Abstract
The diet of juvenile specimens of Raja radula from the Ionian Sea was investigated. The stomach contents of 43 specimens were analysed in order to evaluate composite index (IRI Index of Relative Importance). To assess feeding strategy and trophic niche width, prey-specific biomass plotted against frequency of occurrence and Levins index were used, respectively. The diet of juvenile specimens of R. radula consisted mainly of crustacean decapods followed by algae, polychaetes and molluscs. Among crustaceans, Liocarcinus spp. were the most important prey items, with four species with relatively high IRI values. Most specimens had been feeding on L. pusillus, L. depurator and polychaetes, but the variation of prey-specific biomass indicates that there is not a clear dominance of any prey and the value of niche width indicates a high variability in R. radula diet. Differences between sexes exists only in % IRI values related to Liocarcinus spp. The results indicate that R. radula feeds on a wide range of prey items, although crustaceans were the favoured food source.
Keywords:
Introduction
The rough ray Raja radula (Delaroche, 1809) is an endemic skate (Serena Citation2005), present in the Mediterranean Sea from the west (Stehmann & Bürkel Citation1984) to the east, especially in the Aegean Sea (Stergiou et al. Citation1997; Kallianiotis et al. Citation2000; Torcu & Aka Citation2000; Akyol Citation2003; Yeldan & Avsar Citation2007). Raja radula is endemic of the Mediterranean Sea: although also recorded from outside (probable misidentification), it is present in both western and eastern basins. In Italian waters, R. radula is relatively common along the Sardinian coasts and in the Sicily Strait only (Relini et al. Citation2000), where it inhabits the shallow South Mediterranean plateau on sandy and muddy bottoms down to 350 m. Although the rough ray is caught as by-catch of bottom trawl fisheries (Serena Citation2005), it is not a target species of the commercial fisheries, and data on its biology and ecology are incomplete. Published data are limited and concerned mainly the North Africa population, where this skate is commonly caught and landed at fishing sites in Tunisian waters (Mejri et al. Citation2004) and the Aegean Sea (Kyrtatos Citation1982). Capapé (Citation1974) studied maturity and reproductive cycle of R. radula, while Capapé and Quignard (Citation1975) assessed the fecundity of this species. Preliminary indicative aspects of the diet of R. radula were carried out by Capapé and Azouz (Citation1976) in the Gulf of Tunis, where the authors observed crustaceans and other small invertebrates as major prey items. Moreover, Abd El-Aziz (Citation1986) quantified food items in the waters of Alexandria (Egypt), considering the occurrence of prey categories in relation to size of specimen size and seasons. Here we provide the first data on diet of juveniles of R. radula from the Ionian Sea (South-Central Mediterran Sea), in order to improve the knowledge on this Mediterranean elasmobranch.
Material and methods
A total of 43 individuals were collected as by-catch by artisanal fisheries at depths between 4 and 20 m by means of a trammel net of 20 mm mesh, between May and July 2005, off Portopalo di Capo Passero in the South of Sicily ().
Figure 1. Map of the Ionian Sea, indicating sampling site.
Specimens were sexed and the hypothesis that sex ratio was 1:1 was tested using the χ2 test. Total length (TL) and disc width of each sample were measured to the nearest 0.5 cm; total weight of fresh specimens and preserved stomach contents were weighed to the nearest of 0.1 and 0.01 g, respectively. In order to evaluate TL frequency distribution, specimens were subdivided in length classes (34 mm range). Fish were dissected and sexual maturity stage was defined by macroscopic evaluation of gonads (Stehmann Citation2002); in addition, male maturity was assessed by classification of the claspers size according to Stehmann (Citation2002). The sexual stage of specimens agree with Mejri et al. (Citation2004), who support that males and females from the Gulf of Tunis are adult above 320 and 340 mm disc width, respectively. Stomachs were removed and fixed in 4% formalin solution with seawater and then stored in 80% ethanol. Prey residues found in the stomach were dried on paper towels, counted, weighed and identified to the lowest possible taxonomic level. Prey's frequency of occurrence (%F), percent numerical (%Cn), percent wet weight (%W) values of prey (Hureau Citation1970; Hyslop Citation1980; Cortés Citation1997) were calculated by species and higher taxonomical groups (fish, crustaceans, etc.); the values obtained were used to calculate the Index of Relative Importance (IRI) relativized to 100% (%IRI) of each prey category (Cailliet et al. Citation1986). The feeding strategy of R. radula was studied in terms of specialization and generalization by plotting prey-specific biomass P i = (SB i / SBt i ) × 100 against %F (Amundsen et al. Citation1996), where P i is the prey-specific biomass of prey i, SB i the stomach content biomass of prey i, and SBt i the total stomach content biomass in only those predators with prey i in their stomach. Trophic niche width was analysed by the standard Levin's index B = 1/Σp ij 2, where p ij 2 is the utilization share of resource j by species i. B values vary from 1 (when the species uses only a single resource category) to the numbers of recorded resource categories (when it uses all categories in equal proportions).
Results
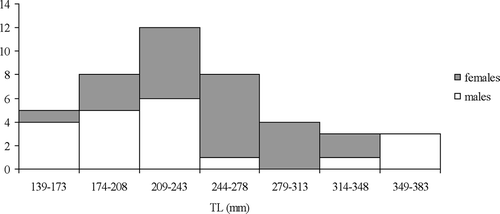
Analysed skates ranged from 139 to 383 mm TL (); the sample had an unimodal TL frequency distribution between 209 and 243 mm. Sex ratio was not significantly different from 1:1 (P > 0.001). From macroscopic evaluation of sexual maturity, we can categorize all specimens as juveniles. In fact we observed undeveloped claspers, small testes and thread-like sperm ducts in males while small and gelatinous ovaries without differentiated oocytes and narrow oviducts were found in females.
Figure 2. Total length frequency distribution of specimens of Raja radula.
Among the 43 stomachs only one was empty and 21 alimentary types were found and divided into four main groups: algae, Crustacea, Polychaeta, and Mollusca. Identification of items to species level has been possible only for crustacean decapods and Mollusca. %IRI values () for prey categories showed that the diet of Raja radula was mainly based on crustaceans, followed by algae, polychaetes and molluscs. Among crustaceans, the decapod Liocarcinus pusillus was the main ingested taxon (%IRI 45.25) followed by Liocarcinus depurator (12.38%) and Liocarcinus arquatus (5.82%); polychaetes and molluscs seem to hold a less important place in the diet (< 2%).
Table I. Diet composition of 43 Raja radula containing food (%F = percentage frequency of occurrence; %Cn = percent numerical composition; %W = percent wet weight composition; IRI = index of relative importance)
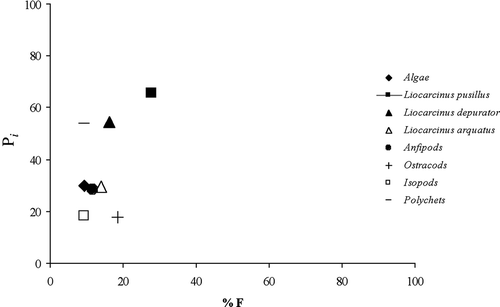
Raja radula shows a poor specialization for L. pusillus, L. depurator and polychaetes (); most specimens had been feeding on these categories, but small amounts of other prey types, such as amphipods, isopods, ostracods and algae, were included in the diet of some individuals. also shows that there is no clear dominance of any prey. The value of niche width (B = 10.6) of juvenile R. radula, calculated on the lowest specific level of preys, indicates a high variability in its diet.
Figure 3. Prey-specific biomass (P i ) plotted against frequency of occurrence (%F).
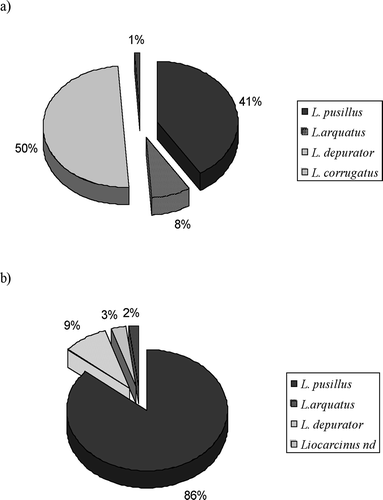
Dietary comparison between males and females was made evaluating variation in % IRI among Liocarcinus species () and some differences came to light: the most important prey items in males was L. pusillus (86%), other Liocarcinus species were relatively less important. In females there is a substantial repartition in %IRI values between L. pusillus (41%) and L. depurator (50%): the latter is less important in the males' diet (3%). L. arquatus was equally divided in males (8%) and females (9%).
Figure 4. %IRI values of Liocarcinus sp. in females (a) and males (b) of Raja radula.
Discussion
The high percentage of full stomachs found in R. radula can be explained both by a physiological increase in metabolism during the warmer seasons when predation is stimulated, and by the fact that skates usually eat frequently and it is common to find them with prey items (Ebert et al. Citation1991; Ellis et al. Citation1996); the high percentage of full stomachs might also be due to the sampling method (during the daytime).
Our analysis of prey categories found in the stomachs of juvenile R. radula, formerly categorized by Konstantinos et al. (Citation2002) as carnivorous with a preference for large decapods, indicated the rough ray as a predator with an extensive preference for the crustaceans. Although Kyrtatos (Citation1982) found small benthic fishes in the diet of R. radula, our results agree with previous studies (Capapé 1976; Abd El-Aziz Citation1986), confirming R. radula as a crustacean feeder. Almost all specimens fed on decapods of the genus Liocarcinus (Portunidae), although very small proportions of other benthic invertebrate categories were found, meaning that these latter were consumed only incidentally. Data about diet of R. radula in Egyptian Mediterranean waters (Abd El-Aziz Citation1986) show that small Crustacea such as crangonids, Mysidiacea and amphipods were the main food resource of smaller fishes; moreover, the contribution of crabs such as Portunidae increased with size. Differences in the relative importance of Portunidae in juvenile specimens of Raja radula between the Ionian Sea and north African coasts may be related to the geographical and environmental characteristics of the different areas.
Differences between sexes in % IRI values related to Liocarcinus spp. cannot be correlated to difference in size, since there is no sexual size dimorphism in juveniles of R. radula that can justify this preference. The small number of collected individuals made it impossible to evaluate this factor.
Unidentified marine vegetals found in the stomach of some specimens may be incidentally ingested during the feeding action, although this presence probably does not represent a problem, as the cellulase activity in the stomachs of elasmobranch fishes is well known (Stickney & Shumway Citation1974).
Ontogenetic dietary shifts have been reported for a large number of skates (Holden & Tucker Citation1974; Gray et al. Citation1997; Platell et al. Citation1998). Changes in diet and predatory efficiency are often related with differences in habitat (Schmitt & Holbrook Citation1984; Stergiou & Fourtouni Citation1991), which require alterations in mouth structures (Castro & Hernández-García Citation1995). It is known that both the mouth dimensions and the kind of dentition may affect the diet of different species of skates (Du Buit Citation1978–1979; Scharf et al. Citation2000). Based on the results of the MEDITS and GRU.N.D. cruises for the bathymetric distribution of R. radula (Relini Citation1995; Bertrand et al. Citation2000; Marano et al. Citation2002), the absence of adults in the sampling coastal area and ontogenetic shifts in diet related to growth (Abd El-Aziz Citation1986) allow us to hypothesize the existence of segregation between the juveniles and adults.
Moreover, the exclusive presence of young individuals and the non-significant differences of sex ratio found in the sample presumes Portopalo di Capo Passero coastal waters as an area of concentrations of juvenile individuals. Here, juveniles, less experienced at foraging and more vulnerable to predation, may find a broad availability of trophic resources due to reduction of direct intraspecific competition (Castro Citation1993).
Our results, although derived from few specimens in one season, provide the first focused information on trophic ecology of Raja radula from the Ionian Sea, representing a substantial step towards improving the knowledge of an endemic element of the endangered Mediterranean elasmobranch fauna, which will need of more attention in the future in order to avoid its collapse (Vacchi & Notarbartolo di Sciara Citation2000; Ferretti et al. Citation2008).
Related Research Data
References
- Abd El-Aziz , SH . 1986 . Food and feeding habits of Raja species (Batoidei) in the Mediterranean waters of Alexandria . Bulletin of the Institute Oceanography and Fisheries. Arab Republic Egypte , 12 : 265 – 276 .
- Akyol , O. 2003 . Retained and trash fish catches of beach-seining in the Aegean coast of Turkey . Turkish Journal of Veterinary and Animal Science , 27 : 1111 – 1117 .
- Amundsen , PA , Gabler , HM and Staldvik , FJ. 1996 . A new approach to graphical analysis of feeding strategy from stomach contents data modification of Costello (1990) method . Journal of Fish Biology , 48 : 607 – 614 .
- Bertrand , J , De , Gil , Sola , L , Papakostantinou , C , Relini , G and Souplet , A. 2000 . Contribution on the distribution of elasmobranchs in the Mediterranean (from the MEDITS surveys) . Biologia Marina Mediterranea , 7 : 385 – 399 .
- Cailliet , G , Love M , S. and Ebeling , AW. 1986 . Fishes: A field and laboratory manual on their structure, identification, and natural history , 194 Belmont, CA : Wadsworth Publishing Company .
- Capapé , C. 1974 . Contribution a la biologie des Rajidae des cotes tunisiennes. II. Raja radula Delaroche, 1809. Répartition géographique et bathymétrique, sexualité, reproduction . Archives de l'Institut Pasteur, de Tunisi , 51 : 211 – 228 .
- Capapé , C and Azouz , A. 1976 . Etude du regime alimentaire de deux Raies communes dans le golfe de Tunis: Raja miraletus Linné, 1758 et R. radula Delaroche, 1809 . Rapport Commission Internationale pour l'Exploration de la Mer Méditerranèe , 23 : 43 – 44 .
- Capapé , C and Quignard , JP. 1975 . Essai d’évaluation de la fécondité chez les sélaciens ovipares: cas de Raja miraletus Linné, 1758 et de R. radula Delaroche, 1809 des cotes tunisiennes . Archives de l'Institut Pasteur de Tunisi , 52 : 263 – 276 .
- Castro , JI. 1993 . The shark nursery of Bulls Bay, South Carolina, with a review of the sharks nurseries of the southeastern coast of United States . Environmental Biology of Fishes , 38 : 37 – 48 .
- Castro , JJ and Hernández-García , V. 1995 . Ontogenetic changes in mouth structures, foraging behaviour and habitat use of Scomber japonicus and Illex coindetii . Scientia Marina , 59 : 347 – 355 .
- Cortés , E. 1997 . A critical review of methods of studying fish feeding based on analysis of stomach contents: Application to elasmobranch fishes . Canadian Journal of Fisheries and Aquatic Sciences , 54 : 726 – 738 .
- Du Buit , MH . 1978–1979 . Remarques sur la denture des raies et sur leur alimentation . Vie et Milieu , 28–29 : 165 – 174 .
- Ebert , DA , Cowley , PD and Compagno , JLV. 1991 . A preliminary investigation on the feeding ecology of skates (Batoidea: Rajidae) off the west coast of Southern Africa . South Africa Journal of Marine Science , 10 : 71 – 81 .
- Ellis , JR , Pawson , MG and Shackley , SE. 1996 . The comparative feeding ecology of six species of shark and four species of ray (Elasmobranchii) in the north-east Atlantic . Journal of the Marine Biological Association UK , 76 : 89 – 106 .
- Ferretti , F , Myers , RA , Serena , F and Lotze , HK. 2008 . Loss of large predatory sharks from the Mediterranean Sea. Conservation . Biology , 22 : 952 – 964 .
- Gray , AE , Mulligan , TJ and Hannah , RW. 1997 . Food habits, occurrence, and population structure of the bat ray, Myliobatis californica in Humboltd Bay, California . Environmental Biology of Fishes , 70 : 133 – 143 .
- Holden , MJ and Tucker , RN. 1974 . The food of Raja clavata Linnaeus 1758, Raja montagui Fowler 1910, Raja naevus Müller and Henle 1841 and Raja brachyura Lafont 1873 in British waters . Journal du Conseil International pour l'Exploration de la Mer , 35 : 189 – 193 .
- Hureau , JC. 1970 . “ Biologie compare de quelques poissons antarctiques (Nototheniidae). Bulletin de l'Institute Ocèanographique Monaco Universitè de Paris, Paris ” . In Thèse d'Etat 244
- Hyslop , EJ. 1980 . Stomach contents analysis a review of methods and their application . Journal of Fish Biology , 17 : 411 – 429 .
- Kallianiotis , A , Sophronidis , K , Vidoris , P and Tselepides , A. 2000 . Demersal fishes and megafaunal assemblages on the Cretean continental shelf and slope (NE Mediterranean): Seasonal variation in species density, biomass and diversity . Progress in Oceanography , 46 : 429 – 455 .
- Konstantinos , I , Stergiou , I and Karpouzi , VS. 2002 . Feeling habits and trophic levels of Mediterranean fish . Reviews in Fish Biology and Fisheries , 11 : 217 – 254 .
- Kyrtatos , NA. 1982 . “ Investigation on fishing and biology of the most important fishes of the region around the Aegean Sea ” . In Island of Tinos. Thalassographica 5 (special publication) 88
- Marano , CA , Manfrin Piccinetti , G , Pasolini , P , Tinti , F and Ungaro , N. 2002 . Annotated checklist of the skates (Chondrichthyes, Rajidae) in the Adriatic Sea . Biologia Marina Mediterranea , 10 : 856 – 862 .
- Mejri , H , Ben Souissi , J , Zaouali , J , El Abed , A , Vergne , Y , Guélorget , O and Capapé , C. 2004 . On the recent occurrence of Elasmobranch species in Tunis southern lagoon (Northern Tunisia, Central Mediterranean) . Annales Series Historia Naturalis , 14 : 143 – 158 .
- Platell , ME , Potter , IC and Clarke , KL. 1998 . Resource partitioning by four species of elasmobranchs (Batoidea: Urolophidae) in coastal waters of temperate Australia . Marine Biology , 131 : 719 – 734 .
- Relini , G. 1995 . La fauna ittica batiale del Mediterraneo con particolare riferimento ai campionamenti dello strascico . Biologia Marina Mediterranea , 2 : 177 – 183 .
- Relini , G , Biagi , F , Serena , F , Belluscio , A , Spedicato , MT Rinelli , P . 2000 . I selaci pescati con lo strascico nei mari italiani . Biologia Marina Mediterranea , 7 : 347 – 384 .
- Scharf , FS , Juanes , F and Rountree , RA. 2000 . Predator size–prey size relationships of marine fish predators: Interspecific variation and effects of ontogeny and body size on trophic-niche breadth . Marine Ecology Progress Series , 208 : 229 – 248 .
- Schmitt , RJ and Holbrook , SJ. 1984 . Ontogeny of prey selection by black surfperch Embiotoca jacksoni (Pisces: Embiotocidae): The roles of fish morphology, foraging behaviour, and patch selection . Marine Ecology Progress Series , 18 : 225 – 239 .
- Serena , F. 2005 . Field identification guide to the sharks and rays of the Mediterranean and Black Sea , 136 Rome : FAO Species Identification Guides for Fishery Purposes Rome .
- Stehmann , M. 2002 . Proposal of a maturity stage scale for oviparous and viviparous cartilaginous fishes (Pisces, Chondrichthyes) . Archive of Fishery and Marine Research , 50 : 23 – 48 .
- Stehmann , M and Bürkel , DL. 1984 . Rajidae , Edited by: Whitehead , PJP , Bauchot , ML , Hureau , JC , Nielsen , J and Tortonese , E . 163 – 196 . Paris : UNESCO .
- Stergiou , KI and Fourtouni , H. 1991 . Food habits, ontogenetic diet shift and selectivity in Zeus faber Linnaeus, 1758 . Journal of Fish Biology , 39 : 589 – 603 .
- Stergiou , KI , Politou , CY , Chrstou , ED and Petrakis , G. 1997 . Selectivity experiments in the NE Mediterranean: The effect of trawl codend mesh size on species diversity and discards. ICES . Journal of Marine Science , 54 : 774 – 786 .
- Stickney , RR and Shumway , SE. 1974 . Occurrence of cellulose activity in the stomachs of fishes . Journal of Fish Biology , 6 : 779 – 790 .
- Torcu , H and Aka , Z. 2000 . A study on the fishes of Edremit Bay (Aegean Sea) . Turkish Journal of Zoology , 24 : 45 – 61 .
- Vacchi , M , di , Notarbartolo and Sciara , G. 2000 . I pesci cartilaginei dei mari italiani, una risorsa che richiede urgenti misure di tutela . Biologia Marina Mediterranea , 7 : 296 – 311 .
- Yeldan , H and Avsar , D. 2007 . Length–weight relationship for five elasmobranch species from the Cilician Basin shelf waters (Northeastern Mediterranean) . Journal of Applied Ichthyology , 23 : 713 – 714 .