
ABSTRACT
The structure of the shell wall in the Ordovician Orthoceras differs from that in the extant Nautilus. It consists of two equal thick layers: an outer probably nacreous layer and an inner prismatic layer. The two layers are separated from each other by a thin intermediate sub-layer that is rich in organic substance and has indistinct boundaries. The intermediate sub-layer and the inner prismatic layer contain a hitherto undetected pore-canal network that consists of vertical and horizontal canals. Horizontal canals are arranged in a single layer in the outermost part of the intermediate sub-layer. The vertical canals are arranged in parallel rows, and the canals in each row open into one of the horizontal canals. The canals have thin walls of calcified organic fibres. The walls are preserved in the intermediate sub-layer, whereas in the inner prismatic layer, they are partially dissolved. The canal network is absent in the shell wall of the extant Nautilus, but the horizontal canals occur in the shell wall of the Carboniferous orthocerid-like coleoid Mitorthoceras and the bactritid-like coleoid Shimanskya. Thus, the pore-canal network had an important, still unknown, function for the animal, and it existed, without changes, at least 150 ma.
Material and methods
The pore-canals were studied in the genus Orthoceras (family Orthoceatidae, revised by Kröger Citation2004) from the Aseri and Lasnamägi Stages of the Middle Ordovician of Baltoscandia. Three specimens of O. regulare Schlotheim, 1820 (specimens Mo 195101, Mo 195103, Mo 195104) and one of Orthoceras sp. (specimen Mo 195105), collected at Kandel, Estonia; one of Orthoceras sp. (specimen Mo 3274), collected at Odensholm, Estonia; three of Orthoceras sp. (specimens Mo 195102, Mo 160547, Mo 154276) and one of O. scabridum Angelin, 1880 (specimen Mo 154229), collected from the localities Folkeslunda, Runsten and Stora Bäck on the Island of Öland, Sweden were investigated. The pore-canals in Orthoceras were compared with pores in the extant Nautilus pompilius Linné1758 (specimen Mo 195106) from the Solomon Islands. The material is deposited at the Swedish Museum of Natural History, Stockholm. The shell structure was studied in incipient light using a Wild Photomakroskop M400 at the Swedish Museum of Natural History, Stockholm.
Description
Shell and vertical pores in extant Nautilus
The shell in Nautilus consists of three calcareous layers: the outer spherulitic-prismatic layer, the nacreous layer, and the inner prismatic layer (B). The inner prismatic layer is only about 0.1 mm thick and contains numerous vertical pores about 0.05 mm in diameter (A). About 100 pores occur on 1.0 mm2. The pores can only be clearly distinguished at the apertural margin in fully-grown shells where the inner prismatic layer is thickened and impregnated by black organic substance (Ward Citation1987; Mutvei & Doguzhaeva Citation1997, fig. 9D, E; Pl. 8:1). The pores contain small, finger-shaped extensions from the mantle epithelium that are attached to the inner shell surface on the body-shell attachment band (Stenzel Citation1964; Doguzhaeva & Mutvei Citation1986, D; Mutvei & Doguzhaeva Citation1997, fig. 10).
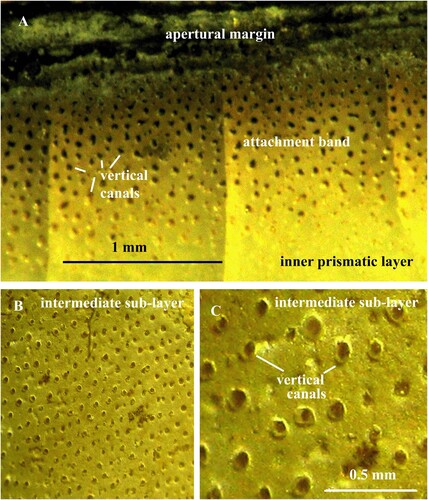
Fig. 1. A. Nautilus pompilius. Specimen no. Mo 195106, Solomon Islands. The shell-body attachment band at the apertural margin to show vertical canals that housed epithelial extensions. B, C. Orthoceras regulare. Specimen no. Mo 195101, Kandel. B. Thin, calcified, intermediate sub-layer with vertical pore-canals; the canals are arranged in parallel rows. C. Same specimen in higher magnification. Note the thin calcareous walls of the vertical canals.
Shell and pore-canal network in Orthoceras
The shell in Orthoceras consists of two equally thick layers: an outer, probably nacreous layer and an inner prismatic layer. The layers are separated by a thin intermediate sub-layer that has indistinct boundaries (A). The sub-layer is about 0.2 mm thick and consists of organic fibres (A; 10C). In most specimens, the fibres are calcified and form a thin calcareous layer (B; 3; 4). The pore-canal network occurs in the intermediate sub-layer and the inner prismatic layer and consists of vertical canals that open into horizontal canals (A).
Fig. 2. A. Orthoceras sp. Specimen no. Mo 195102, Runsten. Vertical pore-canals in the intermediate sub-layer and inner prismatic layer; the intermediate sub-layer is rich in organic substance. B. Orthoceras regulare. Same specimen as in B. Thin, calcified, intermediate sub-layer with vertical pore-canals between the nacreous and inner prismatic layers.
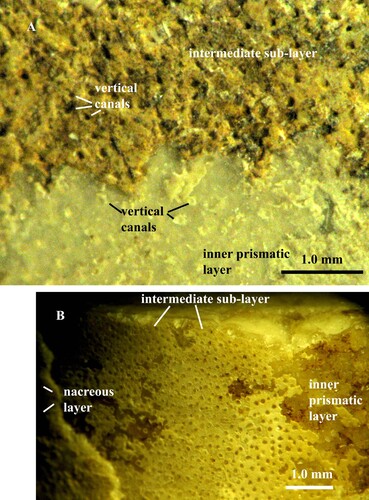
Fig. 3. A, B. Orthoceras scabridum. A, B. Specimen no. Mo 154229, Runsten. A. Intermediate sub-layer with vertical pore-canals B. Canals in higher magnification; note the granular crystalline structure of the intermediate sub-layer
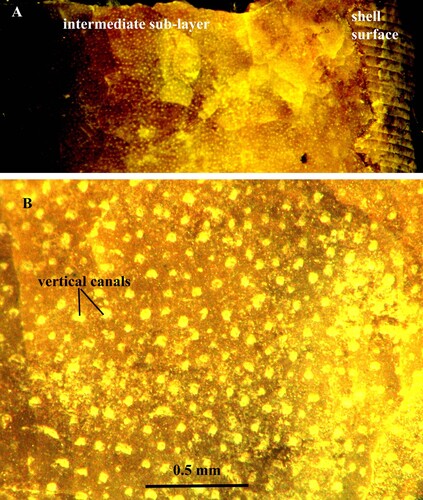
Fig. 4. A, B. Orthoceras sp. Specimen no. Mo 3274, Odensholm. A. Fragments of calcified intermediate sub-layer with numerous vertical pore-canals, arranged in rows. B. Same layer in higher magnification.

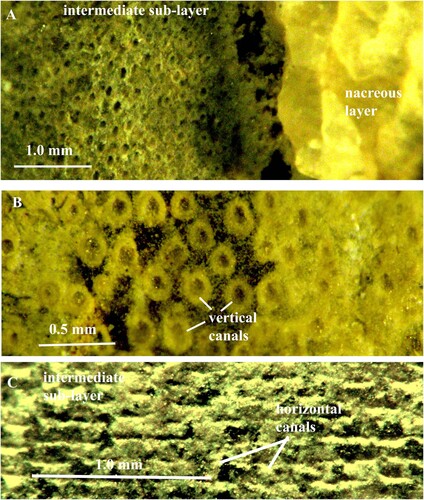
Fig. 5. A, B. Orthoceras sp. Specimen no. Mo 160547, Folkeslunda. A. Intermediate sub-layer with vertical pore-canals. B. Vertical canals in higher magnification; each canal has a wall of globular organic elements. C. Orthoceras sp. Specimen no. Mo 154276, S. Bäck. Intermediate sub-layer with horizontal pore-canals; the canal walls are partially decalcified and consist of globular organic elements; note that the canals are open and empty.
Vertical pore-canals
Already Kröger (Citation2004, p. 60) noticed that the inner surface of the shell wall in Orthoceras regulare and O. bifeovatum is “sometimes slightly irregularly punctured”. The vertical pore-canals extend from the intermediate sub-layer through the inner prismatic layer and open into the body chamber (A). The canals are arranged in parallel rows that are often oriented parallel to the shell aperture (B; 4A, B; 9A, B; 11A). The intervals between the canals in each row and between the rows vary from 0.2 to 0.1 mm. The canals in each row open into one horizontal canal (E; 9A, B; 10A; 11A). The canals have circular cross-sections and diameters between 0.05 and 0.1 mm. There are 40–130 canals in 1.0 mm2 (B; 2A, B; 3B; 4B; 5A; 10C). The canals have thin walls that are only preserved in the intermediate sub-layer (C; 5B). In the re-crystallized inner prismatic layer, the walls are partially dissolved, and the canals can be recognized as white spots or strings with irregular outlines (A; 11F). All canals are left open and empty despite of diagenesis (C; 2A, B; 3B; 5B; 10C).
Fig. 6. A, B, C, D, E. Orthoceras sp. Same specimen as in C. Intermediate sub-layer with horizontal pore-canals; note that the globular organic element in the canal walls are connected to each other by strings; the canals are open and empty; in E the vertical canals open into the horizontal canals.
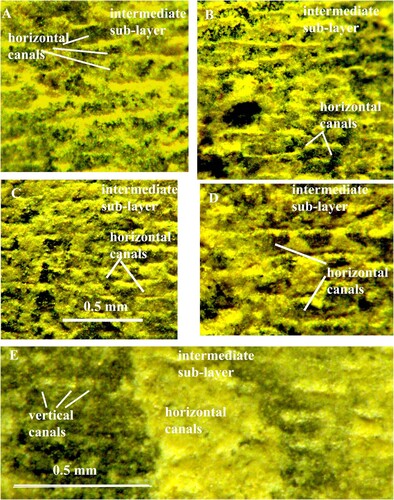
Fig. 7. Orthoceras scabridum. Same specimen as in A. Organic walls of the horizontal pore-canals in higher magnification.

Fig. 8. Orthoceras scabridum. Same specimen as in B. Organic walls of the horizontal pore-canals in higher magnification.

Fig. 9. A, B, C. Orthoceras regulare. Specimen no. Mo 195103, Kandel. A, B. Intermediate sub-layer with horizontal and vertical pore-canals; the vertical canals open into the horizontal canals. C. Intermediate sub-layer with horizontal canals. D. Mitorthoceras perfilosum, Carboniferous orthocerid-like coleoid with horizontal pore-canals (tube-like furrows). Same specimen as in Mutvei & Mapes Citation2019, C. Note the similarity of the horizontal pore-canals in Orthoceras (C) and Mitorthoceras (D).

Fig. 10. A, B. Orthoceras regulare. Specimen no. Mo 195104, Kandel. A. Intermediate sub-layer with vertical pore–canals that open into horizontal pore-canals. B. Horizontal pore-canals divided into sections of variable length. C. Orthoceras sp. Specimen no. Mo 195105, Kandel. Intermediate sub-layer with vertical pore-canals; note that the sub-layer consists of organic particles.
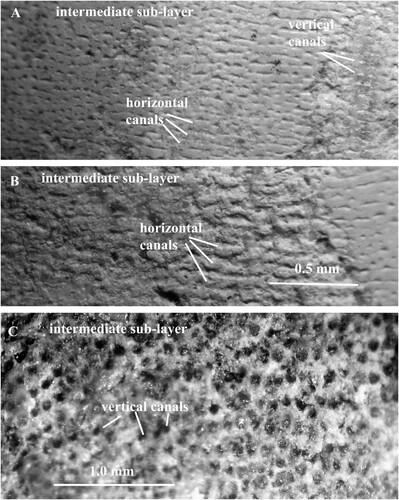
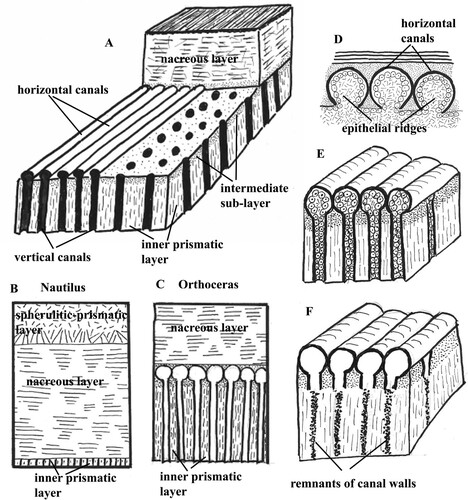
Fig. 11. Schematic presentation of the pore-canals. A. Section of the shell wall in Orthoceras to show arrangement of the horizontal and vertical pore-canals. B. Shell wall in the extant Nautilus. The inner prismatic layer is thin and contains short pores. C. Shell wall in Orthoceras. The inner prismatic layer is thick and contains the pore-canal network that is absent in Nautilus. D. Epithelial ridges on the mantle surface that secreted the walls of the horizontal pore-canals in Orthoceras. E. Horizontal and vertical pore-canals in Orthoceras before diagenesis. F. Horizontal and vertical pore-canals after diagenesis; the walls of the vertical canals are partially dissolved in the inner prismatic layer.
Horizontal pore-canals
The horizontal pore-canals are less frequently preserved than the vertical canals. They form a single layer that is situated in the outermost part of the intermediate sub-layer. The canals have mostly straight course, often parallel to the apertural shell margin (C; 6; 9A–C; 11A). In a shell of O. regulare, the horizontal canals are not continuous but divided into sections of variable length (B). The adjacent canals have usually equal dimensions, and the lateral sides of their walls are in contact to each other (C; 9A, C). There are 7–13 canals in one mm. As in the vertical canals, the cavities in the horizontal canals are left open and empty (C; 6D). Each canal has a thin wall that is composed of calcified organic fibres and has a finely granular structure (A–C). In the canal walls that were partially or completely decalcified during diagenesis, the fibres are well preserved and consist of small globular elements, connected to each other with short strings (C; 6A–D; 7; 8). All canals are left open and empty despite diagenesis.
Discussion
The pore-canal network in Orthoceras has hitherto not been detected despite numerous studies of the shell structure. This depends on the unusual mode of preservation of the canals. The canals were only detected in the organic-rich intermediate sub-layer in longitudinally broken shell walls. The conditions required to find the pore-canals are therefore highly variable and still poorly understood.
Walls of horizontal and vertical canals consist of calcified organic fibres that were secreted by epithelial extensions from the mantle (E). It is difficult to understand the formation of the horizontal canals. The mantle must have formed parallel epithelial ridges, each of which secreted the wall of a horizontal canal (D). Similar epithelial ridges also secreted the walls of the horizontal canals (termed “tube like furrows”, “horizontal rods”) in the Carboniferous coleoids Mitorthoceras (D; Mutvei & Mapes Citation2019, B) and Shimanskya (Mutvei et al. Citation2012). Then the mantle formed finger-like, vertical, epithelial extensions that were connected to the horizontal extensions and secreted the walls of the vertical pore-canals as the inner prismatic layer was deposited. The vertical canals extend from the intermediate sub-layer through the inner prismatic layer and open into the body chamber (A). In the re-crystallized inner prismatic layer, the canal walls were partially dissolved and the canals can be recognized as dots or strings of irregular shapes and sizes (A; 11F). During the growth of the shell, the openings of the vertical canals into the body chamber were successively covered and closed by the myo-adhesive layer in the posterior part of the body chamber.
Conclusion
As described in previous papers (Mutvei Citation2002, Citation2015a, Citation2015b), the siphuncular structure in Orthoceras is of the calciosiphonate type. The chitinous membranes in the connecting ring are partially calcified and perforated by numerous pores. This indicates that the animal used its siphuncle to change the volumes of gas and liquid in the shell chambers for vertical migrations. Because of its long shell, Orthoceras was not capable of jet-powered swimming (Mutvei Citation2002). As a contrast, the extant Nautilus swims with the aid of jet propulsions and has a different, simpler, siphuncular structure. The chitinous membranes in the connecting ring do not contain pores (Mutvei Citation2010), and the animal uses the siphuncle mainly to maintain neutral buoyancy (Ward Citation1987; Ward & Greenwald Citation1981; Dunstan et al. Citation2011). It is probable that the pore-canals in Orthoceras contributed to the regulation of buoyancy for vertical migrations. If the canals contained low- density liquid, the increase of the buoyancy of the shell would be small, and only the vertical canals could have filled this function. If the pore-canals contained gas, the animal could have attained neutral buoyancy without the aid of cameral liquid. This would explain why the canals have too complicated structures and why they existed without changes for a long period of time. More structural studies of the shell wall are needed to understand the function of the pore-canals.
Acknowledgements
The author thanks Dr. D.J. Peterman, Wright State University, Bayton, USA, and the unnamed reviewer, for valuable comments.
Disclosure statement
No potential conflict of interest was reported by the author(s).
References
- Doguzhaeva, L.A. & Mutvei, H., 1986: Functional interpretation of inner shell layers in triassic ceratid ammonites. Lethaia 19, 195–209.
- Dunstan, A.J., Ward, P.D., Marshall, N.J. & Solan, M., 2011: Vertical distribution and migration patterns of Nautilus pompilius. PLoS ONE 6(2), e16311.
- Kröger, B., 2004: Revision of middle Ordovician orthoceratacean nautiloids from Baltoscandia. Acta Palaentologica Polonica 49, 57–74.
- Mutvei, H., 2002: Connecting ring structure and its significance for classification of the orthoceratid cephalopods. Acta Palaeontologica Polonica 47, 157–168.
- Mutvei, H., 2015a: Characterization of two new superorders nautilosiphonata and calciosiphonata and a new order cyrtocerinida of the subclass nautiloidea; siphunal structure in the ordovicin nautiloid bathmoceras (cephalopoda). GFF 137, 164–174.
- Mutvei, H., 2015b: Siphuncular structure in calciosiphonate nautiloid orders actinocerida, orthocerida and barrandeocerida (cephalopoda). GFF 138, 1–11.
- Mutvei, H. & Doguzhaeva, L.A., 1997: Shell structure and ontogenetic growth in Nautilus pompilius L. (mollusca,: cephalopoda). Palaeontogrphica 246, 33–52.
- Mutvei, H. & Mapes, R.A., 2019: Carboniferous orthocerid-like coleoid Mitorthoceras with unique shell structure (new order tuborthocerida, coleoidea, cephalopoda); comparison with bactritid-like coleoid Shimanskya. GFF 141, 121–132.
- Mutvei, H., Dunca, E. & Weitschat, W., 2010: Siphuncular structure in the recent Nautilus, compared with that in mesozoic nautilids and ammonoids from Madagascar. GFF 132, 161–166.
- Mutvei, H., Mapes, R.H. & Doguzhaeva, L.A., 2012: Shell structures in Carboniferous bactritid-like coleoids (Cephalopoda) from south central USA. GFF 134, 201–216.
- Stenzel, H.B., 1964: Living Nautilus. In R.C. Moore (ed.): Treatise on Invertebrate Paleontology, Part K, (Mollusca 3), K59–K93, Geological Society of America and University of Kansas Press. Boulder, CO.
- Ward, P.D., 1987: The Natural History of Nautilus. Allen and Unwin, Boston, MA.
- Ward, P.D. & Greenwald, L., 1982: Chamber refilling in Nautilus. Journal of the Marine Biological Association of the United Kingdom 62, 469–475.