
ABSTRACT
An Early Ordovician fauna of linguliform brachiopods and euconodonts is described from the Alum Shale Formation in northernmost Västerbotten County, northern Sweden. The fauna was recovered from a single carbonate concretion collected at an exposure near the northern shore of Lake Storvindeln. Despite of the fragmentary and tectonized preservation of the obtained fossils, at least eight different taxa of linguliform brachiopods and four species of euconodonts could be identified. Among the brachiopods, the ephippelasmatid Pomeraniotreta biernatae Bednarczyk, 1986 is most common; it is associated among others with the acrotretids Eurytreta cf. sabrinae (Callaway, 1877) and Ottenbyella sp., and a spinose specimen potentially representing the zhanatellid Thysanotos. Two of the brachiopod taxa appear to present new species, Pomeraniotreta n. sp. and Mytoella? n. sp., but additional material would be needed for a formal description. Euconodont specimens are rare and comprise coniform elements of Drepanodus arcuatus Pander, 1856, Drepanoistodus aff. amoenus (Lindström, 1955) sensu Löfgren (1994), Paroistodus numarcuatus (Lindström, 1955) and Rossodus aff. manitouensis Repetski & Ethington, 1983. Evaluation of the stratigraphic distribution of the different taxa of brachiopods and euconodonts under consideration of the local geology, suggests that the investigated sample represents a level in the upper part of the Tremadocian (Tr2) part of the Alum Shale Formation. The fauna represents the northernmost occurrence of age diagnostic Early Ordovician (Tremadocian) fossils in Sweden.
Introduction
Cambrian to Early Ordovician sedimentary rocks are preserved in a narrow zone along the eastern Caledonian margin in central and northern Sweden and Norway. In Sweden, this includes the central part of the province of Jämtland and northwards the province of Lapland. The Lower Paleozoic sedimentary regions in Lapland have traditionally been divided into southern Lapland, which essentially encompass Västerbotten County, and northern Lapland, which encompass Norrbotten County (Kulling Citation1960). This arbitrary division results from the former being less well studied than the latter (Martinsson Citation1974).
Extensive mapping has elucidated the regional geology of Lapland and revealed that Lower Paleozoic strata are present in both the narrow autochthonous zone at the Caledonian front as well as the Lower Allochthon (Kulling Citation1942, Citation1955, Citation1960, Citation1964, Citation1982; Gee et al. Citation1974; Greiling et al. Citation1999; Eliasson et al. Citation2001a, Citation2001b, Citation2003; Greiling & Kathol Citation2021 and references therein). Kulling (Citation1960, p. 161) pointed out that some of the sedimentary rocks in the former may have been thrusted, and some have now been shown to be part of the Lower Allochthon.
The age of the lower Cambrian succession is constrained by trilobites and microfossils (Ahlberg & Bergström Citation1983; Moczydłowska Citation1991; Moczydłowska et al. Citation2001; Cederström et al. Citation2012). Middle Cambrian (Miaolingian) fossils along the Caledonian front are known from the localities Tallträsk and Kyrkberget about 27 km north-west of the town of Storuman, Västerbotten County (Holm Citation1890, p. 262; Kulling Citation1942, p. 42, Citation1955, p. 148). Alum Shale with calcareous concretions north of Storuman has thus by extension generally been considered middle Cambrian in age (Kulling Citation1942, Citation1955, Citation1964; Kautsky Citation1945; Lilljequist Citation1973; Willdén Citation1980; Andersson et al. Citation1985; Greiling et al. Citation1999; Eliasson et al. Citation2001a, Citation2001b, Citation2003). Kulling (Citation1955, p. 147) reported alum shale with concretions without fossils just north of Lake Storvindeln. Concretions with paradoxidid trilobite fragments of middle Cambrian age have been recorded in the Lower Allochthon at Lake Gautojaure north-west of Arjeplog (Kulling Citation1982, p. 63) and Kautsky (Citation1945, p. 133) suggested that the middle Cambrian was represented in the Laisvall area just north of Arjeplog.
The black shales of the Alum Shale Formation in Lapland have broadly been considered Middle Cambrian to the Early Ordovician in age but precise age determination has essentially been lacking (Kulling Citation1960, p. 161). The northernmost localities yielding age diagnostic Early Ordovician (Tremadocian) fossils are situated ca. 60 km SSW of Storuman, near Vilhelmina, Västerbotten County (Kulling Citation1955, p. 151) where for instance the Tremadocian Bjørkåsholmen Formation is exposed at Granhöjden and overlain by shales of the Tøyen Formation (Mörtsell Citation1888; pers. observation by Ebbestad). In the Laisvall area, Norrbotten County, a greywacke sequence, possibly equivalent to the Föllinge and Norråker formations further south (Gee et al. Citation1974; Karis Citation1998) is generally considered Ordovician in age but any fossils to corroborate this have not been found (Lilljequist Citation1973; Greiling & Garfunkel Citation2007) (see for locations).
Figure 1. Maps showing locality where the sample was collected (star in C). The shaded area in Fig. A shows the County of Västerbotten. The sample was found ca. 560 m NNE of Jillesnåle farm, situated close to the northern shore line of the northwestern end of Lake Storvindeln (Fig. C) in northwestern Västerbotten County, Sweden.
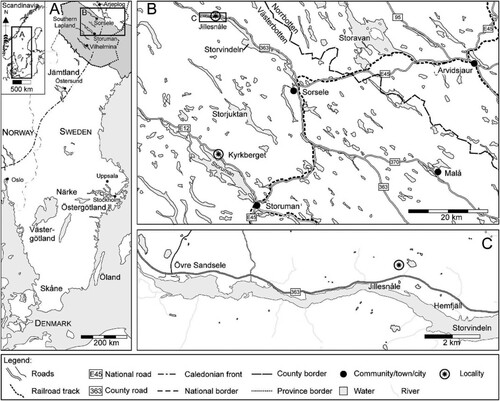
Only recently (since 2001), age indicative fossils have been recovered from the sediments along the Caledonian front north of Storuman through systematic investigations and sampling by Thomas Weidner. This paper describes linguliform brachiopods and euconodonts recovered from a small carbonate concretion in the Alum Shale Formation at Jillesnåle on the northern shore of Lake Storvindeln in southern Lapland in northernmost Västerbotten County (). This is the northernmost occurrence of this fauna hitherto documented in Sweden, and although fragmentary it can be compared to similar faunas further south in Scandinavia and provides a precise age determination for these strata.
Material and geological setting
The area around Lake Storvindeln has been re-mapped by Eliasson et al. (Citation2001a, Citation2001b, Citation2003) and more recently by Greiling & Kathol (Citation2021) documenting outcrops of the Alum Shale Formation on the north side of Lake Storvindeln mainly along the shore, in rivulets, and road cuts. Rare carbonate concretions occur in the shale, but most of these are devoid of macrofossils (trilobites). One sample from Hemfjäll (C) has yielded trilobites of the Acadoparadoxides pinus – Pentagnostus praecurrens Zone of the Acadoparadoxides (Baltoparadoxides) oelandicus Superzone (Weidner et al. Citation2021). The material described in the present report comes from a single fist-sized concretion collected high up in the cliff exposure at Jillesnåle. The concretion drew the attention of T. Weidner during reconnaissance work in 2015 as its lithofacies appeared different from the other concretions recovered in the vicinity. Because it was devoid of macrofossils it was sent to the acid preparation lab in Lund, Lund University, Sweden, for extraction of phosphatic microfossils and the 250–1000 µm fraction yielded abundant but fragmentary microfossils. The material is housed in the paleontological collections at the Museum of Evolution, Uppsala University (PMU).
Age of the fauna
The sample from Jillesnåle yielded at least eight different taxa of linguliform brachiopods and four species of euconodonts. Despite a relatively diverse association, an age estimation based on brachiopods is aggravated by various factors. Most of the taxa are poorly preserved and/or represented by a low number of specimens. Hence, their assigned systematic position is not unequivocal (e.g., aff. Acrotreta sp., Obolinae gen. and sp. indet.). In addition, some of the taxa are likely to represent new species (e.g., Mytoella? n. sp., Pomeraniotreta n. sp.). Essentially only three of the brachiopod taxa can provide useful biostratigraphic information, i.e., Eurytreta cf. sabrinae (Callaway, Citation1877), Ottenbyella sp., and Pomeraniotreta biernatae Bednarzcyk, Citation1986.
Eurytreta sabrinae (Callaway, Citation1877) has originally been described from the Tremadocian of the Anglo-Welsh Basin where it is known from the base of the Ordovician (Rhabdinopora flabelliformis Biozone) to the mid-Tremadocian (Conophrys salopiensis Biozone) (see Sutton et al. Citation2000). Similar and potentially conspecific forms have been described from the middle Tremadocian of France and the latest Cambrian (Cordylodus andresi conodont zone) of Oaxaquia, Mexico (see summary in Streng et al. Citation2011). Occurrences of the genus Eurytreta (sensu Streng et al. Citation2011) on Baltica were so far restricted to E. chabakovi from the Tremadocian of the South Ural Mountains and potentially an unnamed species from the late Tremadocian of southern Sweden (see Systematic paleontology).
Ottenbyella Popov & Holmer, Citation1994 has originally been described from mid-Tremadocian limestones in southern Sweden (Popov & Holmer Citation1994) and has since been reported from deposits in Iran, Laurentia, Kazakhstan, and potentially Oaxaquia ranging from the Furongian to the Floian (Holmer et al. Citation2005; Popov et al. Citation2009; Streng et al. Citation2011). On Baltica, Ottenbyella is so far restricted to southern Scandinavia.
Pomeraniotreta biernatae Bednarzcyk, Citation1986 is known exclusively from Baltica where it occurs from the mid-Tremadocian (Paltodus deltifer Biozone) in southern Scandinavia to the early Floian of northern Poland (see Systematic Paleontology). In Scandinavia it is the most common species in associations of linguliform brachiopods from the Bjørkåsholmen and Köpingsklint formations (Popov & Holmer Citation1994). Collectively, the brachiopod fauna suggests a mid to late Tremadocian age of the Jillesnåle sample.
In comparison to the linguliform brachiopods, euconodonts are scarce with only six recovered specimens. Still, the four identified species allow a rather precise age assignment. The small fauna includes Drepanodus arcuatus Pander, Citation1856, Drepanoistodus aff. amoenus (Lindström, Citation1955) sensu Löfgren (Citation1994), Paroistodus numarcuatus (Lindström, Citation1955), Rossodus aff. manitouensis Repetski & Ethington, Citation1983, and Genus et species indet. Except for the long-ranging species Drepanodus arcuatus, the identified conodont taxa are restricted to the Tremadocian and have been reported previously from the Ørnberget Formation of the Norwegian Caledonides (Bruton et al. Citation1989; Rasmussen Citation2001), the Bjørkåsholmen Formation of Norway (Rasmussen Citation2001), and from strata just below or within the Ceratopyge Limestone and Köpingsklint Formation of Sweden (e.g., Lindström Citation1971; van Wamel Citation1974; Bagnoli et al. Citation1988; Sturkell Citation1991; Löfgren Citation1993, Citation1997b; Löfgren et al. Citation1999). The conodont assemblage shows that the limestone concretion from the Alum Shale Formation at Jillesnåle correlates with a level within the upper part of the Paltodus deltifer Zone or – less probable due to the occurrence of Rossodus aff. manitouensis – the lowermost subzone of the Paroistodus proteus Subzone, i.e., a late mid or early late Tremadocian age.
The widespread, dark fissile mudstones of the Alum Shale Formation are terminated by the first persistent development of carbonates in Baltoscandia. On Öland, in the south eastern part of Sweden, the Alum Shale Formation is followed by glauconite-rich sand- and siltstones of the Djupvik Formation, which is overlain by the glauconitic carbonates of the Köpingsklint Formation (Stouge Citation2004). The base of the latter was shown by Frisk & Ebbestad (Citation2008) to correspond to the first occurrence of the trilobite Ceratopyge acicularis, the zonal fossil of the Ceratopyge acicularis Trilobite Zone as redefined by Pärneste et al. (Citation2013). In the districts of Västergötland, Scania and southern part of Västerbotten in Sweden and in the Oslo Region of Norway the Alum Shale Formation is overlain by the Bjørkåsholmen Formation (Mörtsell Citation1888; Tjernvik Citation1956; Ebbestad Citation1997, Citation1999). In the latter area, the occurrence of Ceratopyge acicularis coincides with the base of the formation while Ceratopyge forficula occurs below in the Alum Shale Formation (Ebbestad Citation1999). In the Swedish provinces of Östergötland, Närke and Dalarna, the Bjørkåsholmen Formation is missing altogether. Based on conodonts belonging to the Paltodus deltifer Conodont Zone, the first thin limestone interval following the Alum Shale Formation in the autochthon of Jämtland was identified by Sturkell (Citation1991) as the Ceratopyge Limestone (=Bjørkåsholmen Formation) (see for correlations).
Figure 2. Simplified correlation chart of the Tremadocian strata of Norway and Sweden. The references to geographical directions with each locality column indicate the areas relative position on the Baltoscandian platform. Sedimentologically the western carbonate/glauconite facies represent distal ramp environments while the eastern are proximal ramp environments (Egenhoff et al. Citation2010). The star in the last column indicates inferred position of the sample from Jillesnåle. Stage Slices and conodont zonations are largely based on Bergström et al. (Citation2009). See text for further references.
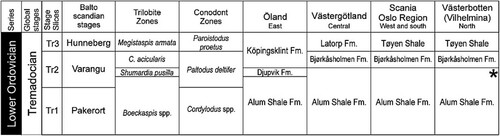
Bergström et al. (Citation2009) used the Paltodus deltifer Conodont Zone to define the base of the Tremadocian 2 Stage Slice. It has been shown to span the Ceratopyge acicularis interval, that is the Bjørkåsholmen Formation and lower part of the Köpingsklint Formation (Lindström Citation1971; Sturkell Citation1991; Löfgren Citation1993; Rasmussen Citation2001; Stouge Citation2004), but also being present in the upper part of the Alum Shale Formation (Lindström Citation1971; van Wamel Citation1974). Hence, based on (1) the brachiopod and conodont contents, (2) the consistent development of the transition from the Alum Shale Formation to the limestones of the Bjørkåsholmen Formation as far north as Vilhelmina, and (3) the lack of any such limestones with trilobites of the Ceratopyge acicularis Zone, the Jillesnåle sample most likely represent a level close to the upper boundary of the Tremadocian (Tr2) part of the Alum Shale Formation.
Systematic paleontology
PHYLUM BRACHIOPODA
SUBPHYLUM LINGULIFORMEA Williams et al., Citation1996
CLASS LINGULATA Gorjansky & Popov, Citation1985
ORDER LINGULIDA Waagen, Citation1885
SUPERFAMILY LINGULOIDEA Menke, Citation1828
FAMILY OBOLIDAE King, Citation1846
SUBFAMILY OBOLINAE King, Citation1846
Mytoella Mergl, Citation2002
Diagnosis. – See Mergl (Citation2002).
Type species. – M. krafti Mergl, Citation2002, p. 28–29, from the Middle Ordovician (Darriwilian) Šárka Formation, near Osek, Czech Republic.
Other species included. – Lingulella pusilla Želízko, Citation1921 from the Lower Ordovician (Floian) of Bohemia, Czech Republic (according to Mergl Citation2002).
Remarks. – Hansen (Citation2008) described “Mytoella? robusta” from the Upper Ordovician (Katian) of the Oslo Region based on few poorly preserved specimens. Provided illustrations and description, however, are ambiguous and do in our opinion not allow a comparison with Mytoella. Furthermore, Hansen (Citation2008, p. 24) assigned two specimens as holotype, a ventral and a dorsal valve. The two specimens co-occur on one slab but are not conjoined and hence do not unambiguously represent a single individual. Accordingly, we consider the name “Mytoella? robusta” as unavailable as its introduction is not in agreement with ICZN Article 16.4.1 (Ride et al. Citation1999).
Occurrence. – Early and Middle Ordovician of Bohemia (e.g., Mergl Citation2002) and questionable form the Early Ordovician of Baltica (this study).
Mytoella? n. sp.
A–E, 6H
Material. – One incomplete ventral valve in addition to three fragments that cannot be assigned to either valve.
Description. – Valves characterized by coarse and somewhat irregular, concentric rugae, also expressed on internal shell surface; no micro-ornamentation seen on external shell surface. Type of shell ultrastructure uncertain as observed shell laminae are recrystallized; shell laminae subparallel to shell surface (C, 4H). Largest preserved specimen, an incomplete ventral valve, measures ca. 1 mm in length (A); its preserved outline suggests an originally elongate elliptical outline and a total length of not more than 1.2 mm. Ventral valve strongly convex with relatively smooth, i.e., rugae-free, apical external surface (larval shell?); transition between apical and rugose part of the shell not clearly demarcated. Internally, ventral valve characterized by poorly elevated pseudointerarea and shallow pedicle groove. Propareas inclined towards the pedicle groove and divided by faint flexure line (C). Anterior margin of proparea evenly convex, while lateral margin is straight but with distinct concave flexure near corner with anterior margin (B). No other internal features observed. Features of dorsal valve unknown.
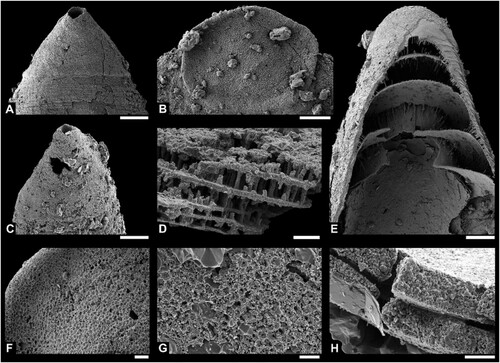
Figure 3. A–E. Mytoella? n. sp. A–C. Ventral valve (PMU38359) in external, internal and lateral view. D, E. Shell fragments PMU38361 and PMU38376 in external and internal view, respectively. F. Obolinae gen. and sp. indet. (PMU38385), exterior of exfoliated ventral valve. G. Thysanotos? sp. (PMU38365), recrystallized shell fragment with marginal spines. Scale bar equals 100 µm, except G (200 µm).

Remarks. – Among the known oboline genera, characters of Mytoella Mergl, Citation2002 agree best with the material described herein. This includes the development of a low ventral pseudointerarea, an elongate elliptical outline, relatively strong convexity of the valves, an external surface void of micro-ornamentation, and the absence of distinct pallial markings. The coarse rugae developed in Mytoella? n. sp. have not been described for the genus suggesting that the single valve might represent a new species. Mergl (Citation2002) described Mytoella as a “minute” genus but with maximum sizes of up to 9 mm, the Bohemian taxa are distinctly bigger than the Jillesnåle specimen.
The coarse concentric rugae of Mytoella? n. sp. remind to some extend of Teneobolus gracilis Mergl, Citation1995 from the Early Ordovician of Bohemia (Mergl Citation1995, Citation2002). Teneobolus is, however, characterized by a deep, anteriorly projecting pedicle groove (Mergl in Holmer & Popov Citation2007, p. 2542). Rebrovia Popov & Khazanovitch in Popov et al., Citation1989 from the Furongian of Estonia and Russia has like Mytoella a low ventral pseudointerarea but lacks flexure lines.
Obolinae gen. and sp. indet.F
Material. – 3 valves.
Remarks. – In addition to the taxa described herein in more detail, the sample from Jillesnåle also contained several fragmentary and/or deformed valves that belong to at least two additional oboline taxa. Their state of preservation is, however, too poor to allow any confident systematic treatment. A potential ventral valve is exemplarily illustrated (F).
FAMILY ZHANATELLIDAE Koneva, Citation1986
Thysanotos Mickwitz, Citation1896
Synonymy and diagnosis. – See Holmer & Popov (Citation2000).
Type species. – Obolus siluricus Eichwald, Citation1840a, p. 195–196, from the Lower Ordovician Leetse Formation (Hunneberg Regional Stage), Paldiski, northern Estonia.
Other species. – T. multispinulosus Popov, Ghobadi Pour & Hosseini, Citation2008 from the upper Lower Ordovician Lashkarak Formation of Iran. Other species previously assigned to Thysanotos represent according to Popov & Holmer (Citation1994) variations of the type species. However, some authors maintain at least one of the species, T. primus (Koliha, Citation1924) from the Trenice Formation of Bohemia, as a distinct species (e.g., Mergl Citation2002; Candela et al. Citation2021).
Remarks. – There is some confusion and inconsistencies in the literature concerning the year of publication as well as the actual publication of the formal description of the type species of Thysanotos, Obolus siluricus. Karl Eduard von Eichwald, the author of the type species published his studies mainly in German or Russian, but also in French, and commonly the same work could have been re-published several times, as a translation, as special offprints, or as extended translations. These multiple publications of the same study, which could have been published in different years and not necessarily would match exactly in content, are the source of this confusion. When Mickwitz (Citation1896, p. 194–195) introduced the genus Thysanotos, he referred to Eichwald (Citation1843, p. 5–8, pl. 1, fig. 15) as the source of the protonym. This was followed subsequently by, e.g., Walcott (Citation1912), Sdzuy (Citation1955), and Rowell (Citation1965). It was Gorjansky (Citation1969) who pointed out that Eichwald already in 1840 published a description together with the proposed name Obolus siluricus (Eichwald Citation1840a, p. 195). Accordingly, subsequent taxonomic publications on Thysanotos used 1840 as the year of original publication (e.g., Havlíček Citation1982; Popov & Holmer Citation1994; Holmer & Popov Citation2000; Popov et al. Citation2008). However, commonly these publications referred to the wrong publication in their reference lists, i.e., Eichwald (Citation1840b), which is Eichwald's original description of the Stratigraphy of Estonia written in German. The publication Gorjansky (Citation1969), refers to, Eichwald (Citation1840a), is written in Russian and was most likely intended to become the Russian translation of the original German text. As outlined by Gorjansky (Citation1969, p. 32), during the preparation of the Russian translation, Eichwald seemed to have continued to work on the text. Hence, the German version only contains the description of a potential new species of Obolus from the green sands at Baltischport (=Paldiski) (Eichwald Citation1840b, p. 168–169), whereas the Russian version in addition also mentions the proposed new name Obolus siluricus (Eichwald Citation1840a, p. 195). It should be noted that the publication “Eichwald (Citation1840a)” is a special offprint of what was also published in the Journal of Medical and Natural Sciences in two parts (Eichwald Citation1840c and Eichwald Citation1841). Here, the second part includes the description and mentioning of Obolus siluricus (Eichwald Citation1841, pp. 61–62). Mickwitz (Citation1896) probably was not aware of these earlier publications, but used the publication in which he thought Obolus siluricus was first described and illustrated, i.e., Eichwald (Citation1843). As Obolus siluricus was illustrated for the first time in Eichwald (Citation1843) in which only the German description was referred to, i.e., “Eichwald (Citation1840b)”, (Eichwald Citation1843, p. 6), Mickwitz's error is understandable. Additionally, also the publication “Eichwald (Citation1843)” exists as a special offprint, which was already published in 1842 (Eichwald Citation1842). Interestingly, in another publication from 1840, potentially preceding Eichwald (Citation1840a, Citation1840b), Eichwald proposed a new species of Obolus from the same locality and stratigraphic level as O. siluricus, which he referred to as “Obolus semisulcatus” (Eichwald Citation1840d, p. 6). Eichwald's corresponding publication in Russian with a similar title (Eichwald Citation1840e) does not mention this name. As no description was provided for “Obolus semisulcatus” by Eichwald (Citation1840d), the affinity of the species is unclear. Accordingly, the name has to be considered as unavailable and presents a nomen nudum.
Late Tremadocian occurrences of the species Thysanotos primus (Koliha, Citation1924) in Bohemia (Czech Republic) were for a long time regarded as the oldest record of the genus Thysanotos (e.g., Bednarczyk Citation1999; Mergl Citation2002). Occurrences on the eastern European Platform (Hunneberg Local Stage) were supposed to be younger (Popov & Holmer Citation1994) although more recent correlation schemes put the lower part of the Hunneberg Stage in the upper Tremadocian (Pärnaste et al. Citation2013). The view of a late Tremadocian appearance of Thysanotos was, however, recently challenged by reports of Thysanotos sp. from the early Tremadocian of the Avalonian Brabant Massif in Belgium (Candela et al. Citation2021). T. multispinulosus Popov, Ghobadi Pour & Hosseini, Citation2008 from the upper Lower Ordovician (Floian) of Iran represents the youngest report of the genus (Popov et al. Citation2008; Popov & Cocks Citation2017).
Occurrence. – The genus Thysanotos is known from the Early Ordovician of Baltica (Estonia, Poland, South Urals) (e.g., Gorjansky Citation1969; Popov & Holmer Citation1994), northwestern peri-Gondwana (Germany, Czech Republic) (e.g., Sdzuy Citation1955; Havlíček Citation1982; Mergl Citation2002), Avalonia (Candela et al. Citation2021), and Iran (Popov et al. Citation2008). The specimen described below might represent the first occurrence of the genus in Sweden.
Thysanotos? sp.
G
Material. – One shell fragment.
Description. – Shell fragment rectangular in shape bearing four solid spines along one of its longer sides. Fragment about 1.1 × 0.45 mm in size, excluding spines. Near base, spines are subrectangular in cross-section, measuring ca. 150 µm in width; spines taper distally, reaching lengths of more than 130 µm. Individual spines not conjoined at their bases but separated from each other by a distance of 90–155 µm, with spine midpoints 260–320 µm apart. No micro-ornamentation on either side of the shell observed; shell ultrastructure unclear.
Remarks. – The specimen is interpreted to potentially represent a fragment of the margin of the shell or of a distinct growth lamella of Thysanotos. The affiliation to Thysanotos is based on the presence of solid, marginal spines, which are in size and arrangement comparable to those described for the genus. In other taxa known to have similar marginal spines, these are smaller and arranged more densely. This includes the elkaniids Keskentassia multispinulosa Popov & Holmer, Citation1994 from the Lower Ordovician (Floian) of southern Kazakhstan (Popov & Holmer Citation1994) and Broeggeria fimbriata (Hadding, Citation1913) from the Middle Ordovician of Norway (e.g., Candela & Hansen Citation2010), the acrothelid Orbithele spp. reported from the Miaolingian to Middle Ordovician of mainly Baltica and peri-Gondwana (e.g., Popov & Holmer Citation1994; Holmer & Popov Citation2000; González-Gómez Citation2005a; Popov et al. Citation2008), and the obolid Barbatulella lacunosa Williams & Lockley, Citation1983 from the Late Ordovician (Williams & Lockley Citation1983). Reported distances between spines in these taxa do not exceed 150 µm, except for Barbatulella for which distances around 250 µm have been described in large valves (Williams & Lockley Citation1983), which is similar to measured distances in Thysanotos? sp. Spines in Barbatulella are, however, connected at their bases generating a typical zigzag pattern and most likely were poorly mineralized or even entirely organic (Williams & Lockley Citation1983). Distances between spines in Thysanotos increase with shell size and can reach more than 700 µm (Popov & Holmer Citation1994). Hence, the genus Thysanotos appears to be the best comparison for the fragment from Jillesnåle. The lack of pitted micro-ornament typical for the genus could be explained by the recrystallized state of the fragment or that it represents an exfoliated shell lamina. However, due to the rather incomplete and fragmentary nature of the specimen, which represents only a minor part of the original valve, in addition to the lack of complemental diagnostic characters of Thysanotos, the specimen is only questionably assigned to the genus.
ORDER ACROTRETIDA Kuhn, Citation1949
SUPERFAMILY ACROTRETOIDEA Schuchert, Citation1893
FAMILY ACROTRETIDAE Schuchert, Citation1893
Genus Eurytreta Rowell, Citation1966
Diagnosis. – See Streng et al. (Citation2011).
Type species. – Acrotreta curvata Walcott, Citation1902, p. 584, from the Lower Ordovician (Tremadocian?) Pogonip Limestone, Eureka district, Nevada, USA.
Other species included. – E. ahmadii Ghobadi Pour, Kebriaee-Zadeh & Popov, Citation2011 from an unnamed Lower Ordovician (Tremadocian) lithostratigraphic unit following the Mila Formation, northern Iran, and E. harringtoni Mergl & Herrera in Mergl et al., Citation2015 from the Furongian (Cambrian Stage 10) Lampazar Formation, Salta Province, northwestern Argentina, in addition to the species listed by Streng et al. (Citation2011).
Remarks.. – As outlined by Streng et al. (Citation2011), most of the Baltic specimens previously assigned to Eurytreta by Popov & Holmer (Citation1994), i.e., E. minor and E. cf. sabrinae, might not belong to Eurytreta. They differ from typical species of Eurytreta in having an invariably procline rather than catacline or apsacline ventral valves. Specimens illustrated by Popov & Holmer (Citation1994, fig. 89) as Eurytreta sp. a from the Bjørkåsholmen Formation of southern Sweden might belong to Eurytreta. The lack of a description and illustrations of the ventral interior, however, hamper a confident generic assignment as well as meaningful comparison with the specimens described below. Hence, the specimens described below as Eurytreta cf. sabrinae might represent the first specimens of Eurytreta (sensu Streng et al. Citation2011) from Scandinavia.
Several other species attributed to Eurytreta have been excluded from the genus as their characters were not compatible with the genus diagnosis (Streng et al. Citation2011). This would also apply to the species Eurytreta subtriangularis Hansen & Holmer, Citation2011 from the late Floian of Spitsbergen. Its widely spaced dorsal cardinal muscle scars and distinct interridge of the ventral pseudointerarea are not typical for Eurytreta, but might be representative of a new genus.
Occurrence. – Eurytreta sensu Streng et al. (Citation2011) is known from the Furongian to Early Ordovician of successions in Laurentia, eastern Avalonia, Kazakhstan, Iran, Baltica, Oaxaquia, and Argentina (see above and Streng et al. Citation2011 for details).
Eurytreta cf. sabrinae (Callaway, Citation1877)
A, B, D
n 1994 Eurytreta cf. sabrinae (Callaway, Citation1877): Popov & Holmer, p. 104–106, figs. 86I, 87, 88.
Figure 4. A, B, D. Eurytreta cf. sabrinae (Callaway, Citation1877). A, D. Ventral valve (PMU38362) in oblique anterior and lateral view showing apical process and broad vascula lateralia. B. Ventral valve (PMU38363) in posterior view showing larval shell and flat undivided pseudointerarea. C, F, G. Two dorsal valves of aff. Eurytreta sp. (C, PMU38374 and F, G, PMU38375) showing subrectangular median buttress and broad triangular median groove. E, H, I. Ottenbyella sp. (PMU38364) in ventral exterior, interior, and lateral view; note subrectangular apical process and narrow vascula lateralia. J, K, Dorsal valve of aff. Acrotreta sp. (PMU38360) with triangular median septum. L. Potential ventral valve of aff. Acrotreta sp. (PMU38386). Scale bar equals 100 µm.

Material. – Three ventral valves.
Description. – Valves conical in shape with apsacline, triangular pseudointerarea. Lateral margins of seemingly undivided, flat pseudointerarea poorly defined, but marked by distinct flexure in one specimen (B). Exterior pedicle opening at apex, enclosed within larval shell. Larval shell ornamented with hemispherical imprints of similar sizes, 2–3 µm in diameter. Ornamentation of concentric fila on postlarval shell poorly preserved. Interior characterized by well developed, ramp-like, triangular apical process on anterior slope. Process broadens anteriorly, demarcated by distinct, broad, baculate vascula lateralia. Apical pits not seen.
Remarks. – Overall, the described characters of the three ventral valves suggest reference to Eurytreta. Somewhat uncharacteristic for Eurytreta is the quite distinct demarcation of the pseudointerarea by flexures observed in one valve. This feature, however, is interpreted to be secondary, being the result of tectonic deformation of the shell. If the described absence of a median division of the pseudointerarea is a genuine feature or due to poor and incomplete preservation is unclear.
Among the known species of Eurytreta, the specimens from Jillesnåle are most similar to E. sabrinae described from the Tremadocian of England and Wales (e.g., Sutton et al. Citation2000), southern France (González Gómez Citation2005b) and potentially Mexico (Streng et al. Citation2011). Specimens described previously from Scandinavia as well as from the South Urals (Popov & Holmer Citation1994) as Eurytreta cf. sabrinae have a procline rather than an apsacline ventral pseudointerarea and are not considered conspecific with the material described above.
aff. Eurytreta sp.
C, F, G
Material. – Three dorsal valves.
Description. – All recovered dorsal valves are fragmentary with only their apical part preserved. External valve surfaces recrystallized showing no features except for one valve, which displays remnants of larval shell ornamentation consisting of hemispherical pits, 2.6–3.5 µm in diameter. Valve interior with well-developed pseudointerarea characterized by broadly triangular median groove and distinct propareas. Subrectangular median buttress with steep anterior slope accompanied directly laterally by cardinal muscle scars. Faint swelling on valve floor anterior to median buttress (G) indicates presence of a median ridge.
Remarks. – The three acrotretid dorsal valves are generally too poorly preserved to establish a proper genus affinity. However, it is likely that they have a taxonomic connection to one of the other Jillesnåle taxa, whose description is based on recovered ventral valves, i.e., Eurytreta cf. sabrinae or Ottenbyella sp. (see above and below). Dorsal valves of Eurytreta and Ottenbyella can appear quite similar, especially when poorly preserved. Nevertheless, the three dorsal valves are tentatively assigned to Eurytreta rather than Ottenbyella based on the strong convexity seen within the preserved apical part. The valves are not assigned to the co-occurring Eurytreta cf. sabrinae as the dorsal valve of E. sabrinae is characterized by a median buttress and dorsal ridge forming a continuous structure (see Sutton et al. Citation2000), which also has been observed in dorsal valves of Eurytreta cf. sabrinae from Oaxaquia (Streng et al. Citation2011). It is possible though, that the three dorsal valves and the ventral valves described as Eurytreta cf. sabrinae represent a distinct taxon.
Ottenbyella Popov & Holmer, Citation1994
Diagnosis. – See Popov & Holmer (Citation1994).
Type species. – Acrotreta carinata Moberg & Segerberg, Citation1906, p. 66, from the Lower Ordovician (Tremadocian) Bjørkåsholmen Formation, Scania, southern Sweden.
Other species included. – See Streng et al. (Citation2011).
Remarks. – Ottenbyella is distinguished from Eurytreta by a high conical ventral valve with an invariably procline pseudointerarea divided by an interridge. In addition, dorsal valves of Ottenbyella are relatively flat and have typically a less distinct median division.
Occurrence. – Furongian to Early Ordovician of Baltica, Kazakhstan, Laurentia, northwestern Gondwana, and potentially Oaxaquia (see Streng et al. Citation2011 for details).
Ottenbyella sp.
E, H, I
Material. – One ventral valve.
Description. – Incomplete ventral valve with larval shell, small parts of pseudointerarea and lateral slopes, and a larger part of the anterior slope preserved (E). Valve broadly conical with procline pseudointerarea, poorly defined on one side (left side of valve) but appears to be delineated by distinct flexure on the other side. Larval shell accentuates apex of shell due to distinct change in inclination of anterior valve slope at transition from larval to the postlarval shell (I). Larval shell ornamented by poorly preserved hemispherical pits measuring 2–3 µm in diameter. External pedicle opening elliptical in outline, situated within larval shell, representing highest point of valve. Postlarval shell ornamented by poorly preserved concentric fila. Valve interior characterized by low, subrectangular apical process, which is situated some distance anterior to internal pedicle opening. Process delineated laterally by deep, narrow, subparallel, baculate vascula lateralia.
Remarks. – The observed characters agree with an accommodation of the valve in Ottenbyella. The flexure on the external shell surface of the ventral valve seemingly demarcating one side of the pseudointerarea and the elliptical outline of the external pedical opening are interpreted to be taphonomic and induced by compression. This is supported by the alignment of the flexure axis with the long diameter of the pedicle opening (E).
The ventral valve with its pronounced apex resembles ventral valves of Ottenbyella carinata as illustrated by Popov & Holmer (Citation1994, fig. 93L, M). However, the relatively broad apical process and the narrow vascula lateralia of Ottenbyella sp. differ from O. carinata and other known species of Ottenbyella.
Acrotreta Kutorga, Citation1848
Diagnosis. – See Holmer & Popov (Citation2000).
Type species. – Acrotreta subconica Kutorga, Citation1848, p. 275–276, from an unknown locality. Most likely from the upper Floian (Billingen Regional Stage) of the uppermost Leetse and lower Toila formations (Mäeküla and Päite members) near St. Petersburg (see Holmer & Popov Citation1994 for details).
Remarks. – Holmer & Popov (Citation1994) revised the genus by re-describing the type species A. subconica Kutorga, Citation1848.
aff. Acrotreta sp.
J, K, L
Material. – Two dorsal valves; one questionable ventral valve.
Description. – External dorsal surface poorly preserved but fine concentric fila locally present. Dorsal interior with anacline pseudointerarea characterized by broad, triangular and relatively long median groove; propareas not seen. Median groove supported anteriorly by median buttress from which a blade-like, triangular median septum arises. Large cardinal muscle scars incompletely preserved lateral to median buttress.
Remarks. – Affinity of the two dorsal valves to Acrotreta is based on their similarity with dorsal valves of Acrotreta korynevskii Holmer & Popov, Citation1994 from the South Urals (Holmer & Popov Citation1994; Popov & Holmer Citation1994). A single ventral valve resembling the apical tip of Acrotreta (L) might support this affiliation.
FAMILY EPHIPPELASMATIDAE Rowell, Citation1965
New diagnosis. – Shell with relatively narrow straight or slightly convex posterior margin and postlarval shell commonly ornamented with concentric rugae. Larval shell ornamented by small, hemispherical imprints of about uniform size. Ventral valve usually narrow and highly conical with a pseudointerarea that is commonly catacline to strongly apsacline, but may be procline. Pseudointerarea poorly defined laterally, variably divided by intertrough or interridge or undivided. External pedicle foramen within larval shell, typically at end of short pedicle tube. Ventral interior characterized by apical process developed as pedicle tube along the posterior slope. Pedicle tube may be supported anterolaterally by shell layers of increased thickness causing an occlusion of the apex. Dorsal valve with distinct pseudointerarea of variable length divided by broad median groove.
Remarks. – A new diagnosis for the family Ephippelasmatidae was necessary as many species have been accommodated within the family that actually did not fulfill the diagnosis as, e.g., provided by Holmer & Popov (Citation2000). Genera currently considered as Ephippelasmatidae are quite diverse when it comes to shape of ventral valves as well as the development of the apical process and the dorsal median septum. This diversity might be partly due to the fact that some ephippelasmatid taxa represent transitional forms between the Acrotretidae and Ephippelasmatidae. Thus, e.g., the typical concentric rugae are less developed in Akmolina Popov & Holmer, Citation1994, which also has a broadly conical ventral valve rather than a highly conical one. Accordingly, the new family diagnosis has to be broad in some aspects. Shell ultrastructure and type of pitting of the larval shell have not been universally reported for all ephippelasmatid taxa. However, it is here implied that taxa of the family have a columnar ultrastructure, as indicated by previous studies (e.g., Holmer Citation1989; Popov & Holmer Citation1994), and comprise a larval shell that is ornamented by small, hemispherical pits of about uniform size, i.e., ca. 1.5–3 µm in diameter (B, F, G).
Genus Pomeraniotreta Bednarczyk, Citation1986
Diagnosis. – Ventral valve conical with steeply procline to apsacline pseudointerarea, which is poorly defined laterally and typically undivided. Apical process fills apex. Dorsal valve flattened, with simple anacline pseudointerarea lacking propareas but with wide and long, shallow median groove. Pseudointerarea supported by robust, rectangular median buttress. Median ridge low and indistinct if present.
Type species. – P. biernatae Bednarczyk, Citation1986 from the Lower Ordovician (Paroistodus proteus Biozone) of the Białogóra 1 borehole, northern Poland.
Other species included. – Anatreta transversa Mei, Citation1993 from the Furongian Fengshan Formation, Hebei, North China; P. holmeri Mergl, Citation1995 from the Lower Ordovician (Floian) of Bohemia.
Remarks. – As already noted by Mergl (Citation1995, Citation2002), the Bohemian P. holmeri is very similar to the nominate species. According to Mergl (Citation2002), P. holmeri is distinguished from P. biernatae by a less slender ventral valve, which also has a more convex anterior slope near the apex, and a dorsal valve with a less pronounced posterior margin, i.e., a less posteriorly projecting larval shell. However, these characters appear quite variable within the specimens of P. biernatae from southern Sweden (Popov & Holmer Citation1994) and might not justify a distinction into two species. The Swedish specimens discussed by Popov & Holmer (Citation1994) show an apical anterior slope of the ventral valve that can be strongly convex, relatively straight, or even concave (compare Popov & Holmer Citation1994, figs. 103L, 104F, 103N). The extent of the dorsal larval shell beyond the posterior shell margin is hard to assess without detailed measurements, but some potentially ontogenetic variation can be seen between small and larger valves in the Swedish material, with smaller valves having a less pronounced posterior margin (compare Popov & Holmer Citation1994, fig. 103M and fig. 103A, 103J). Overall, illustrated specimens of dorsal valves in Popov & Holmer (Citation1994) and Mergl (Citation1995, Citation2002) appear comparable and indistinguishable from each other. Also, the slenderness of the ventral valve is hard to assess without detailed measurements and ontogenetic series, for which complete valves are necessary. It appears that ventral valves assigned to P. holmeri increase quicker in diameter and have a more conical appearance in comparison to the Swedish material. However, the majority of these ventral valves is fragmented along their dorsal margin, i.e., they represent the apical part of originally larger specimens. Larger ventral valves of P. holmeri (Mergl Citation1995, pl. 3, fig. 8b; Mergl Citation2002, pl. 34, fig. 4) appear slenderer in comparison and more similar to ventral valves of P. biernatae. Hence, it appears reasonable to consider P. holmeri as a junior synonym of P. biernatae. However, we refrain from synonymizing the two taxa as the ventral interior is unknown from the type material of B. biernatae as well as from P. holmeri. Furthermore, the ventral larval shell of P. holmeri has a consistently more pronounced external pedicle tube with a distinctly concave posterior and anterior slope in lateral view. Respective slopes in P. biernatae are straight and weakly concave.
Popov & Holmer (Citation1994) considered the late Cambrian Anatreta transversa Mei, Citation1993 of North China as a species of Pomeraniotreta. As A. transversa is the type species of Anatreta Mei, Citation1993, the genus is a subjective junior synonym of Pomeraniotreta. However, a second species assigned to Anatreta by Mei (Citation1993), A. cava, might rather be a species of Quadrisonia (see Popov & Holmer Citation1994). The diagnosis of Pomeraniotreta has been adjusted to accommodate characters of P. transversa.
Occurrence. – Furongian (late Jiangshanian) to Early Ordovician (early Floian) of North China, Baltica and Bohemia.
Pomeraniotreta biernatae Bednarczyk, Citation1986
Figures 5A–J, L–N, 6A, B, D–F
p 1971 “acrotretacean brachiopod”, Poulsen, pl. 1, figs. 1, 2, pl. 2, fig. 1; non pl. 2, figs. 2–4, non pls. 3–6.
1979 Torynelasma lebaensis sp. n., Bednarzcyk, Citation1979, p. 415 [nomen nudum; ICZN, Article 13.1].
1986 Pomeraniotreta biernati [sic] n. sp., Bednarczyk, p. 415–416, text-fig. 2, pl. 2, figs. 1–3, pl. 3, figs. 1–3.
1994 Pomeraniotreta biernatae Bednarczyk, Citation1986: Popov & Holmer, figs. 103, 104.
? 1993 Pomeraniotreta sp., Puura & Holmer, 1993, fig. 7I, J.
? 2002 Pomeraniotreta sp., Holmer & Biernat, 2002, fig. 12B, E.
Material. – About 38 ventral and 35 dorsal valves, most of them incomplete and often tectonically deformed.
Description. – Shell strongly ventribiconvex, typically about circular in outline but transversely ovoid and elongate ovoid valves have been observed as well (L–N). Larval shell ornamented by small, hemispherical imprints of similar size measuring 1.7–2.6 µm in diameter; postlarval shell with fine concentric rugae. Ventral valve highly conical, with steeply procline to apsacline pseudointerarea and steep lateral slopes; anterior slope slightly less steep; pseudointerarea seemingly undivided and poorly defined laterally. Larval shell also conical in shape with a straight, apsacline posterior slope and an initially concave but distally convex anterior slope (D, E). Short external pedicle tube forms apex of larval shell. External pedicle opening entirely within larval shell, circular in outline, measuring 26–31 µm in diameter (n = 5). Internal features indistinct beside of apical process, which is constructed of thick columnar laminae that fill up apical cavity (F, 6E). Process penetrated by pedicle tube which runs along posterior valve slope (F). Dorsal valve flat to slightly concave, characterized by prominent larval shell that seems to project past the posterior margin of the shell. Larval shell convex centrally but with brim like, dorsally folded lateral and posterior margin; anterior margin with transition to postlarval shell less pronounced (I, L). Outline of larval shell in accordance with the respective shell outline, i.e., transversely oval, circular, or elongate oval, having a mean diameter of 180–220 µm (n = 4). Dorsal interior with long pseudointerarea lacking propareas but with broad, anacline median groove, which is supported anteriorly by distinct subrectangular median buttress. Cardinal muscle scars large, occupying space between buttress and posterolateral shell margin. Low median ridge somewhat anterior to median buttress. Other internal features not seen.
Figure 5. A–J, L–N. Pomeraniotreta biernatae Bednarczyk, Citation1986; A, ventral valve (PMU38380), posterior view showing undivided pseudointerarea; B, ventral valve (PMU38377), anterior view; C, ventral valve (PMU38379), oblique lateral view; D, E, apical portions of two ventral valves (PMU38367, PMU38384) showing profile of larval shell (for better display purposes E has been mirrored digitally); F, ventral valve (PMU38378), internal view showing apical process and pedicle tube; G, dorsal valves (PMU38368) with transversely oval larval shell; H, slightly concave dorsal valve (PMU38370) in lateral view, I, dorsal valve (PMU38373) with elongate oval larval shell; J, apical view of D; L, dorsal view of H; M, dorsal valve (PMU38371) interior view with subrectangular median buttress and faint interridge; N, dorsal valve (PMU38372), secondarily deformed and elongated. K, O, P, Ventral valve of Pomeraniotreta n. sp. (PMU38387) in posterior, right lateral, and left lateral view; note faint intertrough in K. Scale bar equals 100 µm.

Remarks. – Despite the poor preservation, the specimens from Jillesnåle can confidently be identified as P. biernatae. Specimens match the description and illustrations of specimens previously published from Baltica (Poulsen Citation1971; Popov & Holmer Citation1994). The inconsistent shapes of especially the dorsal valves are due to tectonically induced stretching. A primary, about circular commissural outline is understood for all specimens of P. biernatae from Jillesnåle.
The specimens of P. biernatae described by Popov & Holmer (Citation1994) from the Oslo Region, Västergötland, and Öland are all derived from what they called the Bjørkåsholmen Limestone. In modern terminology, this limestone is equivalent to the Bjørkåsholmen Formation (Owen et al. Citation1990) in the Oslo Region and Västergötland and the basal Köpingsklint Formation (Stouge Citation2004) on Öland (see, e.g., Pärnaste et al. Citation2013). The type level of P. biernatae, i.e., an interval with marly limestones of the Białogóra 1 borehole in northern Poland (Bednarczyk Citation1986), was redescribed by Modliński & Szymański (Citation1997) as the Ordagowo Limestone Member of the Słuchowo Shale with Glauconite Formation.
Occurrence. – Early Ordovician of Baltica. Middle Tremadocian (P. deltifer conodont biozone) of Bjørkåsholmen Formation of the Oslo Region, Norway (Poulsen Citation1971; Bednarzczyk Citation1986; Popov & Holmer Citation1994) and Västergötland, Sweden (Popov & Holmer Citation1994), and basal Köpingsklint Formation of Öland, Sweden (Popov & Holmer Citation1994); early Floian of Ordagowo Limestone Member of the Słuchowo Shale with Glauconite Formation (Bednarczyk Citation1986).
Pomeraniotreta n. sp.
Figure 5K, O, P, 6C, G
Material. – Two ventral valves.
Description. – Valve highly conical, with steep, apsacline pseudointerarea and steep, straight lateral slopes; anterior slope slightly less steep, gently convex in lateral view; pseudointerarea poorly defined laterally, divided by faint intertrough (K). Larval shell high, conical in shape with straight, apsacline posterior slope and straight lateral and anterior slopes, ornamented by hemispherical pits measuring 2.3–3 µm in diameter; height of larval shell measures 170 µm, slightly less than its diameter (ca. 180–190 µm) (P, 6C). External pedicle tube at apex of larval shell, circular in outline, measuring 22–27 µm in diameter (n = 2). Internal features poorly preserved, one specimen with remnants of thick laminae in apical cavity indicating the apical process. Dorsal valve unknown.
Remarks. – The two specimens are similar to P. biernatae, but differ in the shape of the larval shell and by having a faint intertrough. Larval shells of P. biernatae are low conical in comparison (A vs. C) and have a distinct concavity on their anterior slope. The larval shell appears to be also the main distinction between P. biernatae and P. holmeri (see Remarks above), hence, it is likely that the two specimens from Jillesnåle represent a new species of Pomeraniotreta that is characterized by its highly conical larval shell and a weakly expressed intertrough.
Figure 6. A, B, Ventral and dorsal larval shell of Pomeraniotreta biernatae (PMU38366, PMU38370). C, Ventral larval shell of Pomeraniotreta n. sp. (PMU38387, same specimen as in K, O, P). D, Columnar shell structure of P. biernatae seen in a dorsal valve (PMU38369). E, Anterior view of exfoliated ventral valve of P. biernatae exposing thick laminae of apical process (PMU38378, same specimen as in F). F, Larval shell pitting in a dorsal valve of P. biernatae (PMU38373, same specimen as in I). G, Larval shell pitting in a ventral valve of Pomeraniotreta n. sp. (PMU38387, detail of P). H, Recrystallized shell laminae of Mytoella? n. sp. (PMU38359; detail of C). Scale bar equals 50 µm (A, B, C, E), 10 µm (D, F), and 5 µm (G, H). Note that A, C and E are depicted to scale.
PHYLUM CHORDATA
INFRAPHYLUM VERTEBRATA Lamarck, Citation1801
CLASS CONODONTA Eichenberg, Citation1930
Remarks. – The insoluble remains from the dissolved calcareous concretion yielded a small conodont fauna including six isolated euconodont elements. Five of these were broken but identifiable while the remaining element was unidentifiable. Because of the very sparse and poorly preserved material at hand, the taxonomical descriptions have been limited to brief paleontological notes. The family-level classification follows that of Sweet (Citation1988).
Conodonts from Ordovician strata of the Lower Allochthon nappes of the Scandinavian Caledonides were previously described by, e.g., Bergström (Citation1971, Citation1988), Bergström et al. (Citation1974), Bruton et al. (Citation1989), Rasmussen & Stouge (Citation1989, Citation1995, Citation2018), Rasmussen (Citation2001), Pålsson et al. (Citation2002) and Rasmussen et al. (Citation2021b). Although Tremadocian conodonts are common and well documented from the Baltoscandian inner shelf deposits, especially the Bjørkåshomen Formation, records from the Scandinavian Caledonides are very sparse and include those of Bruton et al. (Citation1989) and Rasmussen (Citation2001) from the Ørnberget and Bjørkåsholmen formations of Norway.
FAMILY ACANTHODONTIDAE Lindström, Citation1970
Drepanodus Pander, Citation1856
Remarks. – Apparatus is quinquemembrate and consists of five different elements (Löfgren & Tolmacheva Citation2003).
Drepanodus arcuatus Pander, Citation1856
A, B
2001 Drepanodus arcuatus Pander, Citation1856: Rasmussen, pp. 70–71, pl. 5: 4–6 (cum. syn.).
Figure 7. Euconodont elements. A, B, Geniculate coniform element of Drepanodus arcuatus Pander, Citation1856 in lateral and lower view (PMU38390). C, D, Nongeniculate coniform element of Drepanoistodus aff. amoenus (Lindström, Citation1955) sensu Löfgren (Citation1994) in lateral and lower view (PMU38381). E–I, Two elements of Paroistodus numarcuatus (Lindström, Citation1955); nongeniculate coniform element in lateral and lower view (PMU38383) (E, F) and geniculate coniform element in lateral, upper and lower view (PMU38388) (G–I). J–M, Asymmetric nongeniculate coniform (acontiodiform) element of Rossodus aff. manitouensis Repetski & Ethington, Citation1983 in upper (J), posterior (K), lower (L), and anterior (M) view (PMU 38389). N, O, Nongeniculate coniform element of Gen. and sp. indet. in lateral and lower view (PMU38382). Note: Outlines of broken cusps in upper view from respective species illustrated in A, C, E and N. Scale bar equals 100 µm.

2003 Drepanodus arcuatus Pander, Citation1856: Löfgren & Tolmacheva, pp. 211–215, figs. 2, 3A–C, E-H, 5K-V, 6M-U, 7H-N, 8A-G (cum. syn.).
Material. – One specimen.
Remarks. – The illustrated specimen is a geniculate coniform element.
Occurrence. – From the Tremadocian Paltodus deltifer Zone to the Darriwilian Pygodus serra Zone (Löfgren & Tolmacheva Citation2003) or even the uppermost Darriwilian part of the Pygodus anserinus Zone (Rasmussen Citation2001).
FAMILY DREPANOISTODONTIDAE Fåhraeus and Nowlan, Citation1978
Drepanoistodus Lindström, Citation1971
Remarks. – Apparatus is quinquemembrate and comprises four nongeniculate and one geniculate elements (e.g., Stouge & Bagnoli Citation1990; Rasmussen et al. Citation2021a).
Drepanoistodus aff. amoenus (Lindström, Citation1955) sensu Löfgren (Citation1994)
C, D
1955 Drepanodus amoenus sp. n., Lindström, p. 558, pl. 2, fig. 25 (only).
1985 Drepanoistodus cf. forceps (Lindström, Citation1955): Löfgren, 1985, p. 126, fig. 4AF.
1988 Drepanoistodus cf. forceps (Lindström, Citation1955): Bagnoli & Stouge in Bagnoli et al., p. 210, pl. 39, figs. 1–3.
1994 Drepanoistodus aff. D. amoenus (Lindström, Citation1955): Löfgren, p. 1365, figs. 6.10–6.12, 6.17, 6.18.
Material. – One specimen.
Remarks. – The illustrated element is a nongeniculate coniform element with an antero-basal, triangular flare pointing downward in the opposite direction of the cusp. Both the upper part of the cusp and the posterior part of the base are missing, but the rounded lateral sides of the cusp are characteristic for this species. Probably, it is identical with the nongeniculate element shown by Löfgren (Citation1994, fig. 6.12). As pointed out by Löfgren (Citation1994, p. 1365) in her careful taxonomic discussion, the present species is not conspecific with the holotype of “Drepanodus” amoenus illustrated by Lindström (Citation1955, pl. 2, fig. 26), which seems to be a nongeniculate element belonging to Paroistodus numarcuatus, but probably represents a separate species within Drepanoistodus. Because a formal revision and description of this taxon is needed and the single specimen at hand is broken, we have tentatively decided to keep the taxon name in open nomenclature as Drepanoistodus aff. amoenus (Lindström Citation1955) sensu Löfgren (Citation1994). See descriptions by Bagnoli & Stouge in Bagnoli et al. (Citation1988; geniculate element) and Löfgren (Citation1994; nongeniculate elements).
Occurrence. – Mid-Tremadocian, from the upper part of the Paltodus deltifer Zone to the D. aff. amoenus Subzone, which is the lowermost subzone of the Paroistodus proteus Zone (Löfgren Citation1996).
Paroistodus Lindström, Citation1971
Remarks. – Apparatus is basically bimembrate and consists of a geniculate, coniform element together with nongeniculate coniform elements, which make a curvature transition series. Löfgren (Citation1997a) distinguished between one geniculate and six nongeniculate elements.
Paroistodus numarcuatus (Lindström, Citation1955)
E–I
1997a Paroistodus numarcuatus (Lindström, Citation1955): Löfgren, pp. 921–922, text-figs. 2O–U, 3A–G, 4A–K (cum. syn.).
2001 Paroistodus numarcuatus (Lindström, Citation1955): Rasmussen, pp. 106–108, pl. 12:13–17 (cum. syn.).
Material. – Two specimens.
Remarks. – The present material comprises one nongeniculate element (E, F) and one geniculate element (G–I). The nongeniculate element is characterized by a recurved cusp and a short base and is similar to the specimen figured by Bruton et al. (Citation1989; Q) from the Tremadocian Ørnberget Formation of the Norwegian Caledonides. The geniculate element is broken but is characterized by a pointed antero-basal extension, which is comparable to that of the specimen shown by Löfgren (Citation1997a; text-fig. 4A). See Löfgren (Citation1997a) for description.
Occurrence. – Mid-Tremadocian, from the upper part of the Paltodus deltifer Zone to the lowermost subzone of the Paroistodus proteus Zone, the D. aff. amoenus Subzone (Löfgren Citation1997a, Citation1997b).
FAMILY OISTODONTIDAE Lindström, Citation1970
Rossodus Repetski & Ethington, Citation1983
Remarks. – Apparatus is quadrimembrate (Repetski & Ethington Citation1983).
Rossodus aff. manitouensis Repetski & Ethington, Citation1983
J–M
aff. 1983 Rossodus manitouensis sp. n., Repetski & Ethington, pp. 293–300, figs. 1A–V, 2A–T, 3A–R, 4A–D.
aff. 2020 Rossodus manitouensis Repetski & Ethington, Citation1983: Zhang, figs. 9.2–9.9.
Material. – One specimen.
Remarks. – The specimen is an asymmetric coniform (acontiodontiform) element. It is broken but is characterized by a concave outline of the cusp on the posterior side, which is similar to that of the Rossodus manitouensis Repetski & Ethington elements shown by Landing et al. (Citation2012, fig. 6q) and Zhang (Citation2020, figs. 9.2 and 9.6). The specimen at hand may differ from R. manitouensis sensu stricto by a weaker developed carina on both the posterior and anterior sides of the cusp although the specimen is too fragmented to confirm this safely. The genus Rossodus and the species R. manitouensis occur widespread on the Laurentia paleocontinent but only sporadically in Baltica (Löfgren et al. Citation1999).
Occurrence. – Rossodus manitouensis occurs rarely within the Paltodus deltifer Zone of Baltica (Sweden and Norway) slightly below or within the Bjørkåsholmen Formation (Löfgren et al. Citation1999).
Family indet.
Gen. and sp. indet.
N, O
Remarks. – The specimen is a fragmented nongeniculate coniform element, where parts of the cusp and the base are missing. The poor preservation does not allow for an identification even to the family level.
Acknowledgments
We are indebted to Professor Reinhard Greiling, Karlsruhe Institute of Technology, who found the carbonate concretion and donated it for this study. Git Klintvik Ahlberg, Lund University, is thanked for processing the sample in the acid lab in Lund. MS acknowledges financial support from the Swedish Research Council. Valuable comments by two anonymous reviewers improved the initial manuscript.
Disclosure statement
No potential conflict of interest was reported by the author(s).
Additional information
Funding
References
- Ahlberg, P. & Bergström, J., 1983: Lower Cambrian trilobites from southern Swedish Lapland. Geologiska Föreningens i Stockholm Förhandlingar 104, 241–246.
- Andersson, A., Dahlman, B., Gee, D.G. & Snäll, S., 1985: The Scandinavian alum shales. Sveriges Geologiska Undersökning Ca56, 1–50.
- Bagnoli, G., Stouge, S.S. & Tongiorgi, M., 1988: Acritarchs and conodonts from the Cambro-Ordovician Furuhäll (Köpingsklint) section (Öland, Sweden). Rivista Italiana de Paleontologica e Stratigrafia 94, 163–248.
- Bednarczyk, W., 1979: Upper Cambrian to Lower Ordovician conodonts of the Łeba Elevation, NW Poland, and their stratigraphic significance. Acta Geologica Polonica 29, 409–442.
- Bednarczyk, W., 1986: Inarticulate brachiopods from the Lower Ordovician in northern Poland. Annales Societatis Geologorum Poloniae 56, 409–418.
- Bednarczyk, W., 1999: Significance of the genus Thysanotos Mickwitz 1896 for the Ordovician stratigraphy of east-central Europe. Bulletin of the Polish Academy of Sciences, Earth Sciences 47, 15–25.
- Bergström, S.M., 1971: Conodont biostratigraphy of the Middle and Upper Ordovician of Europe and eastern North America. In W.C. Sweet & S.M. Bergström (eds.): Symposium on conodont biostratigraphy. Geological Society of America Memoir 127, 83–161.
- Bergström, S.M., 1988: On Pander’s Ordovician conodonts: distribution and significance of the Prioniodus elegans fauna in Baltoscandia. Senckenbergiana lethaea 69, 217–251.
- Bergström, S.M., Riva, J. & Kay, M., 1974: Significance of conodonts, graptolites, and shelly faunas from the Ordovician of western and north-central Newfoundland. Canadian Journal of Earth Sciences 11, 1625–1660.
- Bergström, S.M., Chen, X., Gutiérrez-Marco, J.C. & Dronov, A.V., 2009: The new chronostratigraphic classification of the Ordovician System and its relations to major regional series and stages and δ13C chemostratigraphy. Lethaia 42, 97–107.
- Bruton, D.L., Harper, D.A.T. & Repetski, J.E., 1989: Stratigraphy and faunas of the Parautochthon and Lower Allochthon of southern Norway. In R.A. Gayer (ed.): The Caledonide Geology of Scandinavia, 231–241. Graham & Trotman, London.
- Callaway, C., 1877: On a new area of Upper Cambrian rocks in south Shropshire with description of a new fauna. Quarterly Journal of the Geological Society of London 33, 652–672.
- Candela, Y. & Hansen, T., 2010: Brachiopod associations from the Middle Ordovician of the Oslo Region, Norway. Palaeontology 53, 833–867.
- Candela, Y., Marion, J.-M., Servais, T., Wang, W., Wolvers, M. & Mottequin, B., 2021: New linguliformean brachiopods from the lower Tremadocian (Ordovician) of the Brabant Massif, Belgium, with comments on contemporaneous faunas from the Stavelot-Venn Massif. Rivista Italiana di Paleontologia e Stratigrafia 127, 383–395.
- Cederström, P., Ahlberg, P., Babcock, L.E., Ahlgren, J., Høyberget, M. & Nilsson, C.H., 2012: Morphology, ontogeny and distribution of the Cambrian Series 2 ellipsocephalid trilobite Strenuaeva spinosa from Scandinavia. GFF 134, 157–171.
- Ebbestad, J.O.R., 1997: Bjørkåsholmen Formation (Upper Tremadoc) in Norway: regional correlation and trilobite distribution. In S. Stouge (ed.): WOGOGOB-94 Symposium. GEUS Rapport 1996/98, 27–35.
- Ebbestad, J.O.R., 1999: Trilobites of the Tremadoc Bjørkåsholmen Formation in the Oslo Region, Norway. Fossils and Strata 47, 1–118.
- Eichenberg, W., 1930: Conodonten aus dem Culm des Harzes. Palaeontologische Zeitschrift 12, 177–182.
- Eichwald, E., 1840a: O siluriyskoy sisteme plastov v estlyandii. Iz 1-y i 2-y knigi Zhurnala vrachebnykh i yestestvennykh nauk, Meditsinskoy Akademii, osobenno napechatano. 243 pp.; St. Petersburg. [On the strata of the Silurian system in Estonia. From the 1st and 2nd book of the Journal of Medical and Natural Sciences, Medical Academy, especially printed].
- Eichwald, E., 1840b: Ueber das silurische Schichtensystem in Esthland. St. Petersburg, 210 pp. https://www.biodiversitylibrary.org/item/253802
- Eichwald, E., 1840c: O siluriyskoy sisteme plastov v estlyandii. Zhurnal vrachebnykh i yestestvennykh nauk 1, 1–134, [On the strata of the Silurian system in Estonia. Journal of Medical and Natural Sciences.].
- Eichwald, E., 1840d: Kurze Anzeige einer geognostischen Untersuchung Esthlands und einiger Inseln der Ostsee. In E. Eichwald (ed.): Die Urwelt Russlands, durch Abbildungen erlæutert, Erstes Heft, 1–24. Druckerei des Journal de Saint-Petersbourg, St. Petersburg.
- Eichwald, E., 1840e: Zapiska o geognosticheskikh issledovaniyakh po beregam i na nekotorykh ostrovakh finskogo zaliva [A note on geognostic research on the shores and on some islands of the Finnish Gulf]. In E. Eichwald (ed.): Pervobytnyy mir Rossii. Tetrad’ 1 [The primitive world of Russia. Book 1], 1–15. Printing house N. Grech, St. Petersburg.
- Eichwald, E., 1841: O siluriyskoy sisteme plastov v estlyandii. Zhurnal vrachebnykh i yestestvennykh nauk 2, 1–109, [On the strata of the Silurian system in Estonia. Journal of Medical and Natural Sciences].
- Eichwald, E., 1842: Neuer Beitrag zur Geognosie Esthlands und Finnlands. In E. Eichwald (ed.): Die Urwelt Russlands, durch Abbildungen erlaeutert. Zweites Heft, 1–138. Kaiserliche Akademie der Wissenschaften, St. Petersburg.
- Eichwald, E., 1843: Neuer Beitrag zur Geognosie Esthlands und Finnlands. In K. E. von Baer (ed.): Beiträge zur Kenntnis des russischen Reiches und der angränzenden Länder Asiens 8, 1–138. Verlag der Kaiserlichen Akademie der Wissenschaften, St. Petersburg.
- Egenhoff, S., Cassle, C., Maletz, J., Frisk, ÅM, Ebbestad, J.O.R. & Stübner, K., 2010: Sedimentology and sequence stratigraphy of a pronounced Early Ordovician sea-level fall on Baltica – The Bjørkåsholmen Formation in Norway and Sweden. Sedimentary Geology 224, 1–14.
- Eliasson, T., Greiling, R.O., Sträng, T. & Triumf, C.-A., 2001a: Bedrock map 23H Stensele NV, scale 1:50 000. Sveriges geologiska undersökning. Ai126.
- Eliasson, T., Greiling, R.O., Sträng, T. & Triumf, C.-A., 2001b: Bedrock map 23H Stensele SV, scale 1:50 000. Sveriges geologiska undersökning. Ai128.
- Eliasson, T., Greiling, R.O. & Triumf, C.-A., 2003: Bedrock map 24H Sorsele SV, scale 1:50 000. Sveriges geologiska undersökning. Ai188.
- Fåhraeus, L.E. & Nowlan, G.S., 1978: Franconian (Late Cambrian) to early Champlainian (Middle Ordovician) conodonts from the Cow Head Group, western Newfoundland. Journal of Paleontology 72, 444–471.
- Frisk, Å & Ebbestad, J.O., 2008: Trilobite bio- and ecostratigraphy of the Tremadocian Djupvik and Köpingsklint formations (A. serratus trilobite Zone) on southern Öland, Sweden. GFF 130, 153–160.
- Gee, D.G., Karis, L., Kumpulainen, R. & Thelander, T., 1974: A summary of Caledonian front stratigraphy, northern Jämtland, southern Västerbotten, central Swedish Caledonides. Geologiska Föreningens i Stockholm Förhandlingar 96, 389–397.
- Ghobadi Pour, M., Kebriaee-Zadeh, M.R. & Popov, L.E., 2011: Early Ordovician (Tremadocian) brachiopods from the Eastern Alborz Mountains, Iran. Estonian Journal of Earth Sciences 60, 65–82.
- González-Gómez, C., 2005a: Linguliformean brachiopods of the Middle-Upper Cambrian transition from the Val D’Homs Formation, southern Montagne Noir, France. Journal of Paleontology 79, 29–47.
- González Gómez, A.C., 2005b: Braquiópodos linguliformes de la transición Cambro-Ordovícica en la vartiente meridional de la Montaña Negra (Languedoc, Francia). Ph. D. thesis. University of Zaragoza, Spain. 132 pp.
- Gorjansky, V.Y., 1969: Bezzamkovyye brakhiopody kembriiskikh i ordovikskikh otlozhenii severo-zapada Russkoi platformy. Materialy po geologiii poleznym iskopayemym severo-zapada RSFSR 6, 176 pp. Leningrad: Nedra. [in Russian].
- Gorjansky, V.Y. & Popov, L.E., 1985: Morfologiya, sistematicheskoe polozhenie i proiskhozhdenie bezzamkovykh brakhiopod c karbonatnoj rakovinoj [The morphology, systematic position and origin of the inarticulate brachiopods with calcareous shells]. Paleontologicheskij Zhurnal 1985(3), 3–14. (In Russian).
- Greiling, R.O. & Garfunkel, Z., 2007: An Early Ordovician (Finnmarkian?) foreland basin and related lithospheric flexure in the Scandinavian Caledonides. American Journal of Science 307, 527–553.
- Greiling, R.O. & Kathol, B., 2021: Beskrivning till berggrundskartorna 25G AMMARNÄS NV, NO, SV & SO. Sveriges Geologiska Undersökning K 681, 1–57.
- Greiling, R.O., Zachrisson, E., Thelander, T. & Sträng, T., 1999: Bedrock map 23G Dikanäs NO, scale 1:50 000. Sveriges geologiska undersökning. Ai124.
- Hadding, A., 1913: Undre Dicellograptusskiffern i Skåne jämte några därmed ekvivalenta bildningar. Lund Universitets Årsskrift, Ny Följd 9, 1–90.
- Hansen, J., 2008: Upper Ordovician brachiopods from the Arnestad and Frognerkilen formations in the Oslo-Asker District, Norway. Palaeontos 13, 1–99.
- Hansen, J. & Holmer, L.E., 2011: Taxonomy and biostratigraphy of Ordovician brachiopods from northeastern Ny Friesland, Spitsbergen. Zootaxa 3076, 1–122.
- Havlíček, V., 1982: Lingulacea, Paterinacea, and Siphonotretacea (Brachiopoda) in the Lower Ordovician sequence of Bohemia. Sborník geologických věd, paleontologie 25, 9–82.
- Holm, G., 1890: Försteningar från Lappland, insamlade af E. Mörtsell. Geologiska Foreningens i Stockholm Förhandlingar 12, 259–267.
- Holmer, L.E., 1989: Middle Ordovician phosphatic inarticulate brachiopods from Västergötland and Dalarna, Sweden. Fossils and Strata 26, 1–172.
- Holmer, L.E. & Biernat, G., 2002: Lingulate brachiopods from Lower Ordovician (Tremadoc) chalcedonites, Holy Cross Mountains, Poland. Acta Palaeontologica Polonica 47, 141–156.
- Holmer, L.E. & Popov, L.E., 1994: Revision of the type Acrotreta and related lingulate brachiopods. Journal of Paleontology 68, 433–450.
- Holmer, L.E. & Popov, L.E., 2000: Lingulata. In A. Williams et al. (ed.): Treatise on Invertebrate Paleontology. Pt. H. Brachiopoda, revised, vol. 2, 30–146. Geological Society of America and University of Kansas, Boulder, CO.
- Holmer, L.E. & Popov, L.E., 2007: Systematic descriptions: Brachiopoda. Updates To Systematic Volumes 2–5. Linguliformea. In A. Williams et al. (ed.): Treatise on Invertebrate Paleontology, Part H (Brachiopoda), Part H, Brachiopoda (revised), Vol. 6: Suppliment, 2532–2579. Geological Society of America and University of Kansas, Boulder, CO.
- Holmer, L.E., Popov, L.E., Streng, M. & Miller, J.F., 2005: Lower Ordovician (Tremadocian) lingulate brachiopods from the House Range and Fillmore Formation, Ibex area, western Utah. Journal of Paleontology 79, 884–906.
- Karis, L., 1998: Jämtlands östliga fjällberggrund. In L. Karis & A.G.B. Stromberg: Beskrivning till berggrundskartan över Jämtlands län. Del 2: Fjälldelen, 1–184. Sveriges geologiska undersökning Ca53, 1–363.
- Kautsky, F., 1945: Die unterkambrische Fauna vom Aistjakk in Lappland. Geologiska Föreningens i Stockholm Förhandlingar 67, 129–211.
- King, W., 1846: Remarks on certain genera belonging to the class Palliobranchiata. Annals and Magazine of Natural History 18, 26–42.
- Koliha, J., 1924: Atremata z krušnohorských vrstev (dα). Palaeontographica Bohemiae 10, 1–61.
- Koneva, S.P., 1986: Novoe semeistvo kembriiskikh bezzamkovykh brakhiopod [A new family of the Cambrian inarticulate brachiopods]. Paleontologicheskij Zhurnal 1986(1), 49–55.
- Kuhn, O., 1949: Lehrbuch der Paläozoologie. Schweizerbart, Stuttgart. 326 pp.
- Kulling, O., 1942: Grunddragen av fjällkedjerandens bergbyggnad inom Västerbottens län. Sveriges Geologiska Undersökning C445, 1–320.
- Kulling, O., 1955: Beskrivning till berggrundskartan över Västerbottens Län. Del 2; Den kaledoniska fjällkedjans berggrund inom Västerbottens län. Sveriges Geologiska Undersökning Ca37, 101–296.
- Kulling, O., 1960: On the Caledonids of Swedish Lapland. In Magnusson, N.H., Thorslund, P., Brotzen, F., Asklund, B. & Kulling, O.: Description to accompany the map of the pre-Quaternary rocks of Sweden. Sveriges Geologiska Undersökning Ba16, 150–177.
- Kulling, O., 1964: Översikt över norra Norrbottenfjällens kaledonberggrund. Sveriges Geologiska Undersökning Ba19, 1–166.
- Kulling, O., 1982: Översikt över södra Norrbottensfjällens kaledonberggrund. Sveriges Geologiska Undersökning Ba26, 1–295.
- Kutorga, S., 1848: Über die Brachiopoden-Familie der Siphonotretaeae. Verhandlungen der Russisch-Kaiserlichen Mineralogischen Gesellschaft zu St. Petersburg 1847, 250–286.
- Lamarck, J.B., 1801: Systême des animaux sans vertèbres, ou tableau général des classes, des ordres et des genres de ces animaux. viii + 432 pp. Chez l’auteur, Paris.
- Landing, E., Adrain, J.M., Westrop, S.R. & Kröger, B., 2012: Tribes Hill–Rochdale formations in east Laurentia: proxies for Early Ordovician (Tremadocian) eustasy on a tropical passive margin (New York and west Vermont). Geological Magazine 149, 93–123.
- Lindström, M., 1955: Conodonts from the lowermost Ordovician strata of south-central Sweden. Geologiska Foreningens i Stockholm Förhandlingar 76, 517–604.
- Lindström, M., 1970: A suprageneric taxonomy of the conodonts. Lethaia 3, 427–445.
- Lindström, M., 1971: Lower Ordovician conodonts of Europe. In W.C. Sweet & S.M. Bergström (eds.): Symposium on Conodont Biostratigraphy, The Geological Society of America Memoir 127, 21–61.
- Lilljequist, R., 1973: Caledonian geology of the Laisvall area, Southern Norrbotten, Swedish Lappland. Sveriges Geologiska Undersökning C691, 1–43.
- Löfgren, A., 1985: Early Ordovician conodont biozonation at Finngrundet, south Bothnian Bay, Sweden. Bulletin of the Geological Institution of the University of Uppsala 10, 115–128.
- Löfgren, A., 1993: Conodonts from the lower Ordovician at Hunneberg, south-central Sweden. Geological Magazine 130, 215–232.
- Löfgren, A., 1994: Arenig (Lower Ordovician) conodonts and biozonation in the eastern Siljan district, central Sweden. Journal of Paleontology 68, 1350–1368.
- Löfgren, A., 1996: Lower Ordovician conodonts, reworking, and biostratigraphy of the Orreholmen quarry, Västergötland, south-central Sweden. GFF 118, 169–183.
- Löfgren, A., 1997a: Reinterpretation of the Lower Ordovician conodont apparatus Paroistodus. Palaeontology 40, 913–929.
- Löfgren, A., 1997b: Conodont faunas from the upper Tremadoc at Brattefors, south-central Sweden, and reconstruction of the Paltodus apparatus. GFF 119, 257–266.
- Löfgren, A.M. & Tolmacheva, T.J., 2003: Taxonomy and distribution of the Ordovician conodont Drepanodus arcuatus Pander, 1856, and related species. Paläontologische Zeitschrift 77, 203–221.
- Löfgren, A., Repetski, J.E. & Ethington, R.L., 1999: Some trans-Iapetus conodont faunal connections in the Tremadocian. Bollettino della Socità Paleontologica Italiana 37, 159–173.
- Martinsson, A., 1974: The Cambrian of Norden. In Holland C.H. (ed.): Lower Palaeozoic Rocks of the World, Vol. 2, Cambrian of the British Isles, Norden, and Spitsbergen, 185–283. Wiley, London.
- Mei, S.-L., 1993: Middle and Upper Cambrian inarticulate brachiopods from Wanxian, Hebei, North China. Acta Palaeontologica Sinica 32, 400–429.
- Menke, C.T., 1828: Synopsis methodica molluscorum generum omnium et specierum earum quae in Museo Menkeano adservantur, 1–91. Pyrmonti: G. Uslar.
- Mergl, M., 1995: New lingulate brachiopods from the Mílina Formation and the base of the Klabava Formation (late Tremadoc–early Arenig), Central Bohemia. Věstník Českého geologického ústavu 70, 101–114.
- Mergl, M., 2002: Linguliformean and craniiformean brachiopods of the Ordovician (Třenice to Dobrotivá Formation) of the Barrandian, Bohemia. Acta Musei Nationalis Pragae, Series B, Historia Naturalis 58, 1–82.
- Mergl, M., Herrera, Z., Villas, E. & Ortega, G., 2015: Lingulate brachiopods from the Lampazar Formation (late Cambrian) of the Cordillera Oriental of northwestern Argentina. Journal of Paleontology 89, 537–552.
- Mickwitz, A., 1896: Über die Brachiopodengattung Obolus Eichwald. Zapiski Imperatorskoĭ akademīi nauk, po Fiziko-matematicheskomu otdielenīiu. 8e série, 1–215. [=Mémoires de l'Académie impériale des sciences de St.-Pétersbourg, Classe des sciences physiques et mathématiques. 8e série].
- Moberg, J.C. & Segerberg, C.O., 1906: Bidrag till kännedomen om ceratopygeregionen med särskild hänsyn till dess utveckling i Fogelsångstrakten. Lunds Universitets Årsskrift, Serie 2 2(7), 1–113.
- Moczydłowska, M., 1991: Acritarch biostratigraphy of the Lower Cambrian and the Precambrian-Cambrian boundary in southeastern Poland. Fossils and Strata 29, 1–127.
- Moczydłowska, M., Jensen, S., Ebbestad, J.O.R., Budd, G.E. & Martí-Mus, M., 2001: Biochronology of the autochthonous Lower Cambrian in the Laisvall-Storuman area, Swedish Caledonides. Geological Magazine 138, 435–453.
- Modliński, Z. & Szymański, B., 1997: The Ordovician lithostratigraphy of the peribaltic depression (NE Poland). Geological Quarterly 41, 273–288.
- Mörtsell, E., 1888: Resenotiser från det fossilförande kambro-siluriska området af Vesterbottens Lappmark. Geologiska Föreningens i Stockholm Förhandlingar 12, 255–258.
- Owen, A.W., Bruton, D.L., Bockelie, J.F. & Bockelie, T.G., 1990: The Ordovician successions of the Oslo Region, Norway. Norges Geologiske Undersøkelse, Special Publication 4, 1–54.
- Pålsson, C., Månsson, K. & Bergström, S.M., 2002: Biostratigraphical and palaeoecological significance of graptolites, trilobites and conodonts in the Middle–Upper Ordovician Andersö Shale: an unusual ‘mixed facies’ deposit in Jämtland, central Sweden. Transactions of the Royal Society of Edinburgh, Earth Sciences 93, 35–57.
- Pander, H.C., 1856: Monographie der fossilen Fische des Silurischen Systems der Russisch-Baltischen Gouvernements. Buchdruckerei der Kaiserlichen Akademie der Wissenschaften, St. Petersburg, 1–91. St. Petersburg.
- Pärnaste, H., Bergström, J. & Zhiyi, Z., 2013: High resolution trilobite stratigraphy of the Lower–Middle Ordovician Öland Series of Baltoscandia. Geological Magazine 150, 509–518.
- Popov, L.E. & Cocks, L.R.M., 2017: The World’s second oldest strophomenoid-dominated benthic assemblage in the first Dapingian (Middle Ordovician) brachiopod fauna identified from Iran. Journal of Asian Earth Sciences 140, 1–12.
- Popov, L. & Holmer, L.E., 1994: Cambrian-Ordovician lingulate brachiopods from Scandinavia, Kazakhstan, and South Ural Mountains. Fossils and Strata 35, 1–156.
- Popov, L.E., Khazanovitch, K.K., Borovko, N.G., Sergeeva, S.P. & Sobolevskaya, R.F., 1989: Opornye razrezy i stratigrafiia kebro-ordovikhskoy fosforitonosnoi obolovoi tolshchi na severo-zapade Russkoy platformy [The key sections and stratigraphy of the Cambrian-Ordovician phosphate-bearing Obolus beds on the north-western Russian platform]. Ministerstvo Geologii SSSR, Mezhvedomstvennyi stratigraficheskii komitet SSSR 18, 1–222.
- Popov, L.E., Ghobadi Pour, M. & Hosseini, M., 2008: Early to Middle Ordovician lingulate brachiopods from the Lashkarak Formation, Eastern Alborz Mountains, Iran. Alcheringa 32, 1–35.
- Popov, L.E., Ghobadi Pour, M., Hosseini, M. & Holmer, L.E., 2009: Furongian linguliform brachiopods from the Alborz Mountains, Iran. Memoirs of the Association of Australasian Palaeontologists 37, 103–122.
- Poulsen, V., 1971: Notes on an Ordovician acrotretacean brachiopod from the Oslo region. Bulletin of the Geological Society of Denmark 20, 265–278.
- Puura, I. & Holmer, L.E., 1993: Lingulate brachiopods from the Cambrian-Ordovician boundary beds in Sweden. Geologiska Föreningens i Stockholm Förhandlingar 115, 215–237.
- Rasmussen, J.A., 2001: Conodont biostratigraphy and taxonomy of the Ordovician shelf margin deposits in the Scandinavian Caledonides. Fossils and Strata 48, 1–180.
- Rasmussen, J.A. & Stouge, S.S., 1989: Middle Ordovician conodonts from allochthonous limestones at Høyberget, southeastern Norwegian Caledonides. Norsk Geologisk Tidsskrift 69, 103–110.
- Rasmussen, J.A. & Stouge, S., 1995: Late Arenig-Early Llanvirn conodont biofacies across the Iapetus Ocean. In J.D. Cooper, M.L. Droser & S.E. Finney (eds.): Ordovician Odyssey: Short Papers for the Seventh International Symposium on the Ordovician System. SEPM, Pacific Section, Book 77, 443–447.
- Rasmussen, J.A. & Stouge, S., 2018: Baltoscandian conodont biofacies fluctuations and their link to Middle Ordovician (Darriwilian) global cooling. Palaeontology 61, 391–416.
- Rasmussen, J.A., Eriksson, M.E. & Lindskog, A., 2021a: Middle Ordovician Drepanoistodus (Vertebrata, Conodonta) from Baltica, with description of three new species. European Journal of Taxonomy 774, 106–134.
- Rasmussen, J.A., Thibault, N. & Rasmussen, C.M.Ø., 2021b: Middle Ordovician astrochronology decouples asteroid breakup from glacially-induced biotic radiations. Nature Communications 12, 6430.
- Repetski, J.E. & Ethington, R.L., 1983: Rossodus manitouensis (Conodonta), A new Early Ordovician index fossil. Journal of Paleontology 57, 289–301.
- Ride, W.D.L., Cogger, H.G., Dupuis, C., Krauss, O., Minelli, A., Thompson, F.C. & Tubbs, P.K. (eds.), 1999: International Code of Zoological Nomenclature (Fourth Edition). International Trust for Zoological Nomenclature, London. xxix + 306 pp.
- Rowell, A.J., 1965: Inarticulata. In R.C. Moore (ed.): Treatise on Invertebrate Paleontology. Pt. H. Brachiopoda 1 (2), H260–H296. Geological Society of America and University of Kansas Press, Boulder, Kansas.
- Rowell, A.J., 1966: Revision of some Cambrian and Ordovician inarticulate brachiopods. University of Kansas, Paleontological Contributions 7, 1–36.
- Schuchert, C., 1893: A classification of the Brachiopoda. The American Geologist 11, 141–167.
- Sdzuy, K., 1955: Die Fauna der Leimnitz-Schiefer (Tremadoc). Abhandlungen der senckenbergischen naturforschenden Gesellschaft 492, 1–74.
- Stouge, S., 2004: Ordovician siliciclastics of Öland, Sweden. Erlanger geologische Abhandlungen Sonderband 5, 91–111.
- Stouge, S. & Bagnoli, G., 1990: Lower Ordovician (Volkhovian–Kundan) conodonts from Hagudden, northern Öland, Sweden. Palaeontographia Italica 77, l–54.
- Streng, M., Mellbin, B.B., Landing, E. & Keppie, J.D., 2011: Linguliform brachiopods from the terminal Cambrian and lowest Ordovician of the Oaxaquia microcontinent (southern Mexico). Journal of Paleontology 85, 122–155.
- Sturkell, E., 1991: Tremadocian Ceratopyge Limestone identified by means of conodonts, in Jämtland, Sweden. Geologiska Föreningen i Stockholm Förhandlingar 113, 185–188.
- Sutton, M.D., Bassett, M.G. & Cherns, L., 2000: Lingulate brachiopods from the Lower Ordovician of the Anglo-Welsh Basin, part 2. Monograph of the Palaeontological Society London 153, 61–114.
- Sweet, W.C., 1988: The Conodonta – Morphology, taxonomy, paleoecology, and evolutionary history of a long-extinct animal phylum. Oxford Monographs on Geology and Geophysics 10, 212 pp. Oxford: Clarendon Press.
- Tjernvik, T.E., 1956: On the early Ordovician of Sweden. Bulletin of the Geological Institutions of Uppsala 35, 108–284.
- Waagen, W.H., 1885: Salt range fossils, Part 4 (2) Brachiopoda: Palæontologia Indica. Memoirs 13, 729–770.
- Walcott, C.D., 1902: Cambrian Brachiopoda: Acrotreta; Linnarssonella; Obolus; with descriptions of new species. Proceedings U.S. National Museum 25, 577–612.
- Walcott, C.D., 1912: Cambrian Brachiopoda. Monographs of the United States Geological Survey 51, 1–872.
- van Wamel, W.A., 1974: Conodont biostratigraphy of the Upper Cambrian and Lower Ordovician of northwestern Öland, south-eastern Sweden. Utrecht Micropaleontological Bulletins 10, 1–126.
- Weidner, T., Nielsen, A.T. & Ebbestad, J.O.R., 2021: Paleozoiska fossil från den undre skollberggrunden vid Vindelälven, Västerbotten. In R.O. Greiling & B. Kathol (eds): Beskrivning till berggrundskartorna 25G AMMARNÄS NV, NO, SV & SO. Sveriges Geologiska Undersökning K 681, 16–18.
- Willdén, M.Y., 1980: Palaeoenvironment of the autochthonous sedimentary rock sequence at Laisvall, Swedish Caledonides. Stockholm Contributions in Geology 33, 1–100.
- Williams, A., Carlson, S.J., Brunton, C.H.C., Holmer, L.E. & Popov, L., 1996: A supra-ordinal classification of the Brachiopoda. Philosophical Transactions of the Royal Society of London B351, 1171–1193.
- Williams, S.H. & Lockley, M.G., 1983: Ordovician inarticulate brachiopods from graptolitic shales at Dob’s Linn, Scotland; their morphology and significance. Journal of Paleontology 57, 391–400.
- Želízko, J.V., 1921: Äquivalente der untersilurischen Euloma-Niobefauna bei Plzenec in Böhmen. Skrifter utgit av Videnskapsselskapet i Kristiania. I, Matematisk-naturvidenskabelig klasse 1921(10), 1–27.
- Zhang, S., 2020: Upper Cambrian and Lower Ordovician conodont biostratigraphy and revised lithostratigraphy, Boothia Peninsula, Nunavut. Canadian Journal of Earth Sciences 57, 1030–1047.