Abstract
Mediterranean sabellid material preserved in the private collection of the Zoological Laboratory of Salento University has been examined in order to revise species of Megalomma genus (Annelida: Polychaeta: Sabellinae) present in the Mediterranean Sea. The description of the new species M. messapicum n.sp. from the Southern coast of the Adriatic Sea, together with the re‐description of M. claparedei (Gravier, Citation1908), a probable lessepsian migrant, are given. The distribution of M. vesiculosum (Montagu, Citation1815), previously reported for the Mediterranean area is discussed at the light of re‐examination of specimens preserved at the Zoological Museum of Berlin (ZMB): Branchiomma vesiculosum var. neapolitana Claparède, Citation1868; Branchiomma kollikeri Claparède, Citation1868; and Sabella lanigera Grube, Citation1846. All the examined taxa are proved to be the same species belonging to Megalomma genus, but differing from M. vesiculosum in some features such as the collar shape and the handle length of thoracic uncini. Megalomma lanigera (Grube, Citation1846) was the name maintained for the Mediterranean taxon, and B. kollikeri was confirmed as junior synonym of this species. All the examined M. vesiculosum material coming from Mediterranean soft‐bottoms, and preserved in the Authors' collection revealed to be M. lanigera. Lastly the presence of M. vigilans is also discussed.
Introduction
The genus Megalomma Johansson, Citation1927 was proposed for species characterized by smooth outer surface of radiole with a single subterminal compound eye, previously considered as belonging to Branchiomma Kölliker, Citation1858. Species remaining within Branchiomma have in fact styloids and numerous paired eyes along radiole. Revisions of Megalomma were performed by Perkins (Citation1984) and Knight‐Jones (Citation1997). More recently, Fitzhugh (Citation2003), and Tovar‐Hernández and Salazar‐Vallejo (Citation2006), describing some new species, discussed about the diagnostic features of the genus. Up to now the genus includes 29 species, of which 18 are distributed in tropical areas. Most of the species show a restricted distribution except for M. vesiculosum (Montagu, Citation1815), originally described for the Kingsbridge Estuary (UK) and subsequently reported worldwide. This species is up to now also the more common species within the Mediterranean area. Other species reported for this area are M. neapolitanum (Claparède, Citation1868) and M. vigilans (Claparède, Citation1868).
The present paper deals with the revision of Mediterranean species of Megalomma genus from the examination of material from the authors' collection. Moreover, in order to clarify the actual distribution of Megalomma vesiculosum, the extra Mediterranean material of this species was compared to Mediterranean specimens, both Claparède's and Grube's type material (ZMB), and more recent material identified as M. vesiculosum, or synonimized with it.
Materials and methods
Most of the examined material has been collected along the Italian coasts since 1989 and was deposited in the private Collection of the Laboratory of Zoology of Lecce (PCZL).
The following material from the Zoological Museum of Berlin (ZMB) were examined: Branchiomma vesiculosum var. neapolitana Claparède Citation1868, catalog number 1544; Branchiomma kollikeri Claparède, Citation1868, catalog number 6387; Sabella lanigera Grube, Citation1846 Holotype T‐ZMB 136; and material from Adriatic Sea, Grube, Citation1861, catalog number Q5226.
Holotype of Megalomma messapicum n. sp. was deposited in the Museum Nacional de Ciencias Naturales de Madrid (MNCN).
In all the species, description of the number of thoracic chaetae and uncini is referred to the third chaetiger, while that of the abdomen are referred to the middle portion of the abdomen.
Taxonomic accounts
Megalomma Johansson, Citation1927
Diagnosis
Medium to large‐size species, with numerous radioles. Radiolar skeleton with 4 or more rows of cells. Palmate membrane and radiolar flanges absent. One or more pairs of radioles, each with a single, terminal compound eye along the inner radiole margin near distal end. Dorsal lips triangular, with radiolar appendage indicate by longitudinal ridge or “midrib” along entire length; distinct blood vessel and radiolar sheath tissue extending through the length of the appendages, without a branchial skeleton extension; dorsal pinnular appendages present or absent. Ventral lips and parallel lamellae present. Posterior peristomial ring collar present. Inferior thoracic notochaete broadly hooded. Abdominal neurochaetal fascicles with elongate narrowly hooded chaetae. Thoracic uncini avicular with numerous very small teeth above the main fang, breast well‐developed, and handle of medium length. Companion chaetae with very thin distal ends perpendicular to the shaft. Abdominal uncini similar to thoracic ones, but with shorter handle.
Remarks
The generic diagnosis follows the previous reported by Knight‐Jones (Citation1983, Citation1997) Knight‐Jones and Walker (Citation1985) Knight‐Jones et al. (Citation1991) and Knight‐Jones and Perkins (Citation1998). All these authors considered the presence of dorsal radiolar appendages recognizing a longitudinal ridge or “midrib” extending through the length of the lips. Fitzhugh (Citation2003) emended the diagnosis, pointing out that the radiolar appendages of the dorsal lips in Megalomma do not have an associated branchial skeleton, but only blood vessel and radiolar sheath tissue supporting the lip. This author also stated that dorsal pinnular appendages can also be absent, as occurs in a new species described in Fitzhugh (Citation2003).
Type species Megalomma vesiculosum (Montagu, Citation1815), Figures ,
Amphitrite vesiculosa Montagu, Citation1815; Sabella vesiculosa Milne‐Edwards Citation1849; Branchiomma vesiculosum Saint‐Joseph 1894; Fauvel, Citation1927.
Material examined
One specimen from South Wales, 2 m depth on sandy bottom, 1989.
Description
Complete specimen with 6 thoracic and 125 abdominal chaetigers. Body length 140 mm, crown length 20 mm. Twenty pairs of radioles. Sub terminal compound eyes on most radioles. Dorsalmost radioles bearing large rounded eyes and a very small free tip, (Figure ). Other radioles with smaller eyes and a longer tip (Figure ). Dorsal lips long with broad base (Figure ), radiolar appendage present. Dorsal pinnular appendages present. Ventral lips with parallel lamellae. Dorsal margin of the collar fused to the fecal groove and forming two folded deep pockets (Figure ). Dorsal lappets not covering the junction between branchiae and peristomium, dorsal lappets reaching the same level of the lateral margin of the collar. Collar slightly higher ventrally (Figure ). Ventral shield of the first chaetigers with a deep anterior notch, remaining shields squared, with a pair of small lateral notches (Figure ). Superior thoracic notochaetae elongate narrowly hooded. Inferior thoracic notochaetae narrowly hooded, 15 per fascicle (Figure ). Thoracic uncini numbering 35, leaving a large gap between torus and ventral shield (Figure ). Thoracic uncini avicular with a well‐developed breast and short handle, and with very small teeth over the main fang (Figure ). Companion chaetae with roughly symmetrical tips, as teardrop‐shaped membrane (Figure ). Abdominal neurochaetae in two transversal rows. Anterior chaetigers with elongate, narrowly hooded chaetae in the anterior row and modified, elongate, narrowly hooded chaetae in the posterior row. Abdominal uncini avicular with a manubrium shorter than the thoracic ones, 35 per fascicle. Tube encrusted with sand.
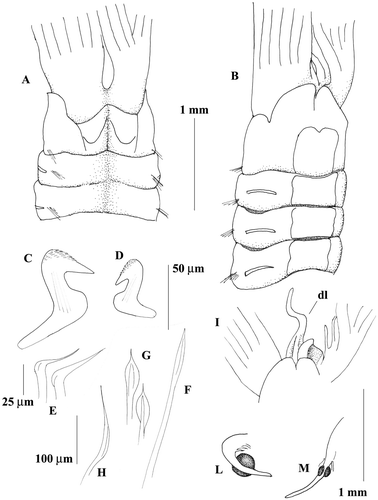
Figure 1 Megalomma vesiculosum: A, anterior end, dorsal view; B, anterior end, ventral view; C, eye from the dorsalmost radiole; D, eyes from other radioles; E, thoracic uncinus; F, companion chaeta; G, inferior thoracic notochaeta.
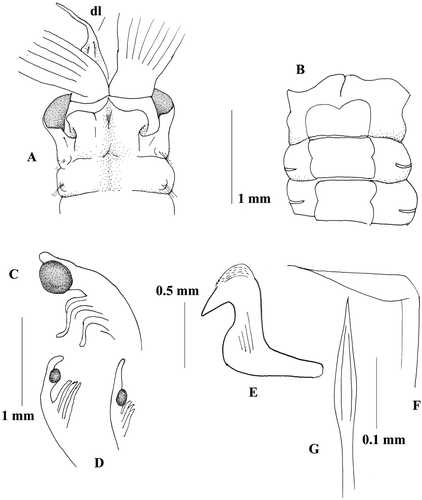
Remarks:
The examined material from South Wales was collected and identified by Dr Knight‐Jones, and kindly given to the University of Lecce collection. It well corresponds to the description of neotype NMWZ 1995.024.
Megalomma lanigera (Grube, Citation1846) Figures , , ,
Sabella lanigera Grube, Citation1846; Branchiomma kollikeri Claparède, Citation1868; Branchiomma vesiculosum Claparède Citation1868; Lo Bianco, Citation1893; ?Soulier, Citation1903; Iroso, Citation1921; Rioja Citation1923; Vatova, Citation1949; Banse, Citation1959; Pronzato & Belloni, Citation1981; Albertelli et al. Citation1983; Gambi et al. 1983–Citation1984; Megalomma vesiculosum Katzmann, Citation1973; Pozar‐Domac, Citation1978; Cantone & Fassari, Citation1982: Farina et al. Citation1985; Bedulli et al. Citation1986; Giangrande & Gambi, Citation1986; Sardà, Citation1991; Megalomma vigilans Flagella et al. Citation2006; Guglielmo et al. Citation2006.
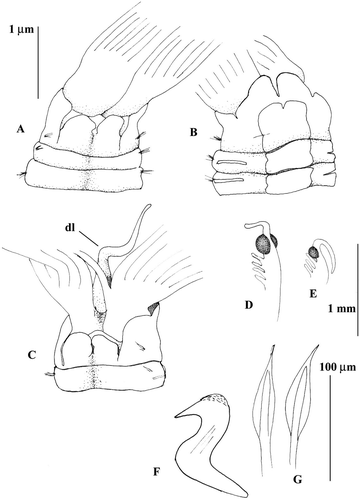
Figure 2 Megalomma lanigera Holotype T‐ZMB 138 (Sabella lanigera): A, anterior end, dorsal view; B, anterior end, ventral view; C, eyes from the dorsalmost radiole; D, eye from another radiole; E, dorsal lip; F, thoracic uncinus; G, companion chaetae; H, inferior thoracic notochaeta.
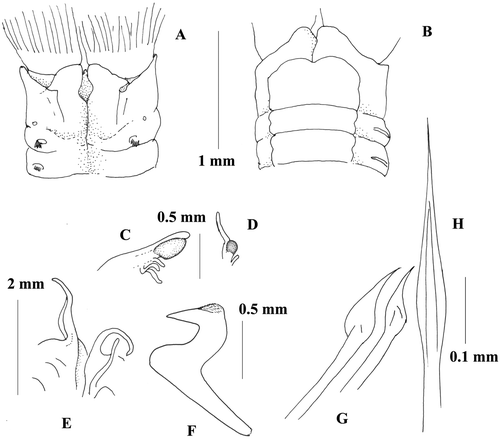
Figure 3 Megalomma lanigera from Venice material: A, anterior end, dorsal view; B, anterior end, ventral view; C, eyes from the dorsalmost radiole; D, eye from another radiole; E, dorsal lip; F, thoracic uncinus.
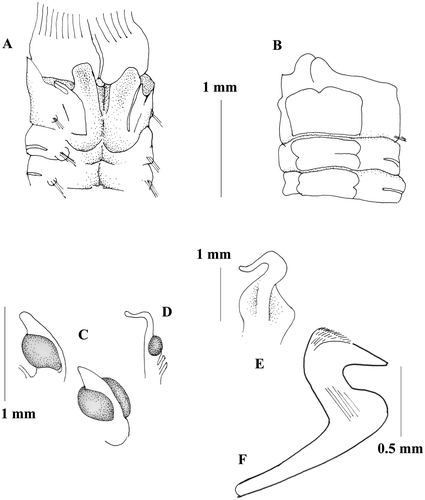
Figure 4 Megalomma lanigera material from the Gulf of Naples: A,B, anterior end, dorsal and ventral views of Claparède's material identified as Branchiomma vesiculosum var. neapolitana ZMB 1544; C,D, anterior end, dorsal and ventral views of Claparède's material identified as Branchiomma kollikeri ZMB 6387; (E,F) anterior end, dorsal and ventral views of recently collected material.
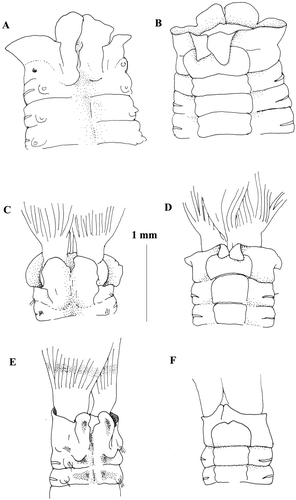
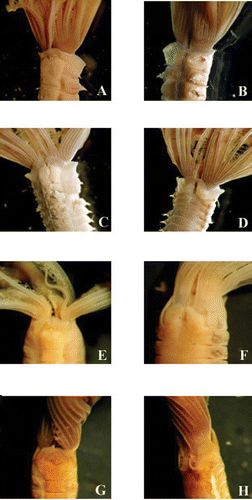
Figure 5 Photographs from differentMegalomma species: A,B, Megalomma vesiculosum; C,D, Megalomma lanigera; E,F, Megalomma claparedei; G,H, Megalomma messapicum n.sp.
Material examined:
Sabella lanigera Grube, Citation1846
Holotype T‐ZMB 136; and Grube, Citation1861, catalog number Q5226.
Material from ZMB: Branchiomma vesiculosum var. neapolitana Claparède Citation1868, catalog number 1544; Branchiomma kollikeri Claparède, Citation1868, catalog number 6387.
Material from the author's collection: one specimen from Salerno Gulf (Tyrrhenian Sea) 1982, 20 m depth on sandy mud bottom; one specimen from Fregene (Tyrrhenian Sea), on sandy bottom, 1989; two specimens from the Venice Lagoon (Adriatic Sea), 5 m depth on muddy sand, 2002; one specimen from Murcia, Spain (Mediterranean Sea) on Posidonia bed; two specimens from the Gulf of Naples, 2004, 11 m depth; one specimen from the Gulf of Naples (Tyrrhenian Sea), 2002, 8 m depth.
Re‐description of holotype
Complete specimen with 7 thoracic and 120 abdominal chaetigers. Body length 85 mm, width 5.8 mm, crown length 18 mm, 28 pairs of radioles in each lobe. Sub terminal compound eyes on most radioles. Dorsalmost radioles bearing large eyes, eyes opaque oval in shape (Figure ) and with a very short free tip. Most of the remaining radioles damaged, but, when present, with smaller and more rounded eyes, and a longer tip (Figure ). Dorsal lips long with broad base (Figure ), radiolar and pinnular appendage present. Ventral lips with parallel lamellae. Vestigial ventral sac. Dorsal margin of the collar fused to the fecal groove and forming two folded deep pockets. Dorsal lappets highly developed covering the junction of the crown, and higher than the lateral margin of the collar, covering the junction as well. Base of the collar dorsally at the pocket level reaching the second chaetiger (Figure ). Collar higher ventrally with highly developed lappets (Figure ). Ventral shield of the first chaetiger squared but with rounded apical margin, following shields rectangular (Figure ). Superior thoracic notochaetae elongate narrowly hooded. Inferior thoracic notochaetae narrowly hooded and with long tip (Figure ). Thoracic uncini leaving a large gap between torus and ventral shield (Figure ). Thoracic uncini avicular with a well‐developed breast and long handle (Figure ). Companion chaetae with roughly symmetrical tips, as teardrop‐shaped membrane (Figure ). Abdominal neurochaetae in two transverse rows. Anterior chaetigers with elongate, narrowly hooded chaetae in the anterior row and modified, elongate, narrowly hooded chaetae in the posterior row. Abdominal uncini avicular with a manubrium shorter than the thoracic ones.
Description of Mediterranean material from the Venice Lagoon
Largest specimen complete with 7 thoracic and 130 abdominal chaetigers. Body length 120 mm, crown length 20 mm. Twenty pairs of radioles. Sub terminal compound eyes on most radioles. Dorsalmost radioles bearing large eyes and a small free tip, eyes oval in shape (Figure ). Other radioles with smaller and more rounded eyes, and a longer tip (Figure ). Dorsal lips long with broad base (Figure ), radiolar appendage present. Dorsal pinnular appendages present. Ventral lips with parallel lamellae. Dorsal margin of the collar fused to the fecal groove and forming two folded deep pockets. Dorsal lappets highly developed covering the junction between the branchiae and the peristomium, and higher than the lateral margin of the collar. Base of the collar dorsally at the pocket level reaching the second chaetiger (Figure ). Collar higher ventrally with highly developed lappets (Figure ). Ventral shield of the first chaetigers squared, following shields rectangular (Figure ). Superior thoracic notochaetae elongate narrowly hooded. Inferior thoracic notochaetae narrowly hooded and with long tip, 45 per fascicle. Thoracic uncini in number of 54, leaving a large gap between torus and ventral shield (Figure ). Thoracic uncini avicular with a well‐developed breast and long handle (Figure ). Companion chaetae with roughly symmetrical tips, as teardrop‐shaped membrane. Abdominal neurochaetae in two transverse rows, with elongate, narrowly hooded chaetae in the anterior row and modified, elongate, narrowly hooded chaetae in the posterior row. Abdominal uncini avicular with a manubrium shorter than the thoracic ones, 50 per fascicle. Tube encrusted with sand.
Remarks
The material from the Venice Lagoon corresponds well to the M. lanigera holotype ( = Sabella lanigera) (Grube, Citation1846) (T‐136). Both are also similar to M. kollikeri (6387) ( = Branchiomma kollikeri) (Claparède, Citation1868) (Figure , D), the only difference being in the slender thoracic segments giving a more squared shape to the ventral shields. Megalomma lanigera and B. kollikeri were previously considered by Hartman (Citation1959) junior synonyms of M. vesiculosum. However, Knight‐Jones (Citation1997) stressed the differences between M. vesiculosum and M. lanigera, suggesting that M. lanigera should be considered a subspecies of M. vesiculosum.
The type locality of M. lanigera is unknown; however, the species was also reported for the Adriatic Sea (Grube Citation1861). Moreover, the South African material of M. vesiculosum of Day (Citation1967), and the material from east Africa is similar to M. lanigera (Knight‐Jones, Citation1997).
The examination of Claparède's material from the Gulf of Naples (catalog number 1544), labeled M. vesiculosum, but probably var. neapolitana (Figure ), does not show any difference in comparison to M. lanigera; by contrast, it is very different to M. vesiculosum.
All the available material coming from soft‐bottom Mediterranean Sea, previously identified as M. vesiculosum, fits well with the above description. Moreover, the examination of recently collected material from the Gulf of Naples and identified as M. vigilans (Flagella et al. Citation2006; Guglielmo et al. Citation2006) proved to be the same species. The specimens are slightly smaller than that collected in the Venice lagoon, but the features agree with that of M. lanigera (Figures ).
The features distinguishing M. vesiculosum from M. lanigera are mainly linked to the collar, which in M. lanigera has higher dorsal lappets covering the base of the branchiae. Moreover, in this species the base of the collar folded pocket reaches the second chaetigers, while those of M. vesiculosum terminate at the first chaetiger (see Figures , , , and ).
The similar size between material of M. vesiculosum from Wales and specimens of M. lanigera from Venice Lagoon allowed us a better comparison also considering other features in separating the two species. Thus, M. lanigera shows: a higher number of thoracic chaetae, thoracic uncini and abdominal uncini; a more developed tip in the inferior thoracic chaetae and a longer handle in the thoracic uncini; a different shape of eyes of the dorsalmost radioles appearing more elongate and a shorter radiolar tip.
Megalomma messapicum n. sp. Figures ,
Type material: Holotype, 1 specimen on MNCN catalog number 16.01/11020, Brindisi (Adriatic Sea) 10 m rocky bottom Gravina leg 1987.
Paratypes, 11 specimens from the same locality PCZL.
Figure 6 Megalomma messapicum n.sp.: A, anterior end, dorsal view; B, anterior end, ventral view; C, thoracic uncinus; D, abdominal uncinus; E, companion chaetae; F, superior thoarcic notochaeta G, inferior thoracic notochaetae; H, abdominal neurochaeta; I, dorsal lip; L, eye from the dorsalmost radiole; M, eye from another radiole; dl = dorsal lip.
Description
The holotype is a complete specimen with 8 thoracic and 45 abdominal chaetigers. Body length 12 mm, width 3 mm. Crown length 5 mm; 9 pairs of radioles in each lobe with three brown bands. Sub terminal compound eyes with a peculiar distribution: dorsalmost radiole bearing large rounded eye and a small free tip (Figure ), second and third and fourth radioles without eyes, fifth, sixth, and seventh with smaller eyes and longer tip (Figure ), eight and ninth without eyes. Dorsal lips long with broad base (Figure ), radiolar appendage present. Dorsal pinnular appendages present. Ventral lips with parallel lamellae. Dorsal margin of the collar fused to the fecal groove low and forming two small pockets with bases not reaching posterior end of first chaetigers (Figure ). Collar slightly higher ventrally with lateral notches and becoming higher ventrally with well‐developed lappets (Figure ). Ventral shield of the first chaetiger squared, two times longer than the following shields, with a little anterior notch (Figure ). Superior thoracic notochaetae elongate narrowly hooded, numbering 5 (Figure ). Inferior thoracic notochaetae very broadly hooded, nearly paleate, numbering 5, and arranged in two rows (Figure ). Thoracic uncini numbering 14, leaving a gap between torus and ventral shield (Figure ); avicular, with a well‐developed breast and short handle (Figure ). Companion chaetae with roughly symmetrical tips, as teardrop‐shaped membrane (Figure ). Abdominal neurochaetae in two transversal rows, with elongate, narrowly hooded chaetae in the anterior row and modified, elongate, narrowly hooded chaetae in the posterior row (Figure ). Abdominal uncini avicular, but differing from the thoracic ones in the shorter main fang, the more numerous teeth surmounting it, and the shorter manubrium (Figure ).
Remarks
Megalomma species are usually inserted into five artificial groups on the bases of whether the mid‐dorsal collar margins are fused to the fecal groove or are unattached, whether the dorso‐lateral collar margin forms pockets or not, and the extent to which the eyes occur on radioles (Knight‐Jones Citation1997). Although artificial, these groups have been useful in species comparison (Nishi Citation1998; Fitzhugh Citation2002, Citation2003). The new described species could belong to the group 1 defined by the fusion of the dorsal margin of the collar to the fecal grove and by the presence of dorso‐lateral pockets. However, the eyes are neither present on most radioles nor in few radioles as occurs in groups 1A and 1B, respectively. The difficulty to place some species within one of the defined groups was already pointed out by Nishi (Citation1998) in describing M. miyukiae. Later, Fitzhugh (Citation2003) stressed the importance to consider the existence of intraspecific variability when describing new species of this genus. Eye distribution not fitting with any existing group is the case of M. messapicum, which, however, shows a very peculiar eye distributional pattern, constantly present in all the examined specimens. This feature mainly distinguishes the new species from all the currently known recognized Megalomma species. Moreover, a collar with very low pockets fused to the fecal groove is not present in any of the species included in both the groups 1A and 1B. Lastly, it must be stressed that the comparison with the species M. vigilans (Claparède Citation1870), described for the Gulf of Naples, was not possible because of the lack of availability type material, and the poor definition of the figures. However, from the original description it is clear that the collar of M. vigilans is similar to that of M. vesiculosum (Claparède, Citation1870), and that the tip of the dorsalmost radioles bearing larger eyes are longer than in the new species.
Etymology
The species was named after the ancient name of the South Apulia where it has been collected.
Megalomma claparedei Gravier, Citation1908 Figures ,
Material examined
One specimen from Brindisi (Adriatic Sea) on rocky bottom, 10 m depth, Gravina leg 1987; four specimens from Brindisi Posidonia bed (matte) 9 m depth, Gravina leg 1987; one specimen from Cerano (Adriatic Sea) on rocky bottom 20 m depth, 2000.
Figure 7 Megalomma claparedei: A, C, anterior end, dorsal view; B, anterior end, ventral view; D, eye from the dorsalmost radiole; E, eye from another radiole; F, thoracic uncinus; G, inferior thoracic notochaetae; dl = dorsal lip.
Description of the Mediterranean material
Largest specimen complete with 8 thoracic and about 60 abdominal chaetigers. Body length 20 mm, width 3 mm. Crown length about 7 mm; 15 pairs of radioles in each lobe. Subterminal compound eyes on most radioles. Dorsalmost radiole bearing large eye and a long free tip (Figure ). Other radioles with smaller eyes and longer tip (Figure ). Dorsal lips long with broad base (Figure ), radiolar appendage present. Ventral lips with parallel lamellae. Dorsal margin of the collar fused to the fecal groove, dorsal lappets ending at the same level of the branchial base, pockets present, but completely separated from the dorsal lappets by a deep cleft (Figure ). Collar becoming higher laterally, showing small notches (Figure ), and ventrally with well‐developed lappets (Figure ). Ventral shield of the first chaetiger quadrangular, longer than the following ones with a little anterior notch; remaining shields rectangular with inconspicuous lateral notches (Figure ). Superior thoracic notochaetae elongate narrowly hooded numbering 4. Inferior thoracic notochaetae very broadly hooded, nearly paleate, numbering 8, arranged in two rows (Figure ). Thoracic uncini numbering of 15, leaving a wide gap between torus and ventral shield (Figure ). Thoracic uncini avicular with a well‐developed breast and short handle (Figure ). Abdominal neurochaetae in two transversal rows, with elongate, narrowly hooded chaetae in the anterior row and modified, elongate, narrowly hooded chaetae in the posterior row. Abdominal uncini avicular with a manubrium shorter than the thoracic ones.
Remarks
Mediterranean material fit well to Gravier's description for material from the Red Sea, especially in the shape of the collar and in the long free tip of the dorsalmost radioles bearing large eyes. In the Red Sea this species was found inside Porites coral (Gravier Citation1908), while in the Mediterranean it was found at different depths on hard bottom substrates and at the base of Posidonia bed where a precoralligenous formation is present.
The species can be considered a lessepsian migrant found, for the moment, only in the South Adriatic Sea.
Concluding remarks
Considering the groups defined by Knight‐Jones (Citation1997) and rearranged by Nishi (Citation1998), all the Mediterranean species revised in the present paper belong to the group 1 A characterized by dorsal collar margin fused to the fecal groove, presence of pockets and occurrence of sub terminal eyes on most radioles. However, the new described species M. messapicum has a peculiar eye distribution not described in any of the existing groups, so stressing the need of update of the present division.
The placement of M. neapolitanum within group 2A remains dubious (differing from 1A only in the dorsal collar margins not fused to the fecal groove) by Knight‐Jones (Citation1997), who also stated the lack of type material. It is highly improbable that this species, originally described as Branchiomma vesiculosum var. neapolitana, might belong to a different group since after Claparède's description (1868), this species distinguishes from M. vesiculosum only for the color pattern. The examined material from the Gulf of Naples (ZMB 1544), identified by Claparède himself, but labeled only as B. vesiculosum, was herein considered to be type material of B. vesiculosum var. neapolitana, because there are no other Claparède's paper or material concerning the Gulf of Naples. This specimen is identical of M. lanigera, so M. neapolitanum becomes a junior synonym of M. lanigera.
Subsequently to Claparède, Lo Bianco (Citation1893) and Iroso (Citation1921) in their studies on the sabellids from the Gulf of Naples, reported B. vesiculosum considering both B. vesiculosum var. neapolitana and B. kollikeri as synonims of that species. Type material is not available for M. vigilans, the last Mediterranean species, so redescription is not provided. After its description, this species was reported by Soulier (Citation1903), and Rioja (Citation1923). However, this species seems to be quite rare, probably because of its particular habitat. From Claparède's (Citation1870) figures it is impossible to define the features of the collar; however, the species has a unique very long free tip in the dorsalmost radiole bearing eye, even longer than that of M. claparedei. Therefore it is likely a valid species.
From the above assumptions several conclusions can be outlined.
Megalomma vesiculosum is not present within the Mediterranean Sea, and it seems to be a species with a Northern hemisphere distribution; therefore, all the findings of this species within tropical and subtropical areas (Day Citation1967; Salazar‐Vallejo Citation1996) must be revised. In addition, all the Mediterranean finding of M. vesiculosum should be assigned to M. lanigera, a species of similar size and colonizing the same habitat: soft bottom sandy mud substrates.
A second conclusion is that Megalomma neapolitanum becomes a junior synonym of M. lanigera.
Regarding the Mediterranean sabellid fauna, two new findings are added both dwelling hard substrates: the new species M. messapicum, and the lessepsian migrant M. claparedei. The two are smaller species than M. lanigera.
Up to now, Megalomma vigilans remains a component of the Mediterranean fauna, but its real status needs to be confirmed.
Acknowledgments
The authors are very grateful to Dr Phyllis Knight‐Jones for having allowed them to look at the type material from different Museums preserved at the University College of Swansea, for discussion and for her precious advice on M. vesiculosum and M. lanigera separation during a work trip at Swansea. This study was financed by MURST (Ministero dell'Università e della Ricerca Scientifica e Tecnologica) and MARBEF (Network of Excellence “Marine Biodiversity and Ecosystem functioning”).
References
- Albertelli , G. , Cattaneo , M. , Della Croce , N. and Drago , N. 1983 . Macrobenthos delle isole di Capraia, Pianosa, Giglio, Montecristo, Giannutri, Elba (Arcipelago Toscano). . Università degli Studi di Genova, Istituto di Scienze Ambientali Marine, Cattedra di Idrobiologia e Pescicoltura, Rapporto Tecnico n. 18
- Banse , K. 1959 . Polychaeten aus Rovinj (Adria). . Zoologischer Anzeiger , 162 : 296 – 313 .
- Bedulli , D. , Bianchi , C. N. , Zurlini , G. and Morri , C. 1986 . “ Caratterizzazione biocenotica e strutturale del macrobenthos delle coste pugliesi. ” . In Indagine ambientale del sistema marino costiero della Regione Puglia , Edited by: Viel , M and Zurlini , G . 227 – 255 . Santa Teresa, La Spezia : ENEA – CREA .
- Cantone , G. and Fassari , G. 1982 . Policheti del Golfo di Catania. . Animalia , 9 : 227 – 237 .
- Claparède , E. 1868 . Les Annélides Chétopodes du Golfe de Naples. . Memoires de la Societe de Physique et d'Histoire Naturelle de Geneve , 19 : 313 – 584 .
- Claparède , E. 1870 . Les Annélides Chétopodes du Golfe de Naples. Seconde partie. . Memoires de la Societe de Physique et d'Histoire Naturelle de Geneve , 20 : 1 – 225 .
- Day , J. H. 1967 . A monograph on the Polychaeta of Southern Africa. Part 2. Sedentaria , London : British Museum (Natural History) .
- Farina , R. , Castelli , A. and Lardicci , C. 1985 . Distribuzione dei policheti sui fondi mobili infralitorali della costa meridionale dell'isola d'Elba (Arcipelago Toscano). . Atti della Società Toscana di Scienze Naturali Modena , 6 : 41 – 46 .
- Fauvel , P. 1927 . Polychètes Sedentaires. . Faune de France , 16 : 1 – 44 .
- Fitzhugh , K. 2002 . Fan worm polychaetes (Sabellidae: Sabellinae) collected during the Thai–Danish Bioshelf Project. . Phuket Marine Biological Center Special Publication , 24 : 353 – 424 .
- Fitzhugh , K. 2003 . A new species of Megalomma Johansson, 1927 (Polychaeta: Sabellidae: Sabellinae) from Taiwan, with comments on sabellid dorsal lip classification. . Zoological Studies , 42 : 106 – 134 .
- Flagella , S. , Guglielmo , R. and Gambi , M. C. 2006 . “ Comunità bentoniche dei fondi sabbiosi in cinque stazioni selezionate lungo le coste della Campania. ” . In Gestione e tutela dell'ambiente marino costiero in Campania , Edited by: Capobianco , L , Del Gaizo , S , Saggiomo , V and Zucaro , E . Napoli : ARPAC .
- Gambi , M. C. , Giangrande , A. and Fresi , E. 1983–1984 . Policheti di fondo mobile del Golfo di Salerno: ipotesi di un modello di distribuzione generale. . Nova Thalassia , 6 (Suppl) : 575 – 583 .
- Giangrande , A. and Gambi , M. C. 1986 . Polychétes d'une pelouse a Cymodocea nodosa (Ucria) Ashers du Golf de Salerno (Mer Thyrrhenienne). . Vie et Milieu , 36 : 185 – 190 .
- Gravier , C. 1908 . Contribution à l'étude des Annélides polychètes de la Mer Rouge. . Nouvelles Archieves du Museum d'Histoire Naturelle de Paris , 10 : 67 – 168 .
- Grube , E. 1846 . Beschreibungen neuer oder weniger bekannter Anneliden. Erster Beitrag. . Archive für Naturgeschichte , 12 : 45 – 59 .
- Grube , E. 1861 . Ein ausflug nach Triest und dem Quarnero, Nicolaische Venabsbuchhundlung. Arch. Naturg. Berlin . 26 : 175
- Gugliemo , R. , Flagella , S. and Gambi , M. C. 2006 . “ Dinamica della comunità a policheti dei fondi sabbiosi in alcune stazioni selezionate del Golfo di Napoli. ” . In Gestione e tutela dell'ambiente marino costiero in Campania , Edited by: Capobianco , L , Del Gaizo , S , Saggiomo , V and Zucaro , E . Napoli : ARPAC .
- Hartman , O. 1959 . Catalogue of the polichetous annelids of the world. . Allan Hancock Foundation Occasional Paper , 23 : 1 – 628 .
- Iroso , I. 1921 . Revisione dei serpulidi e sabellidi del Golfo di Napoli. . Pubblicazioni della Stazione Zoologica di Napoli , 3 : 68 – 85 .
- Johansson , K. E. 1927 . Beiträge zur Kenntnis der Polychaeten‐familien Hermellidae, Sabellidae und Serpulidae. . Zoologiska Bidrag från Uppsala , 11 : 1 – 184 .
- Katzmann , W. 1973 . Contributo alla conoscenza del mare Adriatico (medio Adriatico, fondi mobili tra 10 e 230 m di profondità). . Quaderni del Laboratorio di Tecnologie della Pesca , 1 : 143 – 155 .
- Knight‐Jones , P. 1983 . Contribution to the taxonomy of Sabellidae (Polychaeta). . Zoological Journal of the Linnean Society , 79 : 245 – 295 .
- Knight‐Jones , P. 1997 . Two new species of Megalomma (Sabellidae) from Sinai and New Zealand with redescriptions of some types and a new genus. . Bulletin of Marine Science , 60 : 313 – 323 .
- Knight‐Jones , P. , Knight‐Jones , W. and Ergen , Z. 1991 . Sabelliform polychaetes, mostly from Turkey's Aegean coast. . Journal of Natural History , 25 : 837 – 858 .
- Knight‐Jones , P. and Perkins , T. H. 1998 . A revision of Sabella, Bispira, and Stylomma (Polychaeta, Sabellidae). . Zoological Journal of the Linnean Society , 123 : 385 – 467 .
- Knight‐Jones , P. and Walker , A. J. M. 1985 . Two new species of Demonax (Sabellidae: Polychaeta) from Liverpool Bay. . Journal of Natural History , 19 : 605 – 612 .
- Kölliker , H. 1858 . Über kopfkiemer mit Augen an den Kiemen (Branchiomma dalyelli). . Zeitschrift der wissenschaftliche Zoologie, Leipzig , 9 : 356 – 541 .
- Lo Bianco , S. 1893 . Anellidi tubicoli del Golfo di Napoli. . Atti Accademia di Scienze Fisiche e Matematiche Napoli , 5 : 65 – 81 .
- Milne‐Edwards , H. 1849 . “ Les Annélides avec un Atlas. ” . In Le règne animale, distribue d'après son organisation, pour servir de base a l'histoire naturelle dea animaux, par Georges Cuvier. Par un réunion de disciples de Cuvier , Paris : Fortin, Masson et Cie .
- Montagu , G. 1815 . Descriptions of several new or rare animals principally marine, found on the south coast of Devonshire. . Transactions of the Linnean Society of London , 11 : 18 – 21 .
- Nishi , E. 1998 . A new species of Megalomma (Annelida: Polychaeta: Sabellidae) from Phuket, Thailand. . Pacific Science , 52 : 53 – 60 .
- Perkins , T. H. 1984 . Revision of Demonax Kinberg, Hypsicomus Grube and Notaulax Tauber, with a review of Megalomma Johansson from Florida (Polychaeta: Sabellidae). . Proceedings of the Biological Society of Washington , 97 : 285 – 368 .
- Pozar‐Domac , A. 1978 . Katalog Monogocetinasa (Polychaeta) Jandrana. 1 Sjevererni Isrednij Jadran. . Acta Adriatica , 19 : 1 – 59 .
- Pronzato , R. and Belloni , S. 1981 . Insediamenti zoobentonici su rizomi di Posidonia a diversa profondità. . Bollettino del Museo dell'Istituto di Biologia dell'Università di Genova 48 , 49 : 47 – 54 .
- Rioja , E. 1923 . Estudio sistemático de las especies ibéricas del suborden Sabelliformia. . Trabajos del Museo Nacional de Ciencias Naturales de Madrid Serie Zoología , 48 : 1 – 144 .
- Saint‐Joseph , A. de. 1894 . Les Annélides polychètes des côtes de Dinard. Troisième partie. . Annales des Sciences Naturelles, Zoologie, Série 7 , 17 : 1 – 395 .
- Salazar‐Vallejo , S. I. 1996 . Lista de especies y bibliografia de poliquetos (Polychaeta) del Gran Caribe. . Annales del Instituto de Biología, Universidad Nacional Autónoma de México, Serie Zoología , 67 : 11 – 50 .
- Sardà , R. 1991 . Polychaete commuities related to plant covering in the Mediolittoral and Infralittoral zones of the Balearic Islands (Western, Mediterranean). . PSZNI: Marine Ecology , 12 : 341 – 360 .
- Soulier , A. 1903 . Revision des Annélides de la région de Cette (2e fascicule). . Académie des Sciences et Lettres de Montpellier 2 , 3 : 193 – 278 .
- Tovar‐Hernández , M. A. and Salazar‐Vallejo , S. I. 2006 . Sabellids (Polychaeta: Sabellidae) from the Grand Caribbean. . Zoological Studies , 45 : 24 – 66 .
- Vatova , A. 1949 . La fauna bentonica dell'alto e medio Adriatico. . Nova Thalassia , 1 : 1 – 110 .