and Pollen, 23–37. The Royal Botanic Gardens, Kew, U.K.
- Kovach, W.L. & Batten, D.J., 1989: World-wide stratigraphic occurrences of Mesozoic and Tertiary megaspores. Palynology 13, 247‒277. doi:https://doi.org/10.1080/01916122.1989.9989362.
- Kovach, W.L. & Dilcher, D.L., 1985: Morphology, ultrastructure and paleoecology of Paxillitriletes vittatus sp. nov. from the mid-Cretaceous (Cenomanian) of Kansas. Palynology 9, 85–94. doi:https://doi.org/10.1080/01916122.1985.9989289.
- Kruger, A., Slater, S.M. & Vajda, V., 2021: 3D imaging shark egg cases (Palaeoxyris) from Sweden, new insights into Early Jurassic shark ecology. GFF. doi:https://doi.org/10.1080/11035897.2021.1907442
- Larsson, K., Solakius, N. & Vajda, V., 2000: Foraminifera and palynomorphs from the greensand-limestone sequences (Aptian–Coniacian) in southwestern Sweden. Geologische Jahrbuch fur Geologie und Paläontologie 216, 277–312. doi:https://doi.org/10.1127/njgpa/216/2000/277.
- Larsson, L.M., 2009: Palynostratigraphy of the Triassic–Jurassic transition in southern Sweden. GFF 131, 147‒163. doi:https://doi.org/10.1080/11035890902924828.
- Li, W.B., Batten, D.J., Li, J.G. & Peng, J.G., in press. Mesozoic megaspores and palynomorphs from Tarim Basin, northwest China. Science Press, Beijing.
- Lindström, S. & Erlström, M., 2006: The late Rhaetian transgression in southern Sweden: regional (and global) recognition and relation to the Triassic–Jurassic boundary. Palaeogeography, Palaeoclimatology, Palaeoecology 241, 339–372. doi:https://doi.org/10.1016/j.palaeo.2006.04.006.
- Lindström, S., Erlström, M., Piasecki, S., Nielsen, L.H. & Mathiesen, A., 2017: Palynology and terrestrial ecosystem change of the Middle Triassic to lowermost Jurassic succession of the eastern Danish Basin. Review of Palaeobotany and Palynology 244, 65–95. doi:https://doi.org/10.1016/j.revpalbo.2017.04.007.
- Linné, C., 1751: Carl Linnaei Skånska resa, på höga öfwerhetens befallning förrättad år 1749. Med rön och anmärkningar uti oeconomien, naturalier, antiquiteter, seder, lefnadssätt. Med tillhörige figurer. Med kongl. Maj:ts allernådigste privilegio, uplagd på Lars Salvii kostnad 1751. Stockholm.
- Looy, C.V., Brugman, W.A., Dilcher, D.L. & Visscher, H., 1999: The delayed resurgence of equatorial forests after the Permian–Triassic ecologic crisis. Proceedings of the National Academy of Sciences of the United States of America 96, 13857–13862. doi:https://doi.org/10.1073/pnas.96.24.13857.
- Lund, J.J., 1977: Rhaetic to Lower Liassic palynology of the onshore south-eastern North Sea Basin. Geological Survey of Denmark. II. Series 109, 1–128.
- Lundblad, B., 1948: On some Caytonia plant remains from the Coal mines of Bjuv in Scania (Rhaetic). Svensk Botanisk Tidskrift 42, 84–86.
- Lundblad, B., 1949a: De geologiska resultaten från borrningarna vid Höllviken. Del 3. Microbotanical studies of cores from Höllviken, Scania. Sveriges Geologiska Undersökning C 506, Årsbok 43, 1–17.
- Lundblad, B., 1949b: De geologiska resultaten från borrningarna vid Höllviken. del 4. On the presence of Lepidopteris in cores from “Höllviken II”. Sveriges Geologiska Undersökning C 507, Årsbok 43, 1–11.
- Lundblad, B., 1950a: Studies in the Rhaeto-Liassic floras of Sweden. I. Pteridophyta, Pteridospermae, and Cycadophyta from the Mining District of NW Scania. Kungliga Svenska Vetenskapsakademiens Handlingar, Series 4(1), 1–82.
- Lundblad, B., 1950b: On a fossil Selaginella from the rhaetic of Hyllinge, Scania. Svensk Botanisk Tidskrift 44, 477–487.
- Lundblad, B., 1954: Contributions to the geological history of the Hepaticae. Svensk Botanisk Tidskrift 48, 381–417.
- Lundblad, B., 1956: On the stratigraphical value of the megaspores of Lycostrobus Scottii. Sveriges Geologiska Undersökning C 547, Ärsbok 50, 1–11.
- Lundblad, B., 1958: On the presence of the genus Pseudotorellia (Ginkgophyta) in the Rhaetic of N.W. Scania. GFF 79, 759–765.
- Lundblad, B., 1959: Rhaeto-Liassic floras and their bearing on the stratigraphy of Triassic‒Jurassic rocks. Stockholm Contributions in Geology 3, 83‒102.
- Mädler, K., 1954: Azolla aus dem Quartär und Tertiär sowie ihre Bedeutung für die Taxionomie älterer Sporen. Geologisches Jahrbuch 70, 143‒157.
- Marcinkiewicz, T., 1960: Analiza megasporowa osadów jurajskich okolic Gorzowa Śląskiego-Praszki. Kwartalnik Geologiczny 4, 713‒733. (Russian and English summaries).
- Marcinkiewicz, T., 1962: Megaspory retyku i liasu z wiercenia Mechowo koło Kamienia Pomorskiego i ich wartość stratygraficzna. Instytut Geologiczny, Prace 30, 469‒493. (Russian and English summaries).
- Marcinkiewicz, T., 1971: Stratygrafia retyku i liasu w Polsce na podstawie badań megasporowych. Instytut Geologiczny, Prace 65, 58. (Russian and English summaries).
- Marcinkiewicz, T., 1981. Megaspory [Jura Dolna, Flora]. In L. Malinowska, W. Bielecka & M. Rogalska (eds.): Budowa geologiczna Polski, Atlas skamieniałości przewodnich i charakterystycznych, Mezozoik, Jura. 3(2b; for 1980), 79‒97. Wydawnictwa Geologiczne, Warszawa.
- McLoughlin, S. & Bomfleur, B., 2016: Biotic interactions in an exceptionally well preserved osmundaceous fern rhizome from the Early Jurassic of Sweden. Palaeogeography, Palaeoclimatology, Palaeoecology 464, 86‒96. doi:https://doi.org/10.1016/j.palaeo.2016.01.044.
- McLoughlin, S., Jansson, I.-M. & Vajda, V., 2014: Megaspore and microfossil assemblages reveal diverse herbaceous lycophytes in the Australian Early Jurassic Flora. Grana 53, 22‒53. doi:https://doi.org/10.1080/00173134.2013.848234.
- McLoughlin, S., Tosolini, A.-M.P., Nagalingum, N.S. & Drinnan, A.N., 2002: The Early Cretaceous (Neocomian) flora and fauna of the lower Strzelecki Group, Gippsland Basin, Victoria. Association of Australian Palaeontologists Memoir 26, 1‒144.
- Michelsen, O., Nielsen, L.H., Johannessen, P.N., Andsbjerg, J. & Surlyk, F., 2003: Jurassic lithostratigraphy and stratigraphical development onshore and offshore Denmark. In J.R. Ineson & F. Surlyk (eds.): the Jurassic of Denmark and Greenland. Geological Survey of Denmark and Greenland Bulletin 1, 147–216. doi:https://doi.org/10.34194/geusb.v1.4651.
- Miner, E.L., 1932: Megaspores ascribed to Selaginellites, from the Upper Cretaceous coals of western Greenland. Journal of the Washington Academy of Sciences 22, 497‒506.
- Morris, P.H., Cullum, A., Pearce, M.A. & Batten, D.J., 2009: Megaspore assemblages from the Åre Formation (Rhaetian‒Pliensbachian) offshore mid-Norway, and their value as field and regional stratigraphic markers. Journal of Micropalaeontology 28, 161‒181. doi:https://doi.org/10.1144/jm.28.2.161.
- Murray, N., 1939: The microflora of the Upper and Lower Estuarine Series of the East Midlands. Geological Magazine 76, 478‒489. doi:https://doi.org/10.1017/S0016756800072502.
- Nathorst, A.G., 1875: Om en cycadékotte från den rätiska formationens lager vid Tinkarp i Skåne. Öfversigt Af Kongliga Vetenskapsakademiens Förhandlingar 10, 25–33.
- Nathorst, A.G., 1876: Bidrag till Sveriges fossila flora. Växter från rhätiska formationen vid Pålsjö i Skåne. Kungliga Svenska Vetenskapsakademiens Handlingar 14, 1–82.
- Nathorst, A.G., 1878a: Om floran i Skånes kolförande bildningar 1. Floran vid Bjuf, första häftet. Sveriges Geologiska Undersökning C 27, 1–52.
- Nathorst, A.G., 1878b: Bidrag till Sveriges fossila flora 2. Floran vid Höganäs och Helsingborg. Kungliga Svenska Vetenskapsakademiens Handlingar 16, 1–53.
- Nathorst, A.G., 1878c. Beiträge zur fossilen Flora Schwedens. Über einige rhätische Pflanzen von Pålsjö in Schonen, 34. E Schweizerbart´sche Verlagshandung (E. Koch), Stuttgart.
- Nathorst, A.G., 1879a: Om floran i Skånes kolförande bildningar 1. Floran vid Bjuf, andra häftet. Sveriges Geologiska Undersökning C 33, 53–82.
- Nathorst, A.G., 1879b: Om Spirangium och dess förekomst i Skånes kolförande bildningar. Sveriges Geologiska Undersökning C 36, 1–15.
- Nathorst, A.G., 1886: Om floran i Skånes kolförande bildningar 1. Floran vid Bjuf, tredje häftet. Sveriges Geologiska Undersökning C 85, 83–116.
- Nathorst, A.G., 1902: Beiträge zur Kenntnis einiger Mesozoischen Cycadophyten. Kungliga Svenska Vetenskapsakademiens Handlingar 36, 1‒28.
- Nathorst, A.G., 1906a: Bemerkungen über Clathropteris meniscioides Brongniart und Rhizomopteris cruciata Nathorst. Kungliga Svenska Vetenskapsakademiens Handlingar 41, 1–14.
- Nathorst, A.G., 1906b: Om några Ginkgoväxter från kolgrufvorna vid Stabbarp i Skåne. Kongliga Fysiografiska Sällskapets Handlingar 17, 1–15.
- Nathorst, A.G., 1906c: Über Dictyophyllum und Camptopteris spiralis. Kungliga Svenska Vetenskapsakademiens Handlingar 41, 1–24.
- Nathorst, A.G., 1907: Über Thaumatopteris schenki Nath. Kungliga Svenska Vetenskapsakademiens Handlingar 42, 1–9.
- Nathorst, A.G., 1908a: Paläobotanische Mitteilungen 3. Lycostrobus scotti, eine grosse Sporophyllähre aus den rätischen Ablagerungen Schonens. Kungliga Svenska Vetenskapsakademiens Handlingar 43, 1‒14.
- Nathorst, A.G., 1908b: Paläobotanische Mitteilungen 4. Über die Untersuchung kutinisierter fossiler Pflanzenteile. Kungliga Svenska Vetenskapsakademiens Handlingar 43, 1‒13.
- Nathorst, A.G., 1908c: Paläobotanische Mitteilungen 5. Über Nathorstia Heer. Kungliga Svenska Vetenskapsakademiens Handlingar 43, 14–19.
- Nathorst, A.G., 1908d: Paläobotanische Mitteilungen 6. Antholithus zeilleri n. sp. mit noch erhaltenen Pollenkörnern aus den rhätischen Ablagerungen Schonens. Kungliga Svenska Vetenskapsakademiens Handlingar 43, 20–23.
- Nathorst, A.G., 1909a: Paläobotanische Mitteilungen 8. Über Williamsonia, Wielandia, Cycadocephalus und Weltrichia. Kungliga Svenska Vetenskapsakademiens Handlingar 45, 1–38.
- Nathorst, A.G., 1909b: Über die Gattung Nilssonia Brongn. mit besonderer Berücksichtigung Schwedischer Arten. Kungliga Svenska Vetenskapsakademiens Handlingar 43, 1–40.
- Nathorst, A.G., 1911: Paläobotanische Mitteilungen 10. Über die Gattung Cycadocarpidium Nathorst nebst einigen Bemerkungen über Podozamites. Kungliga Svenska Vetenskapsakademiens Handlingar 46, 1–11.
- Nathorst, A.G., 1912: Paläobotanische Mitteilungen 11. Zur Kenntnis der Cycadocephalus-Blüte. Kungliga Svenska Vetenskapsakademiens Handligar 48, 1–14.
- Neri, M., Kustatscher, E. & Roghi, G., 2018: Megaspores from the Lower Jurassic (Pliensbachian) Rotzo Formation (Monti Lessini, northern Italy) and their palaeoenvironmental implications. Palaeobiodiversity and Palaeoenvironments 98, 97–110. doi:https://doi.org/10.1007/s12549-017-0314-z.
- Norling, E., 1968: On Liassic nodosariid Foraminifera and their wall structures. Sveriges Geologiska Undersokning, Series C 623, Årsbok 61, 1–75.
- Norling, E., 1981: Upper Jurassic and Lower Cretaceous geology of Sweden. GFF 103, 253–269.
- Norling, E., Ahlberg, A., Erlström, M. & Sivhed, U., 1993: Guide to the Upper Triassic and Jurassic geology of Sweden. Sveriges Geologiska Undersökning Ca 82, 1–71.
- Norling, E. & Bergström, J., 1987: Mesozoic and Cenozoic tectonic evolution of Scania, southern Sweden. In P.A. Ziegler (ed.): compressional Intra–Plate Deformations in the Alpine Fore-land. Tectonophysics 137, 7–19. doi:https://doi.org/10.1016/0040-1951(87)90309-X.
- Petersen, H., Lindström, S., Therkelsen, J. & Pedersen, G., 2013: Deposition, floral composition and sequence stratigraphy of uppermost Triassic (Rhaetian) coastal coals, southern Sweden. International Journal of Coal Geology 116, 117–134. doi:https://doi.org/10.1016/j.coal.2013.07.004.
- Pieńkowski, G., 1991: Liassic sedimentation in Scania, southern Sweden: hettangian—Sinemurian of the Helsingborg area. Facies 24, 39–86. doi:https://doi.org/10.1007/BF02536840.
- Potonié, R., 1956: Synopsis der Gattungen der Sporae dispersae I. Teil: sporites. Beihefte zum Geologischen Jahrbuch 23, 103.
- Potonié, R., 1966: Synopsis der Gattungen der Sporae dispersae IV. Teil: nachtrage zuallen Gruppen (Turmae). Beihefte zum Geologischen Jahrbuch 72, 244.
- Pott, C. & McLoughlin, S., 2009: Bennettitalean foliage in the Rhaetian–Bajocian (latest Triassic–Middle Jurassic) floras of Scania, southern Sweden. Review of Palaeobotany and Palynology 158, 117–166. doi:https://doi.org/10.1016/j.revpalbo.2009.08.004.
- Pott, C. & McLoughlin, S., 2011: The Rhaetian flora of Rögla, northern Scania, Sweden. Palaeontology 54, 1025–1051. doi:https://doi.org/10.1111/j.1475-4983.2011.01090.x.
- Punt, W., Hoen, P.P., Blackmore, S., Nilsson, S. & Le Thomas, A., 2007: Glossary of pollen and spore terminology. Review of Palaeobotany and Palynology 143, 1‒81. doi:https://doi.org/10.1016/j.revpalbo.2006.06.008.
- Reinhardt, P. & Fricke, D., 1969: Megasporen aus dem unteren und mittleren Keuper Mecklenburgs. Monatsbericht der Deutschen Akademie der Wissenschaften zu Berlin 11, 399‒411.
- Retallack, G., 1997: Earliest Triassic origin of Isoetes and quillwort evolutionary radiation. Journal of Paleontology 71, 500–521. doi:https://doi.org/10.1017/S0022336000039524.
- Rodrigues, B., Duarte, L.V., Silva, R.L. & Mendonça Filho, J.G., 2020: Sedimentary organic matter and early Toarcian environmental changes in the Lusitanian Basin (Portugal). Palaeogeography, Palaeoclimatology, Palaeoecology 554, 109781. doi:https://doi.org/10.1016/j.palaeo.2020.109781.
- Sah, S.C.D. & Jain, K.P., 1968: Lower Mesozoic megaspores from the Variegated stage of Salt Range (W. Pakistan). The Palaeobotanist 16, 288‒291.
- Schultz, G. & Noll, H., 1987: Die megasporen-assoziation in den unterkretazischen Sedimenten einer Paläokarsthöhle bei Nehden im Sauerland (Rheinisches Schiefergebirge). Palaeontographica B 203, 83‒107.
- Scott, A.C. & Playford, G., 1985: Early Triassic megaspores from the Rewan Group, Bowen Basin, Queensland. Alcheringa 9, 297‒323. doi:https://doi.org/10.1080/03115518508618975.
- Shevchuk, O.A., McLoughlin, S. & Vajda, V., 2020: The first Cretaceous megaspores from Ukraine. Cretaceous Research 118, 1‒13.
- Slater, B.J., McLoughlin, S. & Hilton, J., 2011: Guadalupian (Middle Permian) megaspores from a permineralised peat in the Bainmedart coal measures, Prince Charles Mountains, Antarctica. Review of Palaeobotany and Palynology 167, 140–155. doi:https://doi.org/10.1016/j.revpalbo.2011.07.007.
- Slater, S.M., Taylor, W.A., Batten, D.J., Hill, C.R. & Wellman, C.H., 2015: Morphology and wall ultrastructure of a new and highly distinctive megaspore from the Middle Jurassic of Yorkshire, UK. Review of Palaeobotany and Palynology 216, 33–43. doi:https://doi.org/10.1016/j.revpalbo.2015.01.009.
- Slodownik, M., Vajda, V. & Steinthorsdottir, M., 2021: Fossil seed fern Lepidopteris ottonis from Sweden records increasing CO2 concentration during the end-Triassic extinction event. Palaeogeography, Palaeoclimatology, Palaeoecology 110157. doi:https://doi.org/10.1016/j.palaeo.2020.110157.
- Sorgenfrei, T. & Buch, A., 1964: Deep tests in Denmark, 1935–1959. Danmarks Geologiske Undersøgelse Series III 36, 146.
- Taylor, T.N., Taylor, E.L. & Krings, M., 2009. Palaeobotany, the biology and evolution of fossil plants, 1230. Academic Press, Elsevier, Burlington, MA.
- Torsvik, T. & Cocks, L., 2017. Earth History and Palaeogeography, 317. Cambridge University Press, Cambridge.
- Tralau, H., 1975: An upper Triassic microflora from Vallåkra, southern Sweden. GFF 97, 237–242.
- Troedsson, G., 1951: On the Höganäs Series of Sweden (Rhaeto–Lias). Skrifter Från Mineralogisk- Och Paleontologisk-Geologiska Institutionerna, Lund 7, 1–269.
- Vajda, V., 2001: Aalenian to Cenomanian palynofloras of SW Scania, Sweden. Acta Paleontologica Polonica 46, 403–426.
- Vajda, V., Calner, M. & Ahlberg, A., 2013: Palynostratigraphy of dinosaur footprint-bearing deposits from the Triassic–Jurassic boundary interval of Sweden. GFF 135, 120–130. doi:https://doi.org/10.1080/11035897.2013.799223.
- Vajda, V., Linderson, H. & McLoughlin, S., 2016: Disrupted vegetation as a response to Jurassic volcanism in southern Sweden. Geological Society, London, Special Publications 434, 127–147. doi:https://doi.org/10.1144/SP434.17.
- Vajda, V. & McLoughlin, S., 2005: A new Maastrichtian-Paleocene Azolla species from Bolivia, with a comparison of the global record of coeval Azolla microfossils. Alcheringa 29, 305‒329. doi:https://doi.org/10.1080/03115510508619308.
- Vajda, V., McLoughlin, S., Mays, C., Frank, T.D., Fielding, C.R., Tevyaw, A., Lehsten, V., Bocking, M. & Nicoll, R.S., 2020: End-Permian (252 Mya) deforestation, wildfires and flooding—An ancient biotic crisis with lessons for the present. Earth and Planetary Science Letters 529(Article), 115875. doi:https://doi.org/10.1016/j.epsl.2019.115875.
- Vajda, V. & Raine, J.I., 2003: Pollen and spores in marine Cretaceous/Tertiary boundary sediments at mid-Waipara River, North Canterbury, New Zealand. New Zealand Journal of Geology and Geophysics 46, 255‒273. doi:https://doi.org/10.1080/00288306.2003.9515008.
- Vajda, V. & Wigforss-Lange, J., 2009: Onshore Jurassic of Scandinavia and related areas. GFF 131, 5–23. doi:https://doi.org/10.1080/11035890902975309.
- van der Hammen, T., 1954: Principios para la nomenclatura palinolögica sistemätica. Boletín Geológico 2, 1‒24.