Abstract
The morphology of about half of the valid ampharetid species (25% including types) was examined. The usefulness of morphological characters, including traditional ones and newly proposed ones, for generic diagnoses are discussed in detail. If possible, intraspecific and interspecific variation in external morphological characters, both traditional and newly proposed were examined. Some of the characters that have traditionally been used for generic identification are considered useful at a specific level. This results in a fundamental change of generic diagnoses and a considerable reduction of the number of genera from 90 described to 24 (some genera have uncertain taxonomical status). New diagnoses for all valid genera, a list of genera, a synopsis of proposed generic synonymies and an identification key for genera are provided.
Introduction
Ampharetids have a worldwide marine distribution from the intertidal to 8292 m deep (personal observation). A few (two or three) species are restricted to freshwater systems. Almost 100 genera of Ampharetidae have been described for less than 300 species, although the overview by Reuscher et al. (Citation2009) regarded only about 70 valid. Of the genera described to date, 37 genera are monotypic (Holthe Citation2002), suggesting that ampharetid generic diagnoses are very narrow. Traditionally, since Malmgren (Citation1866) ampharetid taxonomy has been based on meristic and qualitative characters including the number of thoracic segments, the number of branchiae and the presence/absence of paleae. The present classification with its multitude of monotypic genera does not represent a natural system. I aim to re-define generic diagnoses and modify the ampharetid classification so that it reflects the phylogenetic relationships more realistically.
Material and methods
For the present study almost half of the known ampharetid species and more than 20 species yet to be described have been examined from the following collections:
| • | Department of Hydrobiology, Moscow State University, Russia; | ||||
| • | Forschungsinstitut Senckenberg, Abteilung Deutsches Zentrum für Marine Biodiversitätsforschung, Germany; | ||||
| • | Marine Ecological Survey Ltd, Bath, UK; | ||||
| • | N.N. Severtsov Institute of Ecology and Evolution of the Russian Academy of Sciences, Russia; | ||||
| • | Natural History Museum, London, UK; | ||||
| • | P.P. Shirshov Institute of Oceanology of the Russian Academy of Sciences, Russia; | ||||
| • | Thomson Unicomarine Ltd, UK; | ||||
| • | Zoological Institute of Russian Academy of Sciences, St. Petersburg, Russia; | ||||
| • | Zoological Museum of Moscow State University, Russia; and | ||||
| • | Zoologisches Institut und Zoologisches Museum, Universität Hamburg, Germany. | ||||
The total number of specimens examined is in the tens of thousands. The total number of investigated species is about 150 (73 of these were type material), 129 are listed in Table II; here I only mention new species that are described in papers that are already or almost in press. If possible, intraspecific and interspecific variation in external morphological characters, traditional and newly proposed morphological characters were examined.
Specimens were studied with a stereomicroscope, generally stained with methylene blue in water, which helps observation of certain morphological characters and enabled one to find some previously unrecognized characters. The precise concentration does not matter; it should colour the specimen blue (not too dark!) in a few seconds. After staining, specimens were rinsed in order to remove excess dye. Investigation was made in water as alcohol immediately dissolves the staining pattern. A few days after returning the specimens to alcohol the methyl blue dissolves, leaving the specimens in their initial condition. The chaetae of 46 species were examined using scannning electron microscopy (SEM).
The following abbreviations are used:
| • | AU, abdominal unciniger; | ||||
| • | TC, thoracic chaetiger; | ||||
| • | TU, thoracic unciniger. | ||||
Abbreviations with a number refer to an actual segment, i. e. AU2 means the second abdominal segment.
Characters of Ampharetidae and their value for taxonomy
I have previously reviewed the taxonomic value of some characters for the classification of Ampharetidae (Jirkov Citation2001, Citation2009). Additional examination has revealed new insights into previously discussed characters and new characters of taxonomic interest.
Prostomium
The most common types of prostomium were described earlier (Jirkov Citation2001, 2009). Additional types can be distinguished ().
Figure 1. Main types of prostomia.
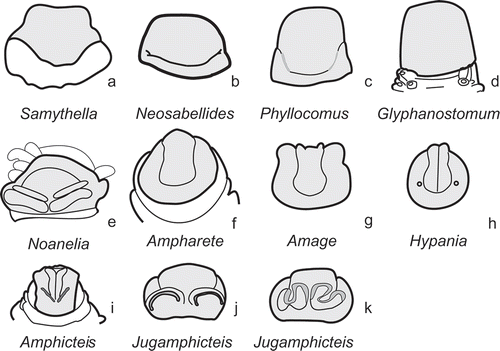
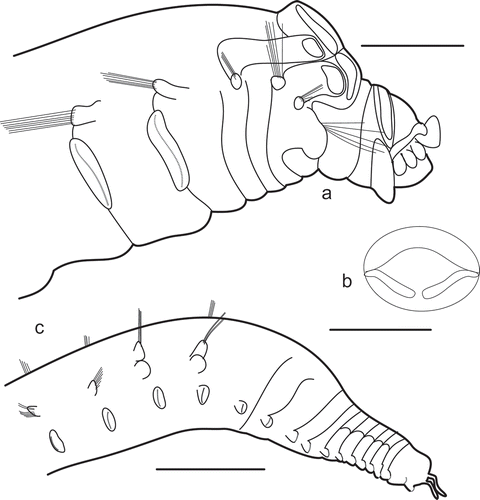
Noanelia-type (, ). The tentacle membrane of this genus is unable retract into the mouth cavity, as with Terebellidae (). The anterior prostomium has a transverse ridge caused by the attachment of tentacles. Such a ridge has not been described in any other ampharetid including specimens with extended tentacles. The prostomium is not separated into parts. A pair of straight or slightly arched anteriorly wide low ridges is present. They range from the median prostomium to the lateral margins without medial gap between them. There are yellow (in fixed specimens) nuchal (?) organs along anterior margins of these ridges. This type of prostomium is known from Noanelia only (see also remarks for the generic diagnosis).
Figure 2. Noanelia hartmanae. a, Lateral view; b, entire prostomium, dorsal view; c, posterior thorax and abdomen, lateral view. Scales: a, 0.5 mm; b, 0.25 mm; c, 1 mm.
Three more types of prostomium can be distinguished: a Neosabellides-type, a Phyllocomus-type, and a Glyphanostomum-type (). For their descriptions, see generic diagnosis below.
Segmentation
Most likely, the uncini of Ampharetidae start on the same segment throughout the family. Some authors argue that the paleae are located in the second segment and uncini start in the sixth segment (Orrhage Citation2001; Reuscher et al. Citation2009), while others suggest third and seventh segment, respectively (Malmgren, 1865; Fauvel, Citation1927). However, for the delimitation of genera, the number of the first uncinigerous segment does not matter if a clear statement is provided to prevent confusion. Herein paleae are considered notochaetae situated in the 2nd segment, irrespective of size.
Recently, Reuscher et al. (Citation2009) suggest counting thoracic chaetigers including the paleal chaetiger (in Terebellomorpha the most commonly accepted definition of thoracic segments is any segments with notochaetae). Some species have one or two achetous segment next to the paleal segment, so it is not possible to count them as chaetigers or even as thoracic segments. In my opinion, the paleal segment and segments with notochaetae need to be counted separately in order to avoid confusion.
Buccal tentacles
The diagnosis of Ampharetidae includes the statement that buccal tentacles are attached inside the mouth (Hilbig Citation2000; Jirkov Citation2001). However, in Noanelia hartmanae and Noanelia sp. nov., the buccal tentacles are attached outside the mouth (). Probably the same situation is in Andamanella Holthe, Citation2002, but the nature of the last taxon is not clear from the original description. Examined types of Glyphanostomum holthei have a ring of papillae in addition to normal buccal tentacles. The variation of the tentacles in Polycirrinae is high; in different species they are attached differently, sometimes they are distinctly different in size, some have a groove, other have not. I suggest that the papillae and tentacles of G. holthei as well as tentacles of Terebellidae are probably homologous.
Jaws
Jaws have been described for three ampharetid species: Gnathampharete paradoxa Desbruyeres, Citation1978; Adercodon pleijeli Mackie, Citation1994, and Ampharete sp. B Uebelacker, Citation1984. All known species share other characters (Ampharete-type prostomium; neuropodia of the first two abdominal chaetigers are of thoracic type, a pair of nephridial papillae behind the branchiae). These characters they share with species of Ampharete sensu lato, so the presence of jaws can be treated as synapomorphy and new species with jaws can be found within previously described species of Ampharete sensu lato. The presence of jaws might be a valuable character for generic diagnoses. However, jaws can be observed only after dissection, so for most species we do not know if they have jaws.
Lower lip
The lower lip of Ampharetidae is usually similar. However, in Lysippe, Samytha, Pterolysippe and Lysippides it is enlarged with numerous longitudinal grooves and extends laterally.
‘Horns’
Here the first segment (the segment preceding the paleal segment) has large (of notopodium size) papillae, the methylene blue staining pattern is dark blue, much darker than the surrounding body. This structure is only known from a recently found undescribed genus.
Modified notopodia
Two new types of modification can be added to previously described types.
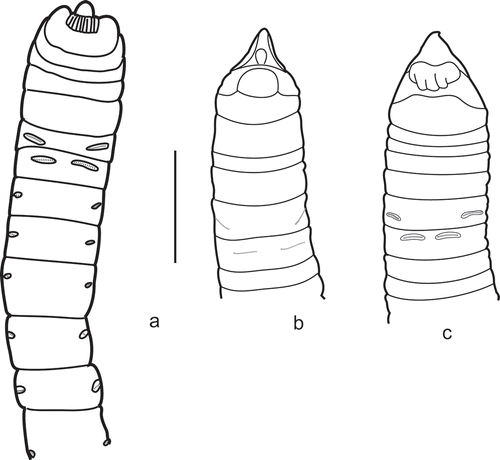
A Noanelia-type (), where there are large papillae between the notopodia and the neuropodia of last two TU in small (5–6 mm long) specimens. These papillae are similar in size to the notopodia. In larger specimens papillae are fused to the notopodia, so these notopodia become larger than others. In all specimens notopodia of last TU are shifted, especially on the 2nd from last TU. However, there are no other differences between the two last TU and other segments (dorsal ridges, bands, etc.). The last two pairs of notopodia are slightly enlarged, but do not differ in shape.
An Auchenoplax-type, where the thorax is sharply subdivided into two regions: (1) anterior 9–10 TS short with well-developed ventral glandular pads, occupying all of the ventral surface between the notopodia of anterior TS; (2) last 5 TU very long, glandular pads not developed. There are no transitional segments. Notopodia of the anterior region are cylindrical, those of the posterior slightly flattened. Notopodia of the posterior part of the thorax are connected by a more or less developed dorsal ridge. Sometimes ridges are developed near the notopodia only.
Modified neuropodia
The size of neuropodia can vary greatly, even between congeners. The neuropodia of TU1 are often the most developed; they gradually decrease in size posteriorly, and their position does not change. Only Auchenoplax shows a remarkable difference. Usually in this genus the neuropodia of TU2 and/or TU1 can be moved ventrally and the rows of their uncini are several times longer than those of the rest TU ().
Figure 3. Neuropodial modification in Auchenoplax. a, Holotype of Auchenoplax crinita, ventral view; b,c, ventral view of two undescribed species of Auchenoplax.
Different types of neuropodia
In most Ampharetidae the shape of thoracic and abdominal neuropodia differs. The uncini of thoracic neuropodia are situated in a furrow; the uncini of abdominal neuropodia at the margin. I propose to call them thoracic and abdominal types. The change of types is very sharp, without any transitional segments. However, the change from the thoracic to the abdominal type of neuropodia does not necessarily match up with the transition from thorax to abdomen. Usually, there are two anterior AU with the thoracic type of neuropodia. This is the case for genera with an Ampharete-type prostomium (Ampharete, Anobothrus, Sosane, Gnathampharete, Lysippe, Eclysippe, Melinnampharete and some others) and for Neosabellides. In Grubianella only the first AU has thoracic-type neuropodia, while in Amage, Amphicteis and Amphysamytha all TU have thoracic-type neuropodia and all AU have abdominal-type neuropodia. In Melinna the last two TU have neuropodia of the abdominal type. In several genera the number of TU varies, but the number of two anterior AU with neuropodia of thoracic-type remains constant: Ampharete always has two, while Amage has none: this means that the number of segments with thoracic-type neuropodia and notopodia varies. Contrarily, in the case of Lysippe, the order of the last segments with thoracic-type neuropodia is constant despite variation in TU; this means that only the number of notopodia varies.
Genera with the simplest types of prostomium like Samythella and Phyllocomus have no difference between thoracic and abdominal neuropodia: all neuropodia are the same type throughout, which, in my opinion, represents the plesiomorphic condition in Ampharetidae. The absence of thoracic neuropodia in Andamanella may be an artefact: in some species (in very small specimens, like the only known specimen of Andamanella) I failed to find neuropodia even after staining.
Uncini: changes along the body
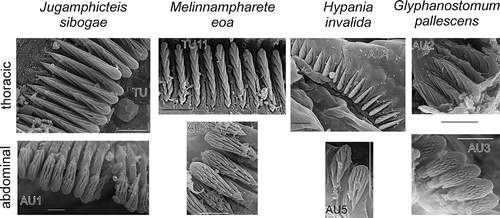
Often the shape of uncini varies among different regions of the body. If so, abdominal uncini are smaller, have more rows of teeth and fewer teeth per row. However, there is no consensus on the taxonomic value of the shape of uncini. Investigation of variation shows that difference in number of rows (2 or 3) and a slight change in number of teeth per row has no importance as this variation can be observed within a single neuropodium (). On the other hand, in some genera the neuropodial uncini of thoracic and abdominal types (which do no not necessary coincide with thorax/abdomen) differ consistently ().
Figure 4. Variation in uncini in a single neuropodium (Lysippe sexcirrata). Scale 10 μm.
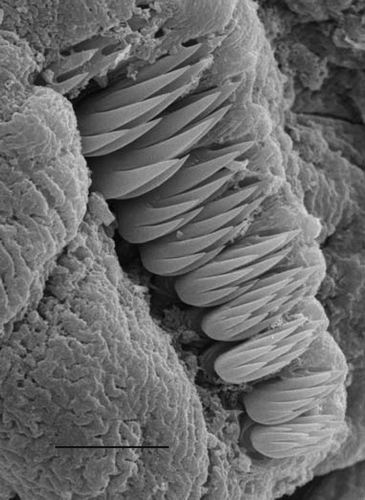
Figure 5. Uncini of neuropodia of thoracic and abdominal types of some genera. All scales 10 μm.
Ventral glandulations
All Ampharetidae build tubes. Ventral glands of anterior segments secrete organic mucus as a matrix for tube construction. Methylene blue staining of glandular fields shows a darker blue than surrounding tissue. Usually there are no distinct ventral shields as in most Terebellinae and Polycirrinae; the glandular surface extends as transversal strip from one neuropodium to another of the same segment through the ventrum. The width of these strips gradually decreases caudally. Among the studied ampharetids only Noanelia has well marked ventral shields like most of the Terebellinae. In Auchenoplax and Eclysippe the thorax is sharply subdivided into two regions: (1) anterior 9–10 TS short with well-developed ventral glandular pads; (2) last 5 TU very long, glandular pads not developed. In all other Ampharetidae all thoracic segments remain similar in length, and the size of glandular pads usually reduce gradually.
Dorsal crest in Melinninae
Usually there is a single dentate crest, but in Melinna elisabethae I have seen up to three crests (one per segment), dorsally decreasing in size and dentation.
Dorsal hooks in Melinninae
Some authors (Fauchald Citation1977; Reuscher et al. Citation2009) propose that Moyanus Chamberlin, Citation1919 differs from Melinna Malmgren by the presence of two pairs of dorsal hooks, rather than one pair. However, Chamberlin (Citation1919) did not state the presence of four hooks, he wrote: ‘Two stout spines, but little exposed, caudad of the branchiae on each side’ (p. 452). Moreover, I have seen several Melinna elisabethae and M. palmata with three hooks (two on one side and one on the other).
Table I contains the synopsis of the proposed taxonomical value of morphological characters of Ampharetidae.
Table I. Taxonomical value of morphological characters in Ampharetidae
Table II. List of examined species.
Remark. Only species are described in Schüller & Jirkov (in prep.) are mentioned as sp. nov
The following characters are considered to be valuable for specific, rather than generic diagnoses: number of branchiae, TU, presence/absence of paleae. However, the degree of variation of these characters within the newly defined genera differs. The most uniform genus is Amphicteis, whose species all have 4 pairs of branchiae, 14 TU, 17 TC, and usually 15 AU; most species have well-developed paleae. On the other hand, Sosane shows great variation of all these characters: it has 9–13 TU, 3–4 pairs of branchiae, paleae present or absent, even the position of modified thoracic segment varies. Other large genera (Amage, Ampharete, Anobothrus) are intermediate in terms of variation.
The branchial arrangement (not number!) is very characteristic for genera (see Jirkov Citation2001, Citation20092009), and may be included in the diagnoses.
Key to genera
The re-evaluation of characters results in emended generic diagnoses and an updated identification key. All previously proposed keys (Malmgren Citation1866; Chamberlin Citation1919; Fauvel Citation1927; Day Citation1964; Fauchald Citation1977; Reuscher et al. Citation2009) have been based on characters such as number of segments, branchiae and presence/absence of paleae, which, in my opinion, are not useful for generic delimitation, as discussed above. This newly proposed key is based only on characters considered valuable on generic level, as discussed above and previously (Jirkov Citation2001, Citation2009).
Only genera that I have examined or that have been described in sufficient detail are included in the key. In some cases I was not able to point out a diagnostic character for genera, but for different reasons (explained below in the generic diagnosis) I would not like to treat them as synonyms. Such genera are separated by commas. Synonyms are in round parentheses after = sign.
| 1. | Three or four anterior segments with vertical rows of minute acicular chaetae. . . . . . . . . . . . . . . Melinninae . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 2
| ||||
| 2. | Hooks (usually one pair) and dorsal crest (usually one and dentate) behind the branchiae present . . . . . . . Melinna, Isolda, Irana, Moyanus
| ||||
| 3. | Prostomium Jugamphicteis-type (,k): trilobed, with transversal nuchal organs, middle lobe simple, anteriorly rounded . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .Jugamphicteis
| ||||
| 4. | Thorax sharply subdivided into two regions: (1) anterior 9–10 TS short with entirely glandular ventrum, (2) last 5 TU very long, glandular pads not developed . . . . . . . . . . . . . . . .5
| ||||
| 5. | Neuropodia of TU2 or/and TU1 enlarged and moved ventrally () . . . . . . . Auchenoplax
| ||||
| 6. | One, usually 5th (4th–6th) from last, pair of posterior notopodia slightly shifted dorsally and connected by low ridge . . . . . . . . . . . . Anobothrus (=Anobothrella, Melythasides, Sosanides)
| ||||
| 7. | – Distinct transversal dorsal ridge between TC3 and TC4 present . . . . . . . Melinnampharete (Eusamytha Hartman, Citation1967 non McIntosh, Citation1885; Eusamythella; Neosamytha)
| ||||
| 8. | Middle lobe of prostomium anteriorly incised or with horns, with or without longitudinal ridges (,h) . . . . . . . . . . . . . . . . . . . . . . . . 9
| ||||
| 9. | Middle lobe of prostomium with longitudinal ridges () Hypania (=Alkmaria, . . . . . . . . . . . . . . . . . . . Hypaniola, Microsamytha, Parhypania)
| ||||
| 10. | Abdominal neuropodia with very long cirri, longer than width of segment . . . . . . . . . . . . . . . . . Samythopsis (= Neopaiwa; Paiwa; Weddelia)
| ||||
| 11. | AU1 with neuropodia of abdominal type, less than 16 AU . . . . . . . . . . . . . . . . . . . . . . . . . . . Amage (= Egamella; Mexamage; Paramage; Phyllampharete)
| ||||
| 12. | 1st segment with ‘horns’ . . . . . . . . Gen. sp. nov.
| ||||
| 13. | Lower lip enlarged, longitudinally grooved . . . . . . . . . . . . . Lysippe (= Lysippides, Paralysippe, Pseudampharete, Pterolysippe, Samytha)
| ||||
| 14. | Numerous jaws present . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . Gnathampharete (= Adercodon)
| ||||
| 15. | 2nd from last notopodia slightly shifted dorsally and connected by low ridge . . . . . . . . Zatsepinia
| ||||
| 16. | Prostomium with pair of tranversal nuchal organs () . . . . . . . . . . . . . . . . Phyllocomus (= Shistocomus; Pavelius, ? Amphisamytha)
| ||||
| 17. | Prostomium laterally with glandular fields . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . Glyphanostomum
| ||||
New proposed diagnosis of genera
Ampharetinae
Diagnosis. Minute acicular chaetae in anterior segments absent.
Amage Malmgren, Citation1866
(= Egamella; Mexamage; Paramage; Phyllampharete)
Diagnosis. Middle lobe of prostomium anteriorly incised or with horns, without longitudinal ridges (). Neuropodia of two types. AU1 with neuropodia of abdominal type, less than 16 AU. Number of AU usually constant for species.
Ampharete Malmgren, Citation1866
(=Asabellides; Parampharete; Pseudosabellides; Pterampharete; Sabellides, ? Amythasides)
Diagnosis. Prostomium Ampharete-type; middle lobe anteriorly rounded (). Neuropodia of two types, their uncini generally similar. Number of AU usually constant for species (most species have 12 or 13 AU). Number of AU with neuropodia of thoracic type always two, while number of TU can be 11 or 12.
Remark. Amythasides seems very similar to Ampharete. The small size of types (or specimens) does not allow a final descision about its taxonomic status.
Amphicteis Grube, 1850
(= Crossostoma; Paramphicteis, Phyllamphictes, Pseudoamphicteis)
Diagnosis. Prostomium Amphicteis-type (): with transversal prominent nuchal organs and with a pair of longitudinal ridges, anteriorly incised. Neuropodia of two types, no AU with thoracic type neuropodia. Thoracic and abdominal uncini generally similar, usually with one row of teeth.
Remark. Always four pairs of branchiae and usually 15 AU.
Anobothrus Levinsen, 1884
(= Anobothrella, Melythasides, Sosanides)
Diagnosis. Prostomium Ampharete-type. One of posterior notopodia (usually 5th from last) slightly shifted dorsally and connected by low ridge. There is a glandular circular band behind parapodia of 1st, 2nd or 3rd TC. There is a pair of nephridial papillae medially behind the branchiae. Neuropodia of two types, their uncini generally similar. Two anterior AU with neuropodia of thoracic type.
Remark. Number of AU usually constant for species (most species have 12 AU).
Auchenoplax Ehlers, 1887
Diagnosis. Prostomium Ampharete-type. Thorax distinctly subdivided into two regions: anterior segments (up to TC-9) several times shorter than posterior one. Neuropodia of TU-2 and/or TU-1 enlarged and moved ventrally (). Notopodia of posterior TU slightly shifted and are connected by more or less developed transversal dorsal ridge. Neuropodia of two types, first two AU with thoracic type ones.
Eclysippe Eliason, 1955
Diagnosis. Prostomium Ampharete-type. Thorax distinctly subdivided into two regions: anterior segments (up to TC-9) several times shorter than posterior one. Anterior neuropodia not enlarged and not moved ventrally. Notopodia of posterior TU slightly shifted and are connected by more or less developed transversal dorsal ridges.
Remark. Genus is very similar to Auchenoplax.
Gen. sp. nov.
Diagnosis. Prostomium Ampharete-type. 1st segment with ‘horns’. No modified noto- and neuropodia. Neuropodia of two types, no AU with thoracic type neuropodia.
Remark. The description of Gen. sp. nov. is currently prepared (Schüller & Jirkov, in prep.).
Glyphanostomum Levinsen, 1884
Diagnosis. Prostomium without lobes, but laterally with glandular fields, often whitish. AU numerous, varying interspecifically. Neuropodia of two types: thoracic (all thoracic and first two abdominal) and abdominal (remaining abdominal). Uncini () of neuropodia of thoracic type with teeth in one row, except lower, paired, uncini of neuropodia of abdominal type with numerous teeth in several rows.
Remark. Lateral prostomial glandular fields are illustrated by Uschakov (Citation1955, figure 138) and Hartman (Citation1969).
Gnathampharete Desbruyères, 1978
(= Adercodon)
Diagnosis. Prostomium Ampharete-type (). Numerous jaws present. Neuropodia of two first abdominal chaetigers of thoracic type, pair of nephridial papillae behind branchiae.
Remarks. Externaly all three species of this genus looks like very small species of Ampharete sensu lato. So examination of the pharyngeal anatomy of Ampharete sensu lato, and especially small species, is needed, as some may belong to Gnathampharete. Examination of the origin of the jaws is warranted.
Grubianella McIntosh, 1885
(= Amagopsis)
Diagnosis. Middle lobe of prostomium anteriorly incised without longitudinal ridges (). AU1 with neuropodia of thoracic type, more than 20 AU.
Remark. Deepwater genus.
Hypania Ostroumow, 1897
(= Alkmaria, Hypaniola, Microsamytha, Parhypania)
Diagnosis. Middle lobe of prostomium anteriorly incised with longitudinal ridges (). Neuropodia and their uncini of two types, uncini of neuropodia of thoracic type with teeth in one row, uncini of neuropodia of abdominal type with numerous teeth in several rows. AU numerous, varying interspecifically.
Remark. Species of this genus occur in fresh or brackish water.
Jugamphicteis Fauchald et Hanckock, 1981
Diagnosis. Prostomium Jugamphicteis-type (,k): trilobed, with transversal prominent nuchal organs, middle lobe simple, anteriorly rounded in small specimens, straight in middle-sized and longitudinally grooved in large. AU1 with enlarged foliaceous rudimental notopodia. Neuropodia and their uncini of two types (): uncini of neuropodia of thoracic-type (all thoracic) with teeth in one row, except upper teeth, uncini of neuropodia of abdominal-type (all abdominal) with numerous teeth in several rows.
Remark. Deepwater genus.
Lysippe Malmgren, Citation1866
(= Lysippides, Paralysippe, Pseudampharete, Pterolysippe, Samytha)
Diagnosis. Prostomium Ampharete-type; middle lobe anteriorly rounded (). Lower lip enlarged, longitudinally grooved. Neuropodia of two types, their uncini generally similar.
Remark. Contrary to Ampharete, number of segments with neuropodia of thoracic type is constant (always 14 uncinigers with neuropodia of thoracic type), while number of TU can be 12–14.
Melinnampharete Annenkova, 1937
(= Eusamytha Hartman, Citation1967 non McIntosh, Citation1885; Eusamythella; Neosamytha)
Diagnosis. Prostomium Ampharete-type. There is a distinct transversal dorsal crest between TC3 and TC4. Neuropodia and their uncini () of two types: uncini of neuropodia of thoracic-type (in type species all thoracic and two anterior abdominal) with teeth in one row, uncini of neuropodia of abdominal-type (in type species starting from AU3) with numerous teeth in several rows.
Remark. All known species have small paleae, 12 TU and 15 AU.
Noanelia Desbruyères et Laubier, Citation1977
Diagnosis. Prostomium Noanelia-type (), buccal tentacles attached outside the mouth, methylene blue staining pattern of ventral glandular shields well marked, while in all other investigated ampharetid species and genera there is no any trace of ventral glandular shields. Neuropodia of two types, thoracic in thorax, AU1 and AU2, remaining AU with neuropodia of abdominal type.
Remarks. The position of this genus within Terebellomorpha remains unresolved. Unlike all other ampharetids, the buccal tentacles are outside the mouth as in Terebellidae. Also, distinct unpaired glandular shields are diagnostic for Terebellinae. On the other hand, moving it to Terebellidae will not clearly resolve the problem. Species of Terebellinae have a double row uncini at least in the posterior thorax, while Noanelia lacks this. Analysis of DNA may help to find the proper position of the genus.
Neosabellides Hessle, 1917
(= ? Amythas Benham, Citation1921)
Diagnosis. Prostomium without lobes but with transversal groove, sometimes lateral ends of groove with more or less deep depressions (nuchal organ?). Contrary to Ampharete-type, this grove does not completely split the prostomium. Neuropodia of two types: thoracic in thorax, AU1 and AU2; remaining AU with neuropodia of abdominal-type.
Remark. The only investigated specimen of Amythas is a holotype of Amythas membranifera Benham, Citation1921. It has everted its tentacle membrane, changing the shape of its prostomium, and was incomplete posteriorly. More specimens are needed to clear the taxonomic position of this genus.
Phyllocomus Grube, 1878
(= Shistocomus; Pavelius, ? Amphisamytha)
Diagnosis. Prostomium without lobes but with a pair of transversal nuchal organs. When tentacles are withdrawn and specimens sufficiently well preserved, nuchal organs can be connected by a groove, and this groove can continue laterally up to the prostomium margin. The number of AU is high and varies individually. All neuropodia and their uncini are of the same type (there is no difference between abdominal- and thoracic-type).
Remark. Specimens known from Phyllocomus, Shistocomus, Pavelius have similar prostomia. As for Amphisamytha, A. vanuatuensis Reuscher, Fiege, Wehe, 2009 has similar prostomia as well as identical types of neuropodia throughout the body.
Samythella Verrill, 1873
Diagnosis. Prostomium simple, without lobes and nuchal organs. Number of AU is high and varies individually. All neuropodia and their uncini of the same type, with teeth in a single row.
Samythopsis McIntosh, 1885
(= Neopaiwa; Paiwa; Weddelia)
Diagnosis. Middle lobe of prostomium anteriorly incised without longitudinal ridges (). Neuropodia of two types: thorax with thoracic-type, abdomen with abdominal-type neuropodia only. Abdominal neuropodia with very long cirri, longer then width of segment.
Remark. Deepwater genus.
Sosane Malmgren, Citation1866
(= Mugga; Muggoides; Sosanella; Sosanopsis; Genus A sensu Uebelacker, Citation1984; ? Melinnata)
Diagnosis. Prostomium Ampharete-type. One of posterior notopodia (usually last, 2nd or 3rd from last) flattened and shifted dorsally. Thorax and 0–2 anterior AU with thoracic0type neuropodia, remaining AU with abdominal-type neuropodia.
Zatsepinia Jirkov, 1986
Diagnosis. Prostomium without ridges, grooves and nuchal organs. Notopodia of 2nd from last TU slightly shifted dorsally and connected by low transversal ridge. Neuropodia of two types: thoracic in thorax, AU1 and AU2 and abdominal in remaining AU.
Melinninae
Diagnosis. Three or four anterior segments with vertical rows of minute acicular chaetae.
Remark. All Melinninae have varying and comparatively large number of AU.
Melinna Malmgren, Citation1866, Isolda, Irana, Moyanus
Diagnosis. There are hooks (usually one pair) and dorsal crest (usually one and dentate) behind the branchiae.
Remarks. Here the difference between these four genera is not found to be sound. Since I have not examined type material of all species, additional characters on the generic level may exist. I suppose for the present time it will be better not to change the taxonomy of this group until we have additional information, especially since most species of this group belong to Melinna, the type genus of the subfamily, which would have be treated as junior synonym of Isolda Müller (in Grube) Citation1858.
Melinnides Wesenberg-Lund, Citation1950
(= Melinnantipoda)
Diagnosis. There is only dorsal crest behind the branchiae, no hooks.
Melinnopsis McIntosh, 1885
(=Amelinna; Melinnexis, Melinnopsides)
Diagnosis. There are neither hooks nor crest behind the branchiae.
Acknowledgements
I am very much in debt to Dr G. Paterson (Natural History Museum, London), Dr Myriam Shüller (Forschungsinstitut Senckenberg, Abteilung Deutsches Zentrum für Marine Biodiversitätsforschung), Dr T.A. Britaev (N.N. Severtsov Institute of Ecology and Evolution of the Russian Academy of Sciences), Dr N. Budaeva (P.P. Shirshov Institute of Oceanology of the Russian Academy of Sciences), Dr D. Hall and Dr T. Worsfold (Thomson Unicomarine Ltd), Dr G.N. Buzhinskaja and S. Potin (Zoological Institute of Russian Academy of Sciences St. Petersburg), and Dr G. Hartman-Schröder (Zoologisches Institut und Zoologisches Museum, Universität Hamburg) for kindly allowing me to examine their materials for the present study.
The visit to the Zoologisches Institut und Zoologisches Museum, Universität Hamburg was possible due to financial support of the DAAD, visiting Natural History Museum, London due to financial supports of the Linnaean Society and the Thomson Unicomarine Ltd, visiting DZMB Wilhelmshaven for the investigation of Antarctic Polychaeta was supported by the grant from the CeDAMar. Special thanks to Emma Delduca (MES Ltd, UK), who corrected my English. Finally I wish to thank an anonymous referee and the editor who spent a lot of time trying to make my paper better.
Related Research Data
References
- Chamberlin , RV. 1919 . The Annelida Polychaeta . Memoirs of the Museum of Comparative Zoology at Harvard College 48 , : 1 – 514 .
- Day , JH. 1964 . A review of the family Ampharetidae (Polychaeta) . Annals of the South African Museum , 48 : 97 – 120 .
- Desbruyères , D and Laubier , L. 1977 . “ Noanelia hartmanae gen. sp. n., Ampharetidae (Annelides Polychetes Sedentaires) abyssal du Golfe de Gascogne ” . In Essays on Polychaetous Annelids, in memory of Dr Olga Hartman , Edited by: Reish , DJ and Fauchald , K . 199 – 207 . Los Angeles, California : A special publication of the Allan Hancock Foundation University of Southern California .
- Fauchald , K. 1977 . The polychaete worms. Definitions and keys to the Orders, Families and Genera . Natural History Museum of Los Angeles County, Science Series , 28 : 1 – 188 .
- Fauvel , P. 1927 . Polychètes sédentaires. Addenda aux errantes, Archiannélides, Myzostomaires , 1 – 494 . Paris : Paul Lechevalier .
- Hilbig , B. 2000 . “ Family Ampharetidae Malmgren, 1867 ” . In Taxonomic atlas of the benthic fauna of the Santa Maria Basin and Western Santa Barbara Channel. The Annelida Part 4. Polychaeta: Flabelligeridae to Sternaspidae , Edited by: Blake , JA , Hilbig , B and Scott , PV . 169 – 230 . Santa Barbara, CA : Santa Barbara Museum of Natural History .
- Holthe , T. 2002 . “ One new genus and three new species of the Ampharetidae (Polychaeta: Terebellida) from the BIOSHELF Project ” . In Proceedings of the International Workshop on the Polychaetes of the Andaman Sea, held 3 June–27 August 1997 at Phuket Marine Biological Center , Edited by: Eibye-Jacobsen , D . 345 – 351 . Thailand : Department of Fisheries .
- Jirkov , IA. 2001 . Polychaeta of the North Polar Basin , 1 – 632 . Moscow : Yanus-K . in Russian
- Jirkov , IA. 2009 . Revision of Ampharetidae (Polychaeta) with modified thoracic notopodia . Invertebrate Zoology , 5 : 111 – 132 .
- Malmgren , AJ. 1866 . Nordiska Hafs-Annulater . Öfversigt af Kongliga Vetenskapsakademiens Förhandlingar , 22 : 355 – 410 .
- Orrhage , L. 2001 . On the anatomy of the central nervous system and the morphological value of the anterior end appendages of Ampharetidae, Pectinariidae and Terebellidae Polychaeta . Stockholm: Acta Zoologica , 82 : 57 – 71 .
- Reuscher , M , Fiege , D and Wehe , T. 2009 . Four new species of Ampharetidae (Annelida: Polychaeta) from Pacific hot vents and cold seeps, with a key and synoptic table of characters for all genera . Zootaxa , 2191 : 1 – 40 .
- Schüller , M. and Jirkov , I. in prep . New Ampharetidae from the Atlantic sector of the deep Southern Ocean and shallow Patagonian waters, excluding species of the Sosane-complex (sensu Jirkov, 1994)
- Uschakov , PV. 1955 . Polychaeta of the Far-Eastern seas of the USSR . Identification key for fauna of the USSR, published by the Zoological Institute of the USSR Academy of Science , 56 : 1 – 445 .
Additional references of original species descriptions or redescriptions
- Annenkova , NP. 1929 . Beiträge zur Kenntnis der Polychaeten-Fauna der USSR. I. Fam. Pectinariidae Quatrefages (Amphictenidae Malmgren) und Ampharetidae Malmgren . Annuaire du Musée Zoologique de l'Acad.des Sciences de l'URSS , 30 : 477 – 502 . in German
- Annenkova , NP. 1931 . Zur Polychaetenfauna von Franz-Joseph Land (Melinnexis gen. nov. arctica sp. nov.) . Zoologischer Anzeiger , 95 : 269 – 272 .
- Annenkova , NP. 1934 . Kurze Übersicht der Polychaeten der Litoralzone der Bering-Insel (Kommador-Inseln) nebst Beschreibung neuer Arten . Zoologischer Anzeiger , 106 : 322 – 331 .
- Annenkova , NP. 1937 . Polychaetous fauna of the Northern part of the Japanese Sea . Explorations des Mers de l'URSS , 23 : 139 – 216 . in Russian, English summary
- Augener , H. 1914 . “ Polychaeta Sedentaria ” . In Die Fauna Südwest-Australiens Edited by: Michaelsen , W and Harmeyer , R . Vol. 5 , 1 – 170 .
- Augener , H. 1918 . Polychaeta . Beiträge zur Kenntnis des Meerresfauna West-Afrikas , 2 : 67 – 625 .
- Augener , H. 1928 . Beitrag zur Polychaetenfauna der Ostsee . Zeitschrift für Morphologie und Ökologie der Tiere , 11 : 102 – 104 .
- Banse , K. 1979 . Ampharetidae (Polychaeta) from British Columbia and Washington . Canadian Journal of Zoology , 57 : 1543 – 1552 .
- Benham , WB. 1921 . Polychaeta . Scientific Reports Australian Antarctic Expedition Zoology and Botany , 6 : 1 – 128 .
- Benham , WB. 1927 . Polychaeta . British Antarctic ‘Terra Nova’ Expedition Natural History Reports, Zoology , 7 : 47 – 182 .
- Caullery , M. 1944 . Polychètes sédentaires de l'expédition du Siboga: Ariciidae, Spionidae, Chaetopteridae, Chloraemidae, Opheliidae, Oweniidae, Sabellariidae, Sternaspidae, Amphictenidae, Ampharetidae, Terebellidae. Siboga-Expeditie . Résultats des explorations zoologiques, botaniques, océanographiques et géologiques , 24 : 1 – 204 .
- Chamberlin , RV. 1919 . The Annelida Polychaeta . Memoirs of the Museum of Comparative Zoölogy at Harvard College , 48 : 1 – 514 .
- Chamberlin , RV. 1920 . “ The Polychaetes Collected by the Canadian Arctic expedition, 1913–18 ” . In Report of the Canadian Arctic expedition 1913–189. Annelids, parasitic worms, Protozoans, etc , Edited by: Polychaeta , B. 1 – 41 . Ottawa : Thomas Mulvey .
- *Claparède , É . 1863 . Beobachtungen über Anatomie und Entwicklungsgeschichte wirbelloser Thiere an der Küste von Normandie angestellt , 1 – 120 . Leipzig : W. Engelmann .
- Day , JH. 1955 . The Polychaeta of South Africa. Part 3. Sedentary species from Cape shores and estuaries . Journal of the Linnean Society of London , 42 ( 287 ) : 407 – 452 .
- Day , JH. 1961 . The Polychaete fauna of South Africa. Part 6. Sedentary species dredged off Cape coasts with a few new records from the shore . Zoological Journal of the Linnaean Society of London , 44 : 463 – 560 .
- Day , JH. 1963 . The Polychaete fauna of South Africa. Part 8: New species and records from grab samples and dredgings. Bulletin of the British Museum (Natural History), Ser . Zoology , 10 : 384 – 445 .
- Desbruyères , D. 1978 . Melythasides laubieri gen. sp. nov. Ampharetidae (Annélides polychètes sédentaires) abyssal de la mer de Norvège . Bulletin du muséum national d'Histoire naturelle, Paris , 3 ( 514 ) : 231 – 238 .
- Desbruyères , D. 1978 . Un Ampharetidae (Annélides polychètes sédentaires) à structure buccale aberrante: Gnathampharete paradoxa gen. sp. n . Compte Rendus des séances de l'Académie des Sciences, Série D , 286 : 281 – 284 .
- Desbruyères , D and Laubier , L. 1977 . “ Noanelia hartmanae gen. sp. n., Ampharetidae (Annélides polychètes sédentaires) abyssal du Golfe de Gascogne ” . In Essays on Polychaetous Annelids in Memory of Dr , Edited by: Reish , DJ and Fauchald , K. 279 – 287 . Olga Hartman, Los Angeles : the Allan Hancock Foundation, University of Southern California .
- Desbruyères , D and Laubier , L. 1996 . A new genus and species of ampharetid polychaete from deep-sea hydrothermal vent community in the Azores triple-junction area . Proceedings of the Biological Society of Washington , 109 : 248 – 255 .
- Ehlers , E. 1887 . Reports on the Annelids. Report on the results of dredging, under the Direction of L. F. Pourtalès, during the years 1868–1870, and of Alexander Agassiz, in the Gulf of Mexico (1877–78), and in the Caribbean Sea (1878–79), on the U.S. coast survey steamer Blake, Lieut. Com. C.D. Sigsbee, U.S.N., and Commander J.R Bartlett, U.S.N . Commanding. Memoirs of the Museum of Comparative Zoology at Harvard College , 15 ( i–vi ) : 1 – 335 .
- Ehlers , E. 1908 . Die Bodensaessigen Anneliden aus den Sammlungen der deutschen Tiefsee-Expedition . Wissenschaftliche Ergebnisse der deutschen Tiefsee-Expedition auf dem Dampfer ‘Valdivia’ 1898–1899 , 16 : 1 – 168 .
- Ehlers , E. 1913 . Die Polychaeten-Sammlungen der deutschen Südpolar-Expedition 1901–1903 . Deutche Südpolar-Expedition , 13 : 397 – 598 .
- Eliason , A. 1955 . Neue oder wenig bekannte schwedische Ampharetiden (Polychaeta) – Göteborgs Königlich vetenskaps- och vitterhetssamhälles handlingar, Ser . Sjätte Följden, ser. B , 6 ( 16 ) : 1 – 17 .
- Fauchald , K. 1972 . Benthic polychaetous annelids from deep water off western Mexico and adjacent areas in the eastern Pacific Ocean . Allan Hancock Monographs in Marine Biology , 7 : 1 – 575 .
- Fauchald , K and Hancock , DR. 1981 . Deep-water polychaetes from a transect off Central Oregon . Allan Hancock Foundation Monograph , 11 : 1 – 73 .
- Grimm , OA. 1877 . The Caspian Sea and its Fauna. Notebook 2. Transactions of the Aralo-Caspian Expedition 2. M . Stasjulevich Press, S-Petersburg , : 1 – 105 . in Russian
- *Grube , A.E. 1858 . Einiges über die Annelidenfauna der Insel Santa Catharina an der brasilianischen Küste . Archiv für Naturgeschichte, Berlin , 24 : 211 – 220 .
- *Grube , AE. 1860 . Beschreibung neuer order wenig bekannter Anneliden. Archiv für Naturgeschichte, Berlin. 4 . Beitrag: Zahlreiche Gattungen , 26 : 71 – 118 .
- *Grube , AE. 1862 . Noch ein Wort über die Capitellen und ihre Stelle im Systeme der Anneliden . Archiv für Naturgeschichte, Berlin , 28 : 366 – 378 .
- *Grube , AE. 1877 . Anneliden-Ausbeute S.M.S. Gazelle. Monatsberichte der Königlich Preußischen Akademie der Wissenschaften zu Berlin , : 509 – 544 .
- *Grube , AE. 1878 . Annulata Semperiana. Beiträge zur Kenntniss der Annelidenfauna der Philippinen nach den von Herrn Prof. Semper mitgebrachten Sammlungen. Memoires de L'Academie Imperiale des Sciences de St.Petersbourg . Serie 7 , 25 ( 8 ) : 1 – 300 .
- Hartley , JP. 1985 . The re-establishment of Amphicteis midas (Gosse, 1855) and redescription of the type material of A. gunneri (M. Sars, 1835) (Polychaeta: Ampharetidae) . Sarsia , 70 : 309 – 315 .
- Hartman , O. 1965 . Deep-water benthic polychaetous annelids off New England to Bermuda and other North Atlantic areas . Allan Hancock Foundation Occasional Paper , 28 : 1 – 378 .
- Hartman , O. 1967 . Polychaetous annelids collected by the USNS Eltanin and Staten Island cruises, chiefly from Antarctic Seas . Allan Hancock Monographs in Marine Biology , 2 : 1 – 387 .
- Hartman , O. 1969 . Atlas of the sedentariate polychaetous annelids from California , 1 – 812 . Los Angeles, CA : California Allan Hancock Foundation University of Southern California .
- Hartman , O. 1971 . Abyssal polychaetous annelids from the Mozambique Basin off southeast Africa, with a compendium of abyssal polychaetous annelids from world-wide areas . Journal of the Fisheries Research Board of Canada , 28 : 1407 – 1428 .
- Hartman , O. 1978 . Polychaeta from the Weddell Sea Quadrant, Antarctica. Biology of the Antarctic Seas VI . Antarctic Research Series , 26 : 125 – 223 .
- Hartman , O. and Fauchald , K. 1971 . Deep-water benthic polychaetous annelids off New England to Bermuda and other North Atlantic areas . Part 2. Allan Hancock Monographs in Marine Biology , 6 : 1 – 327 .
- Hartmann-Schröder , G. and Hartmann , G. 1965 . Zur Kenntnis des Sublitorals der chilenischen Küste unter besonderer Berücksichtigung der Polychaeten und Ostracoden . Mitteilungen aus dem Hamburgischen zoologischen Museum und Institut , 62 : 1 – 384 .
- Hartmann-Schröder , G. 1981 . Zur Kenntnis des Eulitorals der australischen Küsten unter besonderer Berücksichtigung der Polychaeten und Ostracoden. Teil 6. Die Polychaeten der tropisch–subtropischen Westküste Australiens (zwischen Exmouth im Norden und Cervantes im Süden) . Mitteilungen aus dem Hamburgischen zoologischen Museum und Institut , 78 : 19 – 96 .
- *Haswell , WA. 1883 . A monograph of the Australian Aphroditea . Proceedings of the Linnean Society of New South Wales , 7 : 250 – 299 .
- Hessle , C. 1917 . Zur Kenntnis der terebellomorphen Polychaeta. Akademische Abhandlung welche zur Erlangung der Doktorwűrde mit Erlaubnis der Matematisch-Naturwissenschaftlichen sektor der Weitberűhmten philosophichen fakultäzu Uppsala , 39 – 258 . Uppsala : Almquist & Wiksells Boktryckeri-A.-B .
- Hilbig , B. 2000 . “ Family Ampharetidae Malmgren, 1867. Taxonomic Atlas of the Benthic Fauna of the Santa Maria Basin and the Western Santa Barbara Channel Vol. 7 ” . In The Annelida Pt.4 Polychaeta, Flabelligeridae to Sternaspidae , Edited by: Blake , JA , Hilbig , B and Scott , PH . 169 – 230 . Santa Barbara, CA : Santa Barbara Museum of Natural History .
- Hlebovich , VV. 1964 . “ Polychaeta from the northern Greenland Sea and regions of Spitsbergen and Franz-Joseph-Land ” . In Arctic and Antarctic Institute Transactions Vol. 259 , 167 – 180 . in Russian
- Holthe , T. 1986 . Polychaeta Terebellomorpha from the north Norwegian Sea and the Polar Sea, with description of Mugga bathyalis sp. n. and Ymerana pteropoda gen. n. and sp. n . Sarsia , 71 : 227 – 234 .
- Horst , R. 1919 . Twee sedentaire Polychaeten uit he brakke water van Nederland . Zoologische Mededeelingen (Leiden) , 5 : 100 – 102 .
- Hutchings , P. 1977 . Terebelliform polychaeta of the families Ampharetidae, Terebellidae and Trichobranchidae from Australia, chiefly from Moreton Bay, Queensland . Records of the Australian Museum , 31 : 1 – 38 .
- Hutchings , PA and Rainer , S. 1979 . The polychaete fauna of Careel Bay, Pittwater, New South Wales, Australia . Journal of Natural History , 13 : 745 – 796 .
- Jirkov , IA. 1997 . Ampharete petersenae sp. nov. (Ampharetidae: Polychaeta) from the Northern Atlantic . Zoologicheskij Zhournal , 76 : 1418 – 1420 . in Russian
- Jirkov , IA. 1985 . Amphicteis ninonae sp. nov. (Polychaeta, Ampharetidae) from the northern waters . Zoologicheskij Zhournal , 64 : 1894 – 1897 . in Russian with English summary
- Jirkov , IA. 1986 . Zatsepinia rittichae gen. et sp. nov. (Polychaeta, Ampharetidae) from the Norwegian and Barents seas . Zoologicheskij Zhournal , 65 : 289 – 291 . in Russian with English summary
- Jirkov , IA. 1994 . Two new species Ampharete (Polychaeta: Ampharetidae) form the North-Western Pacific with discussion of paleae as taxonomic character of Ampharetinae . Zoologicheskij Zhournal , 73 : 28 – 32 . in Russian with English summary
- Jirkov , IA. 1994 . Sosane holthei sp.nov. (Polychaeta: Ampharetidae) form North-Western Pacific with review of Sosane and similar genera . Zoologicheskij Zhournal , 73 : 33 – 38 . in Russian with English summary
- Johnson , HP. 1901 . The Polychaeta of the Puget Sound region . Proceedings of the Boston Society for Natural History , 29 : 381 – 437 .
- Kinberg , JGH. 1866 . Annulata nova . Öfversigt af Königlich Vetenskapsakademiens förhandlingar, Stockholm , 23 : 97 – 103 .
- Kucheruk , NV. 1976 . Polychaeta annelids of family Ampharetidae from deep-water part of Alaska Bay . Transactions of the P.P. Shirshov Institute of Oceanology , 99 : 91 – 101 . in Russian with English summary
- Kuznetsov , AP and Levenstein , RYa. 1988 . Pavelius uschakovi gen. et sp. n. (Polychaeta, Ampharetidae) from the Sea of Okhotsk Paramushir clathrate hydrates field . Zoologicheskij Zhournal , 67 : 819 – 825 . in Russian with English summary
- Laubier , L. 1973 . Uschakovius enigmaticus gen. sp. n., Ampharetidae (Annélides Polychètes sédentaires) aberrant de Méditerranée orientale profonde . Compte Rendus des séances de l'Académie des Sciences, Série D , 277 : 2723 – 2725 .
- Levenstein , RYa. 1978 . Annelida (Polychaeta) from the deep waters of the Pacific region of the Antarctic . Transactions of the P.P. Shirshov Institute of Oceanology , 113 : 73 – 87 . in Russian with English summary
- Levinsen , GMR. 1884 . Systematisk-geografisk Oversigt over de nordiske Annulata, Gephyrea, Chaetognathi og Balanoglossi . Videnskabelige Meddelelser fra Dansk naturhistorisk Forening i Köbenhavn , 1883 : 92 – 350 .
- Mackie , ASY. 1994 . Adercodon pleijeli gen. et sp. nov. (Polychaeta, Ampharetidae) from the Mediterranean Sea. In: Dauvin J-C, Laubier L, Reish D . Actes de la 4eme Conference internationale des Polychetes, Mémoires du muséum national d'Histoire naturelle , 162 : 243 – 250 .
- Mackie , ASY and Pleijel , F. 1995 . A review of the Melinna cristata-species group (Polychaeta: Ampharetidae) in the northeastern Atlantic . Mitteilungen aus dem Hamburgischen zoologischen Museum und Institut , 92 : 103 – 124 .
- Malmgren , AJ. 1866 . Nordiska Hafs-Annulater. Öfvers af. Vetensk . kad. Stockholm, Förh. , 5 : 355 – 410 .
- *Marion , AF. 1876 . Sur les annélides de Marseille . Rev. sci. nat. Montpellier , 4 : 301 – 312 .
- McIntosh , WC. 1885 . Report on the Annelida Polychæta collected by H.M.S. Challenger during the years 1873–76. Report on the Scientific Results of the Voyage of H.M.S. Challenger during the years 1873–76 under the Command of Captain George S. Nares, R.N., F.R.S. and the late Captain Frank Tourle Thomson, R.N . Zoology , 12 ( i–xxxvi ) : 1 – 554 .
- McIntosh , WC. 1914 . Notes from the Gatty Marine Laboratory, St. Andrews. No. 36. 1. On the ventral furrows of the lesser rorqual (Balaenoptera acutorostrata, Lacépède). 2. On some of the species of Prionospio, Malmgren. 3. On the British Amphictenidae. 4 . On the British Ampharetidae. Annals and Magazine of Natural History , 13 : 77 – 110 .
- McIntosh , WC. 1922 . Notes from the Gatty Marine Laboratory, St. Andrews. No. 44. 1. On new and rare Polychaeta, Gephyrea, etc. from various regions. 2 . Recent additions to the British marine Polychaeta (continued). Annals and Magazine of Natural History , 9 ( 49 ) : 1 – 30 .
- Mohammad , MBM. 1971 . Intertidal polychaetes from Kuwait, Arabian Gulf, with descriptions of three new species . Journal of Zoology, London , 163 : 285 – 303 .
- Monro , CCA. 1939 . Polychaeta. Antarctic research expedition 1929–1931 . Adelaide, Australia. Reports – series B (zoology and botany) , 4 ( 4 ) : 89 – 156 .
- Moore , JP. 1906 . Additional new species of Polychaeta from the North Pacific . Proceedings of the Academy of Natural Sciences, Philadelphia , 1906 : 217 – 260 .
- Russell , DE. 1987 . Paeadampharete acutiseries, a new genus and species of Ampharetidae (Polychaeta) from the North Atlantic Hebble area, exhibiting progenesis and broad intraspecific variation . Bulletin of the Biological Society of Washington , 7 : 140 – 151 .
- Salazar-Vallejo , SI. 1996 . Sabellides manriquei new species from the Eastern Pacific, and redescription of Sabellides oculata Webster from the Northwestern Atlantic (Polychaeta: Ampharetidae) . Bulletin of Marine Science , 59 : 142 – 149 .
- Sars , M. 1835 . Beskrivelser og Iagttagelser over nogle moerkelige eller nye i Havet ved den Bergenske Kyst levende Dyr af Polypernes, Acalephernes, Radiaternes, Annelidernes og Molluskernes classer, med en kort Oversigt over de hidtil af Forfatten sammesteds fundne Arter og deres Forekommen , xii – 81 . Bergen : Thorstein Hallagers Forlag hos Chr. Dahl, R. S. .
- Sars , M. 1851 . Beretning om en i Sommeren 1849 foretagen zoologisk Reise i Lofoten og Finmarken . Nyt Magazin for Naturvidenskaberne , 6 : 121 – 211 .
- Sars , M. 1856 . Nye Annelider. Fredrik D. Beyer. Bergen . Fauna littoralis Norvegiae , 2 : 1 – 24 .
- Sars , M. 1865 . “ Fortsatte Bidrag til Kundskaben om Norges Annelider ” . In Forhandlinger fra Videnskabs-Selskabet i Christiania 8 – 20 .
- Schüller , M. 2008 . New polychaete species collected during the expeditions ANDEEP I, II, and III to the deep Atlantic sector of the Southern Ocean in the austral summers 2002 and 2005 — Ampharetidae, Opheliidae, and Scalibregmatidae . Zootaxa , 1705 : 51 – 68 .
- Solis-Weiss , V. 1993 . Grassleia hydrothermalis, a new genus and species of Ampharetidae (Annelida: Polychaeta) from the hydrothermal vents off the Oregon coast (U.S.A.) at Gorda Ridge . Proceedings of the Biological Society of Washington , 106 : 661 – 665 .
- Théel , HJ. 1879 . Les Annélides Polychètes des mers de la Nouvelle-Zemble . Kungliga Svenska vetenskapsakademiens handlingar , 16 : 1 – 75 .
- Treadwell , AL. 1926 . Polychaetous annelids collected by captain R.A.Barlett in Alaska in 1924, with description on new species . American Museum Novitates , 223 : 1 – 8 .
- Uebelacker , JM. 1984 . “ Family Ampharetidae Malmgren, 1867 ” . In Taxonomic guide to the Polychaetes of the Northern Gulf of Mexico. Final Report to the Minerals Management Service, contract 14-12-001-29091 , Edited by: Uebelacker , JM and Johnson , PG . Mobile, AL : Barry A. Vittor & Associates Inc. VII, 51-1–51-32 .
- Uschakov , PV. 1952 . Bathypelagic and deep-water Polychaeta from Pacific waters off Kamchatka . Investigations of Far-Eastern Seas of the USSR , 3 : 103 – 112 . in Russian
- Uschakov , PV. 1955 . “ Polychaeta of the Far Eastern Seas of the USSR. Identification keys for USSR fauna published by the Zoological institute of Science Akademia of USSR 56: 1–446 (in Russian) ” . In English edition 1965. Polychaeta of the Far Eastern Seas of the USSR , 1 – 419 . Jerusalem : Israel Program for Scientific Translations .
- Verrill , AE. 1873 . Report upon the invertebrate animals of Vineyard Sound and the adjacent waters, with an account of the physical characters of the region . Report of the United States Commission for Fisheries, 1871–72. , : 295 – 778 .
- *Wagner , N. 1885 . Die Wirbellosen des Weissen Meeres , 1 – 171 . Leipzig : Wilhelm Engelmann .
- Wesenberg-Lund , E. 1949 . Polychaetes of the Iranian Gulf . Danish Scientific Investigations in Iran , 4 : 247 – 400 .
- Wesenberg-Lund , E. 1950 . Polychæta . The Danish Ingolf-Expedition , 4 ( 14 ) : 1 – 92 .
- Williams , SJ. 1987 . Taxonomic notes on some Ampharetidae (Polychaeta) from Southern California . Bulletin of the Biological Society of Washington , 7 : 251 – 258 .
- Wirén , A. 1883 . Chaetopoder fran Sibiriska Ishafvet och Berings Haf insamlade under Vega-Expeditionen 1878–79 . Vega-exped. vetenskapliga Iakttagelser , 2 : 283 – 428 .
- Zottoli , RA. 1982 . Two new genera of deep-sea polychaete worms of the family Ampharetidae and the role of one species in deep-sea ecosystems . Proceedings of the Biological Society of Washington , 95 : 48 – 57 .