Abstract
Male age influence on male mating success, courtship, mating activities, fecundity, fertility, longevity, percentage of egg–larval hatchability and percentage of larva–adult viability has been studied in Drosophila ananassae. It was noticed that females of D. ananassae were able to discriminate males on the basis of male age and preferred to mate with old-aged males more frequently than younger or middle-aged males using a female choice experiment. Old-aged males significantly mated faster, performed greater courtship activities, copulated longer and inseminated more females in a given unit of time, showed greater fecundity, fertility, percentage of egg–larval hatchability, percentage of larva–adult viability, and females mated to old-aged males lived shorter than those mated to younger or middle-aged males. Thus in D. ananassae females discriminate males on the basis of age and females mated to old-aged males had significantly greater reproductive success than that of females mated to younger or middle-aged males.
Introduction
The age of mating individuals is apparently a key factor in reproduction with effects on their sexual performance and progeny production (Pervez et al. Citation2004). The investigations on the effects of age on life events in insects include mate choice, mating incidence, activity levels and reproduction. The mate choice in insects is predominantly influenced by the age of mates. It was widely held that females should prefer to mate with older males in species where males provide only sperm to females, because viability selection leads to older males with higher genotype quality than young males (Trivers Citation1972; Manning Citation1985; Andersson Citation1994). In support of this hypothesis, a simulation model revealed a strong preference for older males under different eco conditions (Kokko & Lindstrom Citation1996). Several empirical studies have suggested that females do indeed prefer to mate with older males for qualitative and quantitative progeny production (Zuk Citation1988; Manning Citation1989; Simmons & Zuk Citation1992; Simmons Citation1995). However, some have argued that males of young and intermediate ages are better mates, as they have the highest breeding values for fitness and less prone to deleterious mutations (Hansen & Price Citation1995; Beck & Powell Citation2000).
In species where males do not provide parental care or any direct benefits to the mated female except providing sperm and accessory gland protein, even in such species female shows a preference for mating with a particular age class of individuals. The species of the genus Drosophila is one such genus and comprises more than 3000 Drosophila species. Males do not provide parental care or provide material benefits to mated females. Very few attempts have been made to study the role of male age in mating success in Drosophila. It has been found that females of Drosophila prefer to mate with older males more frequently than middle-aged or younger males (Somashekar & Krishna Citation2011). However, more studies are required using different species of Drosophila.
Therefore, the present study has been under taken in D. ananassae, a domestic cosmopolitan species belonging to the bipectinata complex of the ananassae subgroup of the melanogaster species group (Bock & Wheeler Citation1972) to study male age influence on male reproductive success.
Materials and methods
Experimental stock
Experimental stock of D. ananassae was established from an isofemale line (Polymorphic for inversion) collected at semi-domestic localities at Mysore, India. In each generation 40 flies (20 males and 20 females) were cultured per quarter pint milk bottles (250 ml) containing wheat cream agar medium and were maintained at 21°C ± 1°C and a relative humidity of 70% (main experimental stock). This procedure was continued for three generations to allow them to acclimatize to the laboratory condition. At the fourth generation eggs were collected from these flies using Delcour's procedure (Citation1969). The first batch of eggs collected for 3 h were discarded. Subsequent collected eggs in 1 h duration were used in the experiment to study male age influence on male reproductive performance. Eggs (100) were seeded in a vial containing wheat cream agar medium. When adults emerged from these eggs, virgin female and unmated males were isolated within 3 h of their eclosion and were aged in a vial in groups of four flies and individually until they were used in the experiment. Before assigning male age classes, we studied the longevity of male of D. ananassae by transferring individually unmated males into a vial containing wheat cream agar medium once a week and maintaining them in the same condition. This process was continued until their death and longevity was recorded. A total of 50 replicates were made and mean longevity data showed 63 ± 2 days. Since mean longevity of D. ananassae ranges from 60 to 65 days, we assigned days for young-, middle- and old-aged male as follows [young age male (two to three days), middle-aged male (24–25 days), and old-aged male (46–47 days)]. In addition to these male age classes, we also collected virgin five- to six-day-old females from the main experimental stock for use in the present experiment.
Mate choice tests
Five- to six-day-old virgin females and unmated young, middle-, and old-aged males were used to study male age influence on male mating success using a female mate choice experiment. A female along with two males of different male age classes (we painted India ink onto sons of young age males in one trial and sons of old-aged males in an alternate trial on the thorax region) were individually transferred into an Elens–Wattiaux mating chamber (Citation1964) and observation was made for 1 h. When mating occurred, pairs in copulation were aspirated out from the mating chamber. The rejected male in the female mate choice experiment was also transferred to a new vial. A total of 50 of trials were made separately for each combination of female mate choice experiment and Chi-square analysis was applied to the data. We also measured wing length of 50 selected and rejected males in each combination of the female mate choice experiment following the procedure of Hegde and Krishna (Citation1997) and a Paired t-test was applied to the data.
Male courtship displays
We used unmated young, middle- and old-aged males and five- to six-day-old virgin females to study male age influence on male mating activities. A female along with a male (young/middle/old) were individually transferred into an Elens–Wattiaux mating chamber and observed for 1 h. Pairs unmated within 1 h were discarded. We recorded mating latency (time between introduction of male and female together into mating chamber until initiation of copulation of each pair) and copulation duration (time between initiation of copulation to termination of copulation of each pair). We also quantified courtship acts such as tapping, scissoring, vibration, licking, circling, ignoring, extruding and decamping following the procedure of Hegde and Krishna (Citation1997). The behavior of male and female was recorded simultaneously but separately by two observers for 1 h; the number of pairs mated was also recorded.
Fitness evaluation
Soon after copulation as above, the mated female was individually aspirated into a new vial containing wheat cream agar medium to check whether insemination had occurred (by observing larval activity). The mated male was allowed to mate with a second female (virgin five- to six-days-old); if mating occurred with the second female, we allowed the pair to complete copulation and checked for insemination as above. This process was continued and the number of females inseminated by each male in 1 h was recorded as male mating ability. All the mated females by each male were individually transferred to a new vial once in 24 h until their death to study fecundity (number of eggs laid), fertility (progeny production) and longevity. After counting the eggs as above, eggs (100) were taken randomly using a spatula to study egg–larval hatchability and were seeded separately for each of the three male age classes in a small Petri dish containing wheat cream agar medium. The number of first-instar larvae that emerged from the above eggs was counted to account for percentage of egg–larval hatchability. For studying larval–adult viability, 100 first-instar larvae were collected as above and placed in a Petri dish and the number of adult flies that emerged from these larvae were also counted to account for percentage of larval–adult viability.
Statistical analysis
A total of 50 trials were used separately for each of the three male age classes. One-way ANOVA followed by Tukey's Honest post-hoc test (Tukey's test) was carried out on data of mating activities, courtship activities, fecundity, fertility, longevity, percentage of egg–larval hatchability and percentage of larval–adult viability using the SPSS 10.0 Programme.
Results
Females of D. ananassae generally chose to mate with the older males in trials involving two males and one female. [In 82% of cases, older males were successful in trials involving young and old-aged males and a female (χ2=20.48; df=1; P<0.05; N = 50). In 72% of cases, middle-aged males were successful in trials involving young and middle-aged males and a female (χ2=9.68 df=1; P<0.05; N = 50). In 66% of cases, old-aged males were successful in crosses involving middle- and old-aged males and a female (χ2=5.12; df=1; P<0.05; N = 50)].
The mean wing length of rejected males was slightly greater than that of the mean wing length of selected males in all the combinations studied () Paired t-tests carried out on mean wing length data of selected and rejected males showed no difference in size.
Table I. Mean wing length (in mm) of selected and rejected males of D. ananassae in female choice experiment (values are mean±SE)
and show mean values of courtship, mating and fitness data. It was noticed that old-aged males had the lowest mating latency, copulated longest, performed the greatest male courtship activities (tapping, scissoring, vibration, licking, circling) male mating ability, fecundity, percentage of egg–larval hatchability, percentage of larval–adult viability and fertility, while the reverse trend was found in the above characters in young males. Females showed the least rejection responses (ignoring, extruding and decamping) to old-aged males, while they showed greater rejection responses to young males. Significant variation was noticed. Tukey's test showed that old-aged males had significantly lesser mating latency, greater copulation duration, mating activities, fecundity, fertility, percentage of egg–larval hatchability, percentage of larval–adult viability than young or middle-aged males except vibration. Vibration of young and middle-aged males was found to be insignificant by Tukey's test. Similarly, middle-aged males had significantly greater mating latency, copulation duration, mating activities, fecundity, percentage of egg–larval hatchability, percentage of larva–adult viability, and fertility compared to young males. Females showed significantly greater rejection responses to young males compared to old- and middle-aged males
Figure 1. Male age influence on female fitness traits of D. ananassae (values are mean ±SE). Different letter on bar graph indicates significant variation by Tukey's test. YAM, young male (2–3 days); MAM, middle-aged male (24–25 days); OAM, old-aged male (46–47 days)
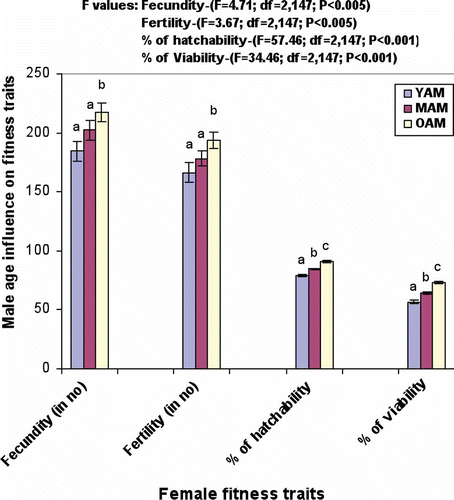
Table II. Male age influence on male courtship and mating activities of D. ananassae (values are mean±SE)
Correlation matrices calculated between male age and male reproductive performance () showed that male age was positively correlated with copulation duration, male courtship activities, male mating ability, fecundity, fertility, percentage of egg–larval hatchability, and percentage of larva–adult viability, while male age was negatively correlated with male mating latency, female courtship activities (ignoring, extruding and decamping) and female longevity.
Table III. Pearson correlation between male age and male reproductive performance of D. ananassae.
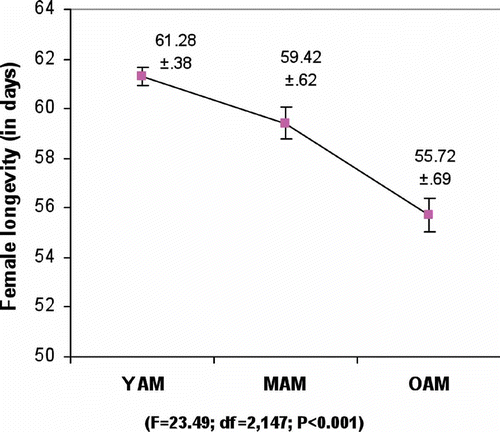
shows average female longevity. It reveals that females mated with young males lived longest followed by females mated with middle-aged males, and females mated with old-aged male lived the shortest length of time. This suggests that female survival decreased with increased male age. Significant variation was noticed among different male age classes. Tukey's test on average female longevity data showed that females mated with young males had significantly greater longevity compared to females mated with old- and middle-aged males.
Figure 2. Male age influence on female longevity of D. ananassae (values are mean ±SE). YAM, young male (2–3 days); MAM, middle-aged male (24–25 days); OAM, old-aged male (46–47 days).
Discussion
Most compelling studies in Drosophila suggest that females of Drosophila select male on the basis of male size (). Male age is another trait used by females to select a mate. In the present study, females of D. ananassae were able to discriminate males on the basis of male age, and they preferred to mate more frequently with old-aged males than with young or middle-aged males. This confirms the earlier studies in D. melanogaster and D. simulans (Moulin et al. Citation2001) and in D. pseudoobscura (Avent et al. Citation2008). They also found that females of D. pseudoobscura prefer to mate with old-aged males over young males. This result is also consistent with some previous observation in other insects and birds (Zuk Citation1988; Conner Citation1989; Hassalquist et al. Citation1996) and also with the theoretical models of good gene hypothesis (Kokko Citation1997, Citation1998), suggesting that female preference for old-aged males can be an evolutionarily stable strategy (Avent et al. Citation2008). In contrast to this, the studies of Hansen and Price (Citation1995) and Price and Hansen (Citation1998) in D. melanogaster have argued that females prefer to mate with young males more frequently than old-aged males. However, empirical evidence in this regard is absent in Drosophila.
Table IV. Influence of male traits on female fitness in Drosophila.
Insignificant variations were also found in mean wing length of selected and rejected males and even in some combination mean wing length of rejected males were slightly longer than that of selected males (). This suggests that the observed greater mating success of old-aged males was not due to a difference in the mean wing length between young, middle- and old-aged males; instead, it could be attributed to male age itself.
Another potential factor known to influence male mating success in Drosophila is male activities and female receptivity during courtship (Bastock Citation1956; Hegde & Krishna Citation1997). Old-aged males mated faster (as the time is reverse of the speed), copulated longer and performed greater courtship activities than young or middle-aged males (Tables II and III). Through these activities old-aged males convinced the females faster and had greater mating success, because these courtship acts allow the male to communicate with the female through visual, chemical, tactile and auditory stimuli and as a result they increases female receptivity (Speith Citation1966; Tompkins et al. Citation1982; Hegde & Krishna Citation1997). This confirms the earlier studies of courtship in different species of Drosophila suggesting that flies which show greater activities during courtship have greater mating success than males which show fewer activities (Hegde & Krishna Citation1997). Females of D. ananassae showed fewer rejection activities, i.e. extruding, decamping, ignoring old-aged males compared to young or middle-aged males (Tables II and III). This suggests the influence of male age on female receptivity. This confirms earlier studies of Drosophila where females which show fewer rejection responses have greater receptivity and greater mating success than females which show greater rejection to courting males (Hegde & Krishna Citation1997)
and shows that females mated with old-aged males lived for a significantly shorter period than females mated with young or middle-aged males. This suggests a significant influence of male age on female longevity too. It is not known whether the accessory gland secretion that is transferred to the female during copulation could have carried a greater harmful effect with increasing male age or the delayed mating of males up to 46 days might have caused a greater harmful effect in their ejaculate, resulting in reduction in female longevity. However, these causes have not been tested presently.
The reproductive success of males not only involves success in mating but also includes how many females he could inseminate in a given time, the number of eggs laid and progeny produced (Krishna & Hegde Citation1997). Old-aged males inseminated a greater number of females in a given unit of time and females mated to old-aged males had a greater percentage of egg–larval hatchability, percentage of larval–adult viability, and produced more progenies than females mated with young or middle-aged males, confirming the greater reproductive success of old-aged males.
Thus these studies in D. ananassae suggest that females of D. ananassae discriminate males on the basis of age and older males had greater reproductive success over young or middle-aged males.
Acknowledgements
The authors are grateful to the Professor and Chairman, Department of Studies in Zoology, University of Mysore, for providing facilities. M. Prathibha is also grateful to UGC for awarding teacher fellowship to carry out this work. This work is dedicated to our teacher Prof. S.N. Hegde, Department of Studies in Zoology, University of Mysore, Mysore – 570006.
Related Research Data
References
- Andersson , M. 1994 . Sexual selection , Princeton, NJ : Princeton University Press .
- Avent , TD , Price , TAR and Wedell , N. 2008 . Age based female preference in the fruit fly Drosophila pseudoobscura . Animal Behaviour , 75 : 1413 – 1421 .
- Bastock , M. 1956 . A gene mutation that changes a behaviour pattern . Evolution , 10 : 421 – 439 .
- Beck , CW and Powell , LA. 2000 . Evolution of female mate choice based on male age: Are older males better mates? . Evolutionary Ecology Research , 2 : 107 – 118 .
- Bock , LR and Wheeler , MR. 1972 . Drosophila melanogaster species group . University of Texas Publication , 7213 : 1 – 102 .
- Conner , J. 1989 . Older males have higher insemination success in a beetle . Animal Behaviour , 38 : 503 – 509 .
- Delcour , J. 1969 . A rapid and efficient method of egg collecting . Drosophila Information Service , 44 : 133 – 134 .
- Elens , AA and Wattiaux , JM. 1964 . Direct observation of sexual isolation . Drosophila Information Service , 39 : 118 – 119 .
- Hansen , TF and Price , DK. 1995 . Good genes and old age: Do old mates provide superior genes? . Journal of Evolutionary Biology. , 8 : 759 – 778 .
- Hasselquist , D , Bensch , S and Von Schantz , T. 1996 . Correlation between male song repertoire, extra-pair paternity and offspring survival in the great reed warbler . Nature , 381 : 229 – 232 .
- Hegde , SN and Krishna , MS. 1997 . Size assortative mating in Drosophila malerkotliana . Animal Behavior , 54 : 419 – 426 .
- Kokko , H. 1997 . Evolutionary stable strategies of age-dependent sexual advertisement . Behavioral Ecology and Sociobiology , 41 : 99 – 107 .
- Kokko , H. 1998 . Good genes, old age and life-history trade-offs . Evolutionary Ecology , 12 : 739 – 750 .
- Kokko , H and Lindstrom , J. Evolution of female preference for old males . Proceedings of the Royal Society of London: Series B . Vol. 263 , pp. 1533 – 1538 .
- Krishna , MS and Hegde , SN. 1997 . Reproductive success of large and small flies in D. bipectinata complex . Current Science , 72 : 747 – 750 .
- Krishna , MS and Hegde , SN. 2003 . Influence of body size in mating success in three sympatric species of Drosophila . Italian Journal of Zoology , 70 : 47 – 52 .
- Manning , JT. 1985 . Choosy females and correlates of male age . Journal of Theoretical Biology , 116 : 349 – 354 .
- Manning , JT. 1989 . Age-advertisement and the evolution of the peacock train . Journal of Evolutionary Biology , 2 : 379 – 384 .
- Moulin , BF , Rybak , T , Aubin , J and Jallon , M. 2001 . Compared ontogenesis of courtship song components of males from the sibling species, D. melanogaster and D. simulans . Behavioural Genetics , 31 : 299 – 308 .
- Partridge , L , Ewings , A and Chandler , A. 1987 . Male size and mating success in D. melanogaster. The role of male and female behaviour . Animal Behavior , 35 : 552 – 555 .
- Pervez , A , Omkar and Richmond , AS. 2004 . The influence of age on reproductive performance of the predatory ladybird beetle, Propylea dissecta . Journal of Insect Science , 4 : 22
- Price , DK and Hansen , TF. 1998 . How does offspring quality change with age in male Drosophila melanogaster? . Behavioral Genetics , 28 : 395 – 402 .
- Simmons , LW. 1995 . Correlates of male quality in the field cricket, Gryllus campestris L.: Age, size, and symmetry determine pairing success in field populations . Behavior, Ecology , 6 : 376 – 381 .
- Simmons , LW and Zuk , M. 1992 . Variability in call structure and pairing success of male field crickets, Gryllus bimaculatus: The effects of age, size and parasite load . Animal Behavior , 44 : 1145 – 1152 .
- Somashekar , K and Krishna , MS. 2011 . Evidence of female preference for older males in Drosophila bipectinata . Zoological Studies , 50 : 1 – 15 .
- Speith , HT. 1966 . Mating behaviour of D. ananassae and ananassae-like flies from the Pacific . University of Texas Publication , 6615 : 133 – 145 .
- Tompkins , L , Gross , AC , Hall , JC , Gailey , DA and Siegal , RW. 1982 . The role of female movement in sexual behaviour of D. melanogaster . Behavioral Genetics , 12 : 295 – 307 .
- Trivers , RL. 1972 . “ Parental investment and sexual selection ” . In Sexual selection and the descent of man, 1871–1971 , Edited by: Campbell , B . 136 – 179 . Chicago, IL : Aldine Press .
- Zuk , M. 1988 . Parasite load, body size, and age of wild-caught male field crickets (Orthoptera: Gryllidae) effects on sexual selection . Evolution , 42 : 969 – 976 .