Abstract
Nest site preference of common dormouse was studied in two different habitat types of central Italy: a mesophilous deciduous forest dominated by Fagus sylvatica, and a thermophilous deciduous oak wood with Quercus cerris and Q. pubescens. In each site, 50 nest boxes were fixed to trees and monitored monthly for two years (May 1998–April 2000). Moreover, in both study sites, 19 habitat variables were measured within a 3‐m radius circular plots centred on each nest box. We hypothesised that specific habitat features may induce common dormice to choose a particular site for nesting. Therefore, we compared habitat variables, recorded at the neighbourhood of either occupied or unoccupied nest boxes, in order to identify the importance of factors in nest site preference. In both beech and oak woodlands, the choice of nest site was affected by understorey vegetation structure rather than species diversity. In particular, the common dormouse prefers sites with high understorey density probably for the presence of route‐ways through branches and visual protection from predators.
Introduction
An accurate description of habitat requirements is a key part of conservation management for specialised species. Habitat alteration due to forest management can influence several life‐history aspects (demography, behaviour, habitat use, reproductive activity, abundance, etc.) of forest‐dwelling vertebrates (Crawford et al. Citation1981; Mahan & Yahner Citation1996; Taulman et al. 1998; Humes et al. Citation1999; Grialou et al. Citation2000; Castleberry et al. Citation2002).
To assess habitat quality, it is important to understand how species perceive their habitat and what features elicit a settling response by the animals. Habitat preference involves a hierarchical discrimination of habitats within landscapes that provide different patterns of resources that affect survival and reproduction (Johnson Citation1980; Douglass et al. Citation1992; Balcom & Yahner Citation1996; Åberg et al. Citation2003). Habitat structure and floristic composition may be used to discriminate among habitats and possible nest sites (Anderson & Shugart Citation1974; Bergin Citation1992; Bence et al. Citation1999; Wagner et al. Citation2000; Kershner et al. Citation2001).
The common dormouse, Muscardinus avellanarius (Linnaeus 1758) has been observed in a variety of habitat types (Bright & Morris Citation1993; Berg & Berg Citation1999; Sarà et al. Citation2001; Capizzi et al. Citation2002), yet appears to have specific habitat requirements. Deciduous woodlands with well‐developed understorey and shrubby habitats are the best habitats for this species (Gaisler et al. Citation1977; Bright & Morris Citation1990; Gurnell et al. Citation1992; Berg & Berg Citation1998; Juškaitis Citation2003). Nevertheless, common dormouse preferences have been studied mainly in deciduous woodlands of northern Europe, and there is scarce information about habitat features providing the optimal habitat for dormice in southern European countries (Sarà et al. Citation2000; Panchetti Citation2005).
The common dormouse builds globular nests that are probably important for predator avoidance, protection from weather conditions and security for offspring. Arboreal nests are supported by understorey branches or constructed in tree hollows. Nest boxes, which represent artificial substitutes of tree hollows, are usually utilised by the species (Catzeflis Citation1984; Morris et al. 1990; Sorace et al. Citation1999; Juškaitis Citation2005). Although some researchers have described the general features of the habitat nearby the arboreal nests (van Laar Citation1979; Hurrell & McIntosh Citation1984; Bright & Morris Citation1991, Citation1992), only few studies have been made to quantify them (Juškaitis Citation1997; Berg & Berg Citation1998).
We hypothesised that some characteristics of the habitat in which common dormice are found induced them to prefer a particular site for nesting. The aim of this study was to identify the habitat features which affected the choice of arboreal nest sites by common dormice in two different habitat types of central Italy. In particular, the present study was designed to verify the association between nest boxes used by common dormice and specific habitat characteristics.
Materials and methods
Study areas
The study was carried out in two different habitat types of Latium Region (central Italy): (1) a mesophilous deciduous wood dominated by beech (Fagus sylvatica) within the Natural Reserve of Lake Vico (42°20′N, 12°11′E, 680 m a.s.l., 40 km from the coastline), and (2) a thermophilous deciduous wood dominated by downy oak (Quercus pubescens) within the Natural Reserve of Castelporziano (41°44′ N, 12°24′ E, sea level, 10 km from the coastline).
The beech wood (BW) was composed by old and well‐spaced beech trees with a dense canopy. Other tree species such as the Italian white maple (Acer obtusatum), turkey oak (Q. cerris) and hornbeam (Carpinus betulus), occurred with few and scattered trees. The shrub layer was scarce and low (about 50 cm height), consisting mainly of butcher's broom (Ruscus aculeatus) with a patchy distribution on the soil surface.
In the deciduous oak wood (OW), downy oaks formed the arboreal layer along with turkey oaks (Quercus cerris), holm oaks (Q. ilex) and occasional trees of Italian oak (Q. frainetto). This woodland was a mosaic of closed wood patches and small clearings, with some stands of evergreen shrubs and trees, mostly represented by heath tree (Erica arborea), strawberry tree (Arbutus unedo), phyllirea (Phyllirea sp.), true myrtle (Myrtus communis), lentisk (Pistacia lentiscus) and evergreen buckthorn (Rhamnus alaternus).
Data collection
Fifty nest boxes were set up in each study site. Nest boxes were placed directly on tree trunks with the entrance hole facing the trunk at 1.5 m above ground level, and spaced approximately at a distance of 20 m from each other, according to a rectangular grid. They had been inspected monthly for two years from May 1998 to April 2000 and always checked at the same time of day, approximately from 9:00 to 11:00, in order to verify the occurrence of dormice or their nests (Panchetti et al. Citation2004).
In order to examine nest‐site preference, from June to September 1999, several habitat variables were measured within 3‐m radius circular plots centred on each nest box (Table ). Variables have been measured in three vegetation strata defined by plant tallness and diameter breast height (DBH): (1) overstorey (DBH⩾7.5 cm), (2) understorey (>2 m tall, DBH<7.5 cm) and (3) shrub layer (<2 m tall) (Dueser & Shugart Citation1978).
Table I. Vegetation variables measured in each 3‐m radius circular plot in both study sites. DBH = diameter breast height; total canopy cover = cumulative percentage of the overstorey and understorey vegetation cover; vegetation units = single stems or trunks, even if some of them belong to the same individual; total density = cumulative density of the overstorey and understorey layers.
In the present study ‘total canopy cover’ was the cumulative percentage of the overstorey and understorey vegetation cover. In both study areas, the same variables were recorded, with few exceptions. In BW total canopy cover was not considered because its values were constant (100%) in proximity of each nest box (Table ). In OW we also calculated the density of the most abundant species and in particular of (1) deciduous oaks (downy oak and turkey oak), (2) holm oak, (3) heath tree and (4) phyllirea (Table ).
Statistical analysis
To assess the occupation rate, a nest box was defined as ‘occupied’ when we found inside (1) one or more dormice, (2) a new nest, or (3) an old nest with evident signs of recent presence of common dormouse (e.g. fresh pellets).
To identify the importance of factors affecting preference of nest sites, we compared variables measured at the immediate neighborhood of occupied nest boxes with those recorded at the immediate neighbourhood of unoccupied nest boxes. First, the non‐parametric chi‐square test was carried out to detect if dormouse‐occupied nest boxes in relation to the plant species. Then, we performed logistic regression with a forward stepwise procedure in order to evaluate the relative importance of habitat variables. Previously, to reduce the number of variables presented to multivariate model, we employed the t‐test with Bonferroni correction: only variables for which significant univariate differences were detected between used and unused nest boxes were retained for logistic regression. Logistic regression employed using separately data of first year, second year and both years, showed similar results with an increased significance level. Therefore, we reported only results obtained using cumulative data. In order to meet the analysis requirements data were opportunely transformed (Tables and ). The statistical analysis was carried out by the SPSS package for Windows (8.0 version) and P‐value for entry and removal of variables was set to 0.05. According to Lewis (Citation2004), logistic regression is the most appropriate and flexible statistical test for analysing factors influencing artificial nest occupation.
Table II. Mean values (±SE) for vegetation variables measured at the neighbourhood of both used and unused best boxes in BW. After Bonferroni correction P‐value was set to 0.0045.
Table III. Mean values (±SE) for vegetation variables measured at nearby used and unused best boxes in OW. After Bonferroni correction P‐value was set to 0.0028.
Results
Beech wood (BW)
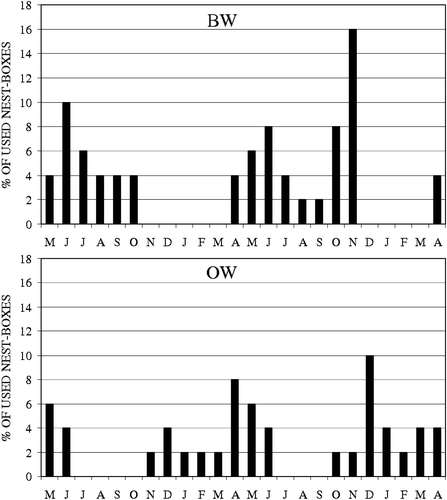
In BW, 48% of nest boxes were used by dormice on the whole: 16% during the first year and 36% in the second one (Figure ). Nest boxes were chosen irrespective of plant species where they were placed (χ2 = 4.64, df = 3, P>0.05).
Figure 1 Percentage of nest boxes occupied each month in BW(above) and in OW (below).
Data analysis with t‐test showed that dormice preferred nest boxes placed in sites with higher vegetation density and cover in the understorey stratum, but lower in the overstorey stratum (Table ).
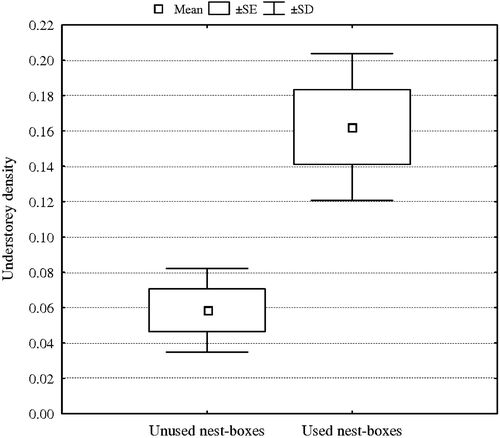
Despite the t‐test analysis, logistic regression analysis indicated the understorey density as the only variable affecting the nest boxes occupation (β±SE = 1.91±0.61, Wald χ2 = 9.98, P = 0.0016; Figure ).
Figure 2 Mean value±SE and SD, of understorey density for used and unused nest boxes in BW site.
Oak wood (OW)
In OW, in both years 14 nest boxes (28%) were used by dormice as nest sites. In particular, 14% of nest boxes were occupied in the first year, and 22% in the second one. The choice of nest box was not related to the plant species where it was placed (χ2 = 5.21, df = 5, P>0.05).
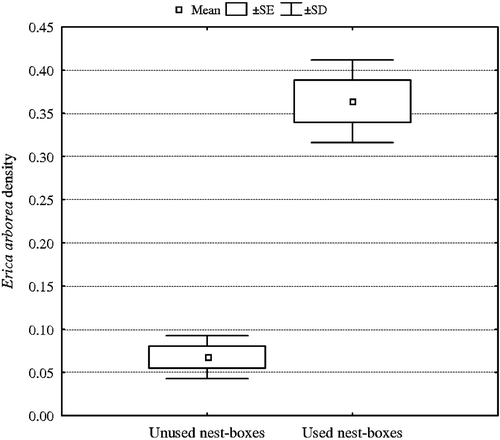
Similarly to BW, analysis with t‐test showed that used nest boxes where located in sites characterised by lower overstorey cover, more developed understorey (i.e. higher cover and density) and higher total vegetation density. Moreover, Q. ilex density and E. arborea density were significantly higher nearby occupied nest boxes, whereas deciduous oak density was significantly lower (Table ). In logistic regression model, only E. arborea density was significantly correlated with the nest box occupation (β±SE = 4.03±1.28, Wald χ2 = 9.86, P = 0.0017; Figure ).
Figure 3 Mean value±SE and SD of Erica arborea density for used and unused nest boxes in OW site.
Discussion
Where tree hollows are available, common dormice select them as nest sites rather than building nests among tangles of vegetation (Bright & Morris Citation1992). For this reason, nest boxes, which mimic tree hollows, are often used by dormice and may cause short‐distance immigration (Morris et al. 1990). Moreover, dormice appear to be relatively long‐lived (5–6 years) in the wild (Juškaitis Citation1999) and so might remember the position of nest boxes (Morris et al. 1990). According to these observations, the increased use of nest boxes, recorded in both study areas from the first year to the second, was expected. Yet our results showed a higher exploitation of nest boxes in beech wood. Considering that common dormice had no previous experience with nest boxes either in beech wood or deciduous mixed oak wood, these differences could depend on population density, but also on forest structure. In fact, nest boxes are more important where the canopy shading is dense, suppressing shrub growth and, consequently, suitable sites for natural woven nests. Moreover, nest boxes are important in woodland where tree hollows are scarce. Both these features were observed jointly in the beech wood of the study area, which turned out to be poor in suitable natural nest sites.
In order to examine nest‐site preference in common dormouse, we compared habitat features at neighbourhood of either used or unused nest boxes. Our results may be affected by the fact that all our nest boxes were located at the lower limit of the range of distance from soil surface known for the natural sites occupied by the species (0.5–16 m height above ground; Bright & Morris Citation1992). However, taking into account that in both study areas nest boxes had the same design, were made with the same material and were tied to the trunk in the same position (height and compass direction), the presence of common dormouse nest into a nest box is a reliable indicator of a suitable nest site for dormice in our experimental setting. In both study areas, our results showed a marked association among occupied nest boxes and specific habitat features. In particular, either in beech or oak wood, nest boxes placed where the overstorey density was higher were avoided by dormice. In contrast, the species preferred sites with a more developed understorey stratum. Even though the understorey foliage mass was not very high in the beech wood, young beech plants were in connection among themselves and with big trees, where the understorey cover reached about 60%. This overlapping among branches probably offers arboreal route‐ways for dormice that, taking into account the reluctance of the species to walk on the ground (Bright & Morris Citation1991, Citation1992; Berg & Berg Citation1998; Bright Citation1998), found a facilitation to move easily in every direction. In abandoned coppice woods, the relation between dormice density and higher understorey growth was already proved by Bright and Morris (Citation1990) in British deciduous woodlands dominated by Quercus robur and Fraxinus excelsior.
Differently from beech wood, oak wood was a heterogeneous habitat consisting of alternated closed tree patches, small clearings and stands of evergreen shrubs or trees. The composition and structure of understorey changed in relation to overstorey. In patches where the arboreal layer was formed by turkey and downy oaks, the understorey was composed mostly by phyllirea, true myrtle, evergreen buckthorn and lentisk. These are evergreen shrubs that generally have a large volume of foliage. In contrast, in patches with low density of deciduous oaks, the understorey was composed mainly by heath tree. This evergreen tree has a vertical growth and forms a close and thick underwood, characterised by null foliage volume from the ground until about 3–4 m height. Leaves are present only in the higher stratum because of canopy shadow, due mainly to overstorey evergreen trees (strawberry tree and holm oak). Our study suggests that, in oak wood, dormice showed a preference in occupying nest boxes placed in areas with high understorey cover mainly consisting of heath trees. These results are in contrast with other studies (Gaisler et al. Citation1977; van Laar Citation1979; Bright & Morris Citation1990; Berg & Berg Citation1998) which showed a preference for areas with high understorey species diversity. A possible explanation of these differences could be the different spatial scale considered. Previous studies were more aimed at studying general habitat preferences in relation to population density; instead, our research was designed to investigate microhabitat preferences within two different deciduous woodlands, in relation to nesting site choice. The correlation between dormouse presence and shrub species diversity observed by previous studies (Gaisler et al. Citation1977; van Laar Citation1979; Bright & Morris Citation1990; Berg & Berg Citation1998) was explained by the requirement of a continuous food supply (Richards et al. Citation1984; Bright & Morris Citation1993; Berg & Berg Citation1998) and not necessary to suitable nest site availability. Telemetry observations clearly showed that the nest sites do not correspond to foraging areas (Panchetti et al., unpublished data), suggesting that the choice of a suitable nest site is led by other habitat characteristic rather than species diversity. Considering that nest position is important for predator avoidance, we can assume that dormice selected areas of high understorey cover to conceal nests and themselves from predator sight, in particular from owls. Nocturnal raptors (Tyto alba and Strix aluco) are habitual predators of common dormouse (Scaravelli & Aloise Citation1994; Juškaitis Citation2004), even though its role in raptors' diet can vary depending on several factors (season, vegetation structure, geographic area and dormouse population density). In fact, predation by owls on common dormouse is not particularly frequent in Sicily where this rodent is rare and mainly localised in the northern ridge and Etna (Sarà & Casamento 1994). Morever, predation by tawny owl was higher in beech woods than in oak woods from southern Italy, probably because of the scarcity of understorey stratum in beech woods (Sarà & Zanca Citation1989).
In our studied oak wood, raptors would have a difficult task in hunting dormice because their nests and route‐ways are protected by heavy dense heath tree scrub; moreover, all potential predators, including marten (Martes martes), would be more easily detected by dormice because of their movements through thick vegetation.
In conclusion, for nests placed at low elevation from the soil surface, the choice of nest site is mainly affected by understorey vegetation structure. As this observation was recurrent in two different habitat types (beech and oak forest), which differ for both vegetation and climatic conditions, it may be probably generalised to every deciduous woodland type.
Acknowledgments
We are grateful to Alessandro De Michelis and Aleandro Tinelli, respectively Director and Research Coordinator of the Natural Reserve of Castelporziano, and to Felice Simmi, Director of the Natural Reserve of Lake Vico, for authorizations and facilities.
References
- Åberg , J. , Swenson , J. and Agelstam , P. 2003 . The habitat requirements of hazel grouse (Bonasia bonasia) in managed boreal forest and applicability of forest stand descriptions as a tool to identify suitable patches. . Forest Ecology and Management , 175 : 437 – 444 .
- Anderson , S. H. and Shugart , H. H. 1974 . Habitat selection of breeding birds in an east Tennessee deciduous forest. . Ecology , 55 : 828 – 837 .
- Balcom , B. J. and Yahener , R. H. 1996 . Microhabitat and landscape characteristics associated with the threatened Allegheny woodrats. . Conservation Biology , 10 : 515 – 525 .
- Bence , S. , Stander , K. and Griffiths , M. 1999 . Nest site selection by the harvest mouse (Mycromis minutus) on arable farmland. . Aspects of Applied Biology , 54 : 197 – 202 .
- Berg , L. and Berg , Å. 1998 . Nest site selection of the dormouse Muscardinus avellanarius in two different landscapes. . Annales Zoologici Fennici , 35 : 115 – 122 .
- Berg , L. and Berg , Å. 1999 . Abundance and survival of hazel dormouse Muscardinus avellanarius in a temporary shrub habitat: A trapping study. . Annales Zoologici Fennici , 36 : 159 – 165 .
- Bergin , T. M. 1992 . Habitat selection by the western kingbird in western Nebraska: A hierarchical analysis. . Condor , 94 : 903 – 911 .
- Bright , P. W. 1998 . Behaviour of specialist species in habitat corridors: Arboreal dormice avoid corridor gaps. . Animal Behaviour , 56 : 1485 – 1490 .
- Bright , P. W. and Morris , P. A. 1990 . Habitat requirements of dormice Muscardinus avellanarius in relation to woodland management in Southwest England. . Biological Conservation , 54 : 307 – 326 .
- Bright , P. W. and Morris , P. A. 1991 . Ranging and nesting behaviour of the dormouse, Muscardinus avellanarius, in diverse low‐growing woodland. . Journal of Zoology, London , 224 : 177 – 190 .
- Bright , P. W. and Morris , P. A. 1992 . Ranging and nesting behaviour of the dormouse, Muscardinus avellanarius, in diverse low‐growing woodland. . Journal of Zoology, London , 226 : 89 – 600 .
- Bright , P. W. and Morris , P. A. 1993 . Foraging behaviour of dormice Muscardinus avellanarius in two contrasting habitats. . Journal of Zoology, London , 230 : 69 – 85 .
- Capizzi , D. , Battistini , M. and Amori , G. 2002 . Analysis of the hazel dormouse, Muscardinus avellanarius, distribution in a Mediterranean fragmented woodland. . Italian Journal of Zoology , 69 : 25 – 31 .
- Casamento , G. and Sarà , M. 1994 . Distribution and ecology of the Gliridae in Sicily: A preliminar account. . Hystrix , 6 : 161 – 168 .
- Castleberry , S. B. , Bohall Wood , P. , Ford , W. M. , Castleberry , N. L. and Mengak , M. T. 2002 . Summer microhabitat selection by foraging allegheny woodrats (Neotoma magister) in managed Forest. . American Midland Naturalist , 147 : 93 – 101 .
- Catzeflis , F. 1984 . Etude d'une population de Muscardins (Muscardinus avellanarius) lors du repos journalier (Gliridae, Rodentia). . Revue Suisse de Zoologie , 91 : 851 – 860 .
- Crawford , H. S. , Hooper , R. G. and Titterington , R. W. 1981 . Songbirds population response to silvicultural practices in central Appalachian hardwoods. . Journal of Wildlife Management , 45 : 680 – 692 .
- Douglass , R. J. , Douglass , K. S. and Rossi , L. 1992 . Ecological distribution of bank voles and wood mice in disturbed habitats: preliminary results. . Acta Theriologica , 37 : 359 – 370 .
- Dueser , R. D. and Shugart , H. H, Jr. 1978 . Microhabitats in a forest floor small mammal fauna. . Ecology , 59 : 89 – 98 .
- Gaisler , J. , Holas , V. and Homolka , M. 1977 . Ecology and reproduction of Gliridae (Mammalia) in Northern Moravia. . Folia Zoologica , 26 : 213 – 228 .
- Grialou , J. A. , West , S. D. and Wilkins , R. N. 2000 . The effects of forest clearcut harvesting and thinning on terrestrial salamanders. . Journal of Wildlife Management , 64 : 105 – 113 .
- Gurnell , J. , Hicks , M. and Whitebread , S. 1992 . “ The effects of coppice management on small mammal populations. ” . In Ecology and management of coppice woodlands. , Edited by: Buckley , G. P . 213 – 232 . London : Chapman and Hall .
- Humes , M. L. , Hayes , J. P. and Collopy , M. W. 1999 . Bat activity in thinned, unthinned, and old‐growth forest in west Oregon. . Journal of Wildlife Management , 63 : 553 – 561 .
- Hurrell , E. and McIntosh , G. 1984 . The Mammal Society Dormouse Survey, January 1975–April 1979. . Mammal Review , 14 : 1 – 18 .
- Johnson , D. H. 1980 . The comparison of usage and availability measurements for evaluating resource preference. . Ecology , 61 : 65 – 71 .
- Juškaitis , R. 1997 . Use of nestboxes by common dormouse (Muscardinus avellanarius L.) in Lithuania. . Natura Croatica , 6 : 177 – 188 .
- Juškaitis , R. 1999 . Life tables for the common dormouse Muscardinus avellanarius in Lithania. . Acta Theriologica , 44 : 465 – 470 .
- Juškaitis , R. 2003 . New data on distribution, habitat and abundance of dormice (Gliridae) in Lithuania. . Acta Zoologica Hungarica , 49 (Suppl. 1) : 55 – 62 .
- Juškaitis , R. 2004 . Local impact of the Tawny owl (Strix aluco) on the common dormice (M. avellanarius) in Lithuania. . Ekològia Bratislava , 23 : 305 – 309 .
- Juškaitis , R. 2005 . The influence of high nestbox density on the common dormouse Muscardinus avellanarius population. . Acta Theriologica , 50 : 43 – 50 .
- Kershner , E. L. , Bollinger , E. K. and Helton , M. N. 2001 . Nest site selection and renesting in blue gray gnatcatcher (Polioptila caerulea). . American Midland Naturalist , 146 : 404 – 413 .
- Lewis , K. P. 2004 . How important is the statistical approach for analysing categorical data? A critique using artificial nests. . Oikos , 104 : 305 – 315 .
- Mahan , C. G. and Yahner , R. H. 1996 . Effects of forest fragmentation on burrow‐site selection by the eastern chipmunk (Tamias striatus). . American Midland Naturalist , 136 : 352 – 357 .
- Morris , P. A. , Bright , P. W. and Woods , D. 1989 . Use of nest boxes by the dormouse Muscardinus avellanarius. . Biological Conservation , 51 : 1 – 13 .
- Panchetti , F. 2005 . Variazioni del ciclo biologico, attività stagionale e distribuzione ecologica del moscardino (Muscardinus avellanarius) nel Lazio [PhD Thesis]. . Dipartimento di Biologia, Università degli Studi “Roma Tre”. 154 p
- Panchetti , F. , Amori , G. , Carpaneto , G. M. and Sorace , A. 2004 . Activity patterns of the common dormouse (Muscardinus avellanarius) in different Mediterranean ecosystems. . Journal of Zoology, London , 262 : 289 – 294 .
- Richards , C. G. , White , A. C. , Hurrell , E. and Price , F. E. F. 1984 . The food of the Common dormouse, Muscardinus avellanarius, in South Devon. . Mammal Review , 14 (1) : 19 – 28 .
- Sarà , M. , Casamento , G. and Spinnato , A. 2001 . Density and breeding of Muscardinus avellanarius L., 1758 in woodlands of Sicily. . Trakya University Journal of Scientific Research, series B , 2 : 85 – 93 .
- Sarà , M. , Spinnato , A. , Casamento , G. and Romano , S. 2000 . Conservazione del Moscardino Muscardinus avellanarius L., 1758 (Mammalia Myoxidae) nel Parco delle Madonie. . Naturalista Siciliano , 24 : 69 – 95 .
- Sarà , M. and Zanca , L. 1989 . Regime alimentare dell'Allocco (Strix aluco) in Sicilia ed Aspromonte. . Avocetta , 13 : 31 – 39 .
- Scaravelli , D. and Aloise , G. 1994 . Predation on dormice in Italy. . Hystrix , 6 : 245 – 255 .
- Sorace , A. , Bellavita , M. and Amori , G. 1999 . Seasonal differences in nest boxes occupation by the Dormouse Muscardinus avellanarius L. (Rodentia, Myoxidae) in two areas of Central Italy. . Ecologia Mediterranea , 25 : 125 – 130 .
- Taulman , J. F. K. , Smith , K. G. and Thill , R. E. 1995 . Demographic and behavioural responses of southern flyng squirrels to experimental logging in Arkansas. . Ecological Applications , 8 : 1144 – 1155 .
- van Laar , V. 1979 . Summer nest sites of the hazel dormouse in North‐Eastern France. . Acta Theriologica , 24 : 517 – 521 .
- Wagner , D. , Feldhamer , G. and Newman , J. 2000 . Microhabitat selection by golden mice (Ochrotomys nuttalli) at arboreal nest sites. . American Midland Naturalist , 144 : 220 – 225 .