
ABSTRACT
Nineteen ichnotaxa, together with algal and invertebrate remains, and various pseudo-traces and sedimentary structures are described from the Torneträsk Formation exposed near Lake Torneträsk, Lapland, Sweden, representing a marked increase in the diversity of biotic traces recorded from this unit. The “lower siltstone” interval of the Torneträsk Formation contains mostly simple pascichnia, fodinichnia and domichnia burrows and trails of low-energy shoreface to intertidal settings. The assemblage has very few forms characteristic of high-energy, soft-sediment, foreshore or upper shoreface environments (representative of the Skolithos ichnofacies).
Uranium-lead (U-Pb) LA-ICPMS analysis of zircon from a thin claystone layer within the “lower siltstone” interval yielded a maximum depositional age of 584 ± 13 Ma, mid-Ediacaran. Most of the zircon is represented by rounded detrital grains that yield dates between 3.3 and 1.0 Ga. Although the age of the basal sandstone-dominated interval of the Torneträsk Formation remains elusive owing to the absence of fossils, the ichnofossil suite from the overlying “lower siltstone” interval lacks deep arthropod trackways, such as Rusophycus and Cruziana, and is suggestive of a very early (Terreneuvian, possibly Fortunian) Cambrian age. The ichnofauna is otherwise similar to early Cambrian trace fossil assemblages from other parts of Baltica, regions further south in modern Europe, and from Greenland.
Introduction
Lower Cambrian sedimentary successions crop out in a thin belt along the eastern front of the Scandinavian Caledonides. The successions, although moderately deformed during the Caledonian Orogeny (Roberts & Gee Citation1985), are virtually unmetamorphosed and are composed predominantly of siliciclastic rocks. In northernmost Sweden and Norway, these strata are assigned to the Dividal Group. This group has a maximum thickness of approximately 200 m (Axheimer et al. Citation2007) and rests non-conformably on a Proterozoic or Archaean crystalline basement. Three major units have been distinguished within the Dividal Group: the Torneträsk Formation, Grammajukku Formation and Alum Shale Formation in ascending stratigraphic order (Føyn Citation1967; Thelander Citation1982; Axhemier et al. Citation2007; Nielsen & Schovsbo Citation2011).
Generally, the Dividal Group has been considered to be impoverished in body fossils (see Skovsted et al. Citationin press [this volume] for details), resulting in some uncertainty regarding the precise age of the formations within the succession. Trilobites from the top of the Grammajukku Formation are referable to upper Stage 4 of the Cambrian (Ahlberg Citation1980a, Citation1980b, Citation1985; Axheimer et al. Citation2007; Skovsted et al. Citationin press [this volume]). However, the underlying Torneträsk Formation has been more difficult to place in a temporal context (Vidal & Moczydłowska Citation1996). Early studies assigned the Torneträsk Formation to the Ediacaran following the discovery of a putative Ediacara-type organism (Föyn & Glaessner Citation1979). However, this taxon, Kullingia concentrica Glaessner, in Föyn & Glaessner (Citation1979), was later interpreted as a scratch circle (Jensen et al. Citation2002) placing significant doubts on the suggested Ediacaran age for the unit.
One of the more detailed investigations of lower Cambrian strata in northern Sweden was undertaken by Jensen & Grant (Citation1998) who documented an assemblage of trace fossils from the Torneträsk Formation near Luobákte in the Abisko area of Lapland. The assemblage included ichnotaxa, such as Treptichnus pedum (Seilacher Citation1955), Curvolithus and Gyrolithes, and the enigmatic body fossil Sabellidites sp., which led Jensen & Grant (Citation1998) to conclude that the majority of the formation was likely to be of early Cambrian age, pre-dating the earliest trilobites (Rovno or Lontovan). However, a more resolved chronostratigraphic position has remained elusive owing to the absence of age-diagnostic fossils in the basal portion of the Torneträsk Formation (Jensen & Grant Citation1998). More recently, Nielsen and Schovsbo (Citation2011) suggested a relatively late early Cambrian age (equivalent to Dominopolian to “Ljubomilan” or Cambrian Stage 3) for the entire Torneträsk Formation, based on a regional sequence stratigraphic analysis, despite the absence of trilobites and arthropod trace fossils.
By their own admission, the trace fossil assemblage documented by Jensen & Grant (Citation1998) was by no means complete as they focused primarily on ichnofossils that were considered stratigraphically important. Here we build on the work of Jensen & Grant (Citation1998) and document a more diverse trace fossil assemblage from the Torneträsk Formation at several localities near Lake Torneträsk in northern Sweden (). We also identify microbially induced sedimentary structures, algal remains and sparse putative invertebrate body fossils from this unit. Through this study, we aim to assess the age constraints on the Torneträsk Formation and increase understanding of the diversity of early animal life and behaviours in Baltica during this crucial earliest Cambrian interval. Our U-Pb LA-ICPMS analysis of zircon from a claystone bed within the “lower siltstone” interval at Luobákte also places the sedimentary succession in a coarse geochronological framework and provides indications of sediment provenance for the Torneträsk Formation.
Figure 1. Maps of the study area. A. Map of Scandinavia showing the distribution of the Caledonian thrust sheets and the study area. B. Geological map of the Lake Torneträsk area, Lapland showing the studied localities. C. Satellite image of the Orddajohka rivulet area, north of Lake Torneträsk, showing the location of the measured section. D. Satellite image of the Mt Luobákte area south of Lake Torneträsk showing the location of the studied composite section compiled at sites 1 and 2. Distribution of rock units after Thelander (Citation2009); satellite images from Google Earth
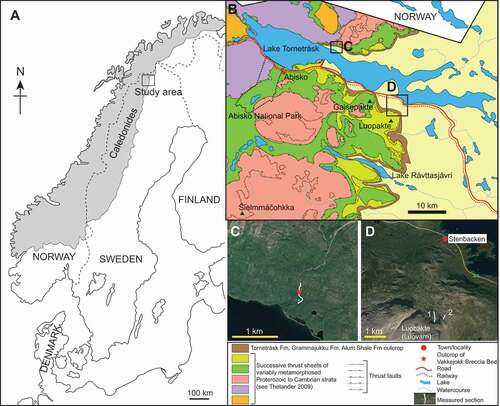
Geological setting
Autochthonous strata at Luobákte and adjacent areas (), now assigned to the Torneträsk, Grammajukku and Alum Shale formations, were first described by Moberg (Citation1908). Subsequent sedimentological and stratigraphic studies by Kulling (Citation1960, Citation1964), Thelander (Citation1982) and Nielsen & Schovsbo (Citation2011), among others, established informal lithostratigraphic subdivisions of these units and summarized their depositional settings within a sequence stratigraphic framework. In addition, Kulling (Citation1930) noted a distinctive boulder-breccia bed intercalated with the otherwise siltstone- and sandstone-dominated successions in the middle part of the Torneträsk Formation to the north of Lake Torneträsk. These earlier studies documented extensive details of the lithological characteristics and depositional settings of these units, so we provide only a short summary of their sedimentary features to provide a palaeoenvironmental context for the body and trace fossil assemblages described below. The Torneträsk Formation was formally established by Thelander (Citation1982) but the constituent “members” were never formally defined, hence, we retain these units under informal nomenclature as “intervals”.
Figure 2. Lithostratigraphic scheme, invertebrate assemblage zones, simplified lithological log (modified from Jensen & Grant Citation1998; Nielsen & Schovsbo Citation2011), and representative field photographs of the Torneträsk Formation to Alum Shale Formation interval exposed on the northern slope of Mt Luobákte, Lapland. A. Basal conglomeratic bed of the “lower sandstone” interval. B. Stacked tabular and wedge-shaped sandstone beds near the top of the “lower sandstone” interval. C. Coarsening-upward package of thinly interbedded siltstones and sandstones of the “lower siltstone” interval. D. Red and green siltstones and fine-grained sandstones in the upper part of the “middle siltstone” interval. E. Repetitive sandstone–siltstone cycles of the “upper sandstone” interval. F. Blocky grey–brown fossiliferous limestone of the upper Grammajukku Formation overlain by contorted organic-rich shales of the Alum Shale Formation
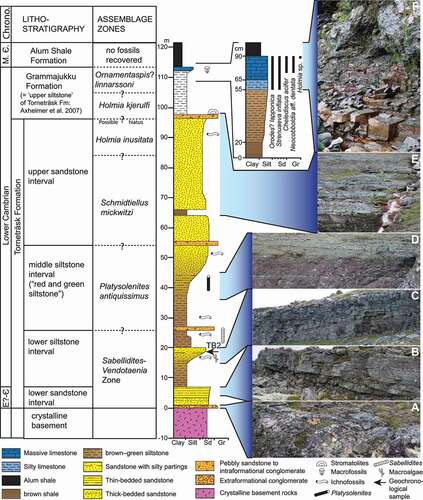
Figure 3. Simplified measured section of the lower part of the Torneträsk Formation exposed in the Orddajohka rivulet, north of Lake Torneträsk, Lapland, showing the stratigraphic position of the Vakkejokk Breccia Bed, trace fossils and wrinkle structures
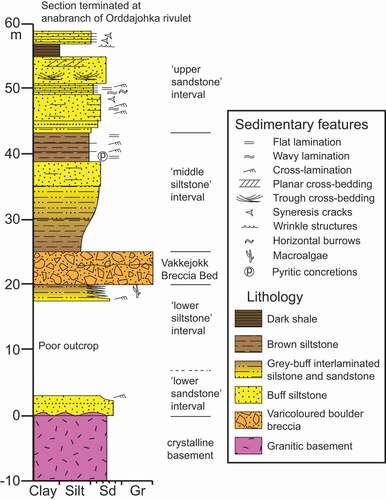
The basal beds of the Torneträsk Formation rest non-conformably on weathered granitic to gneissic basement in the study area. At Luobákte, the base of the formation is characterized by a thin polymict conglomerate (). Stodt et al. (Citation2011) proposed that at least the basal part of this conglomerate was deposited during the mid-Ediacaran Gaskiers glaciation, although we found no evidence of glacigenic features in this deposit in the Torneträsk area. This bed is overlain by a succession(>6 m thick) of quartzose fine- to coarse-grained sandstones and conglomeratic lenses with few or no siltstone partings. These strata are collectively assigned to the informal “lower sandstone” interval. This unit is characterized by extensive high- to low-angle planar cross-bedding () with variable palaeocurrent orientations (Thelander Citation1982). Large-scale straight- to sinuous-crested asymmetrical ripples (), smaller-scale symmetrical ripples, and inclined tabular packages of massive sandstone () are also represented. Thelander (Citation1982, ) also recognized small-scale channel scours within the unit. Although Thelander (Citation1982) inferred a fluvial setting for at least part of this unit, the sheet-like geometries of many beds, common low-angle bedforms, abundant current- and wave-rippled bedding surfaces, and scarcity of siltstone partings suggest a moderate- to high-energy shallow shoreface depositional setting for the “lower sandstone” interval.
Figure 4. Sedimentary features of the Torneträsk Formation at Luobákte (A–D, G–I) and Orddajohka rivulet (E, F). A. High-angle planar cross-bedding; “lower sandstone” interval. B. Large-scale straight to gently sinuous ripples on the surface of a sandstone bed; “lower sandstone” interval. C. Stacked tabular to wedge-shaped low-angle cross-bedded sandstones; “lower sandstone” interval. D. Thinly interbedded fine-grained thin sandstones and siltstones in the upper part of the lower fining-upward cycle of the “lower siltstone” interval. E. Base of the Vakkejokk Breccia Bed sharply overlying thin tabular sandstones and siltstones of the “lower siltstone” interval. F. Chaotically distributed large basement clasts set in a silty matrix within the lower part of the Vakkejokk Breccia Bed. G. Cross-laminated (asymmetrically rippled) fine-grained sandstone bed; “lower siltstone” interval. H. Polymict, conglomeratic bed with angular to sub-rounded clasts in the “upper sandstone” interval. I. Package of siltstones grading up to tabular sandstones incorporating a thin ?bentonitic band (arrowed) in the upper part of the “middle siltstone” interval. Scale bars = 10 cm for A, D–F, I; 10 mm for G, H. Hammer = 30 cm in B, C.

The “lower siltstone” interval of Thelander (Citation1982) rests sharply but conformably on the “lower sandstone” interval – the relatively abrupt contact likely representing the equivalent of a flooding surface and significant deepening of the depositional environment (Nielsen & Schovsbo Citation2011). The “lower siltstone” interval, consisting of two coarsening-upward cycles of grey–brown siltstones to thin-bedded, fine-grained, grey–white sandstones (), probably represents two drowning-shallowing cycles (as opposed to a single cycle interpreted by Nielsen & Schovsbo Citation2011). Flaser bedding, lenticular bedding and symmetrical to asymmetrical ripples are common in the sandstone-dominated upper parts of each sedimentary cycle suggesting a mix of shallow-water wave and current processes (), an interpretation supported by all previous investigations of the succession (Kulling Citation1960; Thelander Citation1982; Nielsen & Schovsbo Citation2011). Bedding surfaces are typically overprinted by an extensive range of trace fossils. The coarsening-upward cycles of the “lower siltstone” interval are interpreted to represent shallowing deposition within relatively quiet-water shoreface up to intertidal settings. The presence of sporadic mudflake conglomerates (Thelander Citation1982) and desiccation features (sand-filled mudcracks with V-shaped profiles that, in some cases, transect ripples and trace fossils) indicate episodic subaerial exposure of bedding surfaces. A thin claystone in the upper part of this interval was sampled for U-Pb geochronology of detrital zircon grains in this study.
To the north of Lake Torneträsk, the “lower siltstone” interval is sharply overlain by a prominent boulder-breccia (), formally defined by Ormö et al. (Citation2017) as the Vakkejokk Breccia Bed, having a thickness of 2–27 m and exposed over a strike length of about 7 km. The breccia incorporates chaotically distributed angular to subrounded clasts of crystalline basement lithologies and minor sedimentary rocks set in a matrix of siltstone to granulestone (). Weakly defined internal partings and fining-upward intervals are locally recognizable in this breccia – the unit as a whole being interpreted as an impact ejecta deposit (Ormö et al. Citation2017). At Luobákte, the Vakkejokk Breccia Bed is not readily recognizable, but has been interpreted to correlate to a thin conglomeratic bed and dolomitic sandstone capping the “lower siltstone” interval (Thelander Citation1982; Jensen & Grant Citation1998).
The “green-grey-red shale” (Moberg Citation1908), “middle shale” (Kulling Citation1960), “middle shale formation” (Kulling (Citation1964), “red and green siltstone” (Thelander Citation1982) or “middle siltstone” interval (Stodt et al. Citation2011; this study) consists of a succession of reddish and greenish siltstones that coarsens upward to thin-bedded fine-grained sandstones (). Thin clay partings in the upper sandy part of this unit () have the appearance of weathered (bentonitic) tuff bands and were sampled for U-Pb geochronology in this study but proved to contain only detrital zircon grains. This unit contains very sparse invertebrate body fossils and is widely traceable in the Scandinavian Caledonides. It is interpreted to have been deposited initially in quiet shelf waters (as part of a transgressive systems tract—and presumably also encompassing a highstand depositional component) but transitioning, in its upper part, to shoreface settings as part of a falling stage systems tract (Thelander Citation1982; Nielsen & Schovsbo Citation2011).
The “upper sandstone” interval is about 35 m thick at Luobákte and, although dominated by tabular sandstones, includes a complex array of lenticular sandstones, persistent siltstone packages, minor conglomeratic lag deposits (), and a nodular phosphorite bed that caps this interval. Lenses of calcareous sandstone, accessory glauconite, various invertebrate traces, sparse body fossils, and extensive ripple-marked bedding planes in this unit suggest predominantly shoreface deposition (Thelander Citation1982; Jensen & Grant Citation1998). Nielsen and Schovsbo (Citation2011) recognized at least two transgressive-regressive cycles within this interval.
The overlying Grammajukku Formation (equivalent to the “upper siltstone” interval of the Torneträsk Formation sensu Thelander Citation1982; see discussion by Nielsen & Schovsbo Citation2011) consists of greenish grey siltstone with local phosphorite nodules and lenses of sandstone and limestone. The upper few metres of this formation consist of thick-banded calcareous shale and dark pyritic limestone (). Brachiopods, hyoliths and trilobites occur at several levels in the siltstones, and the calcareous layers at the top of the formation have yielded a moderately diverse trilobite and Small Shelly Fossil fauna attributable to the Ornamentaspis? linnarssoni Assemblage Zone (Cambrian Stage 4: Axheimer et al. Citation2007; Skovsted et al. Citation2020 [this volume]).
The Grammajukku Formation is sharply overlain by the Alum Shale Formation (), a black (kerogen-rich), pyritic shale unit of middle Cambrian age that is widely distributed across Scandinavia and was deposited in relatively anoxic marine waters. At Luobákte, allochthonous Caledonide slivers, locally yielding a trilobite fauna similar to that of the uppermost Grammajukku Formation (Ahlberg Citation1980a), have been thrust over the ductile Alum Shale Formation, such that the latter has become heavily deformed.
Material and methods
Trace fossils studied from the Torneträsk Formation derive from several sources. Primarily, new material was collected in the field (August, 2020) from outcrop () in the main gully section (site 1) on the northern slope of Mt Luobákte, and from low strike ridges 300 m to the east (site 2). A few specimens were also recovered from the western side of Mt Gaisepakte to the south of Lake Torneträsk, and from the Orddajohka rivulet to the north of the lake (). During this fieldwork, strata of the “lower siltstone” interval were targeted primarily to assess the diversity of ichnofossils within this unit. Available time did not permit thorough quantitative or high-resolution stratigraphic surveys of the ichnoassemblages. Additionally, 39 fossiliferous slabs and synthetic casts from the Torneträsk area were studied from historical collections held in the Palaeobiology Department, Swedish Museum of Natural History, recovered from the field by departmental staff J. Bergström and J. Hagström over the course of several decades. Museum registration numbers and source details for all taxa described below are provided in Supplementary Online File 1.
Trace fossils were photographed in the field and laboratory with a Canon Eos 40D digital camera using low-angle illumination from the upper left, unless stated otherwise. Fossil algae and microbial mat structures were photographed in the laboratory using an Olympus BX51 microscope equipped with a Lumenera Infinity 2 digital camera and an Olympus SZX10 stereomicroscope with an Industrial Digital Camera.
LA-ICPMS methods
Zircon grains were separated from rocks using standard techniques, annealed at 900°C for 60 hours in a muffle furnace, and mounted in epoxy and polished until their centres were exposed. Cathodoluminescence (CL) images were obtained with a JEOL JSM-300 scanning electron microscope and Gatan MiniCL (Supplementary Online File 2). Zircon grains were analyzed by laser ablation inductively coupled plasma mass spectrometry (LA-ICPMS) using an iCAP RQ Quadrupole ICP-MS and Teledyne Photon Machines Analyte Excite+ 193 nm excimer laser ablation system with HelEx II Active two-volume ablation cell. In-house analytical protocols, standard materials, and data reduction software were used for acquisition and calibration of U-Pb dates and a suite of high-field-strength elements (HFSE) and rare-earth elements (REE). Zircon was ablated with a laser spot of 20 µm diameter using fluence and pulse rates of 3 J/cm2 and 10 Hz, respectively, during a 35 second analysis (15 sec gas blank, 20 sec ablation) that excavated a pit c. 20 µm deep. Ablated material was carried by a 0.5 L/min He gas stream in the inner cell and a 1.1 L/min He gas stream in the outer cell. Dwell times were 10 ms for Si, 1 ms for Zr, 5 ms for Hf, 200 ms for 49Ti and 207Pb, 100 ms for 206Pb, 20 ms for 202Hg and 204Pb, 10 ms for 238U, and 10 ms for all other HFSE and REE. Background count rates for each analyte were obtained prior to each spot analysis and subtracted from the raw count rate for each analyte. Ablations pits that appear to have intersected glass or mineral inclusions were identified based on Ti and P.Uranium-lead dates from these analyses are considered valid if the U-Pb ratios appear to have been unaffected by the inclusions. Analyses that appear contaminated by common Pb were rejected based on mass 204 being above baseline. For concentration calculations, background-subtracted count rates for each analyte were internally normalized to 29Si and calibrated with respect to NIST SRM-610 and −612 glasses as the primary standards. Temperature was calculated from the Ti-in-zircon thermometer (Watson et al. Citation2006). Because there are no constraints on the activity of TiO2, an average value in crustal rocks of 0.8 was used.
Data were obtained in two experiments in March 2021 (Supplementary Online File 3). For U-Pb and 207Pb/206Pb dates, instrumental fractionation of the background-subtracted ratios was corrected and dates were calibrated with respect to interspersed measurements of zircon standards and reference materials. The primary standard Plešovice zircon (Sláma et al. Citation2008) was used to monitor time-dependent instrumental fractionation based on two analyses for every 10 analyses of unknown zircon. A secondary correction to the 206Pb/238U dates was made based on results from the zircon standards Seiland (531 Ma, unpublished data, Boise State University) and Zirconia (327 Ma, unpublished data, Boise State University), which were treated as unknowns and measured once for every 10 analyses of unknown zircon. These results showed a linear age bias ofseveral percent that is related to the 206Pb count rate. The secondary correction is considered to mitigate matrix-dependent variations due to contrasting compositions and ablation characteristics between the Plešovice zircon and other standards (and unknowns).
Radiogenic isotope ratio and age error propagation for all analyses includes uncertainty contributions from counting statistics and background subtraction. Errors without and with the standard calibration uncertainty are shown in the data table. For groups of analyses that are collectively interpreted from a weighted mean date, a weighted mean date is first calculated from equivalent dates (probability of fit >0.05) using Isoplot 3.0 (Ludwig Citation2003) with errors on individual dates that do not include a standard calibration uncertainty. A standard calibration uncertainty is then propagated into the error on the date. This uncertainty is the local standard deviation of the polynomial fit to the interspersed primary standard measurements versus time for the time-dependent, relatively larger U/Pb fractionation factor, and the standard error of the mean of the consistently time-invariant and smaller 207Pb/206Pb fractionation factor. These uncertainties are 1.6% (2σ) for 206Pb/238U and 0.4% (2σ) for 207Pb/206Pb. Errors on single analyses without the standard calibration uncertainty are given below. Age interpretations are based on 207Pb/206Pb dates for analyses with 207Pb/206Pb and 206Pb/238U dates >1000 Ma. Otherwise, interpretations are based on 206Pb/238U dates. Analyses with 206Pb/238U dates >1000 Ma and >10% discordance are not considered. Errors are at 2σ.
Results
U-Pb geochronology
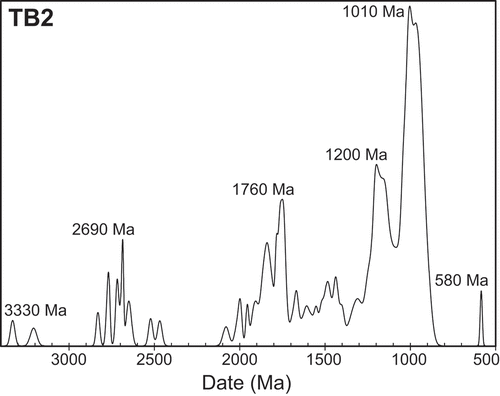
One hundred and forty zircon grains yielded LA-ICPMS dates of 3331 ± 27 to 569 ± 19 Ma. All except one grain yielded dates of 953 ± 51 Ma and older (; Supplementary Online Files 2 and 3). Peaks in the probability density plot are at 2690, 1760, 1200, and 1010 Ma (). The five youngest dates are from a single grain (Supplementary Online File 2) and yield a weighted mean of 584 ± 13 Ma (mean squared weighted deviation = 2.0, probability of fit = 0.09). This is the interpreted maximum depositional age.
Figure 5. Probability density plots of LA-ICPMS U-Pb dates from 140 detrital zircons obtained from sample TB2, representing a thin claystone near the top of the lower upward-coarsening cycle of the “lower siltstone” interval of the Torneträsk Formation at Site 1, Luobákte (). Plotted with Isoplot 3.0 (Ludwig Citation2003)
Microbially induced sedimentary structures
Elephant-skin texture ()
Figure 6. Microbially induced sedimentary structures and algal remains from the Torneträsk Formation. A. Elephant-skin texture formed by a bacterial mat on bedding surface; “upper sandstone interval” of Torneträsk Formation in the Orddajohka rivulet; X00010171a. B. Enlargement of longitudinally striate algal thallus; siltstone under Vakkejokk Breccia, Orddajohka rivulet; X00010211. C. Wrinkled organic remains of algal thallus on dark siltstone; X00010211 (source as above). D. Matted strap-shaped algal thalli; upper part of “middle siltstone” interval, section 1, Luobákte; X00010223. E. Strap-shaped algal thalli from upper part of “lower siltstone” interval, section 1, Luobákte; X00010233. F. Strap-shaped thallus with apparent equal bifurcations; X00010211 (source as above). G. cf. Platysolenites antiquissimus Eichwald, Citation1860, segmented or transversely striate, solid calcareous tube in reddish siltstone; “middle siltstone” interval, section 1, Luobákte. H. Large, flexuous, transversely striate, flattened, tubular Sabellidites cambriensis Yanichevsky, Citation1926; upper part of “lower siltstone” interval, section 1, Luobákte; X00010234. I. Prominent transverse striae on basal part of Sabellidites cambriensis; X00010234 (source as above). Scale bars = 10 mm for A, D, H; 1 mm for B, C, E, F, G, I.

Description
Microbial mat textures (wrinkle structures) preserved on the surfaces of 2–4-cm-thick beds of indurated, homogenous, light grey siltstones, some with medium-scale wavy undulations (). The wrinkle structures cover extensive flat surfaces and individual textural elements are <1 mm high. The wrinkles occur mainly as tufted and elongate mounds 2–3 mm wide, separated by rounded troughs or pits 3–4 mm in diameter. The crests are slightly flattened. Surfaces are locally coated by a thin Fe-enriched film, probably representing pyritic or jarositic veneers resulting in a yellowish-orange appearance.
Remarks
Microbially induced sedimentary structures (MISS) result from biogenic (microbial) modification of sediments (via baffling, trapping and binding grains) to produce distinctive textures and lamina surface features (Noffke Citation2014). Hence, by definition, they can be considered both primary sedimentary structures and microbial ichnofabrics (Noffke Citation2009; Lin et al. Citation2017). The examples from the Torneträsk Formation have morphologies consistent with elephant-skin type wrinkle structures (Hagadorn & Bottjer Citation1997; Peterffy et al. Citation2016). Kinneyia-type morphotypes were not observed. The latter differ from the MISS of this study by forming flat-topped winding crests separatedby mm-scale troughs (Porada & Bouougri Citation2007). No convincing invertebrate trace fossils were seen on the surfaces with wrinkle structures but we can not exclude that stabilization and preservation of some of the indented trails described below were aided by thin microbial films. Rock surfaces with elephant skin textures are best represented in the “upper sandstone” interval of the Torneträsk Formation, several metres above the Vakkejokk Breccia, especially in the Orddajohka rivulet.
Microbial mat elephant-skin textures have been reported typically from shallow marine to intertidal settings (Manten Citation1966; Runnegar & Fedonkin Citation1992; Hagadorn & Bottjer Citation1997; Gehling Citation1999; Weber et al. Citation2012). However, this may partly result from the greater number of studies of such palaeoenvironments. Since cyanobacteria are typically involved in their formation, such mats are normally restricted to the marine photic zone, but they can extend into freshwaters, hypersaline lakes, alkaline lakes and thermal springs (Stal & Caumette Citation1994; Heyl et al. Citation2010). Microbial mats generally establish more readily on sediments with fine grain sizes (Stal et al. Citation1985), which is consistent with our observations. Studies of modern ecosystems have also shown that some microbial mats incorporate anaerobic lithotrophs, such as purple sulphur bacteria, which metabolize sulphur (Brocks et al. Citation2005) and proliferate during the winter at times of low primary productivity and low oxygen levels. Purple sulphur bacteria were possibly present in the studied Cambrian ecosystems, evidenced by the sulphide/sulphate staining of the fossils.
Occurrence. – Moderately common in the upper part of the coarsening-upward cycles of the “lower siltstone” interval and the “upper sandstone” interval at all sites, but best represented in the “upper sandstone” interval at Orddajohka rivulet, north of Lake Torneträsk ().
Systematic palaeontology
PHYLUM OCHROPHYTA Cavalier-Smith, Citation1995CLASS PHAEOPHYCEAE Kjellman, Citation1891
“Vendotaenids” (–F)
Description
Delicate (1–3 mm wide), dichotomously branched, cylindrical or slender ribbon-like thalli with irregular (anisotomous) bifurcations (5B–F). Preserved length up to 20 mm but the thalli complex may cover areas of up to 20 × 20 cm of the bedding surface. Thalli bifurcate at angles of c. 45° (). Surface morphology is mostly laevigate but some examples bear longitudinal striae (). Where the thallus is thick, a rugulate pattern is locally preserved (). Thalli occur as matted overlapping aggregates on some slabs (). These fossils are generally yellowish-orange and contrast strongly with the host sediments, but a few dark, organic-rich examples are also present ().
Remarks
Class Vendophyceae (vendotaenids) was erected by Gnilovskaya et al. (Citation1988) and includes Vendian and Cambrian macroalgae (Gnilovskaya et al. Citation1988; Tewari Citation1999; Xiao & Dong Citation2006). Although the affinity of vendotaenids has been disputed, with some researchers considering them to be the remains of sulfide-oxidizing organotrophic bacteria (Vidal Citation1989), the predominant view is that they represent planktonic macroalgae (e.g., Bykova et al. Citation2020; Wang et al. Citation2020; Xiao et al. Citation2020). We do not discount that some forms included in vendotaenids may represent bacterial sheets; however, we consider the Torneträsk specimens to be macroalgae based on their relatively large maximum width, true dichotomous branching and lack of “chess-board” patterning on the thalli. Based on the lack of anchoring structures we agree with Xiao et al. (Citation2020) that these algae might have had a planktonic habit.
These thalli were possibly dendroid in original growth form but appear to have been disarticulated and transported into matted accumulations of thalli segments. No holdfasts or reproductive structures were identified. The lack of diagnostic characters prevents assignment to any specific taxon, but the studied specimens are broadly similar to various small, branched, ribbon-like, thalloid algae described from the early Palaeozoic (see, e.g., LoDuca et al. Citation2017; Xiao et al. Citation2020). The striking orange colour of some specimens () is possibly the result of post mortem bacterial masking and mineralization during the decay process, whereas the dark examples retain organic matter () suggesting rapid burial.
Occurrence
Several specimens from the upper part of the lowermost coarsening-upward cycle of the “lower siltstone” interval at sites 1 and 2, Luobákte.
PHYLUM ANNELIDA Lamarck, Citation1802CLASS POLYCHAETA Grube, Citation1850ORDER CANALIPALPATA (Rouse & Fauchald, Citation1997)FAMILY SIBOGLINIDAE Caullery, Citation1914Genus Sabellidites Yanichevsky, Citation1926
Sabellidites cambriensis Yanichevsky, Citation1926 (, I)
Description
The single available specimen consists of a straight unbranched tube, c. 2 cm long and 1.5 mm wide, with parallel sides. Thin, closely spaced transverse segments extend over the entire length of the fossil. The fossil and host rock are yellowish to dark grey; there is no sign of mineralization of the organic remains.
Remarks
Sabellidites has generally been considered to represent the remains of organic tube-dwelling annelids that appeared in the late Ediacaran and persisted through the early Cambrian (Urbanek & Mierzejewska Citation1983; Moczydłowska et al. Citation2014; Hybertsen Citation2017). Sabellidites cambriensis has been reported widely in lower Cambrian strata from Baltica (Systra & Jensen Citation2006; Moczydłowska et al. Citation2014), Newfoundland (Landing et al. Citation1989) and China (Yang et al. Citation2006). Sabellidites cambriensis was reported previously from the “lower siltstone” interval of the Torneträsk Formation at Luobákte by Jensen and Grant (Citation1998). Moczydłowska et al. (Citation2014) provided strong evidence that, based on specimens from terminal Ediacaran strata, the morphology, histology and ultrastructure of such fossils, especially the wall architecture of parallel chitinous fibers embedded in a proteinaceous matrix within multilayered sheets, is consistent with affiliation of Sabellidites to siboglinid polychaete worms. However, Georgieva et al. (Citation2019) argued that the ultrastructural characters remain inconclusive with regard to phylogenetic affinities.
Occurrence
The precise stratigraphic position of the isolated specimen is uncertain; it probably derives from the “lower siltstone” interval at Luobákte based on the only previous record ().
INCERTAE SEDISGenus Platysolenites Pander, Citation1851
cf. Platysolenites antiquissimus Eichwald, Citation1860 ()
Description
Solid, cylindrical, smooth, calcareous tube, 11.5 mm long, 1.3 mm wide, divided into segments 0.8–1 mm long.
Remarks
Platysolenites antiquissimus is generally considered to represent very early tubular agglutinated foraminifera, but various other biological affinities have also been postulated (McIlroy et al. Citation2001). The single available Torneträsk Formation specimen appears to have coarser and more regular segments than typical Platysolenites antiquissimus and it might alternatively represent the calcareous filling of a small worm tube or burrow or even a slender cnidarian. Platysolenites antiquissimus is widely recognized in lower Cambrian strata (Streng et al. Citation2005) and has been reported from several exposures of the Dividal Group (Föyn & Glaessner Citation1979), including the “middle siltstone” interval of the section at Luobákte (Jensen & Grant Citation1998). Platysolenites antiquissimus is one of the oldest skeletal fossils from Ediacaran–Cambrian boundary strata in Avalonia and Baltica (McIlroy et al. Citation2001) and is used as a zonal taxon in parts of this region (the Platysolenites antiquissimus Zone defines the Lontova Stage = pre-trilobite part of the Cambrian succession in Estonia: Mens & Pirrus Citation1997).
Occurrence
Precise stratigraphic position uncertain; probably from the “middle siltstone” interval at Luobákte based on the lithology of the isolated sample and the only previous record of the taxon in this section ().
Ichnofossils
Ichnogenus Bergaueria Prantl, Citation1945Bergaueria perata Prantl, Citation1945 (–F)
Figure 7. Trace fossils from the “lower siltstone” interval of the Torneträsk Formation. A–F. Bergaueria perata Prantl, Citation1945; G, H. Bergaueria isp.; I–L. Laevicyclus sp.; M. cf. Rhizocorallium isp. A. Three vertical burrows in positive hyporelief; X00010197. B. Several vertical burrows, one with a raised central knob; X00010183. C. Plug-like tubular burrow in lateral view showing surrounding laminae drawn downwards; X00010197. D. Vertical burrow with central depression in positive hyporelief; X00010221. E. Vertical burrow in epirelief showing dark-stained infill; X00010197. F. Burrow-fill transected by sand-filled desiccation crack; X00010226. G. Large vertical burrow in positive hyporelief, transecting a horizontal sand-filled burrow; X00010184. H. Counterpart of X00010184; photographed in the field. I. Funnel-shaped vertical burrow entrance; specimen photographed in the field. J. Burrow entrance with slightly raised rim; X00010162. K. Funnel-shaped, manganese-stained burrows within thin sandstone bed in lateral view; X00010162. L. Lateral view of funnel-shaped burrow with weakly meniscate fill; X00010162. M. Horizontal, fan-shaped burrow with low-relief spreiten in positive hyporelief with adhering dark siltstone; specimen photographed in the field. Scale bars = 10 mm

Description
Short, vertical, tubular, sand-filled structures that extend up to8 mm beyond the undersurface of sandstone beds to form short cylindrical, plug- or button-like structures in siltstones (, B). The basal surface is generally rounded, but may also be flat or have a central tubercle or depression (, D). These structures are typically 5–21 mm in diameter () but there is also a range of smaller (2.5–5.5 mm in diameter), less regular button-shaped objects that might represent diminutive and ill-preserved specimens. Typical examples are 15–18 mm tall, either lacking an obvious lining, or having a < 1 mm wide rim of mineral-stained, slightly finer grained sandstone, with an outer surface that is smooth or having a weakly concentric ornament (). Laminae of the surrounding sediment are drawn down slightly towards the margin of the burrow (). The upper surface of the bedding is slightly depressed around the dark-stained burrow entrance (). One example is transected by a sand-filled desiccation crack ().
Figure 8. Histograms of burrow diameters for selected trace fossils from the “lower siltstone” interval of the Torneträsk Formation. A. Maximum burrow diameters of Bergaueria perata (black bars: 24 specimens) and Bergaueria isp. (grey bar: 1 specimen). B. Maximum lobe diameters of Treptichnus pedum burrows for 136 specimens
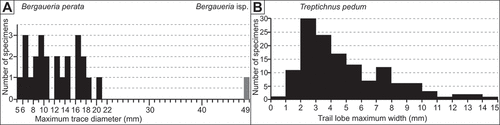
Remarksl
Bergaueria perata differs from B. hemispherica Crimes et al., Citation1977 in having a central depression on the lower surface of the burrow cast and being somewhat deeper than wide (Pemberton et al. Citation1988). Bergaueria radiata Alpert, Citation1973 differs from these species in having prominent radial ridges around the central depression. Since the relative dimensions and surface ornament are markedly variable in the examples from Luobákte (–F, 7A) and other sites (Shitole et al. Citation2019), and the surface features can be easily obscured by adhering sediment or weathering, we consider these ichnotaxa to be probable synonyms – a status alluded to by Jensen (Citation1997). Moreover, Pemberton and Magwood (Citation1990) illustrated examples of these taxa co-preserved in gregarious associations that suggest these ichnospecies represent different preservational states of a single population of trace producers. Although a central depression is not evident in all cases, the examples from Luobákte appear to be structurally consistent with Bergaueria perata Prantl, Citation1945. Equivalent examples at the larger end of the size range were illustrated from 18 m above the base of the Torneträsk Formation at Luobákte by Jensen and Grant (Citation1998). Bergaueria has a long stratigraphic range (?Ediacaran to Cenozoic), occurs in a broad range of bathymetric settings, and is generally interpreted to represent a shallow dwelling burrow of an anemone-like organism (Pemberton et al. Citation1988; Shitole et al. Citation2019).
Occurrencel
Bergaueria perata is common in the heterolithic facies near the top of the lower siltstone interval of the Torneträsk Formation at sites 1 and 2, Luobákte, but occurs as scattered individuals rather than in richly gregarious assemblages as documented in some other Palaeozoic deposits (Pickerill Citation1998).
Bergaueria isp. (, H)
Descriptionl
A single, sand-filled, circular structure (elevated in hyporelief) at the base of a fine-grained sandstone block is available. It is 49 mm in diameter and its centre is raised above the surface of the slab by c. 8 mm. A portion of the margin preserves ring-like wrinkles, but the structure is otherwise featureless (). The counterpart is preserved as a shallow, featureless, hemispherical depression ().
Remarksl
The structure appears to be a short vertical tube or hemispherical (plug-like) fill of Bergaueria type, but is around 2.5–5 times the diameter of typical examples of B. perata from the Torneträsk Formation (). Bergaueria perata burrows of similar size (up to45 mm diameter) were described from lower Cambrian strata of Poland (Orłowski & Żylińska Citation2002) and Pemberton and Magwood (Citation1990) recorded examples up to 55 mm wide from the lower Cambrian Gog Group of western Canada. On this basis the described specimen (NRM X00010184) could be assigned to B. perata but, based on its markedly greater diameter (), hemispherical shape, absence of a central depression, and lack of forms in the ichnoassemblages intermediate with typical B. perata, we retain it as a separate informal ichnospecies. The Torneträsk specimen appears to intersect a large Treptichnus burrow () but otherwise we do not detect any physical relationship with this or other traces in the assemblage. In its size, gross shape, and short tapering burrow form, it has similarities with Guanshanichnus from Cambrian Series 2, Stage 4, strata of Yunnan, China (Weber et al. Citation2012), but it lacks the surficial concentric and radial patterning of that genus. It is likely that it represents the sandy infill of the body position of a large anemone-like organism anchored in the sediment.
Occurrencel
Only recorded in the heterolithic facies near the top of the lower coarsening-upward cycle of the “lower siltstone” interval of the Torneträsk Formation at site 2, Luobákte.
Ichnogenus Laevicyclus Quenstedt, Citation1879Laevicyclus isp. (–L)
Descriptionl
Vertically orientated, cylindrical to funnel-shaped traces that are either partially filled by sandy sediment or hollow with an open funnel-shaped entrance and an iron- or manganese-stained margin (). Where filled, they typically retain a shallow depression on the upper surface of the bed (, J). Weakly defined U-shaped infillings are evident in some burrows (). Funnel-shaped apertures are circular and reach 15 mm in diameter; burrow lengths are typically <20 mm.
Remarksl
These short funnel-shaped burrows are similar to a broad range of forms traditionally included in Monocraterion Torell, Citation1870. However, several authors have pointed out that the type material of that ichnogenus is characterized by surficial radiating burrows around the mouth of the vertical shaft (Jensen Citation1997; Schlirf & Uchman Citation2005; Buatois et al. Citation2017). Hence, many forms assigned previously to Monocraterion have been transferred to Laevicyclus Quenstedt, Citation1879 (Knaust Citation2015). Some examples of this trace fossil approach the morphology of Skolithos, probably owing to the erosional removal of the flared opening of the burrow. The form described here is markedly broader than examples of Monocraterion (=Laevicyclus) from the Cambro-Ordovician of southern Norway (Knaust Citation2004, .7) but is similar to examples from lower Cambrian strata at Trolmen harbour, Västergötland, Sweden, illustrated by Jensen (Citation1997, , D). Laevicyclus is interpreted to be a dwelling burrow of surface- and/or suspension-feeding invertebrates (Knaust Citation2015). It is a typical component of the Skolithos ichnofacies (Knaust Citation2004) generally developed in relatively energetic shallow marine and intertidal environments subjected to regularly shifting sand and episodic erosion, although the examples illustrated herein are small and restricted to just a few sandy beds.
Occurrencel
Moderately common in a few thin sandstone beds of the heterolithic facies near the top of the lower siltstone interval of the Torneträsk Formation at sites 1 and 2, Luobákte.
Ichnogenus Rhizocorallium Zenker, Citation1836cf. Rhizocorallium isp. ()
Descriptionl
Fan-shaped flat or low-relief spreiten of one whorl on the undersurface of a medium-grained sandstone slab (). The spreite consists of weakly developed arcuate ridges and grooves of variable dimensions, the outermost ridge or burrow being slightly more elevated. Gross dimensions are 20 × 14 mm, with individual ridges c. 1 mm wide.
Remarksl
Only a single example of this trace was recorded and its preservation is not ideal. We illustrate the specimen to document one of several rare traces that do not fall clearly within the circumscription of the dominant ichnotaxa recognized within the assemblage. Adhering siltstone and the lack of any apparent burrow extending into the overlying sandstone suggests that this trace was produced by sand casting of a trace developed in the underlying silty sediments.
We acknowledge that identification of this ill-preserved trace is difficult, but we note similarities to Rhizocorallium in its small, roughly U-shaped form, marginal ridge or tube and internal concentric markings. Rhizocorallium generally consists of a simple U-shaped marginal tube with roughly parallel limbs enclosing an area occupied by a spreite (Knaust Citation2013). The most common species, recorded from lower Cambrian to Holocene strata (Rhizocorallium commune Schmid Citation1876) is characterized by sporadically scratched marginal tubes, especially in Mesozoic examples, and an actively filled spreite between the tubes (Knaust Citation2013). The construction of the example from Luobákte is difficult to resolve from the single available specimen and it bears some similarities to Zoophycos, Spirophyton and a few other ichnotaxa depending on interpretation of its three-dimensional architecture. Rhizocorallium generally differs from Zoophycos in having a more U-shaped form and proportionately broader marginal tubes, and was probably generated by several behaviours (feeding, gardening, resting and excreting) of various vermiform groups, such as Sipunculida, Echiurida and Polychaeta (Knaust Citation2013; Zhang et al. Citation2015a, b).
Occurrencel
Only found in the heterolithic facies near the top of the lower siltstone interval of the Torneträsk Formation at site 1, Luobákte.
Ichnogenus Teichichnus Seilacher, Citation1955Teichichnus isp. ()
Figure 9. Trace fossils from the “lower siltstone” interval of the Torneträsk Formation. A. Teichichnus isp.; B. cf. Phycodes palmatus (Hall, Citation1852); C–I. Planolites montanus Richter, Citation1937; J–K. cf. Trichophycus venosus Miller, Citation1879; L. Gyrolithes polonicus Fedonkin, Citation1981. A. Large, isolated, striate, sandstone exichnium; X00010190. B. Low-relief burrows broadening, forking and diverging distally; specimen. C. Very small, smooth, lacriform mounds similar to Lockeia amygdaloides (Seilacher, Citation1953) (in hyporelief: arrowed) feeding into slender straight to irregular, unlined, horizontal burrows filled with silt (Planolites), generally at contrasting orientations to the diagonal streaming lineation; X00010169. D. Slender, straight, unlined, horizontal burrows filled with very fine grained sandstone; X00010182. E. Short, curved, unlined, horizontal burrow filled with very fine grained sandstone; X00010170. F–I. Slender straight to irregular, unlined, horizontal burrows filled with silt (in hyporelief) originating, in some cases, from diminutive lacriform mounds (arrowed); F, G = X00010179; H, I = X00010187. J. Short, longitudinally striate, fine-grained sand-filled burrow with diffuse termini in positive hyporelief; X00010178. K. Short, longitudinally striate, silt-filled burrow in positive hyporelief; X00010182. L. Single whorl of a short, coiled, sand-filled burrow in positive hyporelief; photographed in the field. Scale bars = 10 mm

Descriptionl
Single, large, isolated, sandstone exichnium, straight in plan view, slightly arched in profile view, 123 mm long , 49 mm wide bearing a roughly median major furrow and numerous finer parallel longitudinal striae potentially representing spreiten (). The cast is broken at each extremity, but appears to taper in width slightly at one end.
Remarksl
In its striate patterning and linear and gently curved course, this trace resembles a somewhat smaller specimen from mid-Cambrian strata of China attributed to part of an exichnium of a Teichichnus spreite (Weber et al. Citation2012, ). Orłowski (Citation1989, pl. 17) and Jensen (Citation1997, 1) illustrated additional forms of similar size to the Torneträsk specimen from the lower Cambrian of Poland and south central Sweden as Teichichnus rectus Seilacher, Citation1955 and Teichichnus ovillus Legg, Citation1985, respectively. Gedda (Citation1993) illustrated an additional large, morphologically similar form from middle Cambrian strata of Öland. However, many Teichichnus burrows are substantially smaller (c. 9 mm: Lima & Netto Citation2012) and the arrangement of spreite structures is not well defined on the available specimen so we refrain from assigning it to a formal ichnospecies.
Occurrencel
Jensen and Grant (Citation1998) recorded Teichichnus from the “middle siltstone” and “upper sandstone” intervals of the Torneträsk Formation. The single new specimen was found as a detached block near the lower part of the studied section but may derive from either the upper part of the “lower siltstone” or “middle siltstone” intervals of the Torneträsk Formation at site 1, Luobákte, based on the thick pale brown siltstone preserved adhering to one surface of the structure.
Ichnogenus Phycodes Richter, Citation1850Phycodes palmatus (Hall, Citation1852) ()
Descriptionl
One specimen was identified consisting of divergent burrow lobes, two of which appear to undergo a second rank of division and radiation of lobes (). The structure is large (gross dimensions of 127 × 177 mm), with each increment of radiating lobes measuring c. 45 × 81 mm. The lobes are slightly raised (in hyporelief), are c. 9 mm wide individually, and are infilled by sand that is not noticeably different from the surrounding rock.
Remarksl
The low relief of this specimen suggests that these were relatively shallow excavations (fodinichnia) at the sand-silt interface. The repeated branching is atypical for Phycodes palmatus, but the addition of distal branches was illustrated for an example of this ichnospecies from the upper Torneträsk Formation by Jensen and Grant (Citation1998). The alternative interpretation, that this specimen might represent multiple individual burrow systems that intersected serendipitously, seems less likely owing to the consistency in lobate form and acute divergence angles distally.
Occurrencel
Only found in the heterolithic facies near the top of the “lower siltstone” interval of the Torneträsk Formation at site 1, Luobákte. Phycodes palmatus is, otherwise, a common ichnofossil in Scandinavian early Palaeozoic intertidal to shoreface successions (Clausen & Vilhjálmsson Citation1986; Jensen Citation1997; Jensen & Grant Citation1998; Knaust Citation2004).
Ichnogenus Planolites Nicholson, Citation1873Planolites montanus Richter, Citation1937 (–I)
Descriptionl
Straight to irregular unlined horizontal burrows, <2 mm wide, 5–40 mm long, filled with fine sand or silt that is slightly finer than the surrounding sedimentary rock, preserved in hyporelief on the base of sandstone beds in contact with siltstones (–I). Many of the burrows appear to initiate with a very small (c. 3 × 8 mm), smooth, lacriform mound (in hyporelief; , F, H: arrowed).
Remarksl
These traces are extremely common on the undersides of sandstone beds in the heterolithic upper part of the “lower siltstone” interval of the Torneträsk Formation, but they are easily overlooked owing to their diminutive size, low relief, inconsistent shapes, and presence of similar-sized crenulate structural deformation features at some sites. Vermiform Planolites-type traces (straight to irregular, roughly cylindrical unlined burrows, with mostly structureless fill that is lithologically different from the host material: Pemberton & Frey Citation1982) are common in marine strata throughout the Phanerozoic but can be difficult to differentiate from various similar simple burrows. For example, Jensen (Citation1997) illustrated forms attributed to Cochlichnus isp. and Helminthoidichnites tenuis Fitch, Citation1850 that may overlap in form with the examples of P. montanus from the Torneträsk Formation. Small traces assigned to Belorhaphe from the lower Cambrian of Spain are also similar in size and morphology (Símon López-Villata Citation2019, ). Banks (Citation1970, pl. 1, figs A, B, D) illustrated equivalent unidentified forms (with a lacriform mound extending into a parallel-sided burrow) from lower Cambrian strata of Finnmark, Norway. Knaust (Citation2004, figs. 6.3, 6.4) illustrated essentially identical examples from the Cambro-Ordovician strata of southern Norway, assigning the small lacriform mounds to Lockeia amygdaloides (considered a cubichnion) and the associated fine irregular burrows to Planolites montanus (a repichnion/pascichnion burrow). Such traces may represent the burrowing activities of vermiform animals (Pemberton & Frey Citation1982), very small bivalves (Bromley Citation1996) or similar organisms.
Occurrencel
Common in the heterolithic facies near the top of the “lower siltstone” interval of the Torneträsk Formation at sites 1 and 2, Luobákte.
Ichnogenus Trichophycus Miller & Dyer, Citation1878cf. Trichophycus venosus Miller, Citation1879 (–K)
Descriptionl
Short, raised, straight burrows preserved in positive hyporelief on the undersurfaces of fine-grained sandstone beds excavated into underlying siltstone laminae. The burrows are 42–55 mm long, c. 17 mm wide, and c. 10 mm in maximum thickness but dissipate into the bedding plane at either end. These burrows are characterized by fine but distinctive, parallel, longitudinal striae, 1–2 mm apart (, K).
Remarksl
Although not well preserved, several of these traces in the collection are similar in size and longitudinally striate floor to Trichophycus venosus Miller, Citation1879 (see Jensen Citation1997, ) although they tend to be shorter than the lower Cambrian examples illustrated from Västergötland, central Sweden. They are also generally shorter and more regularly striate than forms attributed to Planolites or Halopoa Torell, Citation1870 (subsequently transferred to Palaeophycus Hall, Citation1847 by Jensen Citation1997), the latter of which is characterized by long, mostly horizontal traces, bearing longitudinal irregular ridges or wrinkles, consisting of several imperfectly overlapping cylindrical probes (Uchman Citation1998). Trichophycus venosus appears to represent the excavation of an epifaunal to shallow infaunal (i.e., semi-infaunal) sediment-feeding invertebrate, the subtle longitudinal striae indicating some regularly disposed ornament on the body surface. Lateral appendage marks or meniscoid backfill structures are not evident.
Occurrencel
Rare in the heterolithic facies near the top of the lower siltstone interval of the Torneträsk Formation at sites 1 and 2, Luobákte.
Ichnogenus Gyrolithes Saporta, Citation1884Gyrolithes polonicus Fedonkin, Citation1981 ()
Descriptionl
Short, coiled, sand-filled burrow of constant (6 mm) width lacking obvious wall structure or ornamentation in positive hyporelief. The entire whorl measures 20 × 29 mm ().
Remarksl
Only a single specimen was identified in the field, preserved in positive hyporelief on the undersurface of a large slab of fine-grained sandstone hosting numerous Treptichnus pedum (Seilacher Citation1955) and cf. Palaeophycus tubularis Hall, Citation1847 burrows. The dense cover of extant lichen on the slab prevented detection of any ornamentation on the burrows. This ichnospecies is relatively common in Cambrian strata elsewhere in Baltica (see Banks Citation1970; Jensen Citation1997; Högström et al. Citation2013 and references therein), but only single specimens were detected in the Torneträsk Formation by Jensen & Grant (Citation1998) and in this study. Such traces are likely to have been produced by polychaetes or arthropods as dwelling and/or feeding burrows (Jensen Citation1997).
Occurrencel
Only found in the heterolithic facies near the top of the lower siltstone interval of the Torneträsk Formation at site 2, Luobákte.
Ichnogenus Treptichnus Miller, Citation1889Treptichnus pedum (Seilacher, Citation1955) (–M)
Figure 10. A–M. Treptichnus pedum (Seilacher Citation1955) Jensen, Citation1997 from the “lower siltstone” interval of the Torneträsk Formation; A–L in positive hyporelief; M in cross-section. A. Slightly curved burrow with weakly divergent segments; X00010192. B. Curved burrow with lobes arrayed on one side of the axis; X00010164. C. Disjointed curved burrow with strongly divergent lateral lobes; X00010164. D. Segmented burrow of commonly aligned lobes; X00010164. E. Curved burrow of commonly aligned lobes; X00010168. F. Broad straight burrow of obliquely oriented lobes; X00010183. G. Slender beaded burrow with a single divergent lateral lobe; X00010210. H. Degraded burrow with irregular course and weakly divergent lateral lobes; X00010176. I. Very slender disjointed burrow forming a narrow loop; X00010186. J. Broadly looped burrow with variably divergent lateral lobes; X00010191. K. Roughly straight burrow, expanding distally to a fan-like array of lobes; X00010189. L. Burrow system of markedly divergent lateral lobes; X00010203. M. Roughly cylindrical silt-filled burrows preserved on the lower side of a thin sandstone bed; X00010203. Scale bars = 10 mm

Descriptionl
Straight () to strongly curved (), rarely sinuous (), or looped (, J), more-or-less horizontal, sand-filled burrow systems comprising a linear beaded burrow or, more commonly, a series of semi-regular segments arranged at acute angles on one (, B, E, K,) or both sides () of the primary burrow direction. Burrows may reach at least 180 mm long and lobes are 1–15 mm wide (), arranged at up to 40° to the main burrow direction (, L); individual lobes are typically <20 mm long and have bluntly tapered termini. Burrows are roughly cylindrical and typically infilled with finer (silty to very fine sandy) material than the surrounding fine-grained sandy sediment ().
Remarksl
Burrows of this type vary greatly in size and orientation, but are distinguished by the short obliquely offset lateral burrow extensions presumably caused by the tracemaker undertaking regular probing of the sediments to one or both sides of the main burrow orientation in search of food. On this basis, Treptichnus pedum is typically interpreted as a fodinichnion produced by a vermiform invertebrate (Buatois et al. Citation1998a; Vannier et al. Citation2010). Kesidis et al. (Citation2019a) recently identified diagnostic transverse and longitudinal markings on T. pedum from the Mickwitzia Sandstone Member of the File Haidar Formation, southern Sweden, that confirm at least some burrows of this type were produced by priapulid scalidophoran worms. The first occurrence of T. pedum has been used as the primary criterion for defining the base of the Cambrian System (Narbonne et al. Citation1987), although recent work has shown that treptichnid burrows of similar form also occur in assemblages containing typical upper Ediacaran fossils (Jensen et al. Citation1998, Citation2000; Gehling et al. Citation2001; Jensen & Runnegar Citation2005; Seilacher et al. Citation2005; Ragozina et al. Citation2008).
Occurrencel
Abundant in the heterolithic facies near the top of the lower siltstone interval of the Torneträsk Formation at sites 1 and 2, Luobákte.
cf. Treptichnus pedum (Seilacher, Citation1955) ()
Figure 11. Trace fossils from the “lower siltstone” interval of the Torneträsk Formation. A. cf. Treptichnus isp.; B–D. Protovirgularia isp.; E. Imbricate-ridged structure; F–I. Radiating and looped burrows. A. Flabellate cluster of horizontal, finger-like, sand-filled, burrow segments, in positive hyporelief; X00010186. B–C. Synthetic mould of straight, bilaterally symmetrical burrow with paired chevron-shaped, lateral lobes; X00010206. D. Bilaterally symmetrical, straight, sand-filled burrow with tapered lateral lobes; X00010212. E. Chevron-shaped array of low, rounded ridges separated by shallow troughs, dissipating into irregular knobs distally, in positive hyporelief; X00010213. F–G. Burrow system in positive hyporelief with (depending on alternative interpretations) a central elevated vertical knob and radiating, branched, recurved, lateral, sand-filled burrows, or serendipitously intersecting burrows; X00010228. H–I. Burrow system with serendipitously intersecting burrows or possibly a central elevated vertical burrow plug with radiating irregularly orientated and sporadically branched horizontal sand-filled burrow arms; X00010230. Scale bars = 10 mm

Descriptionl
Flabellate cluster of prominent finger-like lobes, each consisting of a sand-filled, cylindrical to conical burrow with strongly raised and rounded surface (in hyporelief); lobes 15–30 mm long, 6–12 mm wide, tapering to a blunt apex ().
Remarksl
The single specimen attributable to this form occurs on the same slab as typical Treptichnus pedum burrows, and the two forms may have been generated by the same tracemaker. The flabellate example might represent an extremely condensed coil of T. pedum lobes – a feature seen in some other atypical forms of this ichnotaxon (e.g., Glaesner Citation1969, ); Orłowski Citation1989, , pl. 15, ; Walter et al. Citation1989, ; Seilacher et al. Citation2005, ). Walter et al. (Citation1989, ) also illustrated a specimen under open nomenclature as “Radial hyporelief” that is also notably similar to the specimen of cf. Treptichnus pedum from the Torneträsk Formation. Some forms of Dactyloidites, Dactylophycus and Asterosoma also approach the morphology of the cf. Treptichnus isp. specimen but tend to have a distinctive vertical or oblique source burrow (Muñoz et al. Citation2019) or a more prominently raised central burrow or plug (Joseph et al. Citation2012). We retain this specimen as a separate informal ichnotaxon here, since the trace is incomplete but the segments are relatively large compared to typical T. pedum examples from the Torneträsk Formation, and it remains unclear whether the lobes were part of a tight spiral horizontal trace or a series of radial excavations surrounding a central vertical burrow.
Occurrencel
Only found in the heterolithic facies near the top of the lower siltstone interval of the Torneträsk Formation at site 1, Luobákte.
Ichnogenus Protovirgularia McCoy, Citation1850cf. Protovirgularia isp. (–D)
Descriptionl
Bilaterally symmetrical, straight, sand-filled burrows 42–78 mm long, 16–18 mm wide, represented by a low rounded ridge giving off chevronate to crescentic pairs of lobes at c. 20–30° to the main axis (–D). The lobes, c. 4 mm wide at the central axis, have broadly rounded surfaces and taper to bluntly rounded termini.
Remarksl
Both available specimens, one preserved in positive hyporelief (), the other represented in the collections by a historical polymer mould (, C), lack sufficient details for unequivocal identification, but they appear to have been produced by a bilaterally symmetrical organism with lateral appendages or a probing forked foot (Carmona et al. Citation2010), and may represent a repichnion or fodinichnion. There is no indication of a physical obstacle on the specimens that would produce inorganic chevronate current rills of consistent size along the length of the trace. We acknowledge that additional and better-preserved specimens will be required to confirm affiliation with Protovirgularia, and that some Treptichnus species also approach this morphology in cases where the divergent lobes are tightly paired and subdued in amplitude. Protovirgularia isp. from the lower Cambrian Ocięseki Formation, Poland, is similar to the Torneträsk Formation specimens in size and general form (Orłowski & Żylińska Citation2002). Such traces have been attributed to the motion of cleft-foot molluscs (Seilacher & Seilacher Citation1994; Uchman et al. Citation2005).
Occurrencel
The stratigraphic source of the isolated blocks is uncertain but they probably derive from the heterolithic facies near the top of the lower siltstone interval of the Torneträsk Formation at Luobákte.
Ichnogenus Palaeophycus Hall, Citation1847cf. Palaeophycus tubularis Hall, Citation1847 (–F)
Figure 12. Trace fossils from the “lower siltstone” interval of the Torneträsk Formation. A–F. cf. Palaeophycus tubularis Hall, Citation1847; G–H. Palaeophycus isp.; I–N. Archaeonassa isp.; O. cf. Monomorphichnus isp. A. Densely meshed sand-filled, silt-lined trails in positive hyporelief on the underside of a sandstone bed; X00010229. B. Longitudinally creased portion of a burrow in positive hyporelief; X00010229. C. Markedly curved and apparently branched trails with irregularly ornamented surface in positive hyporelief; X00010216. D. Burrow with acute apparent forking in positive hyporelief; X00010229. E. Longitudinally creased intersecting trails in positive hyporelief; X00010210. F. Longitudinally creased cross-cutting trails in positive hyporelief; X00003532. G. Gently curved, broad, sand-filled trail in positive hyporelief; X00010181. H. Strongly curved, broad, sand-filled trail in positive hyporelief; specimen photographed in the field. I. Slender grooved trail in epirelief; X00010219. J. Deeply grooved trail with raised margins and weak oblique striae on walls; X00010172. K. Strongly curved trail with weakly raised margins in epirelief; X00010187. L. Broadly curved intersecting trails in epirelief; X00010172. M. Synthetic mould of weakly curved trail with weak meniscate ornament; X00010204. N. Broad shallow trail with weakly raised margins in epirelief; X00010164. O. Two sets of parallel ridges and intervening troughs separated by an intervening low ridge; X00010220. Scale bars = 10 mm

Descriptionl
Linear, curvilinear, sinuous or irregular, predominantly bedding-parallel burrows of considerable length (>300 mm) and consistent width (maximum 12 mm), filled by fine-grained sand similar to the host rock, with an ill-defined lining of slightly finer sand or silt (, C, F). Burrows are cylindrical to ribbon-like, have a convex hyporelief at the base of fine-grained sandstone beds, and are generally smooth but a few bear weak longitudinal or oblique lobes or creases (), or contain weakly defined meniscoid backfill features (). Burrows branch very rarely (usually at a very acute angle: , D), but commonly intersect (, F), in some cases forming pervasively bioturbated bedding surfaces of ropey texture (, C).
Remarksl
These are among the most common traces in the lower siltstone unit of the Torneträsk Formation and yet are among the most difficult to assign to any ichnotaxon with consistency. Cambrian examples from various regions that appear to share the morphology of the Torneträsk forms have been assigned previously to such ichnogenera as Palaeophycus Hall, Citation1847 (Bryant & Pickerill Citation1990; Mangano et al. Citation1996; Jensen Citation1997), Halopoa Torell, Citation1870 (Orłowski & Żylińska Citation2002; Knaust Citation2004), Curvolithus Fritsch, Citation1908 (Seilacher et al. Citation2005; McIlroy & Brasier Citation2016), and Didymaulichnus Young, Citation1972 (Crimes & Anderson Citation1985; Walter et al. Citation1989). However, the combination of characters, such as the sporadically lobate base, thin wall lining, weak and sporadic development of meniscate fill of material equivalent to the surrounding matrix, rare branching, and straight to irregular course, is not universally consistent with any of these ichnogenera. The Torneträsk forms also appear to be synonymous with what Jensen & Grant (Citation1998, ) described informally as “winding horizontal burrows” and with those described by Högström et al. (Citation2013, –G) as “trace fossils with a trilobed surface”. These traces are variable in occurrence from isolated trails to densely intersecting ropey aggregations, and although most are parallel to bedding, the hyporelief of some examples shows that they vary in their depth of excavation—a feature illustrated for Palaeophycus tubularis Hall, Citation1847 in some previous studies (e.g., Weber et al. Citation2012, ). The majority of such specimens from the Torneträsk Formation are consistent with the circumscription of P. tubularis in lacking structured infill, ornamented surfaces, and branching. However, a few with creased or lobed ornament () approach Curvolithus isp. (see Buatois et al. Citation1998b, ), Palaeophycus imbricatus (Torell Citation1870) (see Jensen Citation1997), or the “Bure ichnocomplex” of Systra & Jensen (Citation2006, ) in texture. Branching in P. tubularis is atypical but Shahkarami et al. (Citation2017) noted this character in at least a few examples from the Cambrian of Iran. Apparently equivalent burrows have been assigned alternatively to Planolites Nicholson, Citation1873 in some previous studies of Cambrian ichnoassemblages (e.g., Webby Citation1984) but Palaeophycus differs from that ichnogenus by possession of a differentiated wall (Pemberton & Frey Citation1982), although this feature can be very subtle and consistent separation of these taxa remains a matter of continuing debate (Aceñolaza & Yanev Citation2001). Beaconites Vialov, Citation1962 as emended by Keighly & Pickerill (Citation1994) is also similar to Palaeophycus in representing unbranched, mostly horizontal, walled burrows, but the former differs by having an infilling that contrasts with the surrounding lithology and features more prominent meniscate fill (Knaust Citation2004; Gouramanis & McLoughlin Citation2016). We acknowledge that any ichnotaxonomic placement of these burrows will be problematic and we urge a comprehensive revision of the defining characters of simple tubular to ribbon-like burrows.
Occurrencel
Common in the heterolithic facies near the top of the lower siltstone interval of the Torneträsk Formation at sites 1 and 2, Luobákte.
Palaeophycus isp. (, H)
Descriptionl
Large (>250 mm long, 21–43 mm wide), consistently arcuate, hypichnial burrows filled with sand equivalent to the surrounding lithology and lined with a narrow wall of fine sand to silt (, H).
Remarksl
These traces are essentially equivalent in structure and morphology to cf. Palaeophycus tubularis described above but are two to four times larger. They are beset with the same problems of ichnotaxonomic placement as outlined for cf. Palaeophycus tubularis. Both trace types were probably produced by relatively large, vermiform, sediment-feeding invertebrates. Only two, poorly preserved, specimens of Palaeophycus isp. are available and, on this basis, we refrain from segregating them as a formal new ichnospecies.
Occurrencel
Rare; only found in the heterolithic facies near the top of the lower siltstone interval of the Torneträsk Formation at site 1, Luobákte.
Ichnogenus Archaeonassa Fenton & Fenton, Citation1937Archaeonassa isp. (–N)
Descriptionl
Curvilinear, sinuous or looped, horizontal, epichnial furrows with continuous or slightly beaded raised margins (collectively 4–15 mm wide and exceeding 180 mm long: –L). The traces commonly intersect on the bedding plane (). The trace cross-section is typically V-shaped in smaller examples, but flatter () and, in a few cases, has a slightly raised median ridge in larger specimens. Weak oblique () or meniscate striae () are evident on some parts of the furrow wall.
Remarksl
These traces are sparse to moderately common but have few diagnostic features. Bryant and Pickerill (Citation1990) assigned morphological similar traces from the lower Cambrian of Greenland and Norway to Palaeobullia. However, we note that Palaeobullia is considered part of the Scolicia de Quatrefages, Citation1849 complex that is generally considered to have been produced by echinoids (Uchman Citation1995) and, thus, is an untenable assignment for these early Cambrian traces. Walter et al. (Citation1989, ) also assigned very similar winding trails from the lower Cambrian of central Australia to Gordia sp., although examples of that ichnogenus are supposed to be characterized by massive infill (Fillion & Pickerill Citation1990; Wang et al. Citation2009; Hammersburg et al. Citation2018). Psammichnites Torell, Citation1870, as emended by Mángano and Buatois (Citation2002), is distinguished by more prominent transverse ornament and a distinctive median ridge or series of mounds/pits. We tentatively assign the Torneträsk specimens to Archaeonassa Fenton & Fenton, Citation1937 based on their V-shaped cross-sections, flanking ridges and weak oblique striae. Ediacaran Archaeonassa trails from Australia and Russia (Jensen Citation2003), and Carboniferous examples from Ireland (Buckman Citation1994) are especially similar morphologically to the Torneträsk specimens. Kulling (Citation1950, ) illustrated markedly larger but otherwise similar levee-flanked trails from the “upper sandstone” interval of the Torneträsk Formation. Such trails were potentially generated by a range of invertebrate grazers, but small molluscs or mollusc-like animals are especially strong candidates for producers of these traces (Jensen Citation2003).
Occurrencel
Sparse to moderately common in the heterolithic facies near the top of the lower siltstone interval of the Torneträsk Formation at sites 1 and 2, Luobákte.
Ichnogenus Monomorphichnus Crimes, Citation1970cf. Monomorphichnus isp. ().
Descriptionl
A single specimen consists of a series of subdued parallel striae, c. 11 mm long, arranged obliquely on either side of an ill-defined, 2-mm-wide, central ridge (). The striae are 1–2 mm apart, separated by smooth, shallow troughs, and arranged in slightly different orientations on either side of the central ridge. The trace appears to be incomplete and the lateral margins are diffuse.
Remarksl
The origin of this ill-defined structure is ambiguous. It might represent an example of weak streaming lineation or inorganic tool marks, or is possibly indicative of lateral appendage marks flanking a narrow central body trace. However, we note that the low parallel ridges in hyporelief are reminiscent of Monomorphichnus Crimes, Citation1970, especially M. bilinearis Crimes, Citation1970, which commonly has paired or fan-like ridges (Walter et al. Citation1989, 6 C, D; Mangano et al. Citation1996, ; Símon López-Villata Citation2019, ), M. lineatus Crimes et al., Citation1977, which is represented by several parallel ridges (Bryant & Pickerill Citation1990), and M. multilineatus Alpert, Citation1976, occurring as sets of 4–14 parallel ridges (Mangano et al. Citation1996, ; Sharma et al. Citation2018, ). Assignment to any one of these ichnospecies is hampered by poor preservation. Monomorphichnus is regarded as stroke marks of a multidigitate appendage from sideways skimming/raking arthropods (Fillion & Pickerill Citation1990; Jensen Citation1997). At least seven species have been established for this ichnogenus but their consistent differentiation is difficult and the contrasting morphologies probably relate to the number and degree of appendage drag in the sediment (Jensen Citation1997). Some may even have an inorganic origin as lineations or tool marks in uniform-flow environments.
Occurrencel
The stratigraphic source of this isolated specimen is uncertain but probably derives from the heterolithic facies near the top of the lower siltstone interval of the Torneträsk Formation at Luobákte.
Ichnogenera uncertainImbricate-ridged trace ()
Descriptionl
A single trace consisting of an imbricate array (63 mm long and 11 mm wide) of low, rounded ridges, each c. 10 mm long and 3 mm wide, separated by shallow troughs. Ridges dissipate into irregular knobs in the distal part of the trace.
Remarksl
The single available specimen is strongly weathered but is included here for the sake of illustrating the full diversity of trace fossils. In the presumed proximal part the trace, the lateral ridges take on a chevron-like arrangement and approach the morphology of cf. Protovirgularia isp. described above, although the lobes are much narrower. There is no indication of a physical obstacle that would generate leeward divergent ridges abiotically by currents and scouring.
Occurrencel
The stratigraphic source of this isolated specimen is uncertain but probably derives from the heterolithic facies near the top of the lower siltstone interval of the Torneträsk Formation at Luobákte.
Radiating and looped burrows (–I)
Descriptionl
The description of these traces involves a degree of interpretation that can be presented as two alternative morphologies. They represent either sand-filled horizontal burrow systems (preserved in positive hyporelief) consisting of either serendipitously intersecting parallel-sided to beaded, rarely branched, gently curved to strongly recurved burrows; or they could be considered to incorporate a central elevated vertical burrow-fill or plug (6–8 mm in diameter: , I) from which the horizontal burrows emanate (, H). Each horizontal burrow reaches lengths of up to at least 82 mm long and 46 mm wide. The rare cases of apparent branching are at 80–110°. Burrow surfaces are generally smooth and rounded (indicating a cylindrical form) and lack any obvious meniscoid infill or wall lining.
Remarksl
Individually, the horizontal burrows are broadly similar to the form of some Treptichnus pedum and cf. Palaeophycus tubularis Hall, Citation1847 burrows in the same assemblage but the forms described here lack the offset lateral extensions of Treptichnus or the longitudinal creases of cf. P. tubularis. Nevertheless, Palaeophycus-type burrows occur abundantly on the same slab as these radiating burrows and the intersecting and looped forms described here may have been produced by the same organism during more complex feeding behaviour. An alternative interpretation is that these represent genuine radiating burrows originating from a short central vertical tube based on what appears to be a slightly raised plug in the centre of the burrow system and the recurvature of several radial burrows at a consistent distance from the central plug in one specimen (). We draw attention to somewhat similar forms from the lower Cambrian of southern Sweden assigned to Scotolithus mirabilis Linnarsson, Citation1871 (Jensen Citation1997, fig. 58D). We note, however, that the Torneträsk Formation (, H) examples differ by the curvature of the radiating/intersecting burrows. If the described forms consist of burrows radiating from a central shaft, there are also similarities to Radiichnus Karaszewski, Citation1973, but the latter is known only from Jurassic offshore deposits of Poland. Muñoz et al. (Citation2019) recently summarized the diversity and spatio-temporal distribution of radial to rosette-shaped trace fossils noting that this group of ichnofossils occurs as low as the first stage of the Cambrian where they are best represented in shallow to deep shoreface settings.
Occurrencel
Rare; found on isolated slabs presumably derived from the heterolithic facies near the top of the lower siltstone interval of the Torneträsk Formation at site 1, Luobákte.
Ambiguous structures and pseudofossils
Semi-concentric traces ()
Figure 13. Ambiguous structures and pseudofossils from the Torneträsk Formation. A. Semi-concentric traces of ridges and grooves on the underside of a fine-grained sandstone bed; X00010187. B. Isolated large sandstone lobate structure with radiating grooves; X00010174. C. Latex cast of Kullingia concentrica Glaessner in Föyn & Glaessner, Citation1979; cast of specimen SGU30940 held in NRM. D, E. Low-relief discoid structures preserved in ripple troughs on the upper surface of medium-grained ripple-marked sandstone beds of the “lower sandstone” interval; outcrop photos. F. Straight to broadly curved sand-filled desiccation cracks on the underside of a sandstone bed; specimen photographed in the field. G. Short, radiating, sand-filled synaeresis cracks on the underside of a medium-grained sandstone bed; outcrop photo. H. Sand-filled, straight desiccation cracks in siltstone; X00010209. I. Large circular percussion structure with smaller crescentic percussion feature in indurated sandstone; outcrop photo. J. Mass of circular to crescentic percussion structures on bedding plane of indurated sandstone exposed below stream rapids; outcrop photo. Scale bars = 10 mm. A, B, Luobákte site 1, “lower siltstone” interval; C, Luobákte site 1, possibly “middle siltstone” interval; D, E, Luobákte site 2 “lower sandstone” interval; F, H, Luobákte site 1, “lower siltstone” interval; G, Luobákte site 2, “lower siltstone” interval; I, J, Orddajohka rivulet, “upper sandstone” interval

Description
Two structures on the same slab (an irregularly rippled upper surface of fine-grained sandstone) consist of incomplete, slightly raised, circular to semi-circular features 100–150 mm in diameter incorporating successive rings of grooves and ridges (). The grooves and ridges vary in width (reaching up to 15 mm wide) and some ridges are discontinuous, wedging out against neighbouring grooves to produce a semi-concentric pattern.
Remarks
These structures are not as regularly concentric as Kullingia concentrica (previously recorded from the same beds (; Föyn & Glaesner Citation1979; Jensen & Grant Citation1998) but share some features (roughly circular organization, raised central point) that suggest a similar origin. Kullingia concentrica Glaessner in Föyn & Glaessner, Citation1979 was originally interpreted as a probably medusoid cnidarian imprint but may represent scratch marks caused by rotation of some anchored, strap-like object or regularly ornamented tubular sabelliditid-like organism on a soft-sediment surface, perhaps agitated by storm or current action in submarine settings (Jensen et al. Citation2002). The discontinuity of some ridges argues against the semi-concentric traces described here being very regular rotational scratch features. Since neither specimen is complete and a fully circular structure (in contrast to Kullingia) can not be confirmed, these structure may, alternatively, represent small flow or slump structures caused by inter-ripple currents during tidal retreat. Similar semi-concentric structures are evident in the troughs of some modern linguoid ripples on exposed tidal flats (Chakrabarti Citation2005, fig. 18).
Occurrence
Rare in sandstone beds of the heterolithic facies near the top of the lower siltstone interval of the Torneträsk Formation at site 2, Luobákte.
Large lobate grooved structure ()
Description
This structure consists of an isolated, roughly elliptical, sandstone cast, with coarse (2–7 mm wide) grooves originating on one side and radiating, forking, and shallowing to the opposite margin (). The structure is 117 mm long, 96 mm wide and c. 20 mm thick. Indistinct longitudinal striae are evident along some of the grooves.
Remarks
The single specimen of this type was collected from float but likely derives from the thin sandstones in the upper part of the lower siltstone interval. The structure is superficially similar to an excavation mound adjacent to an infaunal burrow. Vaguely similar structures in lower Cambrian strata of Spain have also been attributed to a body fossil imprint of an organism similar to a sand anemone (Símon López-Villalta Citation2019). However, a case could equally be made that this structure from the Torneträsk Formation is a sandstone infilling of a linguoid or lunate ripple trough with drainage rills at its base (compare with, e.g., pl. 30 of Ricci Lucchi Citation1995).
Occurrence
Rare; isolated block presumably derived from a thin sandstone bed near the top of the “lower siltstone” interval of the Torneträsk Formation at site 1, Luobákte.
Flattened elliptical structures (, E)
Description
Elliptical to lobate flattened sand-filled structures within troughs on the upper surfaces of ripple-marked medium- to coarse-grained sandstone (, E). The surfaces of these structures stand about 5 mm above the surrounding bedding plane. They are 75–110 mm long and 67–95 mm wide with a rounded circumference and a margin that is gently curved down to the bedding plane.
Remarks
These simple ill-defined elliptical structures are the only potential trace fossils that we observed in the “lower sandstone” interval of the Torneträsk Formation. A dense cover of extant lichen on both specimens precludes identification of any fine surface features. Although they are superficially similar to some Ediacaran discoid imprints, such as Aspidella (Menon et al. Citation2013; Darroch et al. Citation2016), we detected no central pit or tubercle, and these structures might, alternatively, represent small-scale lobes of sediment that collapsed into troughs from adjacent ripple crests. A disc-shaped structure illustrated by Simón (Citation2017, ) from upper Ediacaran strata of Spain is also superficially similar, though smaller (2 cm diameter), and likewise can not be attributed with confidence to either a sedimentary structure or body imprint.
Occurrence
Rare in the “lower sandstone” interval of the Torneträsk Formation at site 2, Luobákte.
Sand-filled cracks (–H)
Description
These structures are typically represented by long, straight or curvilinear, intersecting, sand-filled, raised ridges on the undersurfaces of sandstone beds resting above siltstones (, H). The infillings have sharply defined margins, V-shaped profiles, and typically form irregular polygons on the undersides of sandstone beds. The structures lack backfill features, are of the same composition as the overlying sandstone, are typically1–6 mm wide, and are variable in length (some exceeding 20 cm). A second category of sand-filled cracks consists of lenticular infillings, 32–75 mm long, 7–15 mm wide, with U-shaped profiles, occurring in isolation or stellate arrangements on the undersurfaces of sandstone beds ().
Remarks
The more slender, elongate, interconnected examples of these ridges represent mostly sand-filled desiccation cracks formed in subaerially exposed siltstone surfaces. A few that enclose more elliptical polygons are similar to Manchuriophycus, earlier considered to represent a body or trace fossil, but now generally considered to constitute subaqueous to subaerial shrinkage cracks, particularly formed in microbial mats (Plummer & Gostin Citation1981; Eriksson et al. Citation2007; Lee et al. Citation2020). The second category is more typical of broad, lenticular, synaeresis cracks that are generated subaqueously by clay deflocculation and lattice contraction caused by changes in the salinity of pore waters (Burst Citation1965; Plummer & Gostin Citation1981; Jensen Citation1997; Knaust Citation2004) or, in some cases, by seismically induced dewatering (Pratt Citation1998). Forms similar to those described here were illustrated from equivalent tidally influenced shoreface deposits of the lower Cambrian Kloftelv Formation (east Greenland) bearing Teichichnus isp. (Jensen et al. Citation2015, ).
Occurrence
Both forms of sand-filled cracks are moderately common in the heterolithic facies near the top of the lower siltstone interval of the Torneträsk Formation at sites 1 and 2, Luobákte.
Percussion structures (, J)
Description
Circular or crescentic structures up to 120 mm diameter preserved on bedding planes of indurated strata in active modern stream channels. The structures are typically defined by a narrow (c. 1 mm wide) marginal indentation in the rock (). In crescentic examples, the indentation is typically in the upstream direction. In a few cases, additional circular crescentic rings may be positioned concentrically within the major ring structure ().
Remarks
These structures are typically found on exposed bedding planes of erosionally resistant (well-indurated) fine-grained sandstones of the Torneträsk Formation, especially immediately downstream of breaks in slope (waterfalls and cascades). Their location, variability in size and shape, and the consistent orientation of crescentic examples leads us to interpret these structures as modern percussion marks on bedding planes caused by fluvial transport of large boulders over waterfalls and rapids. We illustrate these structures here since some isolated examples are somewhat reminiscent of medusoid imprints and might be misinterpreted without careful inspection.
Occurrence
Moderately common to abundant on the exposed upper surfaces of indurated sandstone beds in Orddajohka rivulet, north of Lake Torneträsk.
Biotic diversity
Our reconnaissance survey of the lower part of the Torneträsk Formation reveals a greater diversity of ichnofossils than reported in previous studies, albeit that many of the new forms are represented by a few specimens of equivocal identity (). We hope that the range of traces identified in this study will inspire a broader-scale investigation of ichnofossil diversity within this little-studied unit. We especially highlight the presence of a multitude of small Planolites- and Trichophycus-type burrows and various relatively complex horizontal, Rhizocorallium- and Protovirgularia-like feeding traces that, in association with probable macroalgae, invertebrate body fossils and bacterial mats, indicate a more complex shallow-water ecosystem than previously documented. Owing to limited time available in the field, the size of the slabs, and because the outcrops are only accessible by hiking, we were only able to obtain a modest collection of trace fossils from the Torneträsk Formation. We encourage further high-resolution sampling of this unit to clarify the successive appearances and quantitative representation of ichnofossils through the formation. This should provide a clearer picture of the evolution of biotic diversity and depositional environments through the lower Cambrian succession on the northern margin of Baltica.
Table 1. Body fossils, trace fossils and microbially induced sedimentary structures (MISS) recorded from the Torneträsk Formation near Torneträsk by various authors
Despite its diverse trace fossil record, the Torneträsk Formation is almost devoid of body fossils (), unlike the overlying Grammajukku Formation (Skovsted et al. Citationin press [this volume]). The circular impression Kullingia, originally described as a medusoid Ediacaran body fossil, was later reinterpreted as a scratch-circle tool mark (Jensen et al. Citation2002). In addition, several tubular fossils were described by Moberg (Citation1908) and Kulling (Citation1964). The reported specimens of Hyolithus sp. and Volborthella tenuis were considered dubious by Jensen and Grant (Citation1998). Consequently, the problematic tubular fossils Sabellidites described by Jensen and Grant (Citation1998) from the “lower siltstone” interval (together with an example illustrated herein: , I) and cf. Platysolenites antiquissimus () from the overlying “middle siltstone” interval remain the only credible body fossils (apart from the macroalgae illustrated herein) from the Torneträsk Formation.
Palaeoenvironments
The interbedded sandstones and siltstones near the top of the “lower siltstone” interval of the Torneträsk Formation contain a mix of dwelling and foraging traces and, combined with algal remains, wrinkle structures, and a suite of ripple, synaeresis and desiccation structures, suggest deposition in low-relief and low-energy, shallow subtidal to intertidal settings, which Thelander (Citation1982) attributed to shallowing lagoonal or interdistributary bay environments. Apart from a few thin sandstone beds containing Laevicyclus, there is no evidence of trace fossils typical of high-energy shoreface/foreshore assemblages (e.g., the Skolithos ichnofacies). Assessment of the variation in depositional settings expressed in the Torneträsk Formation must await a future analysis of the vertical and lateral distribution of litho- and bio-facies represented in multiple sections through the unit.
Biomats are inferred to have been common through the Torneträsk Formation based on the presence of wrinkle structures, generally good preservation of traces unaltered by current or wave action, scarcity of surficial trails and tracks, and abundance of presumed shallow under-mat burrowing and dwelling structures (cf. Gehling Citation1999; Gingras et al. Citation2011). Deep and complex burrowing structures are essentially absent from the ichnoassemblage.
Most of the identifiable traces have cosmopolitan distributions, but a few may have palaeogeographic significance. Of these, Mángano and Buatois (Citation2011) argued that the Trichophycus ichnofacies is represented earlier in Baltica and Laurentia compared with Gondwana. Jensen and Grant (Citation1998) found only one specimen of the distinctive spiral trace Gyrolithes in the Torneträsk Formation and we also found just a single specimen. Gyrolithes is a common element in lower Cambrian ichnoassemblages across central and northern Europe (Baltica) and extending into Newfoundland (Systra & Jensen Citation2006). Its scarcity in the Torneträsk Formation might be indicative of the fringe of its geographic range in Baltica or accumulation of unfavourable sedimentary facies in this area. There is, otherwise, little evidence of palaeolatitudinal differentiation of lower Cambrian ichnofossil assemblages, consistent with the findings of Jensen et al. (Citation2013).
Age implications
Geochronological results from detrital zircons are broadly consisted with data obtained by Andresen et al. (Citation2014) from the Dividal Group exposed at various sites along the Scandinavian Caledonides. All but one of the 140 zircon grains in our study yielded LA-ICPMS dates of 953 ± 51 Ma or older, ranging from 3331 ± 27 to 569 ± 19 Ma. These provide various constraints on the provenance of sedimentary grains contributing to the Torneträsk Formation. The single young grain sets the maximum depositional age of the lower Torneträsk Formation to 584 ± 13 Ma (mid-Ediacaran). Peaks in the age profile of detrital zircon grains occur at 2690, 1760, 1200, and 1010 Ma. The oldest population of the detrital zircons probably derives from the oldest rock suite known on the Baltic Shield—the Archaean Pudasjärvi Complex located in Finnish Lapland. These rocks form the Pudasjärvi Granulite Belt (PGB; sensu Nironen et al. Citation2002) and geochronology has revealed that a trondhjemite gneiss from Siurua, Pudasjärvi, “is the oldest primary crustal rock on the Fennoscandian Shield with a minimum conventional U-Pb zircon age of 3.3 Ga” (Mutanen & Huhma Citation2003).
The source of the detrital zircons dated to c. 2.69 Ga is ambiguous. However, a series of roughly coeval Naavala-type grey orthogneisses (dated at 2.69–2.65 Ga) are known to intersect the Kivijärvi gneisses of eastern Baltica (Martin Citation1989) and granitoids of this age occur in the West Troms basement complex (Andresen et al. Citation2014). The detrital zircons yielding an age centred around 1.8 Ga correspond largely to ages available for crystalline complexes formed during the Svecofennian Orogeny (Paleoproterozoic, 2000–1800 Ma; Andersson & Öhlander Citation2004; Andresen et al. Citation2014). The zircon assemblage yielding ages between 1810 and 1650 Ma are probably derived from the Trans-Scandinavian Igneous Belt represented by a series of batholiths extending from northern to southern Sweden (Högdahl et al. Citation2004). Andresen et al. (Citation2014) identified a broad suite of detrital zircons from the Dividal Group with ages similar to the younger populations of detrital grains that we dated to c. 1.2–1.0 Ga. These possibly relate to the Sveconorwegian Orogeny dated to 1140–960 Ma (Andersson et al. Citation2008).
The provenance of the youngest grain, yielding an age of 584 ± 13 Ma, is less certain. A similar small zircon population dated to 570 Ma was reported from the Dividal Group by Andresen et al. (Citation2014). These dates are similar to the age of the Alnö igneous complex of the Sundsvall region dated to 584 ± 7 Ma (Meert et al. Citation2007). However, the zircon dates more likely relate to the Pre-Uralide–Timanide Orogeny, which took place during the Ediacaran–Cambrian transition as a result of the collision between the Timan–Varanger margin of Baltica and parts of the East European Platform (Orlov et al. Citation2011). The crystalline rocks formed during this orogeny have yielded ages of c. 730–510 Ma (Kuznetsov et al. Citation2007, Citation2010; Orlov et al. Citation2011).
The ichnoassemblages from the “lower siltstone” interval of the Torneträsk Formation contain various taxa that are shared with a range of early Cambrian assemblages from Baltica (Banks Citation1970; Jensen Citation1997; Orłowski & Żylińska Citation2002; Systra & Jensen Citation2006; Högström et al. Citation2013), and with areas further south in modern Europe (Crimes et al. Citation1977) and to the west in Greenland (Bryant & Pickerill Citation1990). Some forms, such as Protovirgularia, are notable for making their first appearance in the early Cambrian (Crimes Citation1992).
We found no evidence of body or trace fossils from the “lower sandstone” interval of the Torneträsk Formation. On this basis we can not ascribe a definitive age to this part of the formation. It is at least as old as early Cambrian based on fossil assemblages from overlying strata. Föyn and Glaessner (Citation1979) used the occurrence of Kullingia concentrica in the middle portion of the Torneträsk Formation to argue that these beds were Ediacaran in age. However, if such structures represent tool marks, as suggested by Jensen et al. (Citation2002) and herein, then they are not age diagnostic.
Nielsen & Schovsbo (Citation2011) used sequence stratigraphy to suggest that the base of the Torneträsk Formation is of Dominopolian age (Cambrian Stage 3) and, thus, roughly coeval with the appearance of the first trilobites in southern Sweden and Bornholm. However, this inferred age is at odds with the fossil data, particularly the occurrence of Sabellidites and Platysolenites, which have been used to infer an older Rovnian or Lontovan (Terreneuvian) age for sedimentary sequences in Baltica and Avalonia (Mens & Pirrus Citation1997; Jensen & Grant Citation1998; McIlroy et al. Citation2001; Moczydlowska et al. Citation2014). The notable absence of deep arthropod traces (Rusophycus and Cruziana) in the Torneträsk Formation offers another line of evidence suggesting an older age for the unit, as noted by Jensen & Grant (Citation1998).
Although Rusophycus and Cruziana (that probably represent morphological end-members of deep resting and locomotion/grazing traces: Kesidis et al. Citation2019b) are often referred to the activity of trilobites, their first appearance in the fossil record predates the first trilobite body fossils and the earliest forms were presumably produced by other sclerotized but non-mineralized arthropods (Jensen et al. Citation2013; Kesidis et al. Citation2019b). The first occurrences of Rusophycus are in the Fortunian of Avalonia, China and Laurentia (Jensen et al. Citation2013). In Baltica, the first occurrence of Rusophycus is in the pre-trilobitic (Lontovan, Cambrian Stage 2) lower Breidvika Formation of Finnmark (Banks Citation1970; Högström et al. Citation2013). In southern Scandinavia, Rusophycus is used to define the R. parallelum Zone, which probably correlates with the first appearance of trilobites globally (Jensen et al. Citation2013). In light of the diverse and well-preserved trace fossil assemblage recorded from the “lower siltstone” interval by Jensen & Grant (Citation1998) and herein, the absence of Rusophycus and Cruziana, and the presence of the tubular Platysolenites, the collective fossil evidence is consistent with at least the lower parts of the Torneträsk Formation being older than inferred by Nielsen & Schovsbo (Citation2011). The Torneträsk Formation would thus be of very early Cambrian age, presumably pre-dating the appearance of trilobites (in Cambrian Series 2, Stage 3) and possibly pre-dating the appearance of arthropods (in Terreneuvian, Stage 2). These results are consistent with the conclusions of Jensen & Grant (Citation1998) concerning the Terreneuvian (Fortunian or possibly Stage 2) age of the “lower siltstone” interval. Consequently, the “lower siltstone” interval of the Torneträsk Formation can presumably be correlated with the lower Breidvika Formation of Digermul (Finnmark) rather than the upper Breidvika Formation as suggested by Nielsen & Schovsbo (Citation2011). Correlation of the middle and upper parts of the Torneträsk Formation remains equivocal in the absence of additional biostratigraphic controls; hence, the biozone correlations inferred for this portion of the unit by Axheimer et al. (Citation2007; ) should be considered tentative interpretations.
Trilobites, brachiopods and Small Shelly Fossils from the overlying Grammajukku Formation indicate a Vergalian–Rausvian (Cambrian Stage 4) age (Axheimer et al. Citation2007; Skovsted et al. Citationin press [this volume]). Although the upper part of the Torneträsk Formation (“upper sandstone” interval) has yielded no age-diagnostic fossils, the age discrepancy between the possible Terreneuvian age of the “lower siltstone” interval and the much younger Grammajukku Formation, suggests that the phosphorite-bearing conglomerate at the base of the latter defines a significant depositional hiatus.
Conclusions
This reconnaissance survey of the fossil biota of the Torneträsk Formation reveals a greater diversity of both body and trace fossils than recorded previously from this unit. We highlight this unit as a potential source of additional lower Cambrian fossils that could provide insights into the biotic evolution and variation of depositional settings in northern Baltica during the earliest Cambrian. Owing to an absence of trace or body fossils, we can not exclude an Ediacaran age for the basal sandstone-dominated interval of the Torneträsk Formation. These beds have a minimum age of early Cambrian based on trace- and body-fossil suites in overlying strata. The lower siltstone interval of the Torneträsk Formation contains a broad suite of mostly simple pascichnia, fodinichnia and domichnia burrows and trails but lacks deep arthropod trackways. Furthermore, cyanobacterial mats occur throughout the succession, indicating an age prior to or during the development of early invertebrate grazing communities. This, together with maximum geochronological constraints of 584 ± 13 Ma from LA-ICPMS-dating of a detrital zircon, favours a very early Terreneuvian, possibly Fortunian, Cambrian age for the “lower siltstone” interval of the Torneträsk Formation. The trace fossil assemblages of this interval are generally indicative of low-energy, shallow shoreface to intertidal settings. The upper part of the Torneträsk Formation contains only a few invertebrate trace and body fossils that are not strongly age diagnostic. The coarser nature of the sedimentary facies in this interval suggests deposition in moderate-energy shoreface settings.
Supplementary_Online_File_3.xls
Download MS Excel (1 MB)Supplementary_Online_File_2.pdf
Download PDF (2.4 MB)Supplementary_Online_File_1.pdf
Download PDF (158.9 KB)Acknowledgments
We thank the staff at the Abisko Scientific Research Station for valuable help and advice during fieldwork in August 2020. This work was supported by funding from the Swedish Museum of Natural History and the Swedish Research Council (VR grant number 2018-04527 to S.M.; VR grant number 2019-04061 to VV; VR grant number 2016-04610 to CBS). We thank Sören Jensen and one anonymous reviewer for their insightful comments that improved the manuscript.
Disclosure statement
No potential conflict of interest was reported by the author(s).
Supplementary materials
Supplemental data for this article can be accessed here.
Additional information
Funding
Related Research Data
References
- Aceñolaza, F.G. & Yanev, S.N., 2001: El Ordovícico del sector occidental de Stara Planina (Montes Balcanes), Bulgaria: Icnofósiles e implicaciones paleobiogeográficas. Revista Del Museo Argentino De Ciencias Naturales, N.S 3(1), 55–72. doi:https://doi.org/10.22179/REVMACN.3.110
- Ahlberg, P., 1980a: Early Cambrian trilobites from Mount Luopakte northenSweden. Sveriges Geologiska Undersökning, Series C 765, 1–12.
- Ahlberg, P., 1980b: Early Cambrian trilobites from northern Scandinavia. Norsk Geologisk Tidsskrift 60, 153–159.
- Ahlberg, P., 1985: Lower Cambrian trilobite faunas from the Scandinavian Caledonides - a review. In D.G. Gee & B.A. Sturt (eds.): The Caledonide Orogen - Scandinavia and related areas, 339–346. Wiley & Sons, Chichester.
- Alpert, S.P., 1973: Bergaueria Prantl (Cambrian and Ordovician), a probable actinian trace fossil. Journal of Paleontology 47, 919–924. https://www.jstor.org/stable/1303074
- Alpert, S.P., 1976: Planolites and Skolithos from the Upper Precambrian–Lower Cambrian White-Inyo mountains, California. Journal of Paleontology 49, 508–521.
- Andersson, U.B. & Öhlander, B., 2004: The late Svecofennian magmatism. In K. Högdahl, U.B. Andersson & O. Eklund (eds.): The Transscandinavian Igneous Belt (TIB) in Sweden: a review of its character and evolution. Helsinki: Geological Survey of Finland, Special Paper. Vol. 37, 102–104.
- Andersson, J., Bingen, B., Cornell, D., Johansson, L. & Möller, C., 2008: The Sveconorwegian Orogen of southern Scandinavia: setting, petrology and geochronology of polymetamorphic high-grade terranes. 33 IGC excursion guidebook 51, August 2–5. International Geological Congress, Oslo, 1–83.
- Andresen, A., Agyei-Dwarko, N.Y., Kristoffersen, M. & Hanken, N.-M. 2014: A Timanian foreland basin setting for the late Neoproterozoic–Early Palaeozoic cover sequences (Dividal Group) of northeastern Baltica. In F. Corfu, D. Gasser & D.M. Chew (eds.): New Perspectives on the Caledonides of Scandinavia and related areas, 157–175. Geological Society, London. Special Publications 390. doi:https://doi.org/10.1144/SP390.29
- Axheimer, N., Ahlberg, P. & Cederström, P., 2007: A new lower Cambrian eodiscoid trilobite fauna from Swedish Lapland and its implications for intercontinental correlation. Geological Magazine 144(6), 953–961. doi:https://doi.org/10.1017/S0016756807003597
- Banks, N.L., 1970: Trace fossils from the late Precambrian and lower Cambrian of Finnmark, Norway. In T.P. Crimes & J.C. Harper (eds.): Trace Fossils. Geological journal special issue 3, 19–34. Seel House Press, Liverpool.
- Brocks, J., Love, G., Summons, R., Knoll, A.H., Logan, G.A. & Bowden, S.A., 2005: Biomarker evidence for green and purple sulphur bacteria in a stratified Palaeoproterozoic sea. Nature 437(7060), 866–870. doi:https://doi.org/10.1038/nature04068
- Bromley, R.G., 1996. Trace Fossils. Biology, taphonomy and applications, Second ed., 361. Chapman and Hall, London.
- Bryant, I.D. & Pickerill, R.K., 1990: Lower Cambrian trace fossils from the Buen Formation of central North Greenland: preliminary observations. Grønlands Geologiske Undersøgelse, Rapport 147, 44–62. doi:https://doi.org/10.34194/rapggu.v147.8106
- Buatois, L.A., Mángano, M.G., Maples, C.G. & Lanier, W.P., 1998a: Ichnology of an Upper Carboniferous fluvio-estuarine paleovalley—The Tonganoxie Sandstone, Buildex Quarry, eastern Kansas. Journal of Paleontology 72(1), 152–180. doi:https://doi.org/10.1017/S0022336000024094
- Buatois, L.A., Mángano, M.G., Mikuláš, R. & Maples, C.G., 1998b: The ichnogenus Curvolithus revisited. Journal of Paleontology 72(4), 758–769. http://www.jstor.org/stable/1306701
- Buatois, L.A., Wisshak, M., Wilson, M.A. & Mángano, M.G., 2017: Categories of architectural designs in trace fossils: a measure of ichnodisparity. Earth Science Reviews 164, 102–181. doi:https://doi.org/10.1016/j.earscirev.2016.08.009
- Buckman, J.O., 1994: Archaeonassa Fenton and Fenton 1937 reviewed. Ichnos 3(3), 185–192. doi:https://doi.org/10.1080/10420949409386387
- Burst, J.F., 1965: Subaqueously formed shrinkage cracks inclay. Journal of Sedimentary Petrology 35, 348–353. doi:https://doi.org/10.1306/74D71271-2B21-11D7-8648000102C1865D
- Bykova, N., LoDuca, S.T., Ye, Q., Marusin, V., Grazhdankin, D. & Xiao, S., 2020: Seaweeds through time: morphological and ecological analysis of Proterozoic and early Paleozoic benthic macroalgae. Precambrian Research 350, 105875. doi:https://doi.org/10.1016/j.precamres.2020.105875
- Carmona, N.B., Mángano, M.G., Buatois, L.A. & Ponce, J.J., 2010: taphonomy and paleoecology of the bivalve trace fossil Protovirgularia in deltaic heterolithic facies of the Miocene Chenque Formation, Patagonia, Argentina. Journal of Paleontology 84, 730–738. doi:https://doi.org/10.1017/S0022336000058431
- Caullery, M., 1914: Sur les Siboglinidae, type nouveau d’invertébrés receuillis par l’expédition du Siboga. Comptes rendus hebdomadaires des séances de l’Académie des sciences 158, 2014–2017.
- Cavalier-Smith, T., 1995: Zooflagellate phylogeny and classification. Cytology 37, 1010–1029.
- Chakrabarti, A., 2005: Sedimentary structures of tidal flats: a journey from coast to inner estuarine region of eastern India. Journal of Earth Systems Science 114(3), 353–368. doi:https://doi.org/10.1007/BF02702954
- Clausen, C.K. & Vilhjálmsson, M., 1986: Substrate control of lower Cambrian trace fossils from Bornholm, Denmark. Palaeogeography, Palaeoclimatology, Palaeoecology 56(1–2), 51–68. doi:https://doi.org/10.1016/0031-0182(86)90107-0
- Crimes, T.P., 1970: Trilobite tracks and other trace fossils from the Upper Cambrian of North Wales. Geological Journal 7, 47–68. doi:https://doi.org/10.1002/gj.3350070104
- Crimes, T.P., Legg, I., Marcos, A. & Arboleya, M., 1977: ?Late Precambrian – Lower Cambrian trace fossils from Spain. In T.P. Crimes & J.C. Harper (eds.): Trace Fossils 2, 91–138. Geological Journal Special Issue 9. Seel House Press, Liverpool.
- Crimes, T.P., 1992: Changes in the trace fossil biota across theProterozoic-Phanerozoic boundary. Journal of the Geological Society of London 149(4), 631–646. doi:https://doi.org/10.1144/gsjgs.149.4.0637
- Crimes, T.P. & Anderson, M.M., 1985: Trace fossils from late Precambrian–early Cambrian strata of southeastern Newfoundland (Canada): temporal and environmental implications. Journal of Paleontology 59, 310–343. https://www.jstor.org/stable/1305031
- Darroch, A.A.F., Boag, T.H., Racicot, R.A., Tweed, S., Mason, S.J., Erwin, D.H. & Laflamme, M., 2016: A mixed Ediacaran-metazoan assemblage from the Zaris Sub-basin, Namibia. Palaeogeography, Palaeoclimatology, Palaeoecology 459, 198–208. doi:https://doi.org/10.1016/j.palaeo.2016.07.003
- de Quatrefages, M.A., 1849: Note sur la Scolicia prisca (A. De. Q.), annelide fossile de la Craie. Annales des Sciences Naturelles, Zoologie, Sèrie 3, Zoologie 12, 265–266.
- Eichwald, E., 1860: Lethaea rossica ou palaeontologie de la Russie, 1657. D.E. Schweizerbart, Stuttgart.
- Eriksson, P.G., Porada, H., Banerjee, S., Bouougri, E., Sarkar, S. & Bumby, A.J., 2007: Mat-destruction features. In J. Schieber, P.K. Bose, P.G. Eriksson, S. Banerjee, S. Sarkar, W. Altermann & O. Catuneanu (eds.): Atlas of Microbial Mat Features Preserved within the Clastic Rock Record. Atlases in Geoscience 2, 76–105. Elsevier, New York.
- Fedonkin, M.A., 1981: Belomorskaja biotavenda. Trudy Akademii Nauk SSSR 342, 100.
- Fenton, C.L. & Fenton, M.A., 1937: Archaeonassa, Cambrian snail trails and burrows. American Midland Naturalist 18, 454–456. doi:https://doi.org/10.2307/2420587
- Fillion, D. & Pickerill, R.K., 1990: Ichnology of the Upper Cambrian? To Lower Ordovician Bell Island and Wabana groups of eastern Newfoundland. Palaeontographica Canadiana 7, 1–119.
- Fitch, A., 1850: A historical, topographical and agricultural survey of the County of Washington. Transactions New York Agricultural Society 9, 753–944.
- Føyn, S., 1967: Dividal-gruppen («Hyolithus-sonen») i Finnmark og dens forhold til de eokambrisk–kambriske formasjoner. Norges Geologiske Un Dersøkelse 249, 1–84.
- Föyn, S. & Glaessner, M.F., 1979: Platysolenites, other animal fossils, and the Precambian-Cambrian transition in Norway. Norsk Geologisk Tidsskrift 59, 2–6.
- Fritsch, A., 1908: Problematica silurica, 28. C. Bellman, Prague.
- Gedda, B., 1993: Trace fossils and palaeoenvironments in the Middle Cambrian at Äleklinta, Öland, Sweden. Examensarbeten I Geologi Vid Lunds Universitet 49, 1–11.
- Gehling, J.G., 1999: Microbial mats in terminal Proterozoic siliciclastics: Ediacaran death masks. Palaios 14(1), 40–57. doi:https://doi.org/10.2307/3515360
- Gehling, J.G., Jensen, S., Droser, M.L., Myrow, P.M. & Narbonne, G.M., 2001: Burrowing below the basal Cambrian GSSP, Fortune Head, Newfoundland. Geological Magazine 138(2), 213–218. doi:https://doi.org/10.1017/S001675680100509X
- Georgieva, M.N., Little, C.T., Watson, J.S., Sephton, M.A., Ball, A.D. & Glover, A.G., 2019: Identification of fossil worm tubes from Phanerozoic hydrothermal vents and cold seeps. Journal of Systematic Palaeontology 17, 287–329. doi:https://doi.org/10.1080/14772019.2017.1412362
- Gingras, M.K., Waldron, J.W.F., White, C.E. & Barr, S.M., 2011: The evolutionary significance of a lower Cambrian trace-fossil assemblage from the Meguma terrane, Nova Scotia. Canadian Journal of Earth Sciences 48, 71–85. doi:https://doi.org/10.1139/E10-086
- Glaessner, M.F., 1969: Trace fossils from the Precambrian and basalCambrian. Lethaia 2(4), 369–393. doi:https://doi.org/10.1111/j.1502-3931.1969.tb01258.x
- Gnilovskaya, M.B., Istchenko, A.A., Kolshnikov, C.M., Korenchuk, L.V. & Udalstov, A.P., 1988: Vendotaenids of the East European platform, 143. Leningrad: Nauka.
- Gouramanis, C. & McLoughlin, S., 2016: Siluro-Devonian trace fossils from the Mereenie Sandstone, Kings Canyon, Watarrka National Park, Amadeus Basin, Northern Territory, Australia. Alcheringa 40, 118–128. doi:https://doi.org/10.1080/03115518.2016.1099957
- Grube, A.E., 1850: Die Familien der Anneliden. Archiv Für Naturgeschichte, Berlin 16(1), 249–364.
- Hagadorn, J.W. & Bottjer, D.J., 1997: Wrinkle structures: microbially mediated sedimentary structures common in subtidal siliciclastic settings at the Proterozoic–Phanerozoic transition. Geology 25, 1047–1050.
- Hall, J., 1847: Palaeontology of New York, Volume I. Containing descriptions of the organic remains of the Lower Division of the New York System. (equivalent of the Lower Silurian rocks of Europe), 338. C. Van Benthuysen, Albany.
- Hall, J., 1852: Palaeontology of New York, Volume II. Containing descriptions of the organic remains of the Lower Middle Division of the New York System (equivalent in part to the Middle Silurian rocks of Europe), 362. C. Van Benlhuysen, Albany.
- Hammersburg, S.R., Hasiotis, S.T. & Robinson, R.A., 2018: Ichnotaxonomy of the Cambrian Spence Shale Member of the Langston Formation, Wellsville Mountains, northern Utah, USA. University of Kansas Paleontological Contributions 20, 1–66. doi:https://doi.org/10.17161/1808.26428
- Heyl, K., Woelfel, J., Schumann, R. & Karsten, U., 2010: Microbial mats from wind flats of the southern Baltic Sea. In J. Seckbach & A. Oren (eds.): Microbial Mats. Cellular origin, life in extreme habitats and astrobiology, Vol. 14, 301–319. Springer, Dordrecht. doi:https://doi.org/10.1007/978-90-481-3799-2_16
- Högdahl, K., Andersson, U.B. & Eklund, O., 2004: The Transscandinavian Igneous Belt (TIB) in Sweden: a review of its character and evolution (foreword). Geological Survey of Finland Special Paper 37, 3–4.
- Högström, A., Jensen, S., Palacios, T. & Ebbestad, J.O.R., 2013: New information on the Ediacaran-Cambrian transition in the Vestertana Group, Finnmark, northern Norway, from trace fossils and organic-walled microfossils. Norwegian Journal of Geology 93, 95–106.
- Hybertsen, F., 2017: Distribution of Sabellidites (Annelida?) in the basal Cambrian of the Digermulen Peninsula, Arctic Norway. Examensarbete Vid Institutionen För Geovetenskaper, Degree Project at the Department of Earth Sciences, Uppsala University 396, 1–38.
- Jensen, S., 1997: Trace fossils from the Lower Cambrian Mickwitzia Sandstone, south-central Sweden. Fossils and Strata 42, 1–110.
- Jensen, S., 2003: The Proterozoic and earliest Cambrian trace fossil record; patterns, problems and perspectives. Integrative and Comparative Biology 43(1), 219–228. doi:https://doi.org/10.1093/icb/43.1.219
- Jensen, S., Buatois, L.A. & Mángano, M.G., 2013: Testing for palaeogeographical patterns in the distribution of Cambrian trace fossils. Geological Society, London, Memoirs 38, 45–58. doi:https://doi.org/10.1144/M38.5
- Jensen, S., Gehling, J.G. & Droser, M.L., 1998: Ediacara-type fossils in Cambrian sediments. Nature 393(6685), 567–569. doi:https://doi.org/10.1038/31215
- Jensen, S., Gehling, J.G., Droser, M.L. & Grant, S.W.F., 2002: A scratch circle origin for the medusoid fossil Kullingia. Lethaia 35, 291–299. doi:https://doi.org/10.1111/j.1502-3931.2002.tb00089.x
- Jensen, S. & Grant, S.W.F., 1998: Trace fossils from the Dividalen Group, northern Sweden: implications for Early Cambrian biostratigraphy of Baltica. Norsk Geologisk Tidsskrift 78, 305–317.
- Jensen, S., Harper, D.A.T. & Stouge, S., 2015: Trace fossils from the lower Cambrian Kløftelv Formation, Ella Ø, North-East Greenland. GFF 138, 369–376. doi:https://doi.org/10.1080/11035897.2015.1076029
- Jensen, S. & Runnegar, B.N., 2005: A complex trace fossil from the Spitskop Member (terminal Ediacaran–? Lower Cambrian) of southern Namibia. Geological Magazine 142, 561–569. doi. https://doi.org/10.1017/S0016756805000853
- Jensen, S., Saylor, B.Z., Gehling, J.G. & Germs, G.J.B., 2000: Complex trace fossils from the terminal Proterozoic of Namibia. Geology 28, 143–146.
- Joseph, J.K., Patel, S.J. & Bhatt, N.Y., 2012: Trace fossil assemblages in mixed siliciclastic-carbonate sediments of the Kaladongar Formation (Middle Jurassic), Patcham Island, Kachchh, Western India. Journal Geological Society of India 80, 189–214. doi:https://doi.org/10.1007/s12594-012-0131-y
- Karaszewski, W., 1973: A star-like trace fossil in the Jurassic of the Holy Cross Mts. Bulletin de L’Académie Polonaise des Sciences (Séries des Sciences de la Terre) 21, 157–160.
- Keighley, D.G. & Pickerill, R.K., 1994: The ichnogenus Beaconites and its distinction from Ancorichnus and Taenidium. Palaeontology 37, 305–337.
- Kesidis, G., Budd, G.E. & Jensen, S., 2019b: An intermittent mode of formation for the trace fossil Cruziana as a serial repetition of Rusophycus: the case of Cruziana tenella (Linnarsson 1871). Lethaia 52, 133–148. doi:https://doi.org/10.1111/let.12303
- Kesidis, G., Slater, B.J., Jensen, S. & Budd, G.E., 2019a: Caught in the act: priapulid burrowers in early Cambrian substrates. Proceedings of the Royal Society B 286(1894), 20182505. doi:https://doi.org/10.1098/rspb.2018.2505
- Kjellman, F.R., 1891: Phaeophyceae (Fucoideae). In A. Engler & K. Prantl (eds.): Die natürlichen Pflanzenfamilien. 1, 176–192. Wilhelm Engelmann, Leipzig.
- Knaust, D., 2004: Cambro-Ordovician trace fossils from the SW-Norwegian Caledonides. Geological Journal 39(1), 1–24. doi:https://doi.org/10.1002/gj.941
- Knaust, D., 2013: The ichnogenus Rhizocorallium: classification, trace makers, palaeoenvironments and evolution. Earth-Science Reviews 126, 1–47. doi:https://doi.org/10.1016/j.earscirev.2013.04.007
- Knaust, D., 2015: Siphonichnidae (new ichnofamily) attributed to the burrowing activity of bivalves: ichnotaxonomy, behaviour and palaeoenvironmental implications. Earth-Science Reviews 150, 497–519. doi:https://doi.org/10.1016/j.earscirev.2015.07.014
- Kulling, O., 1930: Studier över den kaledoniska fjällkedjans stratigrafi och tektonik inom norra delen av svenska Lapland (Deutsche zusammenf.). Geologiska Föreningens i Stockholm Förhandlingar 52, 647–673. doi:https://doi.org/10.1080/11035893009448733
- Kulling, O., 1950: Berggrunden söder om Torneträsk. Geologiska Föreningens i Stockholm Förhandlingar 72, 454–471. doi:https://doi.org/10.1080/11035895009455331
- Kulling, O., 1960: Route descriptions. In O. Kulling & P. Geijer (eds.): The Caledonian Mountain Chain in the Torneträsk–Ofoten area, northern Scandinavia: the Kiruna Iron Ore Field, Swedish Lapland. Guide to excursions Nos. A25 and C20, 50–76. International Geological Congress XXI Session. The Geological Survey of Sweden, Uppsala.
- Kulling, O., 1964: Översikt över Norra Norrbottensfjällens Kaledonberggrund. Sveriges Geologiska Undersökning 19, 1–166.
- Kulling, O., 1972: The Autochthonous. In T. Strand & O. Kulling (eds.): Scandinavian Caledonides, 162–177. Wiley-Interscience, London, New York, Sydney, Toronto.
- Kuznetsov, N.B., Natapov, L.M., Belousova, E.A., O’Reilly, S.Y. & Griffin, W.L., 2010: Geochronological, geochemical and isotopic study of detrital zircon suites from late Neoproterozoic clastic strata along the NE margin of the East European Craton: implications for plate tectonic models. Gondwana Research 17, 583–601. doi:https://doi.org/10.1016/j.gr.2009.08.005
- Kuznetsov, N.B., Soboleva, A.A., Udoratina, O.V., Andreichev, V.L. & Hertseva, M.V., 2007: Pre-Ordovician tectonic evolution and volcanoplutonic associations of the Timanides and northern pre-Uralides, northeast part of the East European Craton. Gondwana Research 12, 305–323. doi:https://doi.org/10.1016/j.gr.2006.10.021
- Lamarck, J.-B.D., 1802: Discours d’Ouverture, Prononcé le 27 floréal An 10, au Muséum dHistoire naturelle. Recherches sur l’organisation des corps vivans. Bulletin Scientifique De La France Et De La Belgique. (5th Series) 40, 483–517.
- Landing, E., Myrow, P., Benus, A.P. & Narbonne, G.M., 1989: Appearance of the oldest skeletalized faunas in southeastern Newfoundland. Journal of Paleontology 63, 739–769. doi:https://doi.org/10.1017/S0022336000036465
- Lee, D.-C., Byun, U.H., Kwon, Y.K., Keehm, Y. & Jeong, G.Y., 2020: Manchuriophycus-like elliptical cracks in thin mudstones intercalated with lacustrine sandstone: intrastratal crack formation in water-saturated sediments. Sedimentary Geology 408, 105769. doi:https://doi.org/10.1016/j.sedgeo.2020.105769
- Legg, I.C., 1985: Trace fossils from a Middle Cambrian deltaic sequence, north Spain. In H.A. Curran (ed.): Biogenic Structures: their use in interpreting depositional environments, Vol. 35, 151–165. Society of Economic Paleontologists and Mineralogists Special Publication. doi:https://doi.org/10.2110/pec.85.35.0151
- Lima, J.H.D. & Netto, R.G., 2012: Trace fossils from the Permian Teresina Formation at Cerro Caveiras (S Brazil). Revista Brasileira De Paleontologia 15, 5–22. doi:https://doi.org/10.4072/rbp.2012.1.01
- Lin, X., Peng, J., Du, L., Yan, J. & Hou, Z., 2017: Characterization of the microbial dolomite of the upper Sinian Dengying Formation in the Hanyuan area of Sichuan Province, China. Acta Geologica Sinica 91, 806–821. doi:https://doi.org/10.1111/1755-6724.13311
- Linnarsson, I.G.O., 1871: Geognostiska och paleontologiska iakttagelser ofver Eophytonsandstenen i Västergötland. Kongliga Svenska Vetenskaps-Akademiens Handlingar 9(7), 1–19.
- LoDuca, S.T., Bykova, N., Wu, M., Xiao, S. & Zhao, Y., 2017: Seaweed morphology and ecology during the great animal diversification events of the early Paleozoic: a tale of two floras. Geobiology 15(4), 588–616. doi:https://doi.org/10.1111/gbi.12244
- Ludwig, K.R., 2003: User’s Manual for Isoplot 3.00, 70. Berkeley Geochronology Center, Berkeley, CA.
- Mángano, M.G. & Buatois, L.A., 2002: Carboniferous Psammichnites: systematic re-evaluation, taphonomy and autecology. Ichnos 9, 1–22. doi:https://doi.org/10.1080/10420940190034175
- Mángano, M.G. & Buatois, L.A., 2011: Timing of infaunalization in shallow-marine early Paleozoic communities in Gondwanan settings: discriminating evolutionary and paleogeographic controls. Palaeontologia Electronica 14(2), 1–21. palaeo-electronica.org/2011_2/187/index.html
- Mángano, M.G., Buatois, L.A. & Aceñolaza, G.F., 1996: Trace fossils and sedimentary facies from a Late Cambrian–Early Ordovician tide‐dominated shelf (Santa Rosita Formation, northwest Argentina): implications for ichnofacies models of shallow marine successions. Ichnos 5, 53–88. doi:https://doi.org/10.1080/10420949609386406
- Manten, A.A., 1966: Some problematic shallow-marine structures. Marine Geology 4(3), 227–232. doi:https://doi.org/10.1016/0025-3227(66)90023-5
- Martin, H., 1989: Archaean chronology in the eastern part of the Baltic Shield: a synthesis. Precambrian Research 43(1–2), 63–77. doi:https://doi.org/10.1016/0301-9268(89)90005-3
- McCoy, F., 1850: On some genera and species of Silurian Radiata in the collection of the University of Cambridge. Annals and Magazine of Natural History (Series 2) 6, 270–290.
- McIlroy, D. & Brasier, M.D. 2016: Ichnological evidence for the Cambrian explosion in the Ediacaran to Cambrian succession of Tanafjord, Finnmark, northern Norway. In A.T. Brasier, D. McIlroy & N. McLoughlin (eds.): Earth System Evolution and Early Life: a Celebration of the Work of Martin Brasier, 351–368. Geological Society, London. Special Publications 448. doi:https://doi.org/10.1144/SP448.7
- McIlroy, D., Green, O.R. & Brasier, M.D., 2001: Palaeobiology and evolution of the earliest agglutinated Foraminifera: Platysolenites, Spirosolenites and related forms. Lethaia 34(1), 13–29. doi:https://doi.org/10.1080/002411601300068170
- Meert, J.G., Walderhaug, H.J., Torsvik, T.H. & Hendriks, B.W.H., 2007: Age and paleomagnetic signature of the Alnø carbonatite complex (NE Sweden): additional controversy for the Neoproterozoic paleoposition of Baltica. Precambrian Research 154, 159–174. doi:https://doi.org/10.1016/j.precamres.2006.12.008
- Menon, L.R., McIlroy, D. & Brasier, M.D., 2013: Evidence forCnidaria-like behavior in ca. 560 Ma Ediacaran Aspidella. Geology 41(8), 895–899. doi:https://doi.org/10.1130/G34424.1
- Mens, K. & Pirrus, E., 1997: Cambrian. In A. Raukas & A. Teedumäe (eds.): Geology and Mineral Resources of Estonia, 436. Estonian Academy Publishers, Tallinn.
- Mens, K., Bergström, J. & Lendzion, K., 1990: The Cambrian system on the East European platform. International Union of Geological Sciences, Publication 25, 1–74.
- Miller, S.A., 1879: Description of twelve new fossil species, and remarks upon others. Journal of the Cincinnati Society of Natural History 2, 104–118.
- Miller, S.A., 1889: North American Geology and Palaeontology for the use of Amateurs, Students, and Scientists, 718. Press of Western Methodist Book Concern, Cincinnati.
- Miller, S.A. & Dyer, C.B., 1878: Contribution to Paleontology. Journal of the Cincinnati Society of Natural History 1, 24–39.
- Moberg, J.C., 1908: Bidrag till kännedomen om de kambriska lagren vid Torneträsk. Sveriges Geologiska Undersokning C2I2, 1–30.
- Moczydłowska, M., Westall, F. & Foucher, F., 2014: Microstructure and biogeochemistry of the organically preserved Ediacaran Metazoan Sabellidites. Journal of Paleontology 88(2), 224–239. doi:https://doi.org/10.1666/13-003
- Muñoz, D.F., Mangano, M.G. & Buatois, L.A., 2019: Unravelling Phanerozoic evolution of radial to rosette trace fossils. Lethaia 52(3), 350–369. doi:https://doi.org/10.1111/let.12317
- Mutanen, T. & Huhma, H., 2003: The 3.5 Ga Siurua trondhjemite gneiss in the Archaean Pudasjärvi Granulite Belt, northern Finland. Bulletin of the Geological Society of Finland 75(1–2), 51–68. doi:https://doi.org/10.17741/BGSF/75.1-2.004
- Narbonne, G.M., Myrow, P.M., Landing, E. & Anderson, M.M., 1987: A candidate stratotype for the Precambrian–Cambrian boundary, Fortune Head, Burin Peninsula, southeastern Newfoundland. Canadian Journal of Earth Sciences 24(7), 1277–1293. doi:https://doi.org/10.1139/e87-124
- Nicholson, H.A., 1873: Contributions to the study of the errant annelides of the older Paleozoic rocks. Proceedings of the Royal Society of London 21, 139–147. doi:https://doi.org/10.1098/rspl.1872.0061
- Nielsen, A.T. & Schovsbo, N.H., 2011: The Lower Cambrian of Scandinavia: depositional environment, sequence stratigraphy andpalaeogeography. Earth-Science Reviews 107, 207–310. doi:https://doi.org/10.1016/j.earscirev.2010.12.004
- Nironen, M., Lahtinen, R. & Koistinen, T., 2002: Suomen geologiset aluenimet—yhtenäisempään nimikäytäntöön. Subdivision of Finnish bedrock—an attempt to harmonize terminology. Geologi 54, 8–14.
- Noffke, N., 2009: The criteria for the biogeneicity of microbially induced sedimentary structures (MISS) in Archean and younger, sandy deposits. Earth-Science Reviews 96(3), 173–180. doi:https://doi.org/10.1016/j.earscirev.2008.08.002
- Noffke, N., 2014: Microbially induced sedimentary structures. In R. Amils, et al. (eds.): Encyclopedia of Astrobiology. Springer, Berlin, Heidelberg. doi:https://doi.org/10.1007/978-3-642-27833-4_1004-4
- Orlov, S.U., Kuznetsov, N.B., Miller, E.D., Soboleva, A.A. & Udoratina, O.V., 2011: Age constraints for the Pre-Uralide–Timanide Orogenic Event inferred from the study of detrital zircons. Doklady Earth Sciences 440, 1216–1221. doi:https://doi.org/10.1134/S1028334X11090078
- Orłowski, S., 1989: Trace fossiIs in the Lower Cambrian sequence in the Świętokrzyskie Mountains, central Poland. Acta Palaeontologica Polonica 34, 211–231.
- Orłowski, S. & Żylińska, A., 2002: Lower Cambrian trace fossils from the Holy Cross Mountains, Poland. Geological Quarterly 46, 135–146.
- Ormö, J., Nielsen, A.T. & Alwmark, C., 2017: The Vakkejokk Breccia: an early Cambrian proximal impact ejecta layer in the North-Swedish Caledonides. Meteoritics & Planetary Science 52(4), 623–645. doi:https://doi.org/10.1111/maps.12816
- Pander, C.H., 1851: Sur une découverte de fossiles faite dans la partie inférieure du terrain silurien de Russie. Bulletin de la Societie Geologique de France 8, 251–259.
- Pemberton, S.G. & Frey, R.W., 1982: Trace fossil nomenclature and the Planolites–Palaeophycus dilemma. Journal of Paleontology 56, 843–881.
- Pemberton, S.G., Frey, R.W. & Bromley, R.G., 1988: The ichnotaxonomy of Conostichus and other plug-shaped ichnofossils. Canadian Journal of Earth Sciences 25(6), 866–892. doi:https://doi.org/10.1139/E88-085
- Pemberton, S.G. & Magwood, J.P.A., 1990: A unique occurrence of Bergaueria in the Lower Cambrian Gog Group near Lake Louise, Alberta. Journal of Paleontology 64, 436–440. https://www.jstor.org/stable/1305593
- Peterffy, O., Calner, M. & Vajda, V., 2016: Early Jurassic microbial mats—A potential response to reduced biotic activity in the aftermath of the end-Triassic mass extinction event. Palaeogeography, Palaeoclimatology, Palaeoecology 464, 76–85. doi:https://doi.org/10.1016/j.palaeo.2015.12.024
- Pickerill, R.K., 1998: Bergaueria perata Prantl, 1945 from the Silurian of Cape George, Nova Scotia. Atlantic Geology 25, 191–197.
- Plummer, P.S. & Gostin, V.A., 1981: Shrinkage cracks: desiccation or synaeresis? Journal of Sedimentary Petrology 51, 1147–1156. doi:https://doi.org/10.1306/212F7E4B-2B24-11D7-8648000102C1865D
- Porada, H. & Bouougri, E.H., 2007: Wrinkle structures—a critical review. Earth-Science Reviews 81(3–4), 199–215. doi:https://doi.org/10.1016/j.earscirev.2006.12.001
- Prantl, F., 1945: Dvě zahadné zkameněliny (stopy) z vrstev chrustenických–dδ3. Rozpravy České Akademie Věd a Umění, Třída II 55(3), 1–18.
- Pratt, B.R., 1998: Syneresis cracks: subaqueous shrinkage in argillaceous sediments caused by earthquake-induced dewatering. Sedimentary Geology 117(1–2), 1–10. doi:https://doi.org/10.1016/S0037-0738(98)00023-2
- Quenstedt, F.A., 1879: Petrefactenkunde Deutschlands. Korallen (Die Röhren und Sternkorallen). Abhandlungen 6, 337–496. Fues’s Verlag (R. Reisland), Leipzig.
- Ragozina, A., Dorjnamjaa, D., Krayushkin, A. & Serezhnikova, E., 2008: Treptichnus pedum and the Vendian–Cambrian boundary. 33rd International Geological Congress, 6–14 August 2008, Oslo, Norway, Abstracts. Section HPF 07 Rise and fall of the Ediacaran (Vendian) biota, p. 183.
- Ricci Lucchi, F., 1995: Sedimentographica: photographic atlas of sedimentary structures, 255. Columbia University Press, New York.
- Richter, R., 1850: Aus der thüringischen Grauwacke. Zeitschrift Der Deutschen Geologischen Gesellschaft 2, 198–206.
- Richter, R., 1937: Marken und Spuren aus allen Zeiten. I-II. Senckenbergiana 19, 150–163.
- Roberts, D. & Gee, D.G., 1985: An introduction to the structure of the Scandinavian Caledonides. In D.G. Gee & B.A. Sturt (eds.): The Caledonide Orogen – Scandinavia and related areas, 55–68. John Wiley & Sons Ltd, Chichester.
- Rouse, G.W. & Fauchald, K., 1997: Cladistics and polychaetes. Zoologica Scripta 26(2), 139–204. doi:https://doi.org/10.1111/j.1463-6409.1997.tb00412.x
- Runnegar, B. & Fedonkin, M., 1992: Proterozoic metazoan body fossils. In J. Schopf & C. Klein (eds.): The Proterozoic Biosphere, A Multidisciplinary Study, 369–387. Cambridge University Press, Cambridge.
- Saporta, L.C.J.G.D., 1884: Les organismes problematiques des anciennes mers, 102. Masson, Paris.
- Schlirf, M. & Uchman, A., 2005: Revision of the ichnogenus Sabellarifex Richter, 1921 and its relationship to Skolithos Haldeman, 1840 and Polykladichnus Fürsich, 1981. Journal of Systematic Palaeontology 3(2), 115–131. doi:https://doi.org/10.1017/S1477201905001550
- Schmid, E.E., 1876: Der Muschelkalk des östlichen Thüringen, 20. Fromann, Jena.
- Seilacher, A., 1953: Studien zur Palichnologie, II. Die fossilen Ruhespuren (Cubichnia). Neues Jahrbuch Für Geologie Und Paläontologie, Abhandlugen 98, 87–124.
- Seilacher, A., 1955: Spuren und Fazies im Unterkambrium. In O.H. Schindewolf & A. Seilacher (eds.): Beiträge zur Kenntnis des Kambriums in der Salt Range (Pakistan). Akademie der Wissenschaften und der Literatur zu Mainz, Abhandlung Mathematisch-Naturwissenschaftliche Klasse. Vol. 10, 373–399.
- Seilacher, A., Buatois, L.A. & Mángano, M.G., 2005: Trace fossils in the Ediacaran–Cambrian transition: behavioral diversification, ecological turnover and environmental shift. Palaeogeography, Palaeoclimatology, Palaeoecology 227(4), 323–356. doi:https://doi.org/10.1016/j.palaeo.2005.06.003
- Seilacher, A. & Seilacher, E., 1994: Bivalvian trace fossils: a lesson from actuopalaeontology. Courier Forschung Institut, Senckenberg 169, 5–15.
- Shahkarami, S., Mángano, M.G. & Buatois, L.A., 2017: Ichnostratigraphy of the Ediacaran-Cambrian boundary: new insights on lower Cambrian biozonations from the Soltanieh Formation of northern Iran. Journal of Paleontology 91(6), 1178–1198. doi:https://doi.org/10.1017/jpa.2017.72
- Sharma, M., Pandey, S.K., Ahmad, S., Kumar, K. & Ansari, A.H., 2018: Observations on the ichnospecies Monomorphichnus multilineatus from the Nagaur Sandstone (Cambrian Series 2-Stage 4), Marwar Supergroup, India. Journal of Earth Systems Science 127, 75. doi:https://doi.org/10.1007/s12040-018-0973-9
- Shitole, A.D., Patel, S.J., Jaquilin, K., Joseph, J.K., Jehova, L. & Darngawn, J.L., 2019: Ethological and environmental significance of Bergaueria hemispherica from the Late Cretaceous of the Bagh Group, Western India. Compte Rendu Palevol 18, 287–297. doi:https://doi.org/10.1016/j.crpv.2019.01.002
- Simón, J., 2017: Trace fossils and dubiofossils from the Ediacaran and Cambrian of the Alcudia Anticline, Spain. Estudios Geológicos 73(2), e068. doi:https://doi.org/10.3989/egeol.42724.442
- Símon López-Villalta, J.S., 2019: Fossils from the Azorejo Formation (Lower Cambrian, Central Iberian Zone) in the Guadiana River section at Picón, Ciudad Real. Boletín de la Real Sociedad Española de Historia Natural. Sección biológica 113, 43–65. doi:https://doi.org/10.29077/bol/113/ce06_simon
- Skovsted, C.B., Topper, T.P., McLoughlin, S., Johansson, O., Liu, F. & Vajda, V., this volume: First discovery of Small Shelly Fossils and new occurrences of brachiopods and trilobites from the early Cambrian (Stage 4) of the Swedish Caledonides, Lapland. GFF. doi:https://doi.org/10.1080/11035897.2021.1895303
- Sláma, J., Košler, J., Condon, D.J., Crowley, J.L., Gerdes, A., Hanchar, J.M., Horstwood, M.S.A., Morris, G.A., Nasdala, L., Norberg, N., Schaltegger, U., Schoene, B., Tubrett, M.N. & Whitehouse, M.J., 2008: Plešovice zircon—A new natural reference material for U–Pb and Hf isotopic microanalysis. Chemical Geology 249, 1–35. doi:https://doi.org/10.1016/j.chemgeo.2007.11.005
- Stal, L. & Caumette, P. (eds.), 1994: Microbial Mats – structure, development and environmental significance, 463. Springer, Heidelberg. doi:https://doi.org/10.1007/978-3-642-78991-5
- Stal, L., van Gemerden, H. & Krumbein, W.E., 1985: Structure and development of a benthic marine microbial mat. FEMS Microbiology Letters 74, 305–312. doi:https://doi.org/10.1016/0378-1097(85)90007-2
- Stodt, F., Rice, A.H.N., Björklund, L., Bax, G., Halverson, G.P. & Pharaoh, T.C. 2011: Evidence of late Neoproterozoic glaciation in the Caledonides of NW Scandinavia. In E. Arnaud, G.P. Halverson & G. Shields-Zhou (eds.): The Geological Record of Neoproterozoic Glaciations, 603–611. Geological Society, London. Memoirs 36. doi:https://doi.org/10.1144/M36.58
- Streng, M., Babcock, L.E. & Hollingsworth, J.S., 2005: Agglutinated protists from the lower Cambrian of Nevada. Journal of Paleontology 79(6), 1214–1218. https://www.jstor.org/stable/4095005
- Systra, Y.J. & Jensen, S., 2006: Trace fossils from the Dividalen Group of northern Finland with remarks on early Cambrian trace fossil provincialism. GFF 128(4), 321–325. doi:https://doi.org/10.1080/11035890601284321
- Tewari, V.C., 1999: Vendotaenids: earliest megascopic multicellular algae on Earth. Geoscience Journal 20, 77–85.
- Thelander, T., 1982: The Torneträsk formation of the Dividal Group, northern Swedish Caledonides. Sveriges Geologiska Undersökning C 789, 1–41.
- Thelander, T., 2009: Beskrivning till berggrundskartan Kaledoniderna i norra Sverige, K 222. Sveriges Geologiska Undersökning, Uppsala.
- Torell, O., 1870: Petrificata Suecana Formationis Cambricae. Acta Universitatis Lundensis 6(8), 1–14.
- Uchman, A., 1995: Taxonomy and paleoecology of flysch trace fossils: the Marnoso-arenacea Formation and associated facies (Miocene, Northern Apennines, Italy). Beringeria 15, 1–115.
- Uchman, A. 1998. Taxonomy and ethology of flysch trace fossils: revision of the Marian Książkiewicz Collection and studies of complementary material. Annales Societatis Geologorum Poloniae 68, 105–218.
- Uchman, A., Hanken, N.-M. & Binns, R., 2005: Ordovician bathyal trace fossils from metasiliciclastics in central Norway and their sedimentological and paleogeographical implications. Ichnos 12(2), 105–133. doi:https://doi.org/10.1080/10420940590914534
- Urbanek, A. & Mierzejewska, G., 1983: The fine structure of zooidal tubes Sabelliditida and Pogonophora. In A. Urbanek & A.Y. Rozanov (eds.): Upper Precambrian and Cambrian Palaeontology of the East-European Platform, 100–111. Publishing House Wydawnictwa Geologiczne, Warszawa.
- Vannier, J., Calandra, I., Gaillard, C. & Żylińska, A., 2010: Priapulid worms: pioneer horizontal burrowers at the Precambrian-Cambrian boundary. Geology 38(8), 711–714. doi:https://doi.org/10.1130/G30829.1
- Vialov, O.S., 1962: Problematica of the Beacon Sandstone at Beacon Heights, West Antarctica. New Zealand Journal of Geology and Geophysics 5, 718–732. doi:https://doi.org/10.1080/00288306.1962.10417633
- Vidal, G., 1989: Are late Proterozoic carbonaceous megafossils metaphytic algae or bacteria? Lethaia 22(4), 375–379. doi:https://doi.org/10.1111/j.1502-3931.1989.tb01437.x
- Vidal, G. & Moczydłowska, M., 1996: Vendian–lower Cambrian acritarch biostratigraphy of the central Caledonian fold belt in Scandinavia and the palaeogeography of the Iapetus-Tornquist seaway. Norsk Geologisk Tidsskrift 76, 147–168.
- Walter, M.R., Elphinstone, R. & Heys, G.R., 1989: Proterozoic and Early Cambrian trace fossils from the Amadeus and Georgina basins, central Australia. Alcheringa 13, 209–256. doi:https://doi.org/10.1080/03115518908527821
- Wang, X., Pang, K., Chen, Z., Wan, B., Xiao, S., Zhou, C. & Yuan, X., 2020: The Ediacaran frondose fossil Arborea from the Shibantan limestone of South China. Journal of Paleontology 94(6), 1034–1050. doi:https://doi.org/10.1017/jpa.2020.43
- Wang, Y., Lin, J.-P., Zhao, Y.-L. & Orr, P.J., 2009: Palaeoecology of the trace fossil Gordia and its interaction with nonmineralizing taxa from the early Middle Cambrian Kaili Biota, Guizhou province, South China. Palaeogeography, Palaeoclimatology, Palaeoecology 277(1–2), 141–148. doi:https://doi.org/10.1016/j.palaeo.2009.02.017
- Watson, E.B., Wark, D.A. & Thomas, J.B., 2006: Crystallization thermometers for zircon and rutile. Contributions to Mineralogy and Petrology 151(4), 413–433. doi:https://doi.org/10.1007/s00410-006-0068-5
- Webby, B.D., 1984: Precambrian–Cambrian trace fossils from western New South Wales. Australian Journal of Earth Sciences 31, 427–437. doi:https://doi.org/10.1080/08120098408729302
- Weber, B., Hu, S.X., Steiner, M. & Zhao, F.C., 2012: A diverse ichnofauna from the Cambrian Stage 4 Wulongqing Formation near Kunming (Yunnan Province, South China). Bulletin of Geosciences 87, 71–92. doi:https://doi.org/10.3140/BULL.GEOSCI.1239
- Xiao, S. & Dong, L., 2006: On the morphological and ecological history of Proterozoic macroalgae. In S. Xiao & A.J. Kaufman (eds.): Neoproterozoic Geobiology and Paleobiology, 57–90. Springer, Dordrecht.
- Xiao, S., Chen, Z., Pang, K., Zhou, C. & Yuan, X., 2020: The Shibantan Lagerstätte: insights into the Proterozoic–Phanerozoic transition. Journal of the Geological Society 178, jgs2020–135. doi:https://doi.org/10.1144/jgs2020-135
- Yang, R., Zhang, C., Song, G. & Luo, X., 2006: Vermicular fossils in the Early Cambrian Xidashan Formation in the Quruqtagh region of Xinjiang, China. Progress in Natural Science 16, 559–562. doi:https://doi.org/10.1080/10020070612330035
- Yanichevsky, M., 1926: Sur les restes des Tubicola de l’argile Cambrien. Ežegodnik Vsesoûznogo Paleontologičeskogo Obŝestva 4, 99–112.
- Young, F.G., 1972: Early Cambrian and older trace fossils from the southern Cordillera of Canada. Canadian Journal of Earth Sciences 9(1), 1–17. doi:https://doi.org/10.1139/e72-001
- Zenker, J.C., 1836: Historisch-topographisches Taschenbuch von Jena und seiner Umgebung besonders in seiner naturwissenschaftlicher und medicinischer Beziehung, 338. Wackenholder, Jena.
- Zhang, L.-J., Fan, R.-Y. & Gong, Y.M., 2015a: Zoophycos macroevolution since 541 Ma. Scientific Reports 5, 14954. doi:https://doi.org/10.1038/srep14954
- Zhang, L.-J., Shi, G.R. & Gong, Y.M., 2015b: An ethological interpretation of Zoophycos based on Permian records from South China and southeastern Australia. Palaios 30, 408–425. doi:https://doi.org/10.2110/palo.2013.060