Abstract
Climate change is affecting marine environments all over the world but scientists' attention is mainly devoted to tropical areas. In the Mediterranean Sea, species with a cold affinity are decreasing very fast from 0 to 40 m depth, while warm water species increase. From 2000, several populations of the zoanthid Parazoanthus axinellae (Schmidt 1882) have been showing signs of suffering along the Ligurian coast. Here we report a three‐year monitoring, from June 2001 to September 2003, of a population of P. axinellae on the rocky cliff of the Portofino Promontory (Ligurian Sea). During this span of time the population, which covered an area of several square metres with a density of about 1 polyp cm−2, was severely reduced. In the meanwhile an encrusting sponge, Crambe crambe, rapidly colonized the free substrates abandoned by the zoanthid. Warm water and the massive proliferation of a cyanobacterium attributed to the genus Porphyrosiphon are hypothesized to be the main causes of this disease.
Introduction
The composition of the Mediterranean biocoenoses is very particular because of the paleogeographic and paleoecological history of this basin. The Mediterranean Sea is the subtropical remains of the Tethys Sea, which was alternatively recolonized by species both with cold and subtropical affinity (Krijgsman Citation2002). Moreover, being a semi‐enclosed marginal basin of relatively small volume, it shows very rapid response to climate change.
Recent evidence points to large‐scale warming of the Mediterranean leading to wide mass mortalities, involving numerous benthic species (Cerrano et al. Citation2000; Perez et al. Citation2000), and to substitutions of species having cold affinity with warm species (Chevaldonné & Lejeusne Citation2003). The fact that the fauna of the northwestern basin includes several boreal species could partially explain its high sensitivity towards warm conditions.
Among Cnidaria, the first Mediterranean mass mortality episode was recorded in 1986 and involved three species of sea fans in the Ligurian and Provençal Seas (Rivoire Citation1991). In Provence, Corallium rubrum and Paramuricea clavata were struck (Rivoire Citation1991), while, in the Ligurian Sea, the affected species was Eunicella cavolinii (Bavestrello & Boero Citation1986). In this area P. clavata was affected during 1992, when heavy rainfall and land run‐off led to the formation of a cold and fresh seawater mass that injured thousands of colonies along the Portofino Promontory (Bavestrello et al. Citation1994). In the same period, the same species was involved in two other episodes of mortality along the Provençal coasts (Harmelin & Marinopoulos Citation1994) and in southern Italy: in this last case the presence of mucilage was invoked as the cause of the disease (Mistri & Ceccherelli Citation1995). In the summer of 1999, the widest episode ever reported in the Mediterranean occurred in the Ligurian Sea. It affected many sessile species of filter feeders involving Porifera, Cnidaria, Bryozoa, Mollusca, and Ascidiacea (Cerrano et al. Citation2000; Perez et al. Citation2000). Among these species, the most severely affected group was the gorgonians, particularly the purple sea fan P. clavata, with several million of damaged colonies and a male bias sex ratio (Cerrano et al. Citation2005).
Recently, anomalous seawater temperatures were invoked as a cause of bleaching events also in the Mediterranean basin involving the corals Cladocora caespitosa, Balanophyllia europea and Oculina patagonica (Rodolfo Metalpa et al. Citation2000; Kushmaro et al. Citation2001) and the sponge Petrosia ficiformis (Cerrano et al. Citation2001). Among these cases a pathogenic agent (the bacterium Vibrio shiloi) was recorded only in the case of Oculina patagonica (Kushmaro et al. Citation1997).
From the summer of 2000 we observed, in several localities of the Ligurian Sea, some signs of degeneration in the widely diffused populations of the zoanthid Parazoanthus axinellae, a species until now never reported among the organisms that suffered climate warming in the Mediterranean Sea.
P. axinellae is a zoanthid widely distributed in the NW Mediterranean (Herberts Citation1972), in a depth range from 3 to 250 m. Out of the Mediterranean, the species was recorded on the Atlantic coast of Ireland (Ager Citation2003). The species is characterized by the typical epibiontic life style on the sponges of the genus Axinella. On shadow vertical cliff, the polyps directly live on the primary substratum producing extremely dense populations, reaching coverings of about 100%. P. axinellae is a species that, as reported by Garrabou (Citation1999), shows a high dynamic with successive growth and shrinkage, probably due to a continuous search of the optimal microhabitat.
To document the spotted episodes of necrosis, a population of this species was monitored for three years, during the summer season, on the rocky cliff of the Marine Protected Area of Portofino (Ligurian Sea, Italy).
Materials and methods
From 2000 we visited several shallow populations of P. axinellae both along the coast of Portofino Promontory and Gallinara Island (Figure ), noticing signs of disease during the summer season. Particularly, the population of Punta del Faro (Figure ) was monitored when disease occurred. The study site is located on the northern side of the Portofino Promontory and it is a north‐oriented large rock, projecting from the main cliff, that starts at 3 m depth and stops at 7 m depth with a length of about 10 m. On the north‐oriented side of this rock, with a total surface of about 40 m2, 10 areas of about 400 cm2 were photographed by a digital camera, positioning a scale bar in each photo, in the period June–September, from 2001 to 2003. The photographs were analysed by the ImageJ program obtaining, for each image, data about (i) the size of the colonies of P. axinellae (well identifiable by the yellow colour) and the encrusting demosponge Crambe crambe (red–orange), the main competitor for the substratum in the studied biocoenosis; (ii) the total recovery of the two species; (iii) the number of both polyps and sponge specimens, evaluated obtaining values of density. Data are presented as average ±SE.
Figure 1. Map of the studied areas. Black arrows indicate the position of the populations visited in the present work. White arrow indicates the population monitored for three years.
During the period of observations, samples of cyanobacteria, which covered the colonies of P. axinellae by a purple mat, were collected and fixed in 4% formalin directly in situ for the laboratory analysis.
A white soft mat covering numerous polyps was collected in August 2003 to test the eventual occurrence of pathogenic fungi. In laboratory, samples were rinsed in marine sterile water, soaked in ethyl alcohol for 5 min and then rinsed in sterile water for 5 min. Small pieces of each specimen were placed in Petri dishes containing different solid media (MA, malt extract Difco 20 g l−1, agar 20 g l−1 pH 6.5; MEA, malt extract Difco 20 g l−1, peptone 1 g l−1, glucose 20 g l−1, bacto agar 15 g l−1; TS, bacto soytone 10 g l−1, glucose 40 g l−1, sodium acetate 1 g l−1, sodium benzoate 50 mg l−1, bacto agar 15 g l−1; MYCO, mycosel BBl laboratories). The Petri dishes were incubated at 22°C under dark conditions and inspected daily for up to 1 month for the development of colonies on the agar. Fungi were isolated exclusively from diseased colonies and were identified to the genus level by microscopical morphology. Nomenclature of fungi follows that listed in Ainsworth et al. (Citation1973) and in Kohlenmeyer & Volkmann‐Kohlenmeyer (Citation1991).
Sea water temperature were recorded (by dive computers) during 10 dives in each month of August of the three considered years, at 5 m depth close to the P. axinellae population.
Results
From summer 2001, we have examined several populations of Parazoanthus axinellae along the coast of the Portofino Promontory and of the Gallinara Island (Ligurian Sea) (Figure ). All the examined populations showed signs of suffering. We have frequently observed several polyps stretching their body producing a long filament of tissue (Figure ). This process results in the detaching of the polyps, which drop below onto the sub‐horizontal bottom. Evidence of suffering was never recorded for the specimens living below 10 m depth.
Figure 2. Different phases of the disease ofParazoanthus axinellae. a, Dropping polyps. b, Detail of the algal mats. c,d, Algal mat covering some polyps. e, White fungus hyphae developing on diseased polyps. f,g, Dead polyps just before detachment from the cliff.

In the station studied by photographs, a dense population of P. axinellae (about 1 polyp cm−2) covered the rock. Polyp tufts, comprising 1–50 polyps arising from a common encrusting base, composed the population. The community was also composed of the barnacle Balanus perforatus, often utilized by P. axinellae as substratum, and by several species of sponges, mainly the encrusting C. crambe (Figure ) and the massive Ircinia oros and I. spinosula.
Figure 3. Series of photos from the same area showing the decrease of theP. axinellae covering from a, 2001 to b, 2002 and c, 2003. The decrease in pylops is evident. Crambe crambe colonize the deserted supstratum.

During the early summer of the three years of monitoring, the substratum where the Parazoanthus community lived was widely covered by a purple mat, formed by a dense population of a cyanobacterium (Figure ), attributed to the genus Porphyrosiphon following Giaccone et al. (Citation2003), which gradually extended over the colonies of P. axinellae (Figure ). When covered by the algal mat, the polyps where always observed contracted (Figures , ). From the second half of August of each considered year mats of white fungus hyphae developed over the polyps (Figure ), which quickly degenerated (Figure ). Fungi grew profusely on the damaged polyps, while no strains were recorded on healthy colonies. The most common fungus isolated belonged to the saprophytic species Trichoderma viridens but Aspergillus niger, and the genera Penicillum, Cladosporium, Nigrospora, Chaetomium, and Poecilomyces were also commonly present. All these genera are known to include opportunistic pathogens that have been reported to cause disease in plants and in immuno‐compromised humans.
The average (±SD) temperature recorded during August 2001 was 26.1±0.42°C, during August 2002 was 25±0.5°C, and during August 2003 was 26.85±1.2°C.
These diseases occurred each summer, from 2001 to 2003, leading to a decrease of the P. axinellae percentage covering from 35.13±10.6% in October 2001 to 2.67±2.5% in September 2003 (Figure ). This dramatic decrease of the surface occupied by colonies of P. axinellae made available wide portions of the substratum, allowing a progressive colonisation by encrusting sponges, particularly C. crambe, which increased its covering percentage from 1.9±0.1% in October 2001 to 19.73±4.1% in September 2003 (Figures , ).
Figure 4. Trend ofP. axinellae (continued line) and C. crambe (dotted line) during the three years of monitoring. a, Average percentage of covering (±SE). b, Average size of the colonies and sponge specimens (±SE). c, Average number of colonies and sponge specimens (±SE).
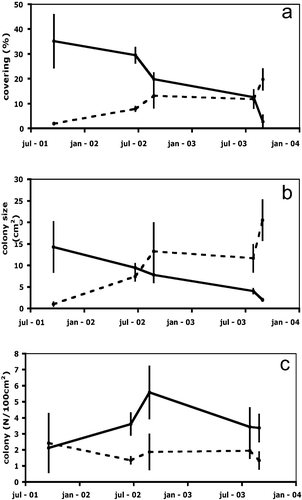
The size of colonies followed a similar trend: while the dead of polyps of P. axinellae resulted in a reduction of the average colony size from 14.24±5.79 cm2 to 1.97±0.27 cm2, the sponges increased their average size from 1.01±0.43 cm2 to 20.47±4.63 cm2 (Figures , ). Due to fragmentation of the zoanthid colonies, their number per 100 cm2 increased from 2.11±0.30 in the first year to 3.36±0.80 in 2003. In the same period the number of specimens of C. crambe per 100 cm2 decreased from 2.42±1.80 in 2001 to 1.33±0.50 in 2003, suggesting the occurrence of fusion among specimens (Figure ).
Discussion
This is the first episode of mass mortality regarding a zoanthid in the Mediterranean Sea. Another episode involving the tropical Palythoa caribaeorum was described on the Brazilian coast (Acosta Citation2001). The comparison between these two phenomena shows strong differences. The disease striking P. caribeaorum is a chronicle phenomenon affecting about 20% of the studied population, producing necrotic zones lower than 5% of the colony surface. The Mediterranean episode is a more acute phenomenon that in three years reduced the coverage of the studied population of more than the 90%.
The disease of P. axinellae also differs from the other mass mortalities that in the last two decades have affected several benthic organisms in the Ligurian Sea. These episodes occurred suddenly: the impressive mass mortality of 1999 involved about the 90% of the sea fans of the Portofino Promontory until 40 m depth in few days (Cerrano et al. Citation2000). The intense but long‐lasting disease of P. axinellae struck only shallow populations, suggesting that for this species the habitat upon 10 m depth is becoming progressively unsuitable. Frequent visual observations, performed by us in the last 20 years indicate that, during this long span of time, the coverage of the substratum by Parazoanthus polyps was constant.
The temperatures recorded during the months of August in the three years of monitoring can be considered higher respect to average temperatures previously known for the same area in the same period (Cerrano et al. Citation2000), which generally never exceed 24°C, suggesting a continuous warming of the Mediterranean sea. On the other hand, this trend is now generally accepted and several authors documented it (Bethoux et al. Citation1990; Turley Citation1999; Gomez & Claustre Citation2003; Laubier Citation2003; Potter & Lozier Citation2004).
Several hypotheses may be invoked to explain the P. axinellae outbreak. It is possible that the primary cause of this disease is the increase in sea water temperature: P. axinellae is a boreal species (Ager Citation2003) and therefore particularly sensitive to global warming. Probably, the fission phenomenon observed in all the shallow populations represents the first reaction of P. axinellae to the water warming. A further increase in temperature determinates a stress so heavy that the polyps become unable to react to the covering by cyanobacteria and by saprophytic organisms. This suffering could be linked to complex biochemical answering to thermal stress as demonstrated by Zocchi et al. (Citation2001) in sponges, where heat stress conditions activate the production of ADP‐rybosil cyclase, which lead to a marked perturbation of the intracellular calcium homeostasis, negatively affecting sponge cell protein synthesis and viability.
Regarding cyanobacteria, massive blooms are generally related to nutrient enrichment, but a temperature increase was also reported as a factor enhancing mat development (Tison et al. Citation1981; Whitton & Potts Citation1982; Snoejis & Prentice Citation1989). In our case, the increase in sea water temperature was concomitant to the development of cyanobacterial mats. Benthic cyanobacteria can produce mats both in freshwater and marine environments. Here, the mats of the filamentous cyanobacterium Porphyrosiphon sp. physically limits the normal functionality of the polyps, avoiding contacts with water flow, similarly to the effects of mucillages on sea fans (Giuliani et al. Citation2005). Moreover, considering that in freshwater and estuarine areas several species of cyanobacteria are also able to produce biotoxins, we cannot exclude that the cyanobacterial mat may release some toxic compound, contributing to the Parazoanthus disease.
As for the other Mediterranean cnidarian diseases, the fungi infesting colonies of P. axinellae represent saprophytic organisms growing on stressed polyps. In fact short‐term exposure to high water temperature can accelerate infection by opportunistic organisms in many stressed sessile invertebrates (Harvell et al. Citation2001). Our observations evidenced a sequential nature of the phenomenon: a first phase characterized by sea water warming and by the occurrence of the cynobacterial mats on polyps of P. axinellae (Figures , ), a second phase with suffering polyps (Figure ) and a third phase with polyps covered by fungi, falling down from the cliff (Figure ).
Regarding the encrusting sponge C. crambe, it is known that it is a slow‐growing species, generally characterized by fast regeneration and long life span (Turon et al. Citation1998). In particular, different behaviours have been recorded between lighted and shaded environments: in this latter C. crambe shows low growth rates, higher survival (Turon et al. Citation1998) and a more efficient production of chemical compounds (Turon et al. Citation1996). For these reasons, in the Mediterranean this species is considered one of the most successful in term of substrate competition (Duran et al. Citation2004). In the case reported here, this species lives in a shaded habitat but revealed fast growth, being the only organism able to cover the area lost by P. axinellae and becoming the new dominant species of the assemblage.
Global warming is producing numerous modifications in organisms' distribution, particularly in a temperate area as the Mediterranean, where species with cold affinity coexist with warm affinity ones. These latter are widening their distribution, limiting the areal of species having cold affinity.
Considering both the high efficiency of benthic suspension feeders in extracting and processing energy from planktonic ecosystems (Gili & Coma Citation1998) and their low growth rate, mortality episodes affecting this functional group may induce long‐term effects on plankton and the structure of benthic communities.
Acknowledgements
This work was supported by the Italian Ministero per l'Università e la Ricerca Scientifica e Tecnologica. The authors thank the staff of the diving centre Continente Blu (Alassio) for field assistance.
References
- Acosta , A. 2001 . Disease in Zoanthids: Dynamics in space and time. . Hydrobiologia , 460 : 113 – 130 .
- Ager OED. 2003. Parazoanthus axinellae . Yellow cluster anemone. Marine Life Information Network: Biology and Sensitivity Key Information Sub‐programme [on‐line]. Plymouth: Marine Biological Association of the United Kingdom. Available online at: http://www.marlin.ac.uk/species/Parazoanthusaxinellae.htm .
- Ainsworth , G. C. , Sparrow , F. K. and Sussman , A. S. 1973 . The fungi, an advanced treatise , New York : Academic Press .
- Bavestrello , G. , Bertone , S. , Cattaneo‐Vietti , R. , Cerrano , C. , Gaino , E. and Zanzi , D. 1994 . Mass mortality of Paramuricea clavata (Anthozoa: Cnidaria) on Portofino Promontory cliffs (Ligurian Sea). . Marine Life , 4 : 15 – 19 .
- Bavestrello , G. and Boero , F. 1986 . Necrosi e rigenerazione in Eunicella cavolinii in Mar Ligure. . Bollettino dei Musei e degli Istituti Biologici dell'Università di Genova , 52 : 295 – 300 .
- Bethoux , J. P. , Gentili , B. , Raunet , J. and Tailliez , D. 1990 . Warming trend in the Western Mediterranean deep water. . Nature , 347 : 660 – 662 .
- Cerrano , C. , Arillo , A. , Azzini , F. , Calcinai , B. , Castellano , L. , Muti , C. , Valisano , L. , Zega , G. and Bavestrello , G. 2005 . Gorgonian population recovery after a mass mortality event. . Aquatic Conservation: Marine & Freshwater Ecosystems , 15 : 147 – 157 .
- Cerrano , C. , Bavestrello , G. , Bianchi , C. N. , Cattaneo‐Vietti , R. , Bava , S. , Moranti , C. , Morri , C. , Picco , P. , Sara , G. , Schiaparelli , S. , Siccardi , A. and Sponga , F. 2000 . A catastrophic mass‐mortality episode of gorgonians and other organisms in the Ligurian Sea (North‐western Mediterranean), summer 1999. . Ecology Letters , 3 : 284 – 293 .
- Cerrano , C. , Magnino , G. , Sarà , A. , Bavestrello , G. and Gaino , E. 2001 . Necrosis in a population of Petrosia ficiformis (Porifera, Demospongiae) in relations with environmental stress. . Italian Journal of Zoology , 68 : 131 – 136 .
- Chevaldonné , P. and Lejeusne , C. 2003 . Regional warming‐indiced species shift in north‐west Mediterranean marine caves. . Ecology Letters , 6 : 371 – 379 .
- Duran , S. , Giribet , G. and Turon , X. 2004 . Phylogeographical history of the sponge Crambe crambe (Porifera, Poeciloslerida): Range expansion and recent invasion of the Macaronesian islands from the mediterranean sea. . Molecular Ecology , 13 : 109 – 122 .
- Garrabou , J. 1999 . Life‐histories traits of Alcyonium acaule and Parazoanthus axinellae (Cnidaria, Anthozoa), with emphasis on growth. . Marine Ecology Progress Series , 178 : 193 – 204 .
- Giaccone , G. , Alongi , G. , Battelli , C. , Catra , M. , Ghirardelli , L. A. , Pezzino , A. and Stefani , S. 2003 . Guida alla determinazione delle alghe del Mediterraneo. Parte I: Alghe azzurre (Cyanophyta o Cyanobacteria) (in ambiente naturale e biodeteriogeni su monumenti lapidei). 92 Pubblicazioni del Dipartimento di Botanica, Università di Catania
- Gili , J. M. and Coma , R. 1998 . Benthic suspension feeders: Their paramount role in littoral marine food webs. . Trends in Ecology and Evolution , 13 : 316 – 321 .
- Giuliani , S. , Virno Lamberti , C. , Sonni , C. and Pellegrini , D. 2005 . Mucilage impact on gorgonians in the Tyrrhenian sea. . Science of Total Environment , 353 : 340 – 349 .
- Gómez , F. and Claustre , H. 2003 . The genus Asterodinium (Dinophyceae) as a possible biological indicator of warming in the western Mediterranean Sea. . Journal of the Marine Biological Association of the United Kingdom , 83 : 173 – 174 .
- Harmelin , J. and Marinopoulos , J. 1994 . Population structure and partial mortality of the gorgonian Paramuricea clavata (Risso) in the North‐Western Mediterranean (France, Port‐Cros Island). . Marine Life , 4 : 5 – 13 .
- Harvell , D. , Kim , K. , Quirolo , C. , Weir , J. and Smith , G. 2001 . Coral bleaching and disease: Contributors to 1998 mass mortality in Briareum asbestinum (Octocorallia, Gorgonacea). . Hydrobiologia , 460 : 97 – 104 .
- Herberts , C. 1972 . Contribution a l'etude biologique de quelques zoantharies tempérès et tropicaux II. Relations entre la reproduction sexuée, la corissance somatique et le bourgeonnement. . Tethys , 4 : 961 – 968 .
- Kohlenmayer , J. and Volkmann‐Kohlenmeyer , B. 1991 . Illustrated key to the filamentous higher marine fungi. . Botanica Marina , 34 : 1 – 61 .
- Krijgsman , W. 2002 . The Mediterranean: Mare Nostrum of earth science. . Earth and Planetary Science Letters , 205 : 1 – 12 .
- Kushmaro , A. , Banin , E. , Looya , Y. , Stackebrandt , E. and Rosenberg , E. 2001 . Vibrio shiloi sp. nov., the causative agent of bleaching of the coral Oculina patagonica. . International Journal of Systematic and Evolutionary Microbiology , 51 : 1383 – 1388 .
- Kushmaro , A. , Rosenberg , E. , Fine , M. and Loya , Y. 1997 . Bleaching of the coral Oculina patagonica by Vibrio AK‐1. . Marine Ecology Progress Series , 147 : 159 – 165 .
- Laubier , L. 2003 . Changement et vulnérabilité des peuplements marins côtiers. . Comptes Rendu Geoscience , 335 : 561 – 568 .
- Mistri , M. and Ceccherelli , V. U. 1995 . Damage and partial mortality in the gorgonian Paramuricea clavata in the Strait of Messina (Tyrrhenian Sea). . Marine Life , 5 : 43 – 49 .
- Perez , T. , Garrabou , J. , Sartoretto , S. , Harmelin , J. G. , Francour , P. and Vacelet , J. 2000 . Mortalité massive d'invertébrés marins: un événement sans précédent en Méditerranée nord‐occidentale. . Comptes Rendu de l'Académie des Sciences Paris, Science Vie , 323 : 853 – 865 .
- Potter , R. A. and Lozier , M. S. 2004 . On the warming and salinification of the Mediterranean outflow waters in the North Atlantic. . Geophysical Research Letters , 31 : L01202 doi: 10.1029/2003GL018161
- Rivoire , G. 1991 . “ Mortalitè du coral et des gorgones en profondeur au large des cotes provençales. ” . In Les Especes marines a protéger le Méditerranée. , Edited by: Bouderesque , C. F , Avon , M and Gravez , V . 53 – 59 . Marseille : GIS Posidonie publications .
- Rodolfo Metalpa , R. , Bianchi , C. N. , Peirano , A. and Morri , C. 2000 . Coral mortality in NW Mediterranean. . Coral Reefs , 19 : 24
- Snoejis , P. J. M. and Prentice , I. C. 1989 . Effects of cooling water discharge on the structure and dynamic of epilithic algal communities in the northern Baltic. . Hydrobiologia , 184 : 99 – 123 .
- Tison , D. L. , Wilde , E. W. , Pope , D. H. and Fliermans , C. B. 1981 . Productivity and species composition of algal mat communities exposed to a fluctuating thermal regime. . Microbial Ecology , 7 : 151 – 165 .
- Turley , C. M. 1999 . The changing Mediterranean Sea—a sensitive ecosystem? . Progress in Oceanography , 4 : 387 – 400 .
- Turon , X. , Becerro , M. A. and Uriz , M. J. 1996 . Seasonal patterns of toxicity in benthic invertebrates: The encrusting sponge Crambe crambe (Poecilosclerida). . Oikos , 75 : 33 – 40 .
- Turon , X. , Tarjuelo , I. and Uriz , M. J. 1998 . Growth dynamics and mortality of the encrusting sponge Crambe crambe (Poecilosclerida) in contrasting habitats: Correlation with population structure and investment in defence. . Functional Ecology , 12 : 631 – 639 .
- Whitton , B. A. and Potts , M. 1982 . “ Marine littoral. ” . In The biology of Cyanobacteria. Botanical Monographs, vol. 19 , Edited by: Carr , N. G and Whitton , B. A . 515 – 542 . Oxford : Blackwell Scientific Publications .
- Zocchi , E. , Carpaneto , A. , Cerrano , C. , Bavestrello , G. , Giovine , M. , Bruzzone , S. , Guida , L. , Franco , L. and Usai , C. 2001 . The temperature‐signaling cascade in sponges involves a heat‐gated cation channel, abscisic acid, and cyclic ADP‐ribose. . Proceedings of the National Academy of Science USA , 98 : 14859 – 14864 .