Abstract
A quantitative study on the sponge spicules present in a series of horizontal core samples, taken at different levels, from a 1.5 m thick living matte within the Posidonia oceanica meadow of Prelo Bay (Eastern Ligurian Sea), has been performed. Sponge spicule amounts were evaluated by microscopical and chemical analyses. From the superficial samples the spicule number decreased until 60–80 cm depth to increase progressively until the deepest studied layer, 140 cm depth. The same results were obtained from the trend of spicule volume and biogenic silica tested by chemical analysis. Among the recognisable spicule types, the most common are oxeas, followed by tylostyles, achantostyles and strongyles. However, oxeas, tylostyles and achantostyles showed the same trend of the entire spicule population, whereas strongyles had an opposite trend with a maximum value at the 80–100 cm depth. According to the hypothesis that the amount of spicules in the sediment is proportional or at least related to the sponge biomass existing in the surrounding area, our data suggest that the sponge assemblage of Prelo Bay should not have been constant in the past 100–200 years. Our hypothesis is that the rapid urban and industrial development that occurred along the Ligurian coast after the Second World War, particularly in the 1960s, had an important impact on the sponge assemblages of the area.
Introduction
Mediterranean marine communities have shown, during the last decades, significant changes in composition and distribution. These modifications are considered directly or indirectly related to the current climatic changes that may influence the spreading capacity of a certain species, therefore modifying its range (Astraldi et al. Citation1995; Morri & Bianchi Citation2001; Bianchi & Morri Citation2004). Moreover, a simplification of the superficial benthic communities has occurred, mainly in consequence of a sharp reduction in structuring organisms (above all sponges, bryozoans and gorgonians) after a number of mass mortality events that took place from 1999 to 2006 (Coma et al. Citation2009). The need to reconstruct past diversity to understand ecosystem change has been stressed by Boero and Bonsdorff (Citation2007), who proposed a Historical Biodiversity Index (HBI).
From a theoretical and practical point of view, it is necessary to state whether these changes are casual and unforeseeable or whether they may be signals of a directional trend, driven by global warming. The interpretation of changes is difficult, especially in the coastal environment where different anthropic disturbances may merge with warming effects and the absence of historical quantitative data on the composition of benthic communities is almost complete. The scant available information is limited to the last two to three decades (Bianchi & Morri Citation2000; Bianchi Citation2007).
The latter problem can be faced through quantitative studies on the remains (e.g. mineral skeletal structures) that benthic organisms leave in the sediment after death. Sponge spicules represent, on this subject, a potential good proxy for the evaluation of long-term sponge community dynamics. In Mediterranean coastal assemblages, sponges are quite abundant. In the last 30 years they have suffered from severe mass mortalities generally attributed to water warming and/or pathogen infections (Gaino et al. Citation1992; Cerrano et al. Citation2000; Coma et al. Citation2009; Garrabou et al. Citation2009). Sponge siliceous spicules resist dissolution and are able to remain in sediments for a long span of time (Rützler & Macintyre Citation1978; Bavestrello et al. Citation1996; Maldonado et al. Citation2005). Nevertheless, the superficial portion of sediment is subjected to a continuous re-suspension (Bavestrello et al. Citation1996) that, together with the known intense bioturbation (Gerino Citation1990), increases the uncertainty of a correct temporal interpretation of data.
A possible remedy for this problem is to use the sediments entrapped into the sea grass meadows and particularly those of Posidonia oceanica. The reduction of water movement by Posidonia leaves reduces resuspension processes in the meadows (Fonseca & Fisher Citation1986; Fonseca Citation1989), entrapping suspended materials (Short & Short Citation1984) and reducing, by friction, wave energy (Fonseca & Cahalan Citation1992). The decreasing of resuspension in Posidonia meadows, experimentally demonstrated by Garcia and Duarte (Citation2001) and Terrados and Duarte (Citation2000) suggests that the sediment settled in these habitats could reflect rather precisely the temporal sequence of its deposition. The Posidonia plant faces sedimentation by a continuous vertical growth of its rhizome (orthotropic growth), with an average rate comprised between 4 and 9 mm·year−1 in undisturbed sites and between 2 and 4 mm·year−1 in polluted ones (Marbà et al. Citation2006).
Therefore, Posidonia meadows represent a sort of natural sediment trap and the vertically growing matte is a reservoir of recent sediment, whose age can be grossly determined.
In this work we have conducted a quantitative study on the sponge spicules present in core samples taken from a 1.5-m thick living matte of Posidonia at Prelo Bay (Eastern Ligurian Sea). All spicule fragments present in a standard amount of sediment, at different levels of the matter, were counted, measured and, when possible, attributed to a specific spicule type. The amount of spicules at the different levels of the core sample was assumed as directly related to the sponge abundance in the surrounding area of the meadow during the deposition period.
Materials and methods
The Prelo Bay, situated on the western side of the Tigullio Gulf (Ligurian Sea), is a small cove facing east, characterised by a sandy bottom with a maximal depth of 15 m. A large and dense meadow of Posidonia oceanica thrives on this bottom, extending continuously from the shore to 7 m depth. From 7 to 15 m depth, the sea grass is present with scattered tufts.
The occurrence of this meadow was previously reported by Issel (Citation1912), but the first bibliographic report is due to Tortonese (Citation1962), who described the meadow as ‘very dense’. Bavestrello (Citation1983) and Balduzzi et al. (Citation1984) provided new and more detailed information, but the first map of the meadow was provided later by Bianchi and Peirano (Citation1995), who also discussed all the relevant bibliographic information to the early 1990s. A more recent and detailed map has been published by Montefalcone et al. (Citation2006). Other recent studies on this meadow include Covazzi Harriague et al. (Citation2006), Giovannetti et al. (Citation2006, Citation2008), Lasagna et al. (Citation2006b) and Montefalcone et al. (Citation2008).
The macrozoobenthic assemblage of this meadow was deeply studied by semi-quantitative methods particularly as to hydroids (Boero Citation1981) and sponges (Pansini & Pronzato Citation1985).
On the southern side of the bay the meadow thrives on an exposed matte, 1.5-m thick that is the result of a cutting done in the 1930s to build a jetty for the access of small boats (Lasagna et al. Citation2006a).
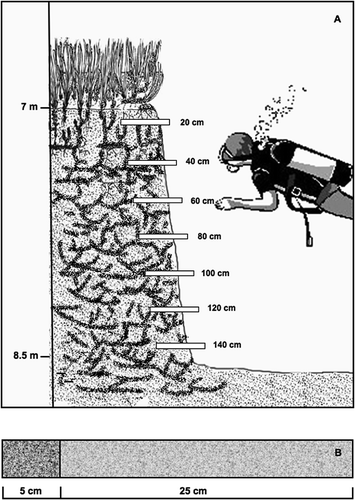
To obtain sediment along the whole thickness of the exposed matte, in July 2009 we collected 7 core samples, 30 cm long and 3 cm in diameter, perpendicular to the exposed matte, regularly spaced from 20 cm under the surface to the bottom (). The first 5 cm of the core (corresponding to the inner part of the matte) were subsampled, stored at –20°C and then dried at 80°C over 24 h.
Figure 1. A, Schematic drawing of the core sample collection in the exposed matte of the Posidonia oceanica meadow of the Prelo Bay (Ligurian Sea). B, Scheme of a core sample; only the first 5 cm of the sample were used to collect data about spicule concentration.
The sediment from each one of these portions was sieved through a mesh of 250 μm to separate Posidonia fibres; then 0.1 g of sediment (in triplicate) was treated by boiling nitric acid to eliminate the carbonate fraction. The resulting fraction has been distributed on three permanent microscope slides for counting and identifying sponge spicule types. The volume of each spicule (or spicule fragment) was obtained from its dimensions. Data were reported to 1 g of original sediment (without Posidonia fibres). Finally, the vertical distribution of the quantity and diversity of the spicule types along the core samples was obtained.
Biogenic silica percentage in the original sediment was chemically measured following DeMaster (Citation1981) and Mortlock and Froelich (Citation1989) procedures. Five millilitres of a solution of H2O2 (10%) were added to approximately 100–200 mg of dry sediment and, after 30 min, 5 ml of HCl (10%) were added to the sediment and the solution of H2O2 (10%). Samples were rinsed with bi-distilled water and after centrifugation (5 min at 5000 rpm) were dried in the oven before alkaline extraction.
The samples (100 mg in triplicate) were resupended in 50 ml of a solution NaOH (1 M) and were immersed in a water bath heated to 100°C for 5 h and periodically shaken throughout the alkaline extraction period (Klein Citation2008). After extraction of biogenic silica from sponge spicules, the samples were centrifuged for 10 min at 4000 rpm and the supernatant was used for the determination of dissolved biogenic silica. The concentration of dissolved silica was measured with the molybdate blue spectrophotometric method at 810 nm (Grasshoff et al. Citation1983). A linear regression was plotted running through the two values corresponding to 2 and 5 h of extraction. The y-intercept of the regression line corresponded to the concentration of biogenic silica in the sample (DeMaster Citation1981).
Results
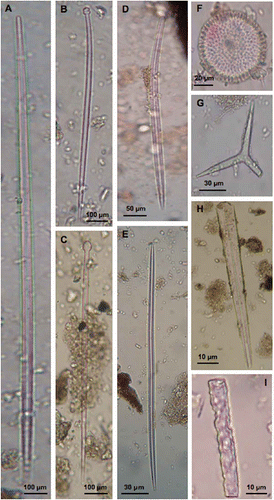
At each level of the core sample sponge spicules or fragments were abundant, generally in a good state of preservation (,G), sometimes showing typical circular erosions (,I). Some spicules could be attributed to the species recorded in the Prelo meadow by Pansini and Pronzato (Citation1985) as strongyloxeas of Tethya citrina Sarà & Melone, 1965 () and tylostyles of Cliona viridis (Schmidt, 1862) ().
Figure 2. A–G, Examples of sponge spicules and H,I, spicule fragments recorded in the sediment of the Posidonia matte. Some spicules can be easily recognised as A, strongyloxea of Tethya citrina or B, tylostyles of Cliona viridis. H,I, some fragments show deep marks of dissolution.
The total number of spicules (fragments + entire spicules) per 1 g of sediment showed significant variations inside the matte. In the most superficial sample the average number of spicules was 4240 ± 800 spicules × g−1 of sediment. This value increased in the 40 cm layer (7040 ± 680 spicules × g−1 of sediment), while it strongly decreased in the two following layers (60 cm, 2200 ± 440 and 80 cm 2360 ± 360 spicules·g−1 of sediment, respectively). In the deeper layers, the spicule number increased again, reaching the value of 11,120 ± 940 spicules·g−1 of sediment at 140 cm ().
Figure 3. A, Vertical trends of concentration per g of sediment of total spicule fragments; B, entire spicules; C, total volume of sponge silica; and D, biogenic silica chemically tested.
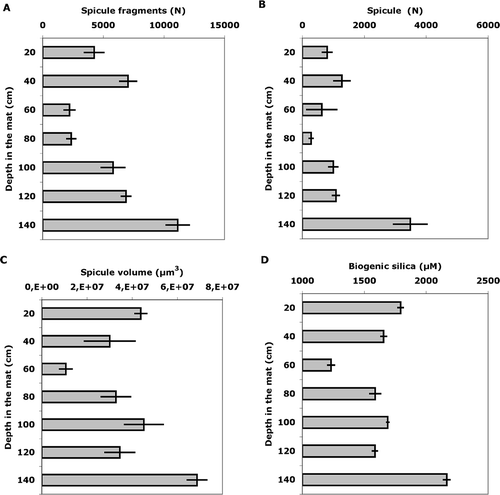
The same trend was obtained by considering only the unbroken spicules, representing from 12% (80 cm) to 31% (140 cm) of the recorded spicular remains ().
The total estimated volume of the recorded spicules showed a decreasing trend from the surface of the matte (about 40 millions of μm3·g−1 of sediment) to 40–60 cm depth (about 10 millions μm3·g−1 of sediment), increasing again towards the deepest part of the sampled matte (about 70 millions of μm3·g−1 of sediment) ().
The volume trend was in agreement with the chemical assessment of the biogenic silica, decreasing by about one-third from the surface (1793 μM·g−1 of sediment) to 60 cm (1230 μM·g−1 of sediment) depth and increasing in the following layers, reaching its maximum (2160 μM·g−1 of sediment) in the deepest layers of analysed sediment ().
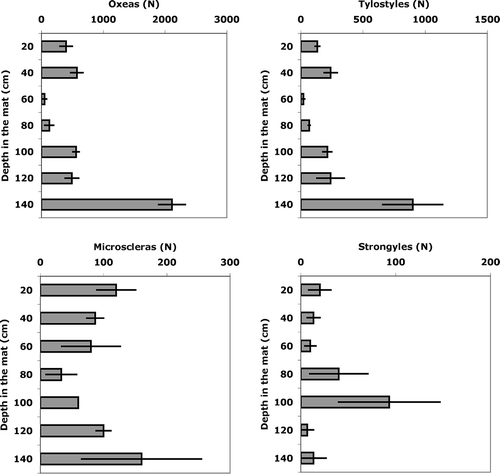
Oxea was the commonest recognisable spicule type, accounting for 55% of the total observed entire spicules. The oxea density showed a trend inside the matte in accordance with the total number of spicules: only at 60–80 cm depth did the number of oxeas significantly decrease (). The same trend was recorded for tylostyles, accounting for the 23% of the total number of spicules; achantostyles, accounting for 8.5%; styles (3.5%); and microscleres (8.1%) (). On the contrary, strongyles, which comprised 2.5% of the total, showed an opposite trend with a maximal density in the 80–100 cm depth layer in the matte.
Figure 4. Vertical trends of concentration per g of sediment of different spicule types. A, oxeas; B, tylostyles; C, microscleres, D, strongyles.
Discussion
Sponge spicules enrich the sediment fraction of biogenic silica when dead animals decay (Rützler & Macintyre Citation1978). Compared to diatom and radiolarian tests, sponge spicules seem to take longer to dissolve, particularly in environments characterised by low pH conditions (Kamatani Citation1971; Conley & Schelske Citation1993; Maldonado et al. Citation2005). In some geographic areas, e.g. Antarctica, they can form 1.5-m thick mats (Koltun Citation1968; Dayton Citation1979; Barthel Citation1992; Cattaneo-Vietti et al. Citation2000).
Type and abundance of sponge spicules in dated sediment core samples were already used to reconstruct the dynamics of freshwater sponge fauna in lakes or lagoons (Paduano & Fell Citation1997; Volkmer-Ribeiro et al. Citation2007). Bavestrello et al. (Citation1996) evaluated the contribution of this form of low-soluble silica to the bottom sediments of different areas of the Ligurian Sea. Microscopical analysis of the fine fraction of the sediments coming from several stations of the Ligurian Sea demonstrated that sponge spicules represent nearly the total BSi, while other siliceous remains, such as diatom and silicoflagellate tests, constitute a negligible fraction. Their amount was variable according to the abundance of sponges in the surrounding communities from about 17,000 to 1000 spicules·g−1 of sediments. These data are in agreement with those coming from the core taken in the Prelo Bay Posidonia meadow ranging from about 11,000 to about 2000 spicules·g−1 of sediment.
In the study by Bavestrello et al. (Citation1996), conducted on detritic bottoms at the base of rocky cliffs inhabited by sponges, far from Posidonia oceanica meadows, spicule concentration continuously decreases from the sediment surface to 80–100 cm depth (the maximal depth examined by core samples), reaching values ranging from 50 to 10% of those recorded at the surface. On the contrary, in the Posidonia meadow, the amount decreases until 40–60 cm to increase in the deeper levels reaching the maximal values at 140 cm depth.
Both in the Ligurian Sea (Bavestrello et al. Citation1996) and in other localities (Inoue Citation1985) it was suggested that the spicules in the sediments are supplied by local sponge fauna as shown by the comparison between recorded spicules in the sediment and sponge species collected in the surroundings. So it is most probable that the spicules recorded in the Posidonia meadow of Prelo Bay derive, in large part, from the rich and diversified sponges recorded on the rhizomes and formerly studied by Pansini and Pronzato (Citation1985).
The assumption that the P. oceanica matte has a regular growth rate evaluated at 4–9 mm·y−1 (Marbà et al. Citation2006) allows a temporal scale to be given to the observed variations. Of course, a more detailed study extended over a larger area would be necessary to obtain more certain results, since our data depict only a possible scenario of the status of the sponge fauna associated to the Posidonia meadow of the Prelo Bay in the last 350–150 years. Particularly evident is the strong decrease of spicules at 40–60 cm depth, corresponding to a time span ranging from 45 to 65 years ago, assuming that the growth rate of the matte is 9 mm·y−1.
This severe decrease of sponge spicules could be tentatively interpreted as a consequence of the human impact during the period of rapid urban and industrial development occurred along the Ligurian coast after the Second World War, which had its maximal impact in the 1960s in the area of the Tigullio Gulf. During this period, for instance, nearly 30% of the original meadow surface of the area disappeared, according to Bianchi and Morri (Citation2000).
Although these results are preliminary, they indicate that sediments entrapped among Posidonia oceanica rhizomes can be a powerful proxy to reconstruct the abundance and diversity of sponge assemblages during a century-long time span.
References
- Astraldi , M , Bianchi , CN , Gasparini , GP and Morri , C. 1995 . Climatic fluctuations, current variability and marine species distribution: A case study in the Ligurian Sea (north-west Mediterranean) . Oceanologica Acta , 18 : 139 – 149 .
- Balduzzi , A , Bavestrello , G , Belloni , S , Boero , F , Cattaneo-Vietti , R , Pansini , M and Pronzato , R. 1984 . “ Valutazione dello stato di alcune praterie di Posidonia del Mar Ligure sottoposte a diverse condizioni di inquinamento ” . In International Workshop on Posidonia oceanica beds , Edited by: Boudouresque , CF , de Grissac , AJ and Olivier , J . 73 – 78 . Marseille : G.I.S. Posidonie publications .
- Barthel , D. 1992 . Do hexactinellids structure Antarctic sponge associations? . Ophelia , 36 : 111 – 118 .
- Bavestrello , G. 1983 . Nuove osservazioni sulle praterie di Posidonia oceanica (L.) Delile del versante occidentale del Golfo Marconi (Riviera Ligure di Levante) . Bollettino dei Musei e degli Istituti Biologici dell'Università di Genova , 50–51 : 99 – 104 .
- Bavestrello , G , Cattaneo-Vietti , R , Cerrano , C and Sarà , M. 1996 . Contribution of sponge spicules to the composition of biogenic silica both in the water column and in the bottom sediments in the Ligurian Sea. P.S.Z.N.I . Marine Ecology , 17 : 41 – 50 .
- Bianchi , CN . 2007 . Biodiversity issues for the forthcoming tropical Mediterranean Sea . Developments in Hydrobiology , 193 : 7 – 21 .
- Bianchi , CN and Morri , C. 2000 . Marine biodiversity of the Mediterranean Sea: Situation, problems and perspectives for future researches . Marine Pollution Bulletin , 40 : 367 – 376 .
- Bianchi , CN and Morri , C. 2004 . Climate change and biological response in Mediterranean Sea ecosystems – A need for broad-scale and long-term research . Ocean Challenge , 13 : 32 – 36 .
- Bianchi , CN and Peirano , A. 1995 . Atlante delle fanerogame marine della Liguria: Posidonia oceanica e Cymodocea nodosa . ENEA, Centro Ricerche Ambiente Marino, La Spezia , : 1 – 146 . 44 pls
- Boero , F. 1981 . Systematics and ecology of the Hydroid population of two Posidonia oceanica meadows. P.S.Z.N.I . Marine Ecology , 2 : 181 – 197 .
- Boero , F and Bonsdorff , E. 2007 . A conceptual framework for marine biodiversity and ecosystem functioning . Marine Ecology , 28 ( Suppl. 1 ) : 134 – 145 .
- Cattaneo-Vietti , R , Bavestrello , G , Cerrano , C , Pansini , M , Mazzella , L and Sarà , M. 2000 . “ The role of sponges of Terra Nova Bay ecosystem ” . In Benthic ecology of Terra Nova Bay , Edited by: Faranda , F . 539 – 549 . Springer-Verlag, Berlin, Heidelberg, New York .
- Cerrano , C , Bavestrello , G , Bianchi , CN , Cattaneo-Vietti , R , Bava , S , Morganti , C , Morri , C , Picco , P , Sara , G Schiaparelli , S . 2000 . A catastrophic mass-mortality episode of gorgonians and other organisms in the Ligurian Sea (NW Mediterranean), summer 1999 . Ecology Letters , 3 : 284 – 293 .
- Coma , R , Ribes , M , Serrano , E , Jimeneza , E , Salat , J and Pascual , J. 2009 . Global warming-enhanced stratification and mass mortality events in the Mediterranean . Proceedings of the National Academy of Sciences , 106 : 6176 – 6181 .
- Conley , DJ and Schelske , CL. 1993 . Potential role of sponge spicules in influencing the silicon biogeochemistry of Florida lakes . Canadian Journal of Fisheries and Aquatic Sciences , 50 : 296 – 302 .
- Covazzi Harriague , A , Bianchi , CN and Albertelli , G. 2006 . Soft-bottom macrobenthic community composition and biomass in a Posidonia oceanica meadow in the Ligurian Sea (NW Mediterranean) . Estuarine, Coastal and Shelf Science , 70 : 251 – 258 .
- Dayton , PK. 1979 . “ Observations of growth, dispersal and population dynamics of some sponges in McMurdo Sound, Antarctica ” . In Colloques internationaux du C.N.R.S. 291. Biologie des spongiaires , Edited by: Lévi , C and Boury-Esnault , N . 271 – 282 . Paris : Editions du Centre National de la Recherche Scientifique .
- DeMaster , DJ. 1981 . The supply and accumulation of silica in the marine environment . Geochimica et Cosmochimica Acta , 45 : 1715 – 1732 .
- Fonseca , MS. 1989 . Sediment stabilization by Halophila decipiens in comparison to other seagrasses . Estuarine, Coastal and Shelf Science , 29 : 501 – 507 .
- Fonseca , MS and Cahalan , JA. 1992 . A preliminary evaluation of wave attenuation by four species of seagrass . Estuarine, Coastal and Shelf Science , 35 : 565 – 576 .
- Fonseca , MS and Fisher , JS. 1986 . A comparison of canopy friction and sediment movement between four species of seagrass with reference to their ecology and restoration . Marine Ecology Progres Series , 29 : 15 – 22 .
- Gaino , E , Pronzato , R , Corriero , G and Buffa , P. 1992 . Mortality of commercial sponges: Incidence in two Mediterranean areas . Bollettino di Zoologia , 59 : 79 – 85 .
- Garcia , E and Duarte , CM. 2001 . Sediment retention by a Mediterranean Posidonia oceanica meadow: The balance between deposition and resuspension . Estuarine, Coastal and Shelf Science , 52 : 505 – 514 .
- Garrabou , J , Coma , R , Bensoussan , N , Bally , M , Chevaldonné , P , Cigliano , M , Diaz , D , Harmelin , JG , Gambi , MC Kersting , DK . 2009 . Mass mortality in Northwestern Mediterranean rocky benthic communities: Effects of the 2003 heat wave . Global Change Biology , 15 : 1090 – 1103 .
- Gerino , M. 1990 . The effects of bioturbation on particle redistribution in Mediterranean coastal sediment. Preliminary results . Hydrobiologia , 207 : 251 – 258 .
- Giovannetti , E , Montefalcone , M , Bianchi , CN , Morri , C and Albertelli , G. 2006 . Structural variability of the epiphytic community in a Posidonia oceanica meadow (Ligurian Sea, NW Mediterranean) . Biologia Marina Mediterranea , 13 : 145 – 148 .
- Giovannetti , E , Montefalcone , M , Morri , C , Bianchi , CN and Albertelli , G. 2008 . Biomassa fogliare ed epifita in una prateria di Posidonia oceanica (Prelo, Mar Ligure): Possibilità di determinazione tramite un metodo indiretto . Atti dell'Associazione Italiana di Oceanologia e Limnologia , 19 : 229 – 233 .
- Grasshoff , K , Ehrhardt , M and Kremling , K. 1983 . Methods of seawater analysis , 2nd , 419 Weinheim : Verlag Chemie .
- Inoue , M. 1985 . An examination of supply processes of sponge spicules to the sediment of the northeastern to eastern part of Sagami Bay . Annual Reports of the Faculty of Education, Iwate University , 44 : 61 – 80 .
- Issel , R. 1912 . Biologia neritica mediterranea. Il benthos animale delle foglie di Posidonia studiato dal punto di vista bionomico . Zoologische Jahrbuecher Systematik , 33 : 379 – 420 .
- Kamatani , A. 1971 . Physical and chemical characteristics of biogenous silica . Marine Biology , 8 : 89 – 95 .
- Klein , AW. 2008 . Analysis of biogenic silica from marine sediment in Maxwell Bay, Antarctica: 258 A look at Holocene climate history , PhD Thesis, Middlebury College .
- Koltun , VM. 1968 . “ Spicules of sponges as an element of the bottom sediments of the Antarctic ” . In SCAR Symposium on Antarctic Ocean , 121 – 123 . Cambridge : Scott Polar Research Institute .
- Lasagna , R , Montefalcone , M , Bianchi , CN , Morri , C and Albertelli , G. 2006a . Morphology of a Posidonia oceanica meadow under altered sedimentary budget . Biologia Marina Mediterranea , 13 : 245 – 249 .
- Lasagna , R , Montefalcone , M , Bianchi , CN , Morri , C and Albertelli , G. 2006b . Approccio macrostrutturale alla valutazione dello stato di salute di una prateria di Posidonia oceanica . Biologia Marina Mediterranea , 13 : 379 – 385 .
- Maldonado , M , Carmona , MC , Velásquez , Z , Puig , A , Cruzado , A , López , A and Young , CM. 2005 . Siliceous sponges as a silicon sink: An overlooked aspect of benthopelagic coupling in the marine silicon cycle . Limnology and Oceanography , 50 : 799 – 809 .
- Marbà , N , Santiago , R , Díaz-Almela , E , Álvarez , E and Duarte , CM. 2006 . Seagrass (Posidonia oceanica) vertical growth as an early indicator of fish farm-derived stress . Estuarine, Coastal and Shelf Science , 67 : 475 – 483 .
- Montefalcone , M , Chiantore , M , Lanzone , A , Morri , C , Albertelli , G and Bianchi , CN. 2008 . Beyond BACI design reveals the decline of the seagrass Posidonia oceanica induced by anchoring . Marine Pollution Bulletin , 56 : 1637 – 1645 .
- Montefalcone , M , Lasagna , R , Bianchi , CN , Morri , C and Albertelli , G. 2006 . Anchoring damage on Posidonia oceanica meadow cover: A case study in Prelo cove (Ligurian Sea, NW Mediterranean) . Chemistry and Ecology , 22 : 207 – 217 .
- Morri , C and Bianchi , CN. 2001 . “ Recent changes in biodiversity in the Ligurian Sea (NW Mediterranean): Is there a climatic forcing? ” . In Structure and processes in the Mediterranean ecosystems , Edited by: Faranda , FM , Guglielmo , L and Spezie , G . 375 – 384 . Milano : Springer Verlag .
- Mortlock , RA and Froelich , PN. 1989 . A simple method for the rapid determination of biogenic opal in pelagic marine sediments . Deep Sea Research Part A , 36 : 1415 – 1426 .
- Paduano , GM and Fell , PE. 1997 . Spatial and temporal distribution of freshwater sponges in Connecticut lakes based upon analysis of siliceous spicules in dated sediment cores . Hydrobiologia , 350 : 105 – 121 .
- Pansini , M and Pronzato , R. 1985 . Distribution and ecology of epiphytic Porifera in two Posidonia oceanica (L.) Delile meadows of the Ligurian sea. P.S.Z.N.I . Marine Ecology , 6 : 1 – 11 .
- Rützler , K and Macintyre , IG. 1978 . Siliceous sponge spicules in coral reef sediments . Marine Biology , 49 : 147 – 159 .
- Short , FT and Short , CA. 1984 . “ The seagrass filter: Purification of estuarine and coastal waters ” . In The estuary as a filter , Edited by: Kennedy , VS . 395 – 413 . New York , NY : Academic Press .
- Terrados , J and Duarte , CM. 2000 . Experimental evidence of reduced particle resuspension within a seagrass (Posidonia oceanica L.) meadow . Journal of Experimental Marine Biology and Ecology , 243 : 45 – 53 .
- Tortonese , E. 1962 . Recenti ricerche in ambienti litorali del mare Ligure . Pubblicazioni della Stazione Zoologica di Napoli , 32 ( suppl ) : 99 – 116 .
- Volkmer-Ribeiro , C , Ezcurra de Drago , I and Parolin , M. 2007 . Spicules of the freshwater sponge Ephydatia facunda indicate lagoonal paleoenvironment at the Pampas of Buenos Aires Province, Argentina . Journal of Coastal Research , 50 : 449 – 452 .