
ABSTRACT
Rare phosphatised fragments within small shelly fossil assemblages from the upper Henson Gletscher Formation (Cambrian, Miaolingian Series, Wuliuan Stage) of North Greenland (Laurentia) are described as Dietericambria hensoniensis n. gen. n. sp. Two pairs of minute cephalic limbs promote comparison with stem-group pentastomids, best known from the late Cambrian (Furongian) Orsten Lagerstätten of Sweden. The North Greenland occurrence is interpreted as the oldest yet described tongue worm and extends the record of pentastomids by about 15 m.y. Dietericambria hensoniensis preserves a unique median axial complex of uncertain function, probably an attachment organ; a mouth has not been identified. Isolated hooks and spicules in the samples from the Henson Gletscher Formation are compared to the grasping hooks and copulatory spicules of the extant pentastomid Raillietiella, although their interpretation is speculative.
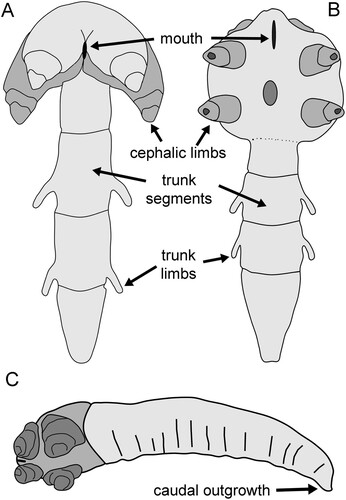
Present-day pentastomids are annulated, vermiform arthropods commonly known as tongue worms (Riley Citation1986; Christoffersen & De Assis Citation2013, Citation2015). Two pairs of cephalic hooks disposed around the mouth facilitate attachment of the parasites to the soft internal tissues of their host, typically the respiratory tracts of tetrapods, but also in some fishes and insects (Christoffersen & De Assis Citation2013, Citation2015). Stem-group pentastomids are particularly well known from the late Cambrian Orsten Lagerstätten of Sweden (Waloszek & Müller Citation1994; Waloszek et al. Citation2006; Castellani et al. Citation2011) but they are also reported from Newfoundland (Waloszek et al. Citation1994) and the Silurian of the UK (Siveter et al. Citation2015). The exceptional, early diagenetic phosphatisation through delicate external encrustation of organisms within the Orsten occurrences (Maas et al. Citation2006) preserves morphologies dissimilar from extant groups, although the diagnostic two pairs of cephalic limbs are well displayed (). Most of the fossil species have few trunk segments and a proportionately large head greater in width than the trunk, although body proportions in Aengapentastomum Waloszek, Repetski & Maas, Citation2006 (C) are more similar to extant forms. Waloszek & Müller (Citation1994) recognised two groups. Members of the “hammer-head” group have greatly enlarged cephalic limbs and include Heymonsicambria Waloszek & Müller, 1994 (A). Boeckelericambria Waloszek & Müller, 1994 is a “round-head” form (B). While some extant pentastomatids may attain a length of about 160 mm, known Orsten specimens rarely exceed a length of 1–2 mm. They have been interpreted as larvae within the meiofauna (Waloszek & Müller Citation1994; Maas et al. Citation2006; Christoffersen & De Assis Citation2015), or even micromorphic adults (Sanders & Lee Citation2010), but their small size may also reflect preservational size limits imposed by the phosphatisation process.
Figure 1. Cambrian stem-group pentastomids from Sweden. A. Heymonsicambria Waloszek & Müller, 1994, a “hammer-head” stem-group pentastomid. B. Boeckelericambria Waloszek & Müller, 1994, a “round-head” stem-group pentastomid. C. Aengapentastomum Waloszek, Repetski & Maas, Citation2006. Schematic sketches redrawn after Waloszek & Müller (Citation1994, fig. 1), Castellani et al. (2011, text-fig. 1) and Waloszek et al (Citation2006, fig. 3).
This paper describes rare vermiform fossils from the middle Cambrian (Miaolingian Series, Wuliuan Stage) Henson Gletscher Formation of North Greenland (). They are interpreted as the oldest described stem-group pentastomids, some 15 m.y. older than records from the late Furongian Orsten of Sweden. The fragmentary remains are phosphatised, although their preservation is much coarser than the exquisitely preserved Orsten material. Interpretation as a stem-group pentastomid reflects general form and the identification of two pairs of minute cephalic limbs in the holotype of the type species, Dietericambria hensoniensis n. gen. n. sp. (), although these are much smaller than the cephalic limbs seen in Orsten species (). The median axial area of the head of Dietericambria hensoniensis also retains a complex of flange-like structures of uncertain function without equivalence in Orsten forms. Detached hooks and spicules from the same Greenland samples are compared to similar sclerotised structures in extant pentastomids, but direct evidence of their association with Dietericambria hensoniensis is lacking.
Figure 2. Geographical and geological background. A. Location of GGU samples 218831 and 271492 in southern Lauge Koch Land with inset of C. B. Greenland showing location (arrow) of J.P. Koch Fjord. C. Løndal region of western Peary Land showing the location of GGU sample 271718. D. Cambrian stratigraphy showing the location of GGU samples in the Henson Gletscher Formation (black dot).
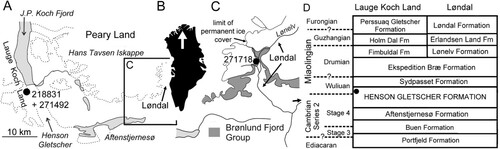
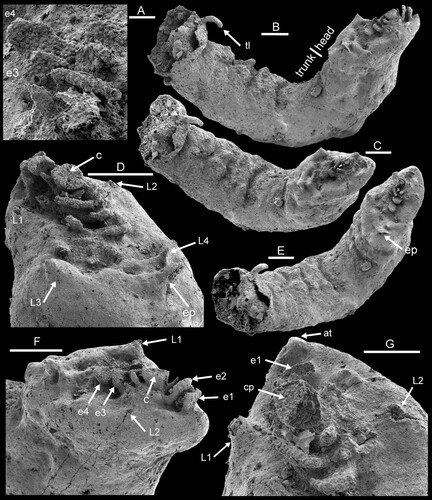
Figure 3. Dietericambria hensoniensis n. gen. n. sp., PMU 39167 from GGU sample 271492, holotype, Henson Gletscher Formation, southern Lauge Koch Land. Cambrian (Miaolingian Series, Wuliuan Stage). A. Detail of C showing ornamentation of median axial complex on ventral surface of head. B,C,E. Oblique lateral and ventro-lateral views of head and trunk. D,F,G. Oblique ventral (D,G) and oblique lateral (F) of head. Abbreviations: at, anterior tubercle; c, central raised disc; cp, papillate surface of central raised disc; e1–e4, flange-like extensions; ep, posterior flange-like extension; L1–L4, cephalic limbs; tl, trunk limb. Scale bars: 20 µm (A), 50 µm (G), 100 µm (B–F).
Background
The Henson Gletscher Formation in the western Peary Land to Lauge Koch Land region () is a constituent formation of a prograding complex of Cambrian–early Ordovician shelf carbonates and siliciclastic sediments, referred to the Brønlund Fjord and Tavsens Iskappe groups (Higgins et al. Citation1991; Ineson & Peel Citation1997; Geyer & Peel Citation2011; Peel et al. Citation2016; Peel & Kouchinsky Citation2022). The highly fossiliferous formation is composed mainly of dark, recessive, limestones, dolostones and mudstones, but a middle member consists of pale fine-grained sandstones.
Fossil assemblages from the Henson Gletscher Formation in southern Lauge Koch Land and Løndal range in age from Cambrian Series 2 (Stage 4) to the Miaolingian Series (Wuliuan Stage; Ptychagnostus gibbus Biozone), but Drumian Stage strata occur to the west in outcrops along the northern coast of North Greenland (Higgins et al. Citation1991; Robison Citation1994; Blaker & Peel Citation1997; Ineson & Peel Citation1997; Geyer & Peel Citation2011). Trilobite assemblages from the Henson Gletscher Formation have a Laurentian aspect but include species, associated with agnostoids, that are important for international correlation with Siberia, the Altai Sayan fold belt and South China (Blaker & Peel Citation1997; Geyer & Peel Citation2011). Additional faunal elements have been described by Clausen & Peel (Citation2012), Peel (Citation2015, Citation2017, Citation2019, Citation2021), Peel et al. (Citation2016) and Peel & Kouchinsky (Citation2022).
Material
Grønlands Geologiske Undersøgelse (GGU) samples 218831 and 271492 were collected at 56.5 m above the base of the Henson Gletscher Formation at its type locality in Lauge Koch Land, where the formation attains a thickness of 62 m, in scours on the top of a 1-m-thick mass flow deposit (82°10′N, 40°24′W; Ineson & Peel Citation1997, fig. 31; Geyer & Peel Citation2011, fig. 3; A). GGU sample 218831 (weight about 200 g) was collected by Peter Frykman on 24th June 1979. GGU sample 271492 (weight about 2 kg) was collected by J.S. Peel on 25th June 1978.
GGU sample 271718 (weight about 5 kg) was collected by J.S. Peel on 15th July 1978 from a thin-bedded, phosphatised, dark dolomitic limestone occurring about 1 m below the top of the formation on the west side of Løndal (82°18′N, 37°03′W; Clausen & Peel Citation2012, fig. 1; C), where the formation has thinned to 47 m.
Specimens were hand picked from residues of limestone dissolved in 10% acetic acid, prior to examination and imaging by scanning electron microscopy. Images were assembled in Adobe Photoshop CS4.
Abbreviations and repositories
GGU prefix indicates a sample made by Grønlands Geologiske Undersøgelse (Geological Survey of Greenland, now a part of the Geological Survey of Denmark and Greenland; GEUS), Copenhagen, Denmark; PMU prefix, a specimen deposited in the palaeontological type collection of the Museum of Evolution, Uppsala University, Sweden.
STEM-GROUP PENTASTOMIDS
Genus Dietericambria new genus
Type species. – Dietericambria hensoniensis n. gen. n. sp. from the Henson Gletscher Formation of North Greenland; Cambrian, Miaolongian Series, Wuliuan Stage.
Diagnosis. – An elongate pentastomid with a complex of steeply inclined flanges in the median axial region of the head. Head oval, of similar width to the trunk, with four minute limbs; trunk with six or more segments and small trunk limbs. Posterior without apparent annulation; tapering to a short caudal outgrowth.
Etymology. – For Dieter Waloszek in recognition of his studies of Swedish Orsten fossils, including stem-group pentastomids.
Discussion. – Dietericambria is readily distinguished from described “hammer-head” and “round-head” Cambrian stem-group pentastomids () by its narrow oval head with minute limbs, greater number of trunk segments, extended posterior section and the complex of steeply inclined flanges in the median axial region of the head (). Aengapentastomum andresi Waloszek, Repetski & Maas, Citation2006 from the lower Ordovician (Tremadoc) of Västergötland, Sweden (but likely reworked from underlying late Cambrian according to Waloszek et al. Citation2006), has a similar elongate annulated form to Dietericambria but with more prominent, robust, cephalic limbs. Additionally, Aengapentastomum andresi lacks the median axial complex of flanges that characterises the head of Dietericambria.
Dietericambria hensoniensis n. gen. n. sp.
Figures 3, 4A–C,E,G,H
Holotype – PMU 39167 from GGU sample 271492, Henson Gletscher Formation, southern Lauge Koch Land, North Greenland. Cambrian, Miaolingian Series, Wuliuan Stage.
Other figured material. – PMU 39168, a section of the trunk from GGU sample 271492, southern Lauge Koch Land; PMU 39169, a posterior fragment from GGU sample 271718, Løndal, Peary Land. All specimens from the Henson Gletscher Formation; Cambrian, Miaolingian Series, Wuliuan Stage. A pair of conjoined hooks from GGU sample 218831 (PMU 36170; D and F) and a sclerite from GGU sample 271492 (PMU 36171; I–K) are discussed below.
Etymology. – From its occurrence in the type section of the Henson Gletscher Formation, southern Lauge Koch Land, adjacent to Henson Gletscher ().
Description. – The holotype is an incomplete specimen about 1.2 mm long, preserving the head and the thorax; the posterior extremity is broken (B,C,E). It is cylindrical in form, with a maximum diameter of about 270 µm, and curved along its length such that the ventral surface is well displayed along the inside of the curve. The head has a length of 350 µm, with convex sides narrowing forward to meet at the pointed anterior end (E, G). It is shallowly convex but the posterior margin is upturned; its maximum width lies near to the posterior margin, which is shallowly convex in plan view. The thorax consists of at least six fused segments or annulations that are delimited on the ventral surface by broad grooves, but obscure on the lateral and dorsal surfaces. The margins of the segments on the ventral surface are raised, with conical tubercles present ventrally (C). A trunk limb extends from the final preserved segment (B, arrow tl) and it is likely that similar limbs or spines arose from the conical tubercles. The ventral surface of the trunk is covered by closely spaced, circular papillae, which pass gradually into smooth lateral areas (B). At its broken posterior margin, the outer surface of the trunk is seen to be preserved as a compact layer overlying coarse, granular and crystalline phosphatised fill of the inner cavity (B,C,E).
The ventral surface of the head preserves a complex array of structures, with smooth peripheral areas surrounding an axial zone ornamented by round papillae 1–2 µm in diameter (A,D; C). The papillate zone grades into the surrounding smooth areas, which display a uniform texture on the ventral surface. In the broken, smooth, conical protruberances, which are interpreted as minute cephalic limbs (D,G and C, arrows L1 and L2), the outer surface is seen to be a thin brittle layer about 1 µm in thickness and overlying an inner granular core. The anterior terminates in a small conical tubercle (G, arrow at). Four small conical cephalic limbs (L1–L4) are recognised but display no preserved traces of segmentation or terminal hooks (D,G and C). The median papillated complex appears to be formed of a series of steeply inclined, flange-like extensions of the ventral surface (F, arrows e1–e4) located mainly anterior (e1 and e2) or posterior (e3 and e4) of a central raised disc (c; D,F,G and C). The rounded edges of these flanges locally appear to be cylinders or tubes (A,D). An additional, partially broken extension (D,E, arrow ep) lies posterior to limbs L3 and L4 but closer to L4.
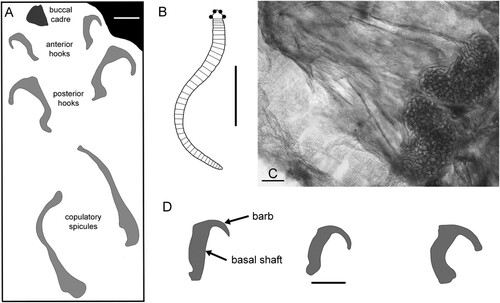
Figure 4. A–C,E,G,H. Dietericambria hensoniensis n. gen. n. sp., Henson Gletscher Formation. Cambrian (Miaolingian Series, Wuliuan Stage). A,B,G. PMU 36169 from GGU sample 271718, Løndal, posterior, with caudal outgrowth (co), trunk limb or spine (tl) and patches of ornamentation beneath phosphatic encrustation. C. PMU 39167 from GGU sample 271492, holotype, head, abbreviations see . E,H. PMU 39168 from GGU sample 271492, fragment of trunk with trunk limbs or spines (tl) and detail of sculpture (H). D,F. Conjoined pair of opposing hooks resembling eupentastomid cephalic hooks. PMU 36170 from GGU 218831. I–K. Spicule with ornamented base, PMU 36171 from GGU sample 271492. Scale bars: 5 µm (H), 10 µm (A,K), 30 µm (B), 50 µm (C,I,J), 100 µm (D–G).
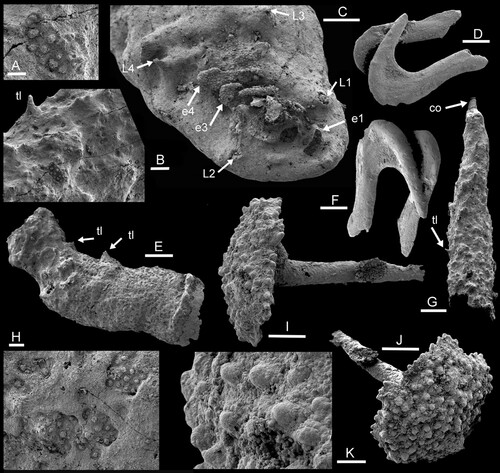
A second specimen consists of a fragment of the trunk with conical bases of the trunk limbs or spines (E, arrows tl). It is broken at the anterior and posterior ends, but a transverse ridge at the anterior end may represent the posterior margin of the head. Most of the surface is covered with diagenetic encrustation but the characteristic papillate ornamentation of small circular tubercles is visible in patches (H). The posterior section is narrower and appears to lack segmentation. In this respect, it is comparable to a third specimen (G) that seems to have a crude honeycomb pattern of ridges, with raised tubercles and short spines but without annulations (B,G, arrow tl). This fragment displays similar papillate ornamentation to other specimens beneath the diagenetic encrustation (A,B) and tapers posteriorly towards a small caudal outgrowth (G, arrow co).
Discussion. – In terms of its overall slender shape, tapering towards the posterior, Dietericambria hensoniensis most closely resembles the late Furongian–?early Ordovician Aengapentastomum andresi amongst described stem-group pentastomids (Andres Citation1989; Waloszek & Müller Citation1994; Waloszek et al. Citation1994, Citation2006; Castellani et al. 2011). However, Dietericambria hensoniensis is distinguished by its much smaller cephalic limbs, which lack segmentation, and the presence of the median axial cephalic complex.
At first glance, some elements of the median axial cephalic complex on the ventral surface of the head appear to be limbs, but closer examination indicates that these elongate rounded surfaces represent the peripheral margins of flanges. In lateral view (F), individual flanges are seen to slope steeply in towards the flat-topped, circular pillar (c). The presence of the same papillate ornamentation of closely spaced, flat-topped tubercles (A) that is seen on the trunk of all three illustrated specimens confirms that the preserved surface of the cephalic flanges is the external surface of the integument and not a mould of its interior. Similar papillation is seen in Heymonsicambria from Västergötland, Sweden, illustrated by Castellani et al. (2011, fig. 9), but on the caudal outgrowth. Equally, the distribution of similar papillae on the trunk of Dietericambria hensoniensis (A,H) indicates that the ornamentation of the median axial cephalic complex is not an impression of a surface to which the head may have been attached.
The position of the mouth has not been located in Dietericambria hensoniensis. The mouth in Boeckelericambria and Aengapentastomum is located forward of the anterior pair of cephalic limbs, while it is located between the inflated basal segments of the anterior limbs in Heymonsicambria (Waloszek & Müller Citation1994; Waloszek et al. Citation1994; Castellani et al. 2011; ). The symmetry of the flanges of the median axial complex might suggest that the circular central pillar (F, arrow c) represented a protruding mouth, but the undamaged part of the flat surface of the structure preserves the external integumental papillation (G, arrow cp).
Trunk limbs in Orsten pentastomids (Waloszek & Müller Citation1994; A,B) are minute structures located on the lateral surfaces in contrast to the ventrally located limbs of Dietericambria hensoniensis (C). In this respect, limb placement in the latter more closely resembles that seen in many tardigrades (Nelson et al. Citation2015; Gross et al. Citation2017). However, the much more robust limbs in the minute stem-group tardigrade from the Miaolingian of Siberia described by Müller et al. (Citation1995) are also disposed laterally (Waloszek et al. Citation2003; Maas et al. Citation2006).
Cambrian pentastomid hooks
Two opposing but diagenetically conjoined hooks from GGU sample 218831 (D,F) invite comparison with the cephalic hooks of eupentastomids, the present-day Pentastomatida (; Christopher & De Assis Citation2015). Hooks are also present as claws at the termination of tardigrade limbs (Nelson et al. Citation2015, fig. 17.8) but generally lack the rounded V-shape with a broader basal shaft and a narrower, tapering barb seen in the Henson Gletscher specimens (for terminology see C). The length of the hooks from the Henson Gletscher Formation, measured along the basal shaft, approaches 500 µm. The hooks are shallowly convex on one side in cross-section (top surface of uppermost hook in D,F), but flattened with an acute margin on the other (upper surface of lower hook). The cross-section thickens as the culmination of the V-shape is approached, with an extension into a thin sheet within the angle of the culmination of the V-shape. The basal termination is broken in both specimens but the tip of the barb is acutely rounded. The hooks are juxtaposed so that the flattened surfaces are in proximity to each other, and likely facing each other in life. As oriented (D), the uppermost hook would lie on the right side of the head, with the lower hook on the left side if the interpretation as cephalic hooks is correct.
Figure 5. Cephalic hooks and copulatory spicules of the extant pentastomid (Eupentastomida) Raillietiella Sambon, Citation1910. A. Sketch of anterior of male Raillietiella frenatus Ali, Riley & Self, Citation1981 showing cephalic hooks and copulatory spicules (drawn from Kelehear et al. Citation2011, fig. 2b). B. Female with cephalic hooks (black dots on head) and multi-annulated trunk (drawn from Riley Citation1986, fig. 2a). C. Copulatory spicules of male Raillietiella orientalis (Hett, Citation1915) (copyright Crystal Kelehear; reproduced from Kelehear et al. Citation2011, fig. 4c). D. Cephalic hooks of Raillietiella frenatus showing size-related transition from pointed to blunt hooks (drawn from Kelehear et al. Citation2011, fig. 3). Scale bars: 100 µm (A,C,D); 10 mm (B).
Cephalic hooks in extant pentastomids are heavily sclerotised and widely used in delimiting species (Riley Citation1986; Christoffersen & De Assis Citation2013, Citation2015). The disposition of cephalic hooks in Raillietiella Sambon, Citation1910, the most diverse genus of Protostomida in terms of number of described species, was illustrated by Kelehear et al. (Citation2011, fig. 2) who noted that the sharpness of the tip of the barb was related to the body size of the animal. Two pairs of cephalic hooks are present in Raillietiella, although males also develop a pair of copulatory spicules (A).
There is no conclusive evidence to indicate that the described, supposed cephalic hooks from the Henson Gletscher Formation belonged to Dietericambria hensoniensis. Indeed, the length of each hook (500 µm) greatly exceeds the width of the head (about 270 µm) in the holotype. However, the ontogenetic size range and stage in ontogenetic development of Dietericambria hensoniensis are not known. The three available fragments of Dietericambria hensoniensis are of similar size, but this may reflect diagenetic bias, postmortal sorting or ecological separation rather than just a similar ontogenetic stage. Maas et al. (Citation2006) considered that a large part of the Orsten assemblages of Sweden had a meiofaunal origin. Other phosphatised small shelly fossils from within the samples from the Henson Gletscher Formation tend to have a similar size range and the GGU samples represent re-deposited sediments in a background of fine argillaceous deposits. Larger specimens of several constituent taxa from the small shelly fossil assemblage are known from unprocessed rock surfaces but are generally not phosphatised.
Waloszek & Müller (Citation1994) and Castellani et al. (2011) considered the most distal part of the segmented cephalic limbs in described Cambrian stem-group pentastomids to be a finger-like claw. Hooks of the type described herein (D,F) have not been described previously in fossil material, although they are present in extant pentastomids ().
Extant males of the pentastomid Raillietiella develop copulatory spicules that are club-shaped rather than V-shaped (A). In some species the base of these spicules may be extended into a platform, the lower surface of which is covered with a prominent raised ornamentation (Riley Citation1986, fig. 5d; Kelehear et al. Citation2011, fig. 4c; C). Rare spicules of similar form are present in GGU sample 271492 (I–K), although their assignment to pentastomids and interpretation as copulatory spicules is speculative.
Mode of life
Present-day eupentastomids generally have a complex developmental cycle associated with their internal parasitic habit, mainly in tetrapods, birds and fishes, but they can occur in humans (Riley Citation1986; Christoffersen & De Assis Citation2013, Citation2015; Tappe & Büttner Citation2009; Klompmaker & Boxhall Citation2015). Insects may be important intermediary hosts (Gałecki & Sokół Citation2019). None of these host groups occur in the Cambrian but it has been suggested that stem-group pentastomids may have been associated with marine conodont-bearing vertebrates at that time (Sanders & Lee Citation2010; Klompmaker & Boxhal Citation2015). Conodont sclerites are often conspicuous in Cambrian samples, including those examined here. Many of the hosts are terrestrial, unlike the marine stem-group species, but eupentastomids occur in both marine and freshwater fishes. Numerous large arthropods and other invertebrate groups that lived in Cambrian seas might have been hosts to pentastomids. In the early Cambrian of North Greenland, specimens of poorly sclerotised lobopodians attain a length of more than 40 cm, while co-occurring arthropods may be half this size (Budd Citation1998; Peel & Ineson Citation2011; Stein et al. Citation2013; Harper et al. Citation2019). However, direct fossil evidence of such associations is lacking. A Silurian pentastomid from the UK was described as a parasite associated with an ostracod by Siveter et al. (Citation2015), although De Baets & Littlewood (Citation2015) considered this to be ectoparasitic rather than endoparasitic. The cephalic limbs and median axial complex of Dietericambria hensoniensis support interpretation as a parasite, but the host is unknown. Neither is it not known if it was attached to the exterior or interior of its host. External attachment may also represent symbiosis, as is the case with tiny worms attached to the exterior of early Cambrian palaeoscolecidans from China described by Cong et al. (Citation2017). However, Zhang et al. (Citation2020) considered encrustations of tubes on the exterior of Cambrian (Stage 4) brachiopods from South China to represent kleptoparasitism. Traces of parasitism or other forms of commensalism may be recorded also in the shells of invertebrates without clear evidence of their perpetrator (Peel Citation2014).
On account of morphological similarities, stem-group pentastomids have been interpreted generally as larvae, although Sanders & Lee (Citation2010) questioned this interpretation. While interpreted as meiofauna (Waloszek & Müller Citation1994; Maas et al. Citation2006; Christoffersen & De Assis Citation2015), their occurrence together with small shelly fossils of similar size in the assemblages from the Henson Gletscher Formation suggests a strong environmental or taphonomic influence on diagenetic phosphatisation. Other species present in the microfossil residues are usually not phosphatised when preserved as macrofossils in hand specimens.
Evolution
Waloszek et al. (Citation2006) established the crown-group Eupentastomida Waloszek, Repetski & Maas, Citation2006 to include present day pentastomids, placing the Cambrian Orsten fossils (Andres Citation1989; Waloszek & Müller Citation1994; Waloszek et al. Citation1994, Citation2006; Castellani et al. 2011) within the pentastomid stem. They concluded that the systematic position of Pentastomida was unresolved, but stressed the fundamental role of morphological data in establishing the ground patterns of evolution. While accepting that many morphological features were arthropodan, they excluded pentastomids from Arthropoda sensu stricta, in the sense of Maas et al. (Citation2004). Sanders & Lee (Citation2010) suggested that the appearance of late Cambrian parasitic pentastomids so soon after that of the first early Cambrian crustaceans contradicted the hypothesis that pentastomids are allied to advanced crustaceans, which may add support for the interpretation of stem-group pentastomids as offshoots of early arthropods (Castellani et alo. 2011). This argument is strengthened by the present extension of the range of stem-group pentastomids by some 15 m.y., back to the early middle Cambrian Wuliuan Stage. However, Sanders & Lee (Citation2010) presented estimated molecular dates for the origin of pentastomids of 490–520 m.y., which encompasses the upper three series of the Cambrian, and proposed a close relationship to branchiurans.
Molecular data presented by Oakley et al. (Citation2013) and others placed present-day pentastomids within the sclerotised arthropods, as eucrustacean euarthropods (Klompmaker & Boxhall Citation2015; Giribet & Edgecombe Citation2019). Wheat & Wahlberg (Citation2013) followed Sanders & Lee (Citation2010) and suggested that Cambrian–Ordovician stem-group pentastomids may represent a stem group of the common ancestor of branchiurans and pentastomids, but this conflicts with the interpretation of Cambrian species as stem-group pentastomids. Christoffersen & De Assis (Citation2013, Citation2015) gave extensive reviews of the history of pentastomid research and recognised a Phylum Pentastomida Huxley, Citation1869 (Pan-Pentastomida of Almeida & Christoffersen Citation1999) that included the Palaeozoic stem-group species together with present-day taxa placed within the crown-group Eupentastomida.
Haug et al. (Citation2021) reviewed the literature based on molecular and morphological data that suggested that eupentastomids are crustaceans. They urged caution in accepting this interpretation due to the absence of morphological features of crustaceans, while accepting that this might result from modification due to the parasitic habit. As regards the stem-group pentastomids, Haug et al. (Citation2021) noted that some showed an astonishing degree of morphological similarity to extant pentastomids, which De Baets et al. (Citation2021) suggested provided unambiguous evidence of parasitism.
Specimens of Dietericambria hensoniensis (, ) from the Henson Gletscher Formation of North Greenland are interpreted as the oldest described stem-group pentastomids. They were collected from the early middle Cambrian (Wuliuan Stage of the Miaolingian Series) and are thus almost 15 m.y. older than stem-group pentastomids described from the late Cambrian (late Furongian Series) by Waloszek & Müller (Citation1994), Waloszek et al. (Citation2006) and Castellani et al. (2011). From an evolutionary perspective, the age difference may explain the minute (incipient?) size of the cephalic limbs of Dietericambria hensoniensis when compared to those of the “hammer-head” and “round-head” groups of stem-group pentastomids described by Waloszek & Müller (Citation1994). Furthermore, the median axial complex of flanges in the head of Dietericambria hensoniensis, which is assumed to be related to the mouth, is not present in the younger forms, where the mouth is usually a simple slit. It likely represents an attachment structure associated with the mouth. Together with the minute cephalic limbs, it may represent an evolutionary stage prior to the development in later stem-group pentastomids (the Orsten pentastomids) of enlarged cephalic limbs with claws or hooks for attachment to the host and a simplified mouth.
Rare hooks (D,F) and spicules (I,J) from the Henson Gletscher Formation closely resemble cephalic hooks and copulatory spicules occurring in the extant eupentastomid Raillietiella. If correctly identified, they represent the oldest known occurrences of such structures, although no direct association with Raillietiella is implied. Neither is there direct evidence of the association of the hooks and spines with Dietericambria hensoniensis, although their size indicates that they would have been associated with much larger individuals than those currently available.
Data archiving statement
This published work and the nomenclatural acts it contains have been registered in Zoobank: zoobank.org:pub:04D7A5AB-6FD4-42D5-B358-09049D13516F
Acknowledgments
Samples were collected during the North Greenland Project (1978–1980) of Grønlands Geologiske Undersøgelse. Joachim T. Haug is thanked for directing me towards pentastomids. Reviews by Carolin Haug and Jean Vannier are gratefully acknowledged.
Disclosure statement
No potential conflict of interest was reported by the author(s).
References
- Ali, J.H., Riley, J. & Self, J.T., 1981: A revision of the taxonomy of the blunt-hooked Raillietiella, pentastomid parasites of African, South-East-Asian and Indonesian lizards, with a description of a new species. Systematic Parasitology 3, 193–207.
- Almeida, W.O. & Christoffersen, M.L., 1999: A cladistic approach to relationships in Pentastomida. Journal of Parasitology 85, 695–704.
- Andres, D., 1989: Phosphatisierte Fossilien aus dem unteren Ordoviz von Südschweden. Berliner Geowissenschaftliche Abhandlungen A 106, 9–19.
- Blaker, M.R. & Peel, J.S., 1997: Lower Cambrian trilobites from North Greenland. Meddelelser om Grønland, Geoscience 35, 1–145.
- Budd, G.E., 1998: Stem-group arthropods from the Lower Cambrian Sirius Passet fauna of North Greenland. In Fortey R.A. & Thomas R.H (eds.): Arthropod Relationships. Systematics Association Special Volume Series 55, 125–138. Chapman & Hall, London.
- Castellani, C., Maas, A., Waloszek, D. & Haug, J.T., 2011: New pentastomatids from the Late Cambrian of Sweden – deeper insight of the ontogeny of fossil tongue worms. Palaeontographica A 293, 95–145.
- Christoffersen, M.L. & De Assis, J.E., 2013: A systematic monograph of the recent Pentastomida, with a compilation of their hosts. Zoologische Mededelingen 87(1), 1–206.
- Christoffersen, M.L. & De Assis, J.E., 2015: Class Eupentastomida Waloszek, Repetski & Mass, 2006. In Vaupel Klein J.C. von, Charmentier-Daures M & Schram F.R (eds.): Treatise on Zoology – Anatomy, Taxonomy, Biology. The Crustacea, vol. 5, 5–75. Brill, Leiden, Boston.
- Clausen, S. & Peel, J.S., 2012: Middle Cambrian echinoderm remains from the Henson Gletcher Formation of North Greenland. GFF 134, 173–200.
- Cong, P., Ma, X., Williams, M., Siveter, D.J., Siveter, D.J., Gabbott, S.E., Zhai, D., Goral, T., Edgecombe, G.D. & Hou, X., 2017: Host-specific infestation in early Cambrian worms. Nature Ecology & Evolution 1, 1465–1469.
- De Baets, K. & Littlewood, D.T.J., 2015: The importance of fossils in understanding the evolution of parasites and their vectors. Advances in Parasitology 90, 1–51.
- De Baets, K., Huntley, J.W., Klompmaker, A.A., Schiffbauer, J.D. & Muscente, A.D., 2021: The fossil record of parasitism: its extent and taphonomic constraints. In De Baets K. & Huntley J.W (eds.): The Evolution and Fossil Record of Parasitism. Topics in Geobiology 50, 1–50. Springer, Cham, Switzerland.
- Gałecki, R. & Sokół, R., 2019: A parasitological evaluation of edible insects and their role in the transmission of parasitic diseases to humans and animals. PloS One 14(7), e0219303. doi:10.1371/journal.pone.0219303.
- Geyer, G. & Peel, J.S., 2011: The Henson Gletscher Formation, North Greenland, and its bearing on the global Cambrian Series 2–Series 3 boundary. Bulletin of Geosciences 86, 465–534.
- Giribet, G. & Edgecombe, G.D., 2019: The phylogeny and evolutionary history of arthropods. Current Biology Review 29, 592–602.
- Gross, V., Minich, I. & Mayer, G., 2017: External morphogenesis of the tardigrade Hypsibius dujardini as revealed by scanning electron microscopy. Journal of Morphology 278, 563–573.
- Harper, D.A.T., Hammarlund, E.U., Topper, T.P., Nielsen, A.T., Rasmussen, J.A., Park, T.S. & Smith, M.P., 2019: The Sirius Passet Lagerstätte of North Greenland: a remote window on the Cambrian explosion. Journal of the Geological Society 176, 1023–1037.
- Haug, J.T., Haug, C. & Nagler, C., 2021: Evolutionary history of crustaceans as parasites. In De Baets K & Huntley J.W (eds.): The Evolution and Fossil Record of Parasitism. Identification and Macroevolution of Parasites. Topics in Geobiology 49, 347–376. Springer, Cham, Switzerland.
- Hett, M.L., 1915: On some new pentastomids from the Zoological Society's Gardens, London. Proceedings of the Zoological Society of London 1915, 115–121.
- Higgins, A.K., Ineson, J.R., Peel, J.S., Surlyk, F. & Sønderholm, M., 1991: Lower Palaeozoic Franklinian Basin of North Greenland. Bulletin Grønlands Geologiske Undersøgelse 160, 71–139.
- Huxley, T.H., 1869: An Introduction to the Classification of Animals, 147 pp. John Churchill & Sons, London.
- Ineson, J.R. & Peel, J.S., 1997: Cambrian shelf stratigraphy of North Greenland. Geology of Greenland Survey Bulletin 173, 1–120.
- Kelehear, C., Spratt, D.M., Dubey, S., Brown, G.P. & Shine, R., 2011: Using combined morphological, allometric and molecular approaches to identify species of the genus Raillietiella (Pentastomida). PLoS ONE 6(9), e24936, 1–10. doi:10.1371/journal.pone.0024936.
- Klompmaker, A. & Boxshall, G., 2015: Fossil crustaceans as parasites and hosts. Advances in Parasitology 90, 233–289.
- Maas, A., Waloszek, D., Chen, J.-Y., Braun, A., Wang, X.-Q. & Huang, D.-Y., 2004: Research on early phylogeny of Arthropoda. Progress in Natural Science 14, 158–166.
- Maas, A., Braun, B., Dong, X., Donoghue, P.C.J., Müller, K.J., Olempska, E., Repetski, J.E., Siveter, D.J., Stein, M. & Waloszek, D., 2006: The ‘Orsten’— more than a Cambrian Konservat-Lagerstätte yielding exceptional preservation. Palaeoworld 15, 266–282.
- Müller, K.J., Walossek, D. & Zakharov, A., 1995: ‘Orsten’ type phosphatised soft-integument preservation and a new record from the middle Cambrian Kuonamka Formation in Siberia. Neues Jahrbuch für Geologie und Paläontologie. Abhandlungen 197, 101–118.
- Nelson, D.R., Guidetti, R. & Rebechi, L., 2015: Phylum Tardigrada. In Thorp F & Rogers D.C (eds.): Thorp and Covich’s Freshwater Invertebrates (fourth edition), 347–380. Academic Press, Cambridge, MA.
- Oakley, T.H., Wolfe, J.M., Lindgren, A.R. & Zaharoff, A.K., 2013: Phylotranscriptomics to bring the understudied into the fold: monophyletic Ostracoda, fossil placement, and pancrustacean phylogeny. Molecular Biology and Evolution 30, 215–233.
- Peel, J.S., 2014: Failed predation, commensalism and parasitism on lower Cambrian linguliformean brachiopods. Alcheringa 39, 149–163.
- Peel, J.S., 2015: Bromalites from the Cambrian (Series 2 and 3) of North Greenland. GFF 137, 181–194.
- Peel, J.S., 2017: A problematic cnidarian (Cambroctoconus; Octocorallia?) from the Cambrian (Series 2–3) of Laurentia. Journal of Paleontology 91, 871–882.
- Peel, J.S., 2019: Sponge spicule assemblages from the Cambrian (Series 2–3) of North Greenland (Laurentia): systematics and biogeography. GFF 141, 133–161.
- Peel, J.S., 2021: In-place operculum demonstrates that the middle Cambrian Protowenella is a hyolith and not a mollusc. Alcheringa 45, 385–394.
- Peel, J.S. & Ineson, J.R., 2011: The Sirius Passet Lagerstätte (Cambrian) of North Greenland. Palaeontographica Canadiana 31, 109–118.
- Peel, J.S. & Kouchinsky, A., 2022: Middle Cambrian (Miaolingian Series, Wuliuan Stage) mollusc and mollusc-like microfossils from North Greenland (Laurentia). Bulletin of the Geological Society of Denmark 70, 69–104.
- Peel, J.S., Streng, M., Geyer, G., Kouchinsky, A. & Skovsted, C.B., 2016: Ovatoryctocara granulata assemblage (Cambrian Series 2–Series 3 boundary) of Løndal, North Greenland. Australasian Palaeontological Memoirs 49, 241–282.
- Riley, J., 1986: The biology of pentastomids. Advances in Parasitology 25, 45–128.
- Robison, R.A., 1994: Agnostoid trilobites from the Henson Gletscher and Kap Stanton formations (middle Cambrian), North Greenland. Bulletin Grønlands Geologiske Undersøgelse 169, 25–77.
- Sambon, L.W., 1910: Untitled text appended to Vaney, M.C., & Sambon, L.W., Preliminary notes on three new species of tongue worms (Linguatulidae), in the collection of the “Museum d'Histoire Naturelle,” Paris. Transactions of the Royal Society of Tropical Medicine and Hygiene 3, 134–154.
- Sanders, K.L. & Lee, M.S.Y., 2010: Arthropod molecular divergence times and the Cambrian origin of pentastomids. Systematics and Biodiversity 8, 63–74.
- Siveter, D.J., Briggs, D.E.G., Siveter, D.J. & Sutton, M.D., 2015: A 425-million-year-old Silurian pentastomid parasitic on ostracods. Current Biology 25, 1632–1637.
- Stein, M., Budd, G.E., Peel, J.S. & Harper, D.A.T., 2013: Arthroaspis n. gen., a common element of the Sirius Passet Lagerstätte (Cambrian, North Greenland), sheds light on trilobite ancestry. BMC Evolutionary Biology 13(99), 2–34. doi:10.1186/1471-2148-13-99
- Tappe, D. & Büttner, D.W., 2009: Diagnosis of human visceral pentastomiasis. PloS Neglected Tropical Diseases 3(2), e320, 1–7. doi:10.1371/journal.pntd.0000320.
- Waloszek, D., 2003: The ‘Orsten’ Window—a three-dimensionally preserved Upper Cambrian meiofauna and its contribution to our understanding of the evolution of Arthropoda. Paleontolological Research 7, 71–88.
- Walossek, D. & Müller, K.J., 1994: Pentastomid parasites from the Lower Palaeozoic of Sweden. Transactions of the Royal Society of Edinburgh, Earth Sciences 85, 1–37.
- Waloszek, D., Repetski, J.E. & Maas, A., 2006: A new Late Cambrian pentastomid and a review of the relationships of this parasitic group. Transactions of the Royal Society of Edinburgh, Earth Sciences 96, 163–176.
- Walossek, D., Repetski, J.E. & Müller, K.J., 1994: An exceptionally preserved parasitic arthropod, Heymonsicambria taylori n. sp. (Arthropoda incertae sedis: Pentastomida), from Cambrian-Ordovician boundary beds of Newfoundland, Canada. Canadian Journal of Earth Science 31, 1664–1671.
- Wheat, C.W. & Wahlberg, N., 2013: Phylogenomic insights into the Cambrian Explosion, the colonization of land and the evolution of flight in Arthropoda. Systematic Biology 62, 93–109.
- Zhang, Z., Strotz, L.C., Topper, T.P., Chen, F., Chen, Y., Liang, Y., Zhang, Z., Skovsted, C.V. & Brock, G.A., 2020: An encrusting kleptoparasite-host interaction from the early Cambrian. Nature Communications 11, 2625. doi:10.1038/s41467-020-16332-3.