Abstract
Six species of Ophelina Ørsted, Citation1843 have been identified in an extensive material covering 145 localities from 575 to 3900 m depth in the Nordic Seas. Ophelina opisthobranchiata Wirén, Citation1901 and O. helgolandiae Augener, Citation1912 were recorded for the first time since their original description about a century ago. O. opisthobranchiata was the most abundant species with more than 18,000 collected specimens. It was recorded between 800 and 3900 m depth and was the only species of Ophelina found deeper than 2000 m. Ophelina cylindricaudata (Hansen, Citation1879) and Ophelina abranchiata Støp-Bowitz, Citation1948 were both widespread from 600 to about 2000 m depth. A new species, Ophelina brattegardi sp. nov., is described from off East Greenland, and another presumably new species, Ophelina ‘B’, from shelf break areas off western Norway is diagnosed but not formally described due to insufficient material. O. cylindricaudata and O. abranchiata are redescribed based on type material and newly collected specimens. Ammotrypane minuta Annenkova, Citation1952 from deep water in the Greenland Sea is newly synonymised with Ophelina helgolandiae. Descriptions are also provided for O. opisthobranchiata and O. helgolandiae. The present study illustrates the importance of examining a relatively large amount of specimens in order to correctly establish characters of fragile structures such as branchiae and anal tube. It is also shown that the number of setigers is stable for all recorded species and consequently represents a character of taxonomic importance.
Introduction
The Nordic Seas (Greenland, Iceland and Norwegian Seas) represent the connection between the northern part of the Atlantic Ocean and the Arctic Ocean. The largest depth (> 4200 m) is found between Iceland and northern Norway. The deep water of the Nordic Seas is separated from deep water of the North Atlantic by a ridge running from Shetland across Iceland to Greenland. In the north, a connection to the deep water of the Arctic Ocean runs through a deep trough between Greenland and Svalbard, the Fram Strait. The water masses below ridge threshold level (650 m water depth) are of Arctic origin and have temperatures below 0°C (Blindheim & Østerhus Citation2005).
Studies of the benthic fauna hitherto conducted in this area have generally indicated that the deep-water fauna of the Nordic Seas is sparse and of low diversity (Stuart & Rex Citation2009). Several groups, however, are poorly studied and very imperfectly known. In several cases most relevant information actually dates back from pioneering expeditions in the nineteenth and early twentieth centuries.
The present study is based on material from epibenthic and infaunal samples collected during 21 scientific cruises in the period 1981–2009 to different parts of the Nordic Seas, and on material from environmental monitoring at the shelf break off western Norway. The investigated localities cover a depth range from 575 to 3900 m. Opheliids from a total of 145 localities have been examined. A majority of the samples were taken in the Norwegian Sea, some are from the Iceland Sea and a handful from the margin between the Norwegian Sea and the Greenland Sea. Based on initial sorting of the samples to family level, opheliids were found to be one of the dominant polychaete groups in the present material.
Five species of Ophelina have previously been reported from deep water habitats in the Nordic Seas. Ophelina cylindricaudata (Hansen, Citation1879) and Ophelina abranchiata Støp-Bowitz, Citation1948, have been regularly recorded (Støp-Bowitz Citation1945, Citation1948; Wesenberg-Lund Citation1950; Eliason Citation1962; Hartmann-Schröder Citation1974), but mainly from shallow waters, and little is known about their occurrence and distribution in the deep sea. The other species, Ophelina opisthobranchiata Wirén, Citation1901, Ophelina helgolandiae Augener, Citation1912, and Ophelina minuta (Annenkova, Citation1952) have not been recorded since their original description. Ophelina opisthobranchiata and O. helgolandiae were both originally described from deep water off Spitsbergen, whereas O. minuta was described from deep water in the Greenland Sea.
In the present study, six species of Ophelina were recorded. These include four of the previously reported species, among them O. opisthobranchiata and O. helgolandiae, and one species which is new to science. All are here described. Redescriptions are provided for O. abranchiata and O. cylindricaudata based on re-examination of type material and additional specimens of good condition from the present material. A lectotype for O. cylindricaudata has been designated. The last species, here designated Ophelina ‘B’, is diagnosed but not formally described due to insufficient material. Based on the original species description, O. minuta is proposed as a junior synonym for O. helgolandiae.
Material and methods
Most of the material included in this study was collected with epibenthic sledges, namely a D-sledge which is a predecessor of the ‘Sneli-sledge’ (Sneli Citation1998), and an RP-sledge (Brattegard & Fosså Citation1991). Material made available from environmental studies and from the MAREANO project was sampled using a box-corer or grab. A station list comprising all included samples is presented in Appendix 1.
Type material of Ophelina abranchiata and Ophelina cylindricaudata was borrowed from the Swedish Museum of Natural History, Stockholm (SMNH) and Bergen Museum, University of Bergen (ZMBN), respectively.
Samples were identified using dissecting and compound microscopes. Staining with methyl blue has been used to aid in identification. A Canon 20D camera, with MP-E 65 mm and EF 100 mm macro lenses, a Leica DM6000B compound microscope and a Leica M205C stereo microscope were used for digital photos of specimens, parapodia and setae. The Leica LAS software was used to take compound images with the ‘Z-stack’ option. Sketch drawings are based on digital photos. Final drawings were prepared in Adobe Illustrator 4.0, and Adobe Photoshop CS 8.0. The material is deposited at the collections of Bergen Museum, University of Bergen and the Museum of Natural History and Archaeology, Norwegian University of Science and Technology, Trondheim (VM).
All body length measurements are excluding anal tube.
Recorded species
Family Opheliidae Malmgren, Citation 1867
Genus Ophelina Ørsted, Citation 1843
Ophelina abranchiata Støp-Bowitz, Citation 1948
, 6A
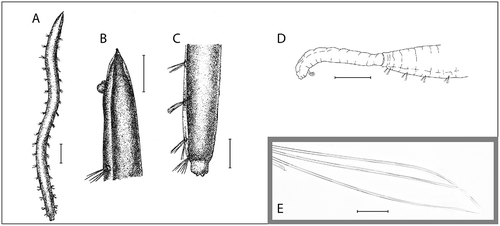
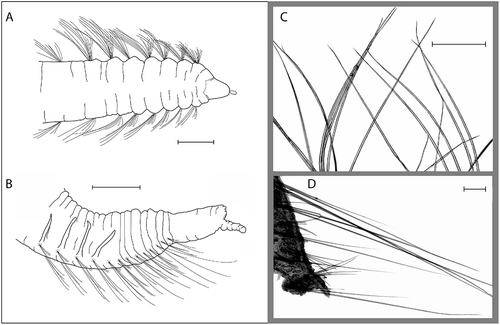
Figure 1. Ophelina abranchiata Støp-Bowitz, Citation1948. A–C, holotype (SMNH type-2118), after Støp-Bowitz Citation1948. A, complete specimen, dorsal view; B, anterior part of body, dorso-lateral view; C, posterior part of body, dorso-lateral view (anal tube missing); D, posterior part of body with anal tube and short (broken) ventral cirrus, lateral view (Environmental monitoring, stn OL-03); E, capillary setae from mid-body setiger (R/V ‘H. Mosby’, stn 83.06.08.2). Scale bars: A,D, 1.0 mm; B,C, 0.5 mm; E, 50 μm.
Ophelina abranchiata Støp-Bowitz, Citation1948, pp. 22–23, fig. 7; Hartman Citation1965, p. 186; Hartman & Fauchald Citation1971, p. 129.
? Ammotrypane abranchiata – Eliason Citation1962, pp. 273–274, fig. 19.
Type material
Holotype (SMNH type-2118) in fairly good condition, anal tube missing, 9 mm long and 0.6 mm wide, with 18 setigerous segments ().
Type locality
East Greenland, 72°28′N 21°48′W, 180 m depth.
Material examined
Holotype, Swedish Arctic Expedition 1899, 24 July 1899. R/V ‘H. Mosby’, stns: 81.03.21.1 (28 specimens); 81.03.22.1 (35); 81.06.04.4 (18); 81.06.06.7 (20); 81.06.07.1 (3); 81.06.07.3 (3); 81.08.13.2 (2); 81.08.15.5 (15); 81.08.15.6 (4); 81.08.16.3 (15); 81.08.16.4 (3); 81.08.16.7 (11); 82.01.21.4 (5); 82.01.21.6 (1); 82.08.15.1 (4); 82.08.19.1 (12); 82.11.26.1 (1); 82.11.27.1 (1); 83.06.08.1 (1); 83.06.08.2 (12); 83.06.17.3 (22); 84.03.15.2 (4); 84.05.23.1 (4); 84.11.20.2 (4); 85.01.08.2 (6); 85.01.08.3 (21); 85.01.08.4 (1); 85.01.12.2 (7); 85.01.12.3 (26); 86.06.13.1 (6); 86.07.25.1 (19); 86.07.27.2 (28); 86.07.30.1 (9); 86.07.31.1 (8); 86.08.15.7 (10); 86.08.16.2 (44); 86.08.16.3 (12); 86.08.17.3 (2); 86.08.17.4 (6); 86.08.17.5 (4); 86.08.17.6 (55); 87.06.12.1 (1); 87.06.13.2 (5); 87.06.17.1 (1). R/V ‘Meteor’, stn: M507/90 (1). MAREANO, stns: R229-27 (1); R231-30 (2); R232-34 (1); R257-43 (1); R259-41 (3); R301-349 (1). Environmental monitoring, stns: V-06 (9); V-14 (1); V-16 (1); OL-01 (1); OL-02 (9); OL-03 (33); OL-04 (14); OL-05 (23); OL-06 (9); OL-07 (17); OL-08 (12); OL-09 (4); OL-10 (8); OL-11 (32); OL-12 (5); OL-13 (25).
Diagnosis
Up to 10 mm long and 0.8 mm wide for 18 setigers. Branchiae absent. Anal tube very long, tubular, with long, thread-like ventral cirrus arising from near posterior end. Last setiger with short cylindrical sleeve for attachment of anal tube (anal tube and ventral cirrus often lost).
Redescription
Holotype 9 mm long and 0.6 mm wide for 18 setigers. Large specimens up to 10 mm long (excluding anal tube) and 0.8 mm wide. All examined specimens with 18 setigerous segments. Ventral and lateral grooves distinct along whole length of body. Colour in alcohol white to light tan, often with reddish pigmentation on the ventral nerve cord.
Prostomium elongate, conical with small acute terminal palpode. Eyes not observed. Nuchal organs rounded, laterally on posterior part of prostomium. Peristomium indistinct. Proboscis tongue-like, asymmetric in the dorsal–ventral plane with distinct ventral groove. Mouth situated below and behind proboscis.
Anterior region of body (first segment?) asetous (). Segmental furrows and annulations indistinct. All setigers shorter than wide; anterior- and posteriormost setigers slightly shorter than mid-body setigers. Branchiae absent. Parapodia rudimentary, biramous, embedded in lateral grooves; no distinct pre- or postsetal lobes. Setae all slender, smooth capillaries (), appear in small numbers in both rami. Notosetae mostly longer than neurosetae.
Last setiger with posterior cylindrical sleeve connecting to anal tube (). Junction between sleeve and anal tube distinct as a deep groove. Anal tube tubular, transparent and transversely ringed, with distal opening with short ventral incision and ventrally situated anal cirrus (). Anal tube up to 1/6–1/8 of total length of worm. Cirrus as long as anal tube or more. Anal tube and cirrus fragile, often lost. Anal tube missing in holotype ().
Methyl blue staining pattern
Dorsal transverse bands on last two setigers and posterior sleeve connecting to anal tube.
Distribution
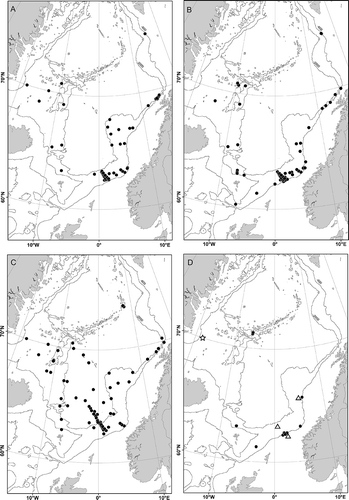
Widely distributed in the Nordic Seas from about 600 to 2000 m depth (). Also recorded from deep water habitats in the Atlantic Ocean (Hartman Citation1965; Hartman & Fauchald Citation1971).
Remarks
This species was originally described with a short cylindrical anal tube with small terminal papillae (Støp-Bowitz Citation1948). However, Eliason (Citation1962) described specimens from the Skagerrak, as ‘?Ammotrypane abranchiata’, with a very long, tubular anal tube, and with a long, threadlike ventral cirrus. He also examined the holotype of O. abranchiata, and concluded that the anal tube is missing in the holotype. Based on re-examination of the holotype we fully agree with Eliason in his conclusion. In the present study, only a small percentage of the specimens examined possessed the anal tube. Of these again, only very few also had the ventral cirrus intact. As in many other opheliids, both the anal tube and the ventral cirrus are fragile structures, which are frequently lost. The specific status of specimens from the Skagerrak is uncertain, however. As noted by Eliason (Citation1962), specimens from the Skagerrak have 17 setigerous segments, in contrast to 18 in specimens from the Nordic Seas. Work is in progress to evaluate the status of the form from Skagerrak.
Blake (Citation2000) described a morphologically similar species, O. farallonensis, from off California in slope depths (1020–3060 m), which he separated from O. abranchiata mainly by the presence of a long anal tube in O. farallonensis. Comparing the description of O. farallonensis (Blake Citation2000), and the description of O. abranchiata provided here, the only obvious difference between the two species is the size, in that adult specimens of O. abranchiata (10 mm long and 0.8 wide) appear to be considerably larger than O. farallonensis (4.5 mm long and 0.35 mm wide). A comparative study of specimens of both species is needed in order to decide upon the validity of O. farallonensis.
Ophelina brattegardi sp. nov.
, 6B
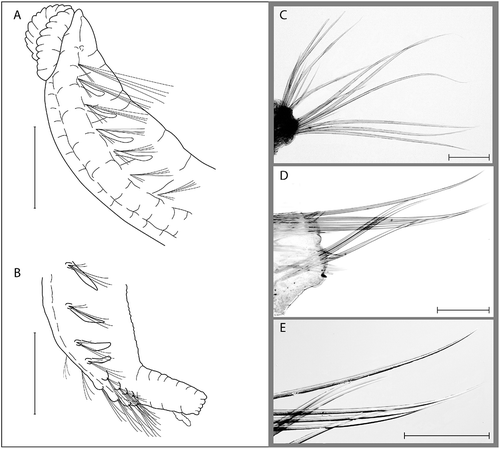
Figure 2. Ophelina brattegardi sp. nov. A,B, holotype (ZMBN 86306); C–E, paratype (ZMBN 86307). A, anterior part of body, ventro-lateral view; B, posterior part of body with anal tube, ventro-lateral view; C, capillary setae from mid-body setiger; D, capillary setae from setiger 28; E, same, close-up. Scale bars: A,B, 0.5 mm; C–E, 100 μm.
Type locality
Iceland Sea, off East Greenland, 69.955N–18.148W, 1600 m depth.
Type material
R/V ‘H. Mosby’, stn 87.06.12.1, holotype (ZMBN 86306) and 11 paratypes (ZMBN 86307).
Diagnosis
Up to 9 mm long and 0.6 mm wide for 28 setigers. Branchiae present on setiger 2–6 and 20–24. Posterior 4 setigers with raised parapodia in a ventral position, with tufts of numerous stiff setae in both rami. Anal tube cylindrical, gradually tapering towards distal end, with small rounded papillae distally. Ventral cirrus short, inserted near post erior end.
Description
Holotype, mature female 8 mm long (excluding anal tube) and 0.6 mm wide. Paratypes ranging from about 6 mm long and 0.4 mm wide to about 9 mm long and 0.6 mm wide. All examined specimens with 28 setigers. Deep ventral groove and distinct lateral grooves along whole length of body. Colour in alcohol white.
Prostomium conical with acute tip, without terminal palpode (). Nuchal organs well developed, more or less circular, laterally on posterior part of prostomium. Peristomium indistinct. Eyes absent. Proboscis tongue-like, about 3 times longer than wide, asymmetric in the dorsal–ventral plane with distinct ventral groove. Mouth situated below and behind proboscis ().
Segmentation indistinct. Mid-body setigers about as long as wide, anterior and posterior setigers shorter. Parapodia distinct, biramous; embedded in lateral grooves on setiger 1–24; no distinct pre- or postsetal lobes. Parapodia on last 4 setigers raised and shifted to a ventral position with numerous stiff setae in both rami (). All setae smooth capillaries (–E). Branchiae cirriform, length up to half body width, present on setigers 2–5 and 20–24.
Anal tube cylindrical, 2.5–3 times longer than wide, gradually tapering towards distal end (). Surface of anal tube with several transverse indistinct furrows. Posterior rim with several small rounded papillae. Ventral cirrus short, wide at base and tapering distally, wrinkled in appearance, inserted about 2/3 the length towards posterior end of anal tube.
Methyl blue staining pattern
No discernable pattern.
Distribution
Only known from the type locality ().
Etymology
The species is named for Torleiv Brattegard in recognition of his contribution to the knowledge of the deep-sea fauna of the Nordic Seas. He also collected the specimens here described as a new species.
Remarks
The present species belongs to a group of species within Ophelina with the 4 posterior parapodia turned ventrally, crowded and pad-like, with cylindrical anal tube, and with 24–28 setigers. Mid-body branchiae are generally reduced in size or lacking. The group encompasses Ophelina cylindricaudata (Hansen, Citation1879) O. breviata (Ehlers, Citation1913), O. chaetifera (Hartman, Citation1965), O. minima Hartmann-Schröder, Citation1974, and one or more forms reported as O. cylindricaudata from North Atlantic deep water (Hartman Citation1965; Hartman & Fauchald Citation1971), the Antarctic (Hartmann-Schröder & Rosenfeldt Citation1989) and southern oceans (Maciolek & Blake Citation2006; as O. cf. cylindricaudata). The new species differs from O. cylindricaudata, O. breviata and O. minima in lacking mid-body branchiae and by having an anal tube with a papillate rim. It differs from Ophelina chaetifera which lacks mid-body branchiae, in that this species has 26 setigers and only two anterior and three posterior pairs of branchiae. The forms reported as O. cylindricaudata have 28 setigers and lack mid-body branchiae, but the number of anterior branchiae is reported to be higher than in the new species and the anal tube has dorsal lobes instead of a papillate rim (Hartman Citation1965; Hartman & Fauchald Citation1971; Hartmann-Schröder & Rosenfeldt Citation1989).
Ophelina cylindricaudata (Hansen, Citation 1879 )
, 6D
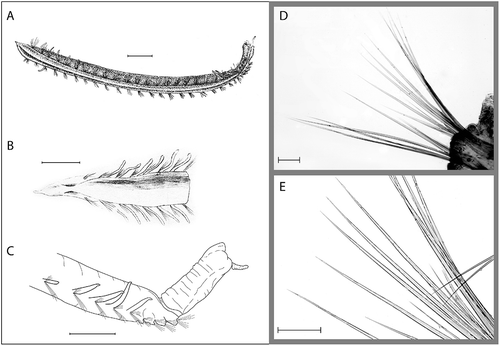
Figure 3. Ophelina cylindricaudata (Hansen, Citation1879). A, complete specimen, lateral view, after Støp-Bowitz Citation1945; B, lectotype (ZMBN 2269), anterior part of body, dorsal view, after Hansen Citation1879; C, posterior part of body with anal tube, lateral view (R/V ‘H. Mosby’, stn 83.06.17.3); D, capillary setae from mid-body setiger (R/V ‘H. Mosby’, stn 84.11.21.2); E, same, close-up. Scale bars: A–C, 1.0 mm; D,E, 100 μm.
Ammotrypane cylindricaudatus Hansen, Citation1879, pp. 8–9, Pl. VI, figs. 1–8; 1882, pp. 36–37, tab VI, figs. 20–28; Wirén Citation1883, p. 406; Levinsen 1887, pp. 295–296; Fauvel Citation1911, p. 32; Citation1914, pp. 245–246; Wesenberg-Lund Citation1950, p. 38; 1953, p. 73.
Ophelina cylindricaudata – Augener Citation1928, p. 747; Støp-Bowitz Citation1945, pp. 49−53, fig. 5; 1948, pp. 18−20, fig. 5; Bakken et al. Citation2010, p. 12.
Type material
The type material consists of two syntypes, fairly well preserved, both with 28 setigerous segments. The larger syntype, here designated as lectotype, has been cut in three pieces with, respectively, 7 (anterior), 12 (middle) and 9 (posterior) setigers. The anterior and posterior fragments agree in detail with illustrations in the original description. The specimen has probably been cut for the purpose of examination and illustration. The smaller specimen (paralectotype) is complete, but lacks branchiae and anal tube. The specimens measure, respectively, 24 mm (including the anal tube of 2 mm length) and 19 mm, width about 1 mm. The specimens have been placed in one vial with reference to the Norwegian North-Atlantic Expedition (NNHE) 1876–78 stns 31 and 87. Here the deepest locality with most stable temperature (NNHE stn 87) is selected as type locality (see below). The two localities are relatively close to each other on the Norwegian shelf break, however.
Type locality
Continental shelf break off the west coast of Norway, 64°02′ N, 05°53′ E, 911 m, mud, bottom temperature −1.0°C (NNHE stn 87 selected as type locality).
Material examined
The Norwegian North-Atlantic Expedition 1876–78 stns 31 and 87, off the west coast of Norway, 763–911 m depth, lectotype (ZMBN 2269) and 1 paralectotype (ZMBN 86308). R/V ‘H. Mosby’, stns: 81.03.21.1 (18 specimens); 81.03.22.1 (1); 81.06.06.7 (4); 81.06.07.1 (4); 81.08.13.2 (1); 81.08.16.3 (1); 81.08.16.4 (3); 81.08.16.7 (26); 82.01.21.2 (12); 82.01.21.4 (8); 82.01.21.6 (2); 82.08.15.1 (2); 82.08.23.1 (15); 82.11.26.1 (22); 82.11.27.1 (4); 83.06.02.1 (36); 83.06.03.1 (2); 83.06.03.2 (170); 83.06.08.1 (6); 83.06.08.2 (8); 83.06.17.3 (106); 84.03.15.2 (18); 84.05.23.1 (235); 84.05.23.2 (19); 84.05.23.3 (17); 84.05.23.5 (4); 84.05.23.7 (3); 84.11.20.2 (21); 84.11.21.1 (1); 84.11.21.2 (69); 85.01.08.1 (246); 85.01.08.2 (22); 85.01.08.3 (21); 85.01.08.4 (2); 85.01.12.2 (3); 85.01.12.3 (5); 86.06.13.1 (2); 86.06.13.4 (1); 86.06.13.5 (1); 86.06.16.1 (1); 86.07.25.1 (4); 86.07.27.2 (1); 86.07.27.5 (1); 86.08.15.5 (81); 86.08.15.7 (1); 86.08.16.2 (5); 86.08.17.5 (19); 86.08.17.6 (67). R/V ‘Meteor’, stns M410/90 (2); M507/90 (1). R/V ‘Jan Mayen’, stn: 848–99 (1). MAREANO, stns: R209-17 (11); R209-18 (8); R232-34 (1); R276-334 (2); R297-346 (1); R351-356 (1). Environmental monitoring, stns: V-06 (1); V-07 (5); V-09 (1); V-16 (1); OL-01 (2); OL-03 (3); OL-04 (1); OL-08 (1); OL-09 (1); OL-10 (3); OL-11 (3).
Diagnosis
Up to 34 mm long and 2 mm wide for 28 setigers. Branchiae present on setigers 2–24, often reduced in midbody setigers; anterior 6–7 pairs and posterior 4 pairs of branchiae characteristically long. Posterior 4 setigers close together and with raised parapodia with tufts of numerous short, stiff setae in both rami. Anal tube about 4 times longer than wide, with ventral groove. Posterior rim smooth, with swollen dorsal rounded papilla overhanging two small ventral papillae.
Redescription
Lectotype 24 mm long and 1 mm wide, in three pieces. Paralectotype 19 mm long and 1 mm wide (anal tube lost). Large specimens up to about 34 mm long and 2 mm wide for 28 setigers. Deep ventral groove and distinct lateral grooves along whole length of body (). Colour in alcohol white, light tan or brown.
Prostomium conical (), with narrow terminal palpode (often lost). Nuchal organs well developed, more or less circular, laterally on posterior part of prostomium. Peristomium indistinct. Eyes not observed. Proboscis tongue-like, asymmetric in the dorsal–ventral plane with distinct ventral groove. Mouth situated below and behind proboscis.
Segmentation indistinct. Midbody setigers about as long as wide, anterior and posterior setigers shorter. Parapodia distinct, biramous; embedded in lateral grooves on setigers 1–24; no distinct pre- or postsetal lobes. Parapodia on last 4 setigers raised and shifted to a ventral position, with numerous short stiff setae in both rami. All setae smooth capillaries (,E).
Branchiae cirriform, present on setigers 2–24, fragile and often lost. Anterior 7 pairs and posterior 4 pairs of branchiae characteristically long, up to half body width. Branchiae in mid-body setigers small and often hardly visible, sometimes apparenty absent in smaller specimens. Type specimens with few rudimentary branchiae present in mid-body setigers.
Anal tube cylindrical, 3–4 times longer than wide (), with ventral groove of body continuing ventrally for whole length. Distal end of anal tube with swollen dorsal papilla overhanging two smaller ventral papillae, one on each side of ventral groove. Ventral cirrus relatively short and thick, inserted between ventral papillae near distal end of anal tube. Ventral cirrus extremely fragile, usually missing.
Methyl blue staining pattern
Prostomial palpode strongly stained. Long branchiae of setigers 2–7 and 20–24 moderately stained as longitudinal central band on lateral surface. Anal tube with distinct patch dorsally on distal end.
Distribution
Widely distributed in the Nordic Seas to about 2000 m depth (). Records of this species from other parts of the world, e.g. the central North Atlantic and Antarctic, remains uncertain (see remarks).
Remarks
Støp-Bowitz (Citation1945) gave a description of the species based on the two syntypes and two specimens from coastal shallow water in southern Norway. He corrected the number of setigers to 28, in contrast to 34 as indicated in the original description by Hansen (Citation1879). He also noted that the indicated length (5 cm) was erroneous and suggested 1.5 cm instead. Later, Støp-Bowitz (Citation1948) reported specimens with 26 or 27 setigers from more shallow water. Specimens from shallow water appear to vary, however, also in other respects (J.A.K., pers. obs.) and may represent different species. In this study, all examined specimens from deep-water habitats were found to have 28 setigers. The variable number of setigers reported by e.g. Hartmann-Schröder (Citation1996) (28–34) does not seem warranted.
In the original description, Hansen (Citation1879) stated that branchiae were lacking in the middle part of the body. This was commented by Wirén (Citation1883) and Levinsen (1887), who noted that specimens from the Kara Sea had rudimentary branchiae, and later by Støp-Bowitz (Citation1945), who considered that branchiae most probably had been lost, and consequently that lack of branchiae in mid-body setigers was not a species specific character. Rudimentary branchiae in some mid-body setigers is also present in the lectotype. In the present study, several specimens lacking mid-body branchiae were found, but there was no consistent pattern and in many cases small scattered branchiae were found on close examination. It is therefore concluded that O. cylindricaudata has branchiae on all setigers except the first and last four.
Ophelina cylindricaudata was reported from North Atlantic deep water by Hartman (Citation1965) and Hartman and Fauchald (Citation1971), and the Antarctic by Hartmann-Schröder and Rosenfeldt (Citation1989) and Maciolek and Blake (Citation2006) (as O. cf. cylindricaudata) using lack of branchiae in mid-body as one of the specific characters. These reports should probably be referred to separate species. In addition, specimens from the Antarctic also lack the prostomial palpode (Maciolek & Blake Citation2006). The reported specimens are close to O. cylindricaudata, however, in other characters, e.g. the number of setigers, the presence of ventrally turned pad-like posterior parapodia and the shape of the anal tube.
Ophelina helgolandiae Augener, 1912
, 6C
Figure 4. Ophelina helgolandiae Augener, Citation1912. A,B, holotype, after Augener, Citation1912. A, anterior part of body, dorsal view; B, posterior part of body with anal tube, lateral view; C, posterior part of body with anal tube, lateral view (R/V ‘H. Mosby’, stn 84.05.23.5); D, capillary setae from mid-body setiger (R/V ‘H. Mosby’, stn 84.05.23.3); E, same, close-up. Scale bars: A–C, 1.0 mm; D,E, 100 μm.
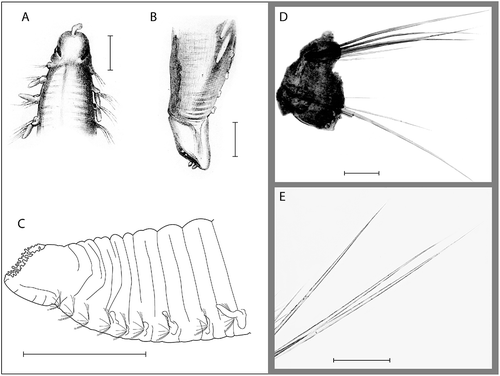
Figure 5. Ophelina opisthobranchiata Wirén, Citation1901 (R/V ‘H. Mosby’, stn 85.01.12.1). A, anterior part of body, dorsal view; B, posterior part of body with anal tube, lateral view; C, capillary setae from mid-body setiger; D, capillary setae from posterior setigers. Scale bars: A,B, 0.5 mm; C,D, 100 μm.
Ophelina helgolandiae Augener, Citation1912, pp. 174–176, figs. 9–11.
Ammotrypane minuta Annenkova, Citation1952, pp. 151–152.
Type locality
Spitsbergen, 1000 m depth.
Material examined
R/V ‘H. Mosby’, stns: 81.03.21.1 (1 specimen); 81.06.06.3 (3); 82.01.21.2 (1); 83.06.03.1 (1); 84.05.23.2 (1); 84.05.23.3 (109); 84.05.23.5 (18); 84.05.23.7 (2); 84.11.21.2 (1); 86.06.13.4 (1). R/V ‘G.O. Sars’, stns: Dive-07 (2); Dive-12 (1); Dive-18 (1).
Diagnosis
Up to 27 mm long and 1.8 mm wide for 33 setigers. Branchiae present from setiger 2 to 27–30. Anal tube short and wide, opening inclined facing upwards, with thickened ventral keel, posterior rim with numerous small cirri. Ventral cirrus absent.
Description
Large specimens up to 27 mm long and 1.8 mm wide for 33 setigers. Deep ventral groove and distinct lateral grooves along whole length of body. Colour in alcohol tan to brown ().
Figure 6. Habitus photos of recorded species. A, Ophelina abranchiata Støp-Bowitz, Citation1948 (R/V ‘H. Mosby’, stn 83.06.08.2); B, Ophelina brattegardi sp. nov., paratype (ZMBN 86307); C, Ophelina helgolandiae Augener, Citation1912 (R/V ‘H. Mosby’, stn 84.05.23.3); D, Ophelina cylindricaudata (Hansen, Citation1879) (R/V ‘H. Mosby’, stn 84.11.21.2); E, Ophelina opisthobranchiata Wirén, Citation1901 (R/V ‘H. Mosby’, stn 85.01.12.1). Scale bars: 1.0 mm.

Figure 7. Maps of the Nordic Seas with records in present study. A, Ophelina abranchiata Støp-Bowitz, Citation1948; B,
Ophelina cylindricaudata (Hansen, Citation1879); C, Ophelina opisthobranchiata Wirén, Citation1901; D, Ophelina helgolandiae Augener, Citation1912 (•), Ophelina brattegardi sp. nov. (
Prostomium quadrangular to conical, with flattened frontal part (). Prostomial palpode relatively large, elongate, clavate. Nuchal organs present as large rounded to oval spots laterally near posterior end of prostomium (). Eyes not observed. Peristomium distinct dorsally and laterally. Proboscis tongue-like, asymmetric in the dorsal-ventral plane with distinct ventral groove. Mouth situated below and behind proboscis.
Body with indistinct segmental furrows. All setigers shorter than wide; anterior and posterior setigers shorter than mid-body setigers, with several more or less distinct annulations. Posterior setigers comparatively high. Parapodia as rounded lobes, biramous, embedded in lateral grooves; no distinct pre- or postsetal lobes. Anterior parapodia with generally more and longer setae than mid-body and posterior parapodia. Setae all smooth capillaries (,E).
Branchiae cirriform, starting on setiger 2, continuing to setigers 27–30. Branchiae of more or less similar length throughout, up to about half body width, fragile and often lost ().
Anal tube cylindrical, shorter than wide, about as long as last 3 setigers. Tube thin-walled, with thickened keel-like ventral structure continuing slightly longer than lateral and dorsal parts (,C, 6C). Opening inclined and facing upwards, rimmed with numerous small cirri. Ventral cirrus not observed.
Distribution
Sporadic, but widely distributed in the Nordic Seas from off western Norway, north of the Faroe Islands and north of the island of Jan Mayen, in 600 to 1300 m depth ().
Remarks
The original description given by Augener (Citation1912) is comprehensive and well illustrated. The specimens examined in the present study agree in detail with the description. The species is particularly characterised by the short anal tube with an inclined opening facing upwards. In addition, the body is relatively high with short segments in the most posterior part.
Annenkova (Citation1952) described Ammotrypane minuta from deep-water in the Greenland Sea. The description is short and without illustrations. The species is stated to have 33 setigers, branchiae from setiger 2 to posterior end, a short anal tube with even margin carrying small spindle-shaped papillae, and no anal cirrus. The description agrees with O. helgolandiae in all characters, except that the number of abranchiate posterior setigers is stated to be 2 instead of 3–6 as observed in the present material. Most likely A. minuta is conspecific with O. helgolandiae (type material was not available for study).
Ophelina opisthobranchiata Wirén, 1901
; 6E
Ophelina opisthobranchiata Wirén, Citation1901, p. 253; Augener Citation1928, p. 747; Støp-Bowitz Citation1945, p. 53; 1948, p. 20.
Type locality
Spitsbergen, at great depths.
Material examined
R/V ‘H. Mosby’, stns: 81.03.21.1 (1 specimen); 81.03.22.1 (2); 81.06.03.5 (120); 81.06.04.4 (8); 81.06.07.3 (208); 81.06.08.1 (78); 81.08.13.2 (10); 81.08.13.5 (5); 81.08.14.1 (43); 81.08.14.5 (483); 81.08.15.2 (412); 81.08.15.4 (117); 81.08.15.5 (6); 81.08.15.6 (3); 81.08.16.7 (1); 82.08.15.1 (10); 82.08.19.1 (2); 82.11.21.2 (∼500); 82.11.21.4 (16); 82.11.24.1 (∼2000); 83.06.08.2 (26); 83.06.09.1 (204); 83.06.10.1 (∼450); 83.06.11.1 (511); 84.03.13.1 (∼600); 84.03.13.2 (7); 84.03.15.2 (84); 84.03.16.2 (350); 84.03.17.1 (2); 84.03.17.2 (∼700); 84.11.20.2 (2); 85.01.08.3 (1); 85.01.08.4 (15); 85.01.09.1 (261); 85.01.10.1 (∼800); 85.01.10.2 (4); 85.01.10.3 (13); 85.01.11.1 (620); 85.01.11.2 (∼1000); 85.01.11.3 (∼1000); 85.01.12.1 (∼1000); 85.01.12.2 (48); 85.01.12.3 (50); 86.06.12.2 (1); 86.07.24.1 (∼500); 86.07.26.1 (306); 86.07.28.1 (∼1000); 86.07.30.1 (51); 86.07.31.1 (395); 86.08.15.7 (1); 86.08.16.2 (46); 86.08.16.3 (112); 86.08.16.5 (∼1000); 86.08.17.1 (1054); 86.08.17.2 (∼700); 86.08.17.3 (∼600); 86.08.17.4 (47); 86.08.17.5 (4); 87.06.10.2 (13); 87.06.12.1 (1); 87.06.13.1 (4); 87.06.13.2 (8); 87.06.15.1 (9); 87.06.15.2 (5); 87.06.16.1 (43); 87.06.17.2 (1); 87.06.18.1 (23). R/V ‘G.O. Sars’, stns: SL-1 (34); SL-2 (1); RP-1 (253). MAREANO, stns: R209-17 (1); R232-33 (1); R259-41 (1); R301-348 (2); R301-349 (3); R322-353 (4); R322-354 (3). Environmental monitoring, stn V-14 (1).
Diagnosis
Up to 20 mm long and 1 mm wide for 38–39 setigers. Anterior setigers with numerous long, slightly curved setae. Branchiae present in posterior part of body, from about setiger 22 to about setiger 31. Anal tube cylindrical with short, thick and wrinkled ventral cirrus, with numerous small terminal cirri.
Description
Large specimens up to about 20 mm long and 1 mm wide for 38–39 setigers. Ventral groove present along whole length of body. Posterior body laterally flattened, usually slightly upwards curved (preserved condition). Colour in alcohol uniformly white or light tan, often with numerous small brownish dots on ventral side of anal tube.
Prostomium conical with distinct terminal palpode (). Peristomium indistinct. Nuchal organs visible as narrow lateral slits or rounded spots near posterior end of prostomium; eyes not observed. Proboscis tongue-like, asymmetric in the dorsal–ventral plane with distinct ventral groove. Mouth situated below and behind proboscis.
Anterior 7 setigers shorter than wide; mid-body setigers about as long as wide; posterior 7–8 setigers all very short. Parapodia biramous. Anterior 7–8 parapodia distinct with well-developed notopodial and neuropodial rounded lobes; with numerous long, slightly posteriorly curved setae (). Mid-body and posterior parapodia smaller, embedded in lateral grooves, with relatively few straight setae. Posterior parapodia with stiff and straight setae, some being considerably longer than the rest (,D). All setae smooth capillaries (,D).
Branchiae present in posterior part of body from about setiger 22 to 31, often lost. Branchiae cirriform, with relatively narrow base and slightly tapering distal end. Posteriormost branchiae usually the longest, about as long as body width.
Anal tube cylindrical, about twice as long as wide (near base) and as long as last 6–7 setigers, with small ventral incision (). Posterior margin with several small cirri, gradually decreasing in length ventrally. Ventral cirrus short, relatively thick, uniformly ringed, arising distally from near ventral incision.
Methyl blue staining pattern
Anterior part of the prostomium uniformly stained; terminal palpode without staining. Ventral part of anal tube distinctly stained.
Distribution
Widely distributed in deep waters from 800 to 3900 m depth in the Nordic Seas ().
Remarks
The original description of O. opisthobranchiata is only a brief diagnosis without illustrations (Wirén Citation1901). Only two characters are mentioned: the presence of long, curved setae in anterior setigers, and branchiae restricted to posterior part of the body (Wirén Citation1901). However, these characters separate O. opisthobranchiata from all other known opheliids in the Nordic Seas. Original material of Ophelina opisthobranchiata could not be traced in any Nordic museum. Although the present specimens cannot be compared to original material, it is held very likely that they represent the same species. The original description only indicates records from ‘great depths off Spitsbergen’.
O. opisthobranchiata appears to be the most common deep-water opheliid in the Nordic Seas. The present material is rather large, consisting of about 18,000 specimens from 73 localities. The species has not been found shallower than 800 m and seems to be restricted to temperatures below 0°C. Støp-Bowitz (Citation1948), having no material at hand, considered the species as dubious. His remarks, combined with few studies of deep-water fauna, may be the reason why the species has not been reported until the present study.
In the present material of O. opisthobranchiata, the anal tube is usually present, while the ventral cirrus was only observed in a small percentage of the numerous specimens examined.
O. opisthobranchiata is similar to Ammotrypanella arctica McIntosh, Citation1879 from the Davis Strait, west Greenland, and three recently described species referred to Ammotrypanella from South Atlantic and Antarctic deep water (Schüller Citation2008). All species have anterior tufts of long setae and branchiae limited to posterior setigers. A. arctica was redescribed by Schüller (Citation2008), and differs from O. opisthobranchiata by lacking ventral cirrus and by having a higher number of setigers. The holotype is an incomplete specimen with 42 setigers. The other species may be separated on anal tube characters. Ammotrypanella cirrosa Müller, 2008 from the Weddel Sea is most close to O. opisthobranchiata in having a similar anal tube with cirrus, branchiae on the same setigers (23–31), and about the same number of setigers (37–42). The maximum number of setigers is higher than in O. opisthobranchiata, but a closer comparison of the two species may be necessary, however, to further clarify any specific differences.
Schüller (Citation2008) redefined the genus Ammotrypanella to include species with long anterior setae and branchiae limited to posterior part of the body. Essentially this implies a splitting of the genus Ophelina (in the wide sense) based on characters which are generally variable. Within Ophelina other characters such as number of setigers and shape and position of posterior parapodia may as well be used to separate between groups of species (e.g. O. cylindricaudata, O. minima and O. brattegardi sp. nov.). Based on the current knowledge there are no valued justification of splitting the genus Ophelina and also which characters that would be of taxonomic value at generic level involving other genera (Eibye-Jacobsen Citation2002). As species in Ophelina display a wide range of variation in morphological characters, a revision of Opheliidae should be based on a phylogenetic analysis. For these reasons, we prefer to retain the species O. opisthobranchiata in the genus Ophelina where it was originally described.
Ophelina ‘B’
Material examined
R/V ‘H. Mosby’, stns: 81.06.06.8 (2 specimens); 82.08.15.1 (1); 84.05.23.7 (11).
Diagnosis
Mature specimens up to 9 mm long and 0.5 mm wide for 25 setigers. Branchiae present on setigers 2–22. Anal tube long and narrow, cylindrical, about 3–4 times longer than wide, with 6–7 small cirri on posterior rim. Ventral cirrus not observed.
Distribution
Shelf break areas off western Norway, 575–1286 m depth ().
Remarks
Ophelina ‘B’ presumably represents an undescribed species, but a formal description of the species is not included here due to unsatisfactory material.
Concluding remarks
The present study demonstrates the lack of knowledge on polychaetes in the deeper part of the Nordic seas. Of the six species of Ophelina recorded, only two may be considered as well known. Two species were recoded for the first time after their original description more than a century ago, whereas one species is described as new to science and one species appears to be new to science (not described here). Further, the results indicate that the species diversity is low in the deep basins, with only a single species of Ophelina represented in samples deeper than about 2000 m. This pattern is in accordance with results from investigations on other invertebrate taxa in the Nordic Seas (e.g. Høisæter Citation2010 and references therein).
Morphological characters in opheliids warrant a comment in relation to their usefulness in descriptions and, in particular, in comparison between related species. Tzetlin and Zhadan (Citation2009) identified four types of mouth structures divergent from the simple non-muscular axial proboscis in opheliids. One of these is a dorsal, asymmetric tongue-like proboscis with a distinct lateral groove, reported to be present in Ophelina species. All recorded species in the present study possess a similar type of proboscis. This supports the view that mouth structures are of taxonomic importance and may represent characters for phylogenetic inferences of opheliids (Tzetlin & Zhadan Citation2009).
The setae of the opheliids are relatively uniform smooth capillaries of limited taxonomical importance (Uebelacker Citation1984; Eibye-Jacobsen Citation2002). However, the recent description of Ophelina margalleffi Sarda et al., Citation2009 demonstrates the presence of hirsute capillaries in the genus Ophelina. All six species recorded in the present study possess smooth capillaries in both rami, and hirsute capillaries were not observed.
Branchiae and anal tubes are very fragile structures that are easily lost. This study illustrates the importance of examining a relatively large amount of specimens in order to correctly establish the characters of such structures, which was the case for O. abranchiata, O. cylindricaudata and O. opisthobranchiata. The study of a large material has also shown that the number of setigers is a good character for several species. Blake (Citation2000) pointed out that the number of setigers is not fixed in the type species of the genus Ophelina (Ophelina acuminata Ørsted, Citation1843), and concluded that the number of setigers is not an important character in the genus. In the present study, the number of setigers was found to be stable for all recorded species. Possibly many reports of a variable number of setigers result from a confusion of similar species, which easily may happen when branchiae and anal tubes are damaged or lost.
Acknowledgements
We are grateful to Torleiv Brattegard and Jon-Arne Sneli for making material from numerous samples from a wide range of localities in the Nordic Seas available for study. Veritas A/S and the ‘Norwegian Deepwater Programme’ are thanked for access to material from deep water monitoring off western Norway. The MAREANO project by Børge Holte and Hannu Koponen is thanked for access to material sampled off Northern Norway. We thank Elin Sigvaldadottir for loan of specimens from the collections of SMNH, and Natalia Budaeva for an English translation of the original description of Ammotrypane minuta Annenkova, Citation1952. Marc Daverdin prepared maps, and Katrine Kongshavn helped make digital photos. Drawings and preparation of the plates were made with help from Beate Helle. This study has been supported by the ‘Norwegian Deepwater Programme’ (http://www.ndwp.org/), and we are grateful to the Norwegian Academy for Sciences and Letters for putting this finance opportunity through. This is contribution #2 from the Norwegian Polychaete Forum.
References
- Annenkova , NP. 1952 . [New species of bristle-worms (Polychaeta)] . Trudy Zoologicheskogo Instituta Akademiia Nauk SSSR , 12 : 148 – 154 .
- Augener , H. 1912 . Beitrag zur Kenntnis verschiedener Anneliden un Bemerkungen über die Nephthys-Arten und deren epitoken Formen . Archiv für Naturgeschichte, Berlin , 78 : 162 – 212 .
- Augener , H. 1928 . Die Polychaeten von Spitzbergen . Fauna Arctica , 5 : 647 – 834 .
- Bakken , T , Kongsrud , JA , Oug , E , Cochrane , SKJ , Moen , TL and Solbakken , BEB. 2010 . Polychaetes from Jan Mayen (Annelida, Polychaeta) . Polar Research , 29 : 1 – 21 .
- Blake , JA. 2000 . “ Family Opheliidae Malmgren, 1867 ” . In Taxonomic atlas of the benthic fauna of the Santa Maria Basin and the Western Santa Barbara Channel , Edited by: Blake , JA and Hilbig , B . Vol. 7 , 145 – 168 . Santa Barbara, CA : Santa Barbara Museum of Natural History . The Annelida, Part 4. Polychaeta: Flabelligeridae to Sternapsidae.
- Blindheim , J and Østerhus , S. 2005 . “ The Nordic Seas, main oceanographic features ” . In The Nordic seas: An integrated perspective , Edited by: Drange , H , Dokken , T , Furevik , T , Gerdes , R and Berger , W . 11 – 37 . Washington, DC : American Geophysical Union .
- Brattegard , T and Fosså , JH. 1991 . Replicability of an epibenthic sampler . Journal of the Marine Biological Association of the U.K. , 71 : 153 – 166 .
- Ehlers , E. 1913 . Die Polychaeten-sammlungen der deutschen Südpolar-Expedition 1901–1903 . Deutsche Südpolar-Expedition , 13 : 397 – 598 .
- Eibye-Jacobsen , D. 2002 . Scalibregmatidae and Opheliidae (Annelida: Polychaeta) collected in the Andaman Sea, Thailand, during the BIOSHELF project . Phuket Marine Biological Center Special Publication , 24 : 57 – 74 .
- Eliason , A. 1962 . Die polychaeten der Skagerak-Expedition 1933 . Zoologiska Bidrag, Uppsala , 33 : 207 – 293 .
- Fauvel , P. 1911 . Annélides Polychètes. Duc d'Orleans’ Campagne Arctique de 1907 , 45 Brussels : Charles Bulens .
- Fauvel , P. 1914 . Annélides polychètes non-pélagiques provenant des campagnes de l'Hirondelle et de la Princesse-Alice (1885–1910) . Résultats des campagnes scientifiques accompliés par le Prince Albert I , 46 : 1 – 432 .
- Hansen , GA. 1879 . Annelider fra den norske Nordhavsexpedition i 1876 . Nyt Magazin for Naturvidenskaberne , 24 : 1 – 17 .
- Hansen , GA. 1882 . “ Annelida ” . In The Norwegian North-Atlantic Expedition 1876–1878. VII. Vol. 3. Zoology , Christiania : Gröndahl & Søn .
- Hartman , O. 1965 . Deep-water benthic polychaetous annelids off New England to Bermuda and other North Atlantic areas . Allan Hancock Foundation Occasional Papers , 28 : 1 – 378 .
- Hartman , O and Fauchald , K. 1971 . Deep-water benthic polychaetous annelids off New England to Bermuda and other North Atlantic Areas. Part II . Allan Hancock Monographs in Marine Biology , 6 : 1 – 327 .
- Hartmann-Schröder , G. 1974 . Polychaeten von Expeditionen der ‘Anton Dohrn’ in Nordsee und Skagerrak . Veröffentlichungen des Institut für Meeresforschung, Bremerhaven , 14 : 169 – 274 .
- Hartmann-Schröder , G. 1996 . Annelida, Borstenwürmer, Polychaeta. 2., neubearbeitete Auflage . Tierwelt Deutschlands , 58 : 1 – 648 .
- Hartmann-Schröder , G and Rosenfeldt , P. 1989 . Die Polychaeten der ‘Polarstern’-Reise ANT III/2 in die Antarktis 1984. Teil 2: Cirratulidae bis Serpulidae . Mitteilungen aus den Hamburgischen Zoologischen Museum und Institut , 86 : 65 – 106 .
- Høisæter , T. 2010 . The shell-bearing, benthic gastropods on the southern part of the continental slope off Norway . Journal of Molluscan Studies , 76 : 234 – 244 .
- Levinsen , GMR. 1887 . “ Kara-Havets Ledorme (Annulata) ” . In Djimphna-Togtets Zoologiske-botaniske Udbytte , Edited by: Lütken , CF . 288 – 303 . Köbenhaven : J. Hagerup .
- Maciolek , NJ and Blake , JA. 2006 . Opheliidae (Polychaeta) collected by the R/V Hero and the USNS Eltanin cruises from the Southern Ocean and South America . Scientia Marina , 70S3 : 101 – 113 .
- Malmgren , AJ. 1867 . Annulata polychaeta Spetsbergiæ, Grönlandiæ, Islandiæ et Scandinaviæ hactenus cognita . Öfversikt af Kongliga Vetenskaps-Akedemiens Förhandlingar , 24 : 127 – 235 .
- McIntosh , WC. 1879 . On the Annelida obtained during the cruise of H.M.S. ‘Valorous’ to Davis Strait in 1875 . Transactions of the Linnean Society of London , 1 : 499 – 511 .
- Ørsted , AS. 1843 . Annulatorum danicorum conspectus. Fasc. 1. Maricolae , 1 – 52 . Hafniae : Librariae Wahlianae .
- Sardá , R , Gil , J. , Taboada , S and Gili , JM. 2009 . Polychaete species captured in sediment traps moored in northwestern Mediterranean submarine canyons . Zoological Journal of the Linnean Society , 155 : 1 – 21 .
- Schüller , M. 2008 . New polychaete species collected during the expeditions ANDEEP I, II, and III to the deep Atlantic sector of the Southern Ocean in the austral summers 2002 and 2005 – Ampharetidae, Opheliidae and Scalibregmatidae . Zootaxa , 1705 : 51 – 68 .
- Sneli , JA. 1998 . A simple benthic sledge for shallow deep-sea sampling . Sarsia , 83 : 69 – 72 .
- Støp-Bowitz , C. 1945 . Les opheliens Norvegiens . Meddelelser fra det Zoologiske Museum, Oslo , 52 : 21 – 61 .
- Støp-Bowitz , C. 1948 . Sur les polychètes arctiques des famillies des Glycériens des Ophéliens, des Scalibregmiens et des Flabelligériens . Tromso Museums Årshefter , 66 : 1 – 58 .
- Stuart , CT and Rex , MA. 2009 . Bathymetric patterns of deep-sea gastropod species diversity in 10 basins of the Atlantic Ocean and Norwegian Sea . Marine Ecology , 30 : 164 – 180 .
- Tzetlin , A and Zhadan , A. 2009 . Morphological variation of axial non-muscular proboscis types in the Polychaeta . Zoosymposia , 2 : 415 – 427 .
- Uebelacker , JM. 1984 . “ Chapter 17: Family Opheliidae Malmgren, 1867b ” . In Taxonomic guide to the Polychaetes of the Northern Gulf of Mexico 5 , Edited by: Uebelacker , JM and Johnson , PG . 1 – 15 . Metairie, LA : Barry, A. Vittor and Associates, Inc .
- Wirén , A. 1883 . “ Chaetopoder från Sibiriska Ishafvet och Berings Haf Insamlade under Vega-Expeditionen 1878–1879 ” . In Vega-Expeditionens Vetenskapliga Iakttagelser (bearbetade af deltagare i resan och andra forskare) , Edited by: Nordenskiöld , NAE . Vol. 2 , 383 – 426 . Stockholm : F&G Beijers Förlag .
- Wirén , A. 1901 . Ueber die waehrend der schwedischen arktischen Expedition von 1898 und 1900 eingesammelten Anneliden . Zoologischer Anzeiger , 24 : 253
- Wesenberg-Lund , E. 1950 . Polychæta . The Danish Ingolf-Expedition , 14 : 1 – 92 .
- Wesenberg-Lund , E. 1953 . The zoology of East Greenland . Polychaeta. Meddelelser om Grønland , 122 : 1 – 170 .