Abstract
Two new species found in high mountain waters of Italy representing the family Enchytraeidae are described. Cernosvitoviella longiducta n. sp. is clearly differentiated from all species in this genus by its exceptionally long sperm duct. Cognettia valeriae n. sp. belongs to a group of species having more than three pairs of pharyngeal glands and male pores situated in segment X, but differs from other species in this group by the shape of spermathecae and the well‐developed seminal vesicles.
Keywords:
Introduction
The majority of species representing the Enchytraeidae family belong to terrestrial fauna, but in high mountain water bodies, both running and stagnant ones, and among oligochaetes, this family usually represents the most abundant and diversified group (Dumnicka Citation1976; Kownacki et al. Citation2000; Malard et al. Citation2001; Lencioni et al. Citation2004; Dumnicka & Boggero Citation2007).
Typically aquatic enchytraeids species are reported only in some genera. The genus Cernosvitoviella is the most closely associated with freshwater habitats while the genus Cognettia is semi‐aquatic (Nielsen & Christensen Citation1959; Rota Citation1995), although terrestrial species are also known (Bauer Citation1993). Besides the high mountain areas, many species from these genera are found in cold or humid climates in northern parts of Europe and Asia (Healy Citation1975; Piper et al. Citation1982; Rota et al. Citation1998; Christensen & Dózsa‐Farkas Citation1999), but due to their high adaptability some species can be found in a range of environments and latitudes (e.g. Cernosvitoviella atrata (Bretscher, 1903) or Cognettia sphagnetorum (Vejdovsky, 1878) (Graefe & Schmeltz Citation1999)).
Our knowledge of the oligochaete fauna of mountain streams and lakes is still limited, which is why new species are still being described from this environment (Martinez‐Ansemil & Collado Citation1996; Dumnicka Citation2004).
The present material was collected in northeastern Italy, mainly within the HIGHEST (Health and Integrity of Glacial Headwater EcoSystems in Trentino) program undertaken in the Stelvio National Park, where the invertebrate fauna of some streams, including Noce Bianco (NB) amongst others, was studied. Additional material was sampled during realization of another project “Caratterizzazione biologica dei torrenti e laghi di Val de la Mare” 2000–2001 (Biological characterization of streams and lakes in the de la Mare Valley) (Lencioni & Maiolini Citation2002).
Materials and methods
Information concerning the water bodies of the studied region as well as methods of sampling was published by Boscaini et al. (Citation2004) and Lencioni et al. (Citation2004). In the Noce Bianco stream on each sampling station (a stream bed 15 m in length), kick samples were taken monthly from June to October 2001 with a pond net (100 µm mesh size). Samples from Lago Lungo were also collected using a pond net. All samples were fixed and preserved in 75% ethanol, segregated in a laboratory using a stereoscopic microscope; whole specimens were mounted in Canada balsam.
Taxonomic accounts
Description of Cernosvitoviella longiducta n. sp.
Type series:
Holotype: a whole mounted specimen, kept in the collection of the Institute of Nature Conservation, Polish Academy of Sciences, Krakow, Poland.
Type locality: The Noce Bianco stream (site NB1bis), Rhaetian Alps, Trentino, Italy, 46°24′N, 10°40′E, 2705 m a.s.l., collected on 15 October 2002, leg. V. Lencioni and B. Maiolini.
Four paratypes were appointed: two whole mounted specimens from the same locality and date as holotype, kept in the collection of the Institute of Nature Conservation, Polish Academy of Sciences, Krakow, Poland. One whole mounted specimen from the Noce Bianco stream (siteNB2bis, drift, 2535 m a.s.l., 10 October 2001, leg. V. Lencioni and B. Maiolini) and one whole mounted specimen from the littoral zone of Lago Lungo (2550 m a.s.l., 1 August 2000, leg. V. Lencioni and B. Maiolini).
Etymology
The name describes (in Latin) the most characteristic feature of this species – namely, the very long sperm duct.
Characteristics
Small species: total length of preserved specimens 2.5–3.5 mm, width in the ante‐clitellar part of the body: 110–125 µm. Number of segments 24–29. Prostomium greater in length than width, with a few conspicuous epidermal gland cells. Setae sigmoid, sharply pointed, with the nodulus situated at the distal one third. Length of dorso‐lateral setae 26–30 µm in anterior bundles, 35–38 µm in posterior ones. The same number of setae (4–6) are present in dorso‐lateral bundles along the whole body. In preclitellar segments 5–6 (7) setae occur in a ventral bundle (length 33–39 µm) while in postclitellar ones, the number of setae per bundle is a little smaller (4–5) although they are the longest (37–43 µm).
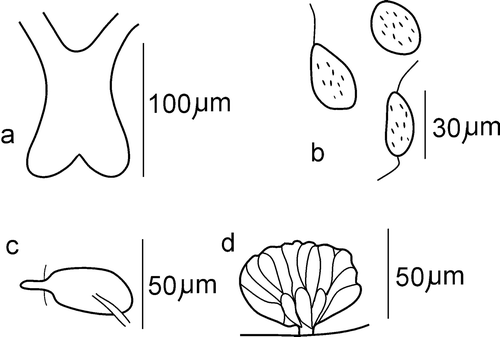
Brain slightly longer than wide (length 82–87 µm), deeply cleft posteriorly (Figure ). Two pairs of pharyngeal glands present: primary glands narrow, united dorsally, situated at 4/5 and 5/6, secondary glands free, spherical, big, situated in V and VI. Coelomocytes of two types: oval, about 19–21 µm long and 10–12 µm wide, and spherical, with a diameter of 15–18 µm; both types sparsely granulated (in fixed specimens) (Figure ), usually not numerous. Chloragogen cells present from V, forming a dense layer from VI. Unpaired nephridia present in 6/7 and 7/8, sometimes also in 8/9.
Figure 1 Cernosvitoviella longiducta n. sp. a, brain; b, coelomocytes; c, sperm funnel.
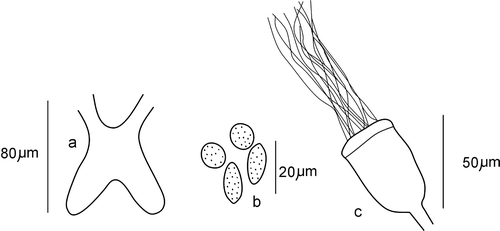
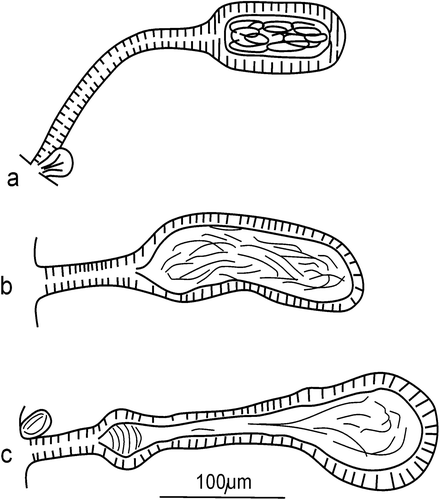
Clitellum on XII–½ XIII, made up of irregularly arranged gland cells. Seminal vesicles in form of a paired compact mass occupying a 1/3–1/2 of XI. Sperm funnels small (length 46–51 µm), slightly longer than wide, cylindrical or faintly barrel‐shaped with distinct but short collars covered in very long spermatozoa (Figure ). Sperm duct very long, 12–15 times longer than the sperm funnel, narrow (mean width 10–11 µm), irregularly coiled in XII (Figure ), with smooth, thin walls. Just behind the sperm funnel its width is a little higher (about 14 µm) with a few small round protrusions (may be glands) visible on the surface (Figure ).
Figure 2 Cernosvitoviella longiducta n. sp. Optical sections of coiled sperm duct (marked by arrows) and a “penial bulb” (pb).
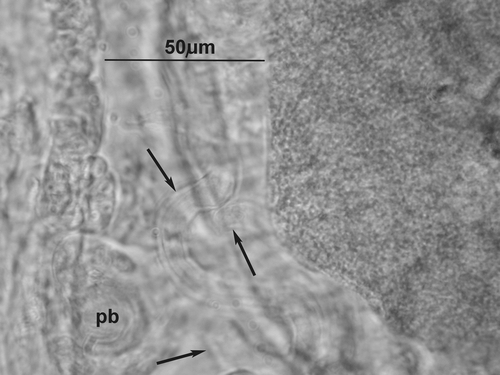
Figure 3 Cernosvitoviella longiducta n. sp. Barrel‐shaped sperm funnel (sf) and beginning of sperm duct with roundnessess (marked by arrows).

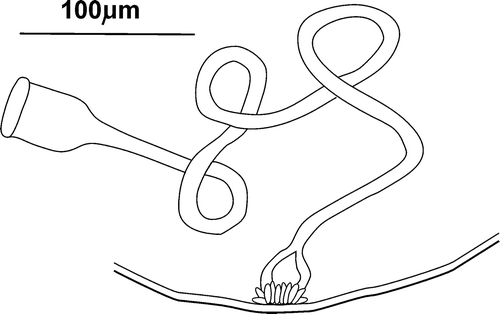
A so‐called “penial bulb”, almost spherical (diameter 40–46 µm), consists of thickened walls in the final part of the sperm duct surrounded by numerous, small glands (Figure ).
Figure 4 Cernosvitoviella longiducta n. sp. male duct with a penial bulb.
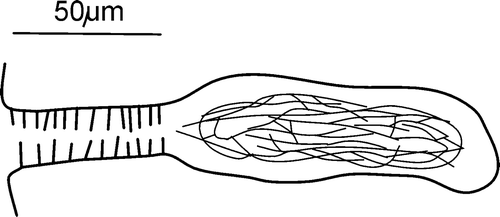
Spermathecae confined to V. Ampullae oval, about 60–70 µm long and 22–26 µm in width with sperm irregularly arranged; ectal ducts stout (diameter 15–18 µm), without glands (Figure ). The length of the ampulla and duct is almost the same or the ampulla is slightly longer.
Figure 5 Cernosvitoviella longiducta n. sp. spermatheca.
Taxonomical remarks
Cernosvitoviella longiducta n. sp. belongs to a group of species having sperm ducts more than five times longer than the length of the sperm funnels. In this genus only four species meet this criterion: Cernosvitoviella atrata, C. microtheca Rota & Healy, Citation1999, C. bulboducta Martinez‐Ansemil & Collado, Citation1996 and C. tridentina Dumnicka, Citation2004. Cernosvitoviella longiducta n. sp. differs from these species in some important features. The sperm duct of the new species is only terminally dilated (Figure ), whereas in C. microtheca, the ectal part of this duct is widened (Rota & Healy Citation1999) and in C. bulboducta, a spherical dilatation of its ental part is present (Martinez‐Ansemil & Collado Citation1996). The male genital apparatus of C. longiducta n. sp. partly resembles that of C. atrata, but these species can be easily distinguished by the shape of the sperm funnels. They are cylindrical with well‐formed collars in C. longiducta n. sp., and funnel‐shaped without distinct collars in C. atrata (Nielsen & Christensen Citation1959). The shape of the sperm funnel is considered the most consistent feature of C. atrata, which is a widespread and changeable taxon (Rota & Healy Citation1999). All the above‐mentioned species (except C. atrata) differ from C. longiducta n. sp. by the shape of the spermathecae. They are long and narrow reaching VII or VIII in C. bulboducta. The spermathecae of C. microtheca and C. tridentina are located in V, as in the new species, but they have spherical ampullae and short (C. microtheca) or long (C. tridentina) ectal ducts (Rota & Healy Citation1999; Dumnicka Citation2004), whereas C. longiducta has oval ampullae of the same length as the ectal ducts. Well‐developed seminal vesicles distinguish the new species from almost all other species of the genus Cernosvitoviella, except C. bulboducta, C. tridentina and C. immota (Knőllner, 1935).
Description of Cognettia valeriae n. sp.
Type series:
Holotype: whole mounted specimen, kept in the collection of the Institute of Nature Conservation, Polish Academy of Sciences, Krakow, Poland.
Type locality: The Noce Bianco stream (site NB3bis), Rhaetian Alps, Trentino, Italy, 46°24′N, 10°40′E, 2270 m a.s.l., 9 October 2001, leg. V. Lencioni and B. Maiolini.
Paratypes: one whole mounted specimen kept in the collection of the Institute of Nature Conservation, Polish Academy of Sciences, Krakow, Poland. The Noce Bianco stream (site NB2bis, replicate 1), Rhaetian Alps, Trentino, Italy, 46°24′N, 10°40′E, 2535 m a.s.l., 19 September 2000, leg. V. Lencioni and B. Maiolini.
Two whole mounted specimens kept in the HIGHEST collection. The Noce Bianco stream (site NB2bis replicate 5), Rhaetian Alps, Trentino, Italy, 46°24′N, 10°40′E, 2535 m a.s.l., 10 October 2001, leg. V. Lencioni and B. Maiolini.
Etymology
Dedicated to Dr Valeria Lencioni, hydrobiologist involved in ecological research on high mountain stream fauna, who collected and kindly provided this material for identification.
Characteristics
Medium size species – length of three complete, preserved specimens 12–15 mm and the number of segments 49–52. Prostomium short, with few big epidermal glands. Setae sigmoid, stout, without a nodulus. Along the whole body length, usually 3 setae per ventral bundle, their length increasing from 62–69 µm in preclitellar segments to 72–77 µm in postclitellar ones. Dorso‐lateral bundles of anterior and posterior segments containing 2 or 3 setae, similar in length to the ventral setae from the respective body part. Ventral setae absent in segment X (genital segment). Five pairs of well developed primary pharyngeal glands at 4/5–8/9, the glands of first two pairs united dorsally, in the remaining pairs glands free. Additionally, two pairs of small secondary glands are present in segments VII and VIII. Brain is longer than wide, slightly incised posteriorly (Figure ).
Figure 6 Cognettia valeriae n. sp. a, brain; b, coelomocytes; c, nephridium; d, penial bulb.
Coelomocytes oval or almost circular, sometimes with hair – like ends, with fine granules (Figure ). Their length varied from 26 to 41 µm and width from 14 to 30 µm.
Chloragogen cells weakly developed, and smaller than coelomocytes. Nephridia observed in segments located behind those with genital organs. The anteseptal part is small, thin, with postseptal part stout, plume‐shaped, sometimes almost spherical (effect of fixation?) with an efferent duct arising in its mid part (Figure ).
Clitellum not developed and gland cells absent on genital segments even in specimens with sperm in the spermathecal ampullae, and with developing eggs. Seminal vesicle probably unpaired well‐developed in segment IX. Sperm funnels cylindrical, 2.5–3 times longer than wide (length 182–210 µm, width 70–82 µm). Its length reaches 2/3 of the body's diameter. Sperm ducts long, 2–2.5 times longer than the sperm funnel, narrow (width 26–28 µm) and irregularly coiled in the genital segment. Small penial bulbs (width 71–76 µm, height 60–64 µm) (Figure ), situated in X. The width of the ectal ducts of the spermathecae 38–43 µm, their length a little shorter than the body's diameter. A small gland (diameter about 35 µm) at the ectal orifice. The ampullae consist of an ectal spherical enlargement (Figure ), which passes into a long connecting tube, wider than the ectal duct, and an almost globular ental chamber (Figure ). Sperm may be present in all parts of the ampulla with the highest concentration in the ectal enlargement (Figure ). Spermathecae confined to V or one ampulla reaching VI.
Figure 7 Cognettia valeriae n. sp. Details of spermatheca (marked by arrows): a, spherical enlargement of ampulla; b, final globular chamber.

Figure 8 Cognettia anomala, C. paxi, C. valeriae n. sp. Comparison of the shape of spermathecae. a, C. anomala (after Dumnicka Citation1976); b, C. paxi (after material from Careser stream, Italy, leg. V. Lencioni and B. Maiolini, det. E. Dumnicka); c, C. valeriae n. sp.
Taxonomical remarks
Cognettia valeriae n. sp. differs from the majority of species belonging to this genus such as: C. cognettii (Issel, 1905), C. lapponica Nurminen, Citation1965, C. hibernica Healy, Citation1975, C. clarae Bauer, Citation1993, C. bisetosa Christensen & Dózsa‐Farkas, Citation1999, C. quadrosetosa Christensen & Dózsa‐Farkas, Citation1999 and C. piperi Christensen & Dózsa‐Farkas, Citation1999 by a higher number of pharyngeal glands and the localization of the genital organs. In C. valeriae n. sp. these organs are located two segments forward, whereas in the remaining species mentioned above they are situated in segments XI–XII (as usual for the Enchytraeidae) (Nielsen & Christensen Citation1959; Nurminen Citation1965; Healy Citation1975; Bauer Citation1993; Christensen & Dózsa‐Farkas Citation1999).
Cognettia valeriae n. sp. belongs to a group of species having more than three pairs of primary pharyngeal glands and genital organs shifted forward. The new species differs from C. sphagnetorum and C. glandulosa (Michaelsen, 1888) by such features as: localization of the spermathecae and penial bulbs, dimension and localization of the seminal vesicle and number of secondary pharyngeal glands (Table ). However; the shape of spermatheca (i.e. ampulla articulated into three sections) is similar for these three species. C. sphagnetorum and C. glandulosa have one large, compact gland at the ectal orifice of the spermatheca, whereas in C. valeriae n. sp. this gland is small.
Table I. Comparison of taxonomic characters in Cognettia species having more than three pairs of pharyngeal glands.
In terms of general appearance the new species is similar to C. anomala (Černosvitov, Citation1928) and C. paxi (Moszyński, Citation1938), but differs from them by the shape of the spermatheca, which is simple (i.e. lacking an ectal enlargement) in these species and more complicated in C. valeriae n. sp. (Figure ).
C. paxi does not have a gland at the ectal orifice of the spermatheca and has a slightly modified posterior body end, resembling a sucker (Moszyński Citation1938). Moreover the shape of its nephridia is atypical for genus Cognettia, whereas in C. valeriae n. sp. the nephridia are similar to those in other Cognettia species. In both the compared species, C. paxi and C. valeriae n. sp., the clitellum was absent in all the specimens studied (Moszyński Citation1938 and personal observation), although the explanation of this fact is difficult (possibly due to the short period of reproduction, or self‐fertilization) and needs detailed investigation.
C. anomala differs from C. valeriae n. sp. by the dimension of the seminal vesicle which is small or absent in the first mentioned species (Černosvitov Citation1928) and well‐developed in C. valeriae n. sp. Moreover, according to Černosvitov's original figure, the sperm funnels of C. anomala are longer than the body diameter (Černosvitov Citation1928), while in C. valeriae n. sp. they are shorter.
Acknowledgments
The HIGHEST Project (Health and Integrity of Glacial Headwater EcoSystems in Trentino, 2001‐2004) was financed by the Autonomous Province of Trento, Italy. The Author would like to thank her Italian colleagues from the Museo Tridentino di Scienze Naturali, who were involved in the field campaigns and sorting of materials in the laboratory.
References
- Bauer , R. 1993 . Cognettia clarae n. sp. – eine neue Enchytraeiden‐Art aus einem ősterreichischen Fichtenwald (Oligochaeta; Enchytraeidae). . Linzer biologische Beitrage , 25 : 685 – 689 .
- Boscaini , A. , Corradini , F. , Lencioni , V. and Maiolini , B. 2004 . Caratterizzazione morfologica e chimico‐fisica di un sistema idrografico alpino (Parco Nazionale dello Stelvio, Trentino). . Studi Trentini di Scienze Naturali, Acta Biologica , 80 : 43 – 49 .
- Černosvitov , L. 1928 . Die Oligochaetenfauna der Karpathen. . Zoologische Jahrbűcher, Abteilung fűr Systematik , 55 : 1 – 28 .
- Christensen , B. and Dózsa‐Farkas , K. 1999 . The enchytraeid fauna of the Palearctic tundra (Oligochaeta, Enchytraeidae). . Biologiske Skrifter , 52 : 1 – 37 .
- Dumnicka , E. 1976 . Oligochaetes (Oligochaeta) of some streams of the High Tatra Mts and of the River Białka Tatrzańska. . Acta hydrobiologica , 18 : 305 – 315 .
- Dumnicka , E. 2004 . A description of Cernosvitoviella tridentina, a new species of Enchytraeidae (Oligochaeta) from the Italian Alps. . Annales de Limnologie – International Journal of Limnology , 40 : 133 – 137 .
- Dumnicka , E. and Boggero , A. 2007 . Freshwater Oligochaeta in two mountain ranges in Europe: The Tatra Mountains (Poland) and the Alps (Italy). . Archiv fűr Hydrobiologie , 163 : 231 – 242 .
- Graefe , U. and Schmeltz , R. M. 1999 . Indicator values, strategy types and life forms of terrestrial Enchytraeidae and other microannelids. . Newsletter on Enchytraeidae , 6 : 59 – 67 .
- Healy , B. 1975 . A description of five new species of Enchytraeidae (Oligochaeta) from Ireland. . Zoological Journal of the Linnean Society , 56 : 315 – 326 .
- Kownacki , A. , Galas , J. , Dumnicka , E. and Mielewczyk , S. 2000 . Invertebrate communities in permanent and temporary high mountain lakes (Tatra Mts). . Annales de Limnologie , 36 : 181 – 188 .
- Lencioni , V. , Dumnicka , E. and Maiolini , B. 2004 . The oligochaete fauna in high mountain streams (Trentino, NE Italy): Ecological and taxonomical remarks. . Studi Trentini di Scienze Naturali, Acta Biologica , 81 : 167 – 176 .
- 2002 . L'ecologia di un ecosistema acquatico alpino (Val de la Mare, Parco Nazionale dello Stelvio). . Natura alpina , 54 : 1 – 96 . Lencioni V, Maiolini B, eds.
- Malard , F. , Lafont , M. , Burgherr , P. and Ward , J. V. 2001 . A comparison of longitudinal patterns in hyporheic and benthic oligochaete assemblages in a glacial river. . Arctic, Antarctic and Alpine Research , 33 : 457 – 466 .
- Martinez‐Ansemil , E. and Collado , R. 1996 . Two new species of freshwater Oligochaeta from the North‐west Iberian Peninsula: Krenedrilus realis sp. nov. (Tubificidae) and Cernosvitoviella bulboducta sp. nov. (Enchytraeidae). . Journal of Zoology, London , 240 : 363 – 370 .
- Moszyński , A. 1938 . Die Fauna des Erdbodens. 1. Marionina paxi ein neuer Borstenwurm von den Haberwiesen. . Beitrage zur Biologie des Glatzer Schneeberges , 4 : 343 – 345 .
- Nielsen , C. O. and Christensen , B. 1959 . The Enchytraeidae. Critical revision and taxonomy of European species. . Natura Jutlandica , 8–9 : 1 – 160 .
- Nurminen , M. 1965 . Enchytraeids (Oligochaeta) from northern Norway and western Lapland. . Annales Zoologici Fennici , 2 : 11 – 15 .
- Piper , S. R. , MacLean , S. F. and Christensen , B. 1982 . Enchytraeidae (Oligochaeta) from taiga and tundra habitats of northeastern U.S.S.R. . Canadian Journal of Zoology , 60 : 2594 – 2609 .
- Rota , E. 1995 . Italian Enchytraeidae (Oligochaeta). I. . Bollettino di Zoologia , 62 : 183 – 231 .
- Rota , E. and Healy , B. 1999 . A taxonomic study of some Swedish Enchytraeidae (Oligochaeta), with descriptions of four new species and notes on the genus Fridericia. . Journal of Natural History , 33 : 29 – 64 .
- Rota , E. , Healy , B. and Erséus , C. 1998 . Biogeography and taxonomy of terrestrial Enchytraeidae (Oligochaeta) in northern Sweden, with comparative remarks on the genus Henlea. . Zoologischer Anzeiger , 237 : 155 – 169 .