Abstract
The food habits of rough ray Raja radula were investigated, with respect to sex, size and season, based on stomach content analyses of specimens collected from commercial fisheries, monthly during 2007 in the Gulf of Gabès (Central Mediterranean Sea). Of 950 stomachs examined, 183 were empty (19.26%). The overall diet of R. radula mainly consisted of crustacean decapods, teleosts and molluscs. Polycheates, sipunculids and echinoderms were minor dietary components. No differences were observed in the major preys consumed between the sexes, but ontogenetic and seasonal changes were found. The dietary trend with size shifted from crustaceans to teleosts. The consumptions of minor preys were independent from both size and season. Quantitative analysis and graphical methods indicated the rough ray to be a specialized feeder.
Introduction
Skates are often taken as a by-catch in demersal fisheries, and historically they have been of low economic worth in the Mediterranean Sea. Recently, however their commercial importance has been increasing and they are now a valuable by-catch of demersal fisheries (Abella & Serena Citation2005). Thus, ecological data of skates are needed for the proper management of their captures.
The rough ray, Raja radula Delaroche 1809, is a relatively small skate endemic to the Mediterranean Sea (Serena et al. Citation2010). The species abound along Tunisian coasts, mainly in the Gulf of Gabès where it is taken as a by-catch by trawlers, demersal gill-nets and longlines and usually captures are marketed.
In spite of the increasing fishing pressure, there is a paucity of information on rough ray life history resulting in an assessment of ‘Data Deficient’ on IUCN Red List assessments for Chondrichthyans in the Mediterranean Sea (Cavanagh & Gibson Citation2007). Relatively little information is available on the reproductive biology and feeding ecology of rough ray. Only two studies have provided quantitative descriptions of the diet of the rough ray in the Mediterranean Sea; in the Gulf of Tunis (Capapé & Azouz Citation1975) and the off water of Alexandria (Egypt) (Abdel-Aziz Citation1986). Anecdotal information on the diet of the species has been reported from the Aegean Sea (Kyrtatos Citation1982). Published information revealed that this species preys mainly on crustaceans, followed by fishes and molluscs.
Food habits are fundamental to understanding the role that the predator plays in the trophic structure of marine community. Thus, the objectives of this study were to: (1) quantify the diet composition, (2) examine potential diet differences by sex, skate size, and season (3) qualitatively assess feeding strategy. This study will to strengthen our knowledge on the feeding biology of R. radula in the Mediterranean Sea.
Materials and methods
Diet composition
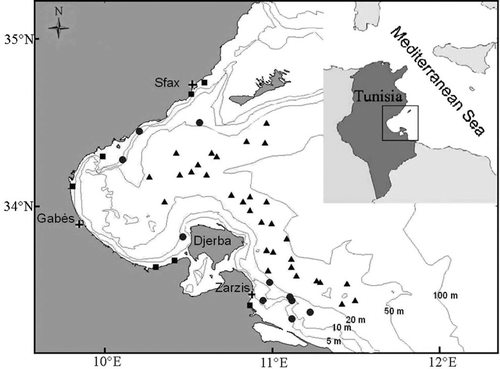
Samples of R. radula were brought regularly twice a month, from January to December 2007 off the Gulf of Gabès, by the commercial trawl using a 22 mm stretched mesh size cod-end and operating over 30 m depth. Additionally, 10 specimens were obtained from longline and demersal gillnets fisheries operating respectively by 30--50 m and 10--40 m depth range, generally on sandy and muddy bottoms (). In longline, the primary bait used was clupeids and this was excluded from the diet analysis. Skates were frozen onboard, subsequently thawed and analysed in the laboratory. Skates were sexed, measured for total length (TL) to the nearest millimetre. The stomachs were then removed, their preys were identified to the lowest possible taxonomic level, counted and weighed in wet condition to the nearest 0.01 g. Vacuity index (VI) was calculated. The variation in VI was tested by a chi-square test over a contingency table of the number of empty stomachs.
Figure 1. Map of the Gulf of Gabès (Tunisia, Central Mediterranean Sea) showing the sampling localities. Triangle (trawls), squared (demersal gillnets) and circle (longline).
The importance of each prey was determined by calculating the index of relative importance (IRI) (Pinkas et al. Citation1971):
Where %F, %N and %M are the percentage contributions of a prey category in terms of frequency of occurrence, number and mass, respectively, in the stomachs with food.
The IRI values were converted to a percentage to facilitate comparisons between prey items (Cortés Citation1997).
Diet shifts
Dietary shifts with sex, predator's size (TL) and season were evaluated using Multivariate analysis of variance (MANOVA) and applying Morisita's dietary overlap index (C H ) (Krebs Citation1989) to test for diet similarities between sexes, size classes and season.
Where pij is the proportion of prey category i (based on %IRI) used by size class j, and pik is the proportion of prey category i used by size class k. The degree of overlap was based on Langton's (Citation1982) scale: low (0–0.29), medium (0.30–0.59) and high overlap (>0.60).
Statistical analyses were carried out considering the main prey categories: Decapods, Isopods, Amphipods, Mysidacea, Stomatopods, demersal fish, pelagic fish, Cephalopods, Gasteropods, Polycheates, Sipunculids and Echinoderms. In addition, specimens were grouped in three size classes: I < 40 cm, II 40--55 cm and III > 55 cm TL. The number of each prey category was considered the dependant variable and season (winter, spring, summer and autumn); sex (F or M) and class (I, II, III) were defined as factors.
The multivariate F value (Wilks' lambda) based on a comparison of the error variance/covariance matrix and the effect variance/covariance matrix was applied to test differences in the diet. An ANOVA was further performed to identify the main prey groups responsible for the major differences among factors: sex, season and size. The significance level adopted was 5%.
Level trophic and feeding strategy
To determine the position of R. radula within the food web, trophic level (TL) was estimated following Cortés (Citation1999) as:
Where TLj is the trophic level of each prey category j, Pj is the proportion of each prey category j (based on %IRI) in the diet of R. radula, and n is the total number of prey categories. The taxonomic categories used to calculate the standardized trophic level of R. radula were Teleosts, Cephalopoda, Mollusca (excluding cephalopods), Decapods, Amphipods (Amphipods and Isopods), Mysids, Other crustaceans (stomatopods), Polychaetes and other invertebrates (sipunculids and echinoderms). The trophic level of each prey category was obtained from Ebert and Bizarro (Citation2007).
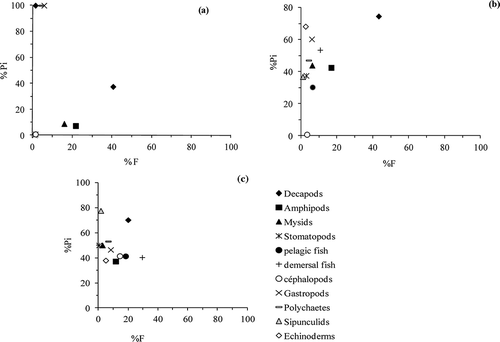
The feeding strategy of R. radula in terms of specialization and generalization was studied by plotting prey specific abundance (Pi) against %F (Amundsen et al. Citation1996). Pi was calculated as the number of a prey i divided by the total number of prey in the stomachs that contained prey i, expressed as a percentage. A single or a few points located at the upper right of the diagram would indicate a specialized diet, but if all prey points were located along or below the diagonal from the upper left to the lower right of the plot, the trophic niche breadth would be broad. This graphical representation was also used to analyse prey importance (Amundsen et al. Citation1996).
Prey diversity (H') was calculated for each size class using Shannon–Weiner diversity index (Krebs Citation1989).
Where Pi is the proportion of individuals in the ith species.
Results
Diet composition
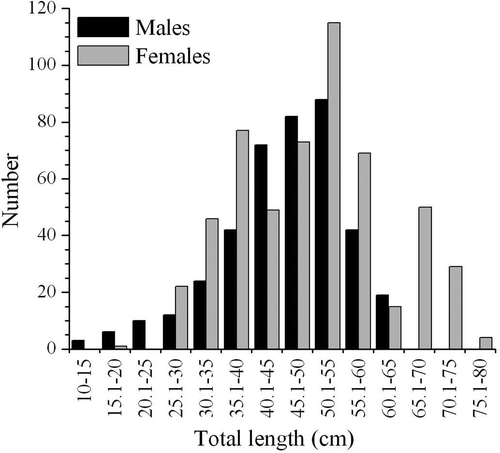
Stomach contents of 550 females and 400 males were analyzed in this study. The total length varied between 13.4–65 cm for males and 18.5–80 cm for females, with skates between 35 cm and 60 cm TL being the most common (). Of the 950 stomachs examined, 183 were empty, giving a vacuity index of 19.26%. The VI of males (14.50%) and females (22.73%) were significantly different (χ2 = 10.08, d.f. = 1, P < 0.05). This percentage did not varied over the year (χ2 = 7.60, d.f. = 3, P > 0.05). Minimum VI was found in spring (14.29%), and maximum of 24.07 during the autumn, while in samples collected in winter and summer percentage of empty stomachs was 19.51% and 20.27% respectively.
Figure 2. Raja radula. Length-frequency distribution of males and females R. radula caught in the Gulf of Gabès.
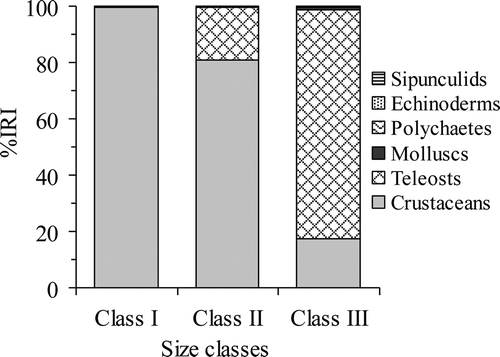
Raja radula was found to prey on a wide variety of invertebrates and teleosts (). The average number of prey and mass per stomach were 2.78 ± 2.75 g and 6.23 ± 8.05 g. Crustaceans were the most frequently observed prey, constituting 63.80% of the total %IRI, followed by teleosts (%IRI = 34.66), while other taxa; cephalopods, gastropods, polychaetes, sipunculids and echinoderms were minor components of the diet (). The majority of preys were considered to be demersal, although pelagic were also found.
Table I. Raja radula. Diet composition of 767 R. radula off the Gulf of Gabès (%F frequency of occurrence; %N percentage in number; %M percentage in mass; %IRI index of relative importance of prey items)
Among crustaceans, decapods occurred most frequently (37.55%) and contributed most to the wet mass (28.49%), while Metapenaeus monoceros (Fabricius, 1798) predominate as a single prey species in terms of occurrence (18.38%), number (20.40%) and mass (16.70%), followed by Sicyonia carinata (Brünnich, 1768). Amphipoda, Isopoda and Mysidacea were important in term of occurrence and number (). The consumed teleosts consist mainly of clupeids and sparids (). Cephalopods were of lower importance in term of occurrence, number and mass ().
Diet shifts
The MANOVA results demonstrated the existence of differences in the diet according to size, season and sex (). However, the ANOVA performed for each dependent variable indicated that there is no difference in the major prey consumed between the sexes and that the minor ones were dependable for the statistically significant of sex (). On the other hand, Morisita diet overlap index between males and females was 0.92, indicating a high level of similarity between their diets.
Table II. MANOVA table of Wilks' lambda and group response to ANOVA. (DF = degrees of freedom; W. L = value of Wilks Lambda; F = approximate F value; H. df = Hypothesis df; E. df = Error df; Sig = significant)
Pelagic and demersal fishes and decapods were responsible for the statistically difference among size classes, but cephalopods and demersal fish were responsible for the difference between season (). The consumption of teleosts increased with increasing size however an inverse trend occurred for crustaceans, which were consumed more frequently by smaller skates (). There was little overlap between class I and class III (CH = 0.17), moderate overlap between class II and class III (CH = 0.42) and a high degree of overlap between class I and class II (CH = 0.83), suggesting a similarity between the diet of classes I and II. On the other hand, the diet overlaps were biologically significant between all seasons (CH > 0.60).
Figure 3. Raja radula. Composition of R. radula diet among size classes based on the percentage index of relative importance (% IRI) values of the major prey groups.
Level trophic and feeding strategy
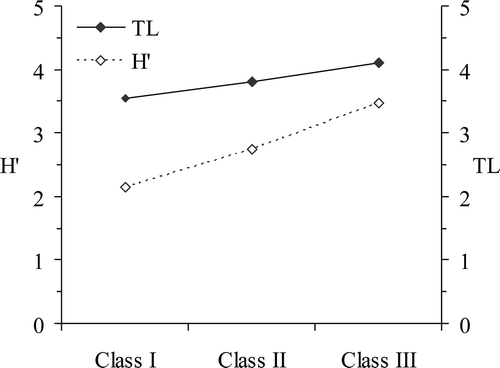
The estimated TL for the whole population of R. radula was 3.91. The trophic level of R. radula increased with increasing body length. Skates of class I and class II were identified as secondary consumers (TL < 4) whereas individuals of class III were tertiary consumers (TL > 4) (). The overall dietary diversity (H') for the entire samples was 3.47. Shannon-Wiener prey diversity values showed increasing prey diversity from classes I to II and II to III ().
Figure 4. Raja radula. Prey diversity (histogram), (Shannon–Wiener index, H') and Trophic level (TL) for each size classes.
The plotting prey-specific abundance against frequency of occurrence (%Pi-%F) showed a progressive change in diet of R. radula with increased size, from one dominated prey consisted of crustacean decapods in the class I to a mixed dietary composed mostly by decapods, amphipods and demersal fishes in class II and class III (). The graphical method indicated a specialization on crustaceans of class I (). The diet of classes II and III consisted of a mixed dietary composed of crustacean decapods and amphipods and teleosts with a dominance of crustaceans in class II and demersal fish in class III (,c). Over 40% of the class II preyed on crustaceans, with 85% Pi, indicating a specialised feeding strategy. On the other hand 30% and 18% of the class III preyed respectively on demersal and pelagic fishes, with 40% Pi, suggesting also a specialised feeding strategy. Raja radula is a specialist feeder with an ontogenetic change on preferred prey from crustaceans to teleosts ().
Figure 5. Raja radula. Feeding strategy and prey importance in the diet of each size class and the all sample of R. radula as determined by the graphical method of Amundsen et al. (Citation1996). (%Pi = Prey-specific abundance; %Fi = frequency of occurrence; a: juveniles, b: subadults, c: adults.)
Discussion
The percentage of empty stomachs of R. radula captured in the Gulf of Gabès as well as in the Gulf of Tunis (VI = 24.64%; Capapé & Azouz Citation1975), was relatively high compared to the percentage on dietary studies of small skates (Smale & Cowley Citation1992; Ellis et al. Citation1996; Scenna et al. Citation2006; Rinewalt et al. Citation2007). The high percentage of empty stomachs may reflect a short period of feeding followed by periods of rapid digestion.
The diet of R. radula in the Gulf of Gabès was dominated by crustaceans representing more than 50% of the total %IRI, and can be classified as a “main food” while the teleosts were secondary preys (Rosecchi & Nouaze Citation1987). Other preys groups were of minor importance and represented incidental food. Although regional differences are observed in prey importance, our results are in agreement with data for specimens from the Gulf of Tunis, Aegean Sea and Egypt water (Capapé & Azouz Citation1975; Abd El-Aziz 1986). Geographic differences in diet have been documented in many elasmobranch species (Yamaguchi & Taniuchi Citation2000; Bethea et al. Citation2006). The differences in the diet between areas are mainly due to different distribution, abundance, density and availability of the prey. Moreover, migratory patterns associated with reproduction are common among elasmobranchs and the occupation of new niches may result in differences in diet (Wetherbee & Cortés Citation2004).
The study shows that in the Gulf of Gabès, as well as in the Gulf of Tunis and Egypt water, decapods were the most abundant prey types among crustaceans in the stomach of R. radula. The dominance of decapods in skate diets has also been demonstrated across geographic regions for several skates (Ebert et al. Citation1991; Orlov Citation2003; Braccini & Perez Citation2005; Dolgov Citation2005).
The stomach contents indicate that this species is an active predator foraging mainly on pelagic and demersal preys while pelagic preys were also found. The ability of skates to hunt in the water column and even reach the sea surface is suggested (Muto et al. Citation2001). However, Beretovski (Citation1989) concluded that skates could not feed in the water column because of their morphology and suggested that pelagic fish traumatized by trawls in the fishing grounds were subsequently preyed upon by skates. The occurrence of pelagic preys in the diet of demersal elasmobranch species may derive from scavenging off the discard of commercial fisheries (Simpfendorfer et al. Citation2001; Saidi et al. Citation2009).
Diets of males and females almost completely overlapped. This indicates that males and females occupy similar areas or encounter similar prey. Similar findings have been reported for other skates (Braccini & Perez Citation2005; Scenna et al. Citation2006; San Martín et al. Citation2007).
Ontogenetic changes observed in this study, have been described for the species and several other skates (Capapé & Azouz Citation1975; Brickle et al. Citation2003).
Ontogenetic shifts in foraging behaviour were observed because small individuals had a specialized diet of crustaceans, medium individuals and large individuals showed specialization on crustaceans and teleosts respectively. These ontogenetic shifts in feeding biology indicate that R. radula has sufficiently plastic foraging behaviour to change from one strategy to another. Diet shift with size is a pattern widely observed in elasmobranchs (Lucifora et al. Citation2009). This shift can be associated with changes in morphometric characteristics, food preference with growth and swimming capacities of the species, as has been observed for other rajids (Farias et al. Citation2006). This shift indicates an ontogenetic shift in diet potentially associated with the energetic requirements of the adults. In terms of diet overlap, adults may rely on differences in the utilization of food resources to avoid intraspecific competition, partitioning the available food resources.
Seasonal variation in the diet of R. radula was noticed in this study. In addition, values of Morisita's index indicated high dietary overlap between seasons. A similar pattern was reported for the species in Egypt water where crustaceans dominated the diet throughout the year (Abd El-Aziz 1986). While, Capapé and Azouz (Citation1975) noticed a seasonal stability in food especially of the zoological groups. The observed seasonal changes in the relative importance of preferred prey probably reflect seasonal environmental changes influencing the distribution and abundance of the main prey (Muto et al. Citation2001; Braccini & Perez Citation2005).
Trophic levels estimated in this study agree well with values reported by Stergiou & Karpouzi (Citation2002) and Ebert & Bizzarro (Citation2007) for the species suggesting that R. radula is a secondary consumer. Nevertheless, the trophic level of R. radula increased with increasing body length and adults are tertiary consumers occupying higher trophic levels (TL > 4) suggesting that they forage on teleosts and crustaceans.
Our results suggest that R. radula is a specialized feeder, crustaceans being the dominant prey of class I and class II, teleosts being the dominant prey of class III. Moreover R. radula has a narrow niche breadth. As in this study, Capapé and Azouz (Citation1975) and Abd El-Aziz (1986) found that large specimens of R. radula were active predators and preyed on alternative prey of crustaceans and teleosts. A generalist predator has a broad dietary niche, whereas the food niche of a specialist is narrow. Many skates are classified as specialist feeders (Ebert et al. Citation1991; Braccini & Perez Citation2005; Scenna et al. Citation2006; San Martín et al. Citation2007), whereas, several others species are considered to be generalist feeders (Smale & Cowley Citation1992; Koen et al. Citation2001). On the other hand, the index of importance and Shannon-Weiner diversity index indicated that a wider variety of prey items were consumed as rays change in size suggesting that large specimens of R. radula are a specialized predator.
Acknowledgements
We are grateful to the commercial fishermen who kindly brought us the catches. Thanks also to the anonymous reviewers for comments that greatly improved the submitted manuscript.
References
- Abdel-Aziz , SH. 1986 . Food and feeding habits of Raja species (Batoidei) in the Mediterranean waters of Alexandria . Bulletin of Institute of Oceanography and Fisheries , 12 : 265 – 276 .
- Abella , AJ and Serena , F. 2005 . Comparison of elasmobranch catches from research trawl surveys and commercial landings at port of Viareggio, Italy, in the last decade . Journal of Northwest Atlantic Fishery Science , 35 : 345 – 356 .
- Amundsen , PA , Gabler , HM and Staldvik , FJ. 1996 . A new approach to graphical analysis of feeding strategy from stomach contents data-modification of the Costello (1990) method . Journal of Fish Biology , 48 : 607 – 614 .
- Berestovosky , EG. 1989 . Feeding habits of Raja radiate and R. fyllae in the Barents and Norwegian Seas . Voprosy Ikhtiologii , 29 : 994 – 1002 .
- Bethea , DM , Carlson , JK , Buckel , JA and Satterwhite , M. 2006 . Ontogenetic and site-related trends in the diet of the Atlantic sharpnose shark Rhizoprionodon terranovae from the Northeast Gulf of Mexico . Bulletin of Marine Science , 78 : 287 – 307 .
- Braccini , JM and Perez , JE. 2005 . Feeding habits of the sandskate Psammobatis extenta (Garman, 1913): sources of variation in dietary composition . Marine and Freshwater Research , 56 : 395 – 403 .
- Brickle , P , Laptikhovsky , V , Pompert , J and Bishop , A. 2003 . Ontogenic changes in the feeding habits and dietary overlap between three abundant rajid species on the Falkland Islands’ shelf . Journal of the Marine Biological Association of the United Kingdom , 83 : 1119 – 1125 .
- Capapé , C and Azouz , A. 1975 . Etude du régime alimentaire de deux raies communes dans le golfe de Tunis: Raja miraletus Linne, 1758 et R. radula, Delaroche, 1809 . Archives de l'Institut Pasteur de Tunis , 52 : 233 – 250 .
- Cavanagh , RD and Gibson , C. 2007 . Overview of the Conservation Status of Cartilaginous Fishes (Chondrichthyans) in the Mediterranean Sea , Vol. vi , 42 Malaga , , Spain : IUCN .
- Cortés , E. 1997 . A critical review of methods of studying fish feeding based on analysis of stomach contents: application to elasmobranch fishes . Canadian Journal of Fisheries and Aquatic Sciences , 54 : 726 – 738 .
- Cortés , E. 1999 . Standardized diet compositions and trophic levels of sharks . ICES Journal of Marine Science , 56 : 707 – 717 .
- Dolgov , AV. 2005 . Feeding and food consumption by the Barents Sea skates . Journal of Northwest Atlantic Fishery Science , 35 : 495 – 503 .
- Ebert , DA and Bizarro , JJ. 2007 . Standardized diet compositions and trophic levels of skates (Chondrichthyes: Rajiformes: Rajoidei) . Environmental Biology of Fishes , 80 : 221 – 237 .
- Ebert , DA , Cowley , PD and Compagno , LJV. 1991 . A preliminary investigation of the feeding ecology of skates (Batoidea: Rajidae) off the west coast of southern Africa . South African Journal of Marine Science , 10 : 71 – 81 .
- Ellis , JR , Pawson , MG and Shackley , SE. 1996 . The comparative feeding ecology of six species of shark and four species of ray (Elasmobranchii) in the north-east Atlantic . Journal of the Marine Biological Association of the United Kingdom , 76 : 89 – 106 .
- Farias , I , Figueiredo , I , Moura , T , Gordo , LS , Neves , A and Serra-Pereira , B. 2006 . Diet comparison of four ray species (Raja clavata, Raja brachyura, Raja montagui, and Leucoraja naevus) caught along the Portuguese continental shelf . Aquatic Living Resources , 19 : 105 – 114 .
- Koen , AM , Crespo , EA , Garcia , NA , Pedraza , SN , Mariotti , PA , Beron , VB and Mora , NJ. 2001 . Food habits of Dipturus chilensis (Pisces: Rajidae) off Patagonia, Argentia . ICES Journal of Marine Science , 58 : 288 – 297 .
- Krebs , CJ. 1989 . Ecological methodology , New York : Harper Collins .
- Kyrtatos , NA. 1982 . Investigation on fishing and biology of the most important fishes of the region around the Aegean Sea, Island of Tinos 88 Thalassographica 5(specl. publ.)
- Langton , RW. 1982 . Diet overlap between Atlantic cod, Gadus morhus, silver hake, Merluccius bilinearis, and fifteen other northwest Atlantic finfish . Fishery Bulletin , 80 : 745 – 759 .
- Lucifora , LO , Garcia , VB , Menni , RC , Escalante , AH and Hozbor , NM. 2009 . Effects of body size, age and maturity stage on diet in a large shark: ecological and applied implications . Ecological Research , 24 : 109 – 118 .
- Muto , EY , Soares , LSH and Goitein , R. 2001 . Food resource utilization of the skates Rioraja agassizii (Muller & Henle, 1841) and Psammobatis extenta (Garman, 1913) on the continental shelf off Ubatuba, southeastern Brazil . Revista Brasileira de Biologia , 61 : 217 – 238 .
- Orlov , AM. 2003 . Diets, feeding habits and trophic relations of six deep-benthic skates (Rajidae) in the western Bering Sea . Journal of Ichthyology & Aquatic Biology , 7 : 45 – 60 .
- Pinkas , LM , Oliphant , S and Iverson , ILK. 1971 . Food habits of albacore, bluefin tuna, and bonito in Californian waters . California Fish and Game , 152 : 1 – 105 .
- Rinewalt , CS , Ebert , DA and Cailliet , GM. 2007 . Food habits of the sandpaper skate, Bathyraja kincaidii (Garman, 1908) off central California: seasonal variation in diet linked to oceanographic conditions . Environmental Biology of Fishes , 80 : 147 – 163 .
- Rosecchi , E and Nouazé , Y. 1987 . Comparaison de cinq indices alimentaires utilisés dans l'analyse des contenues stomacaux . Revue des Travaux de l'Institut des Pêches Maritimes , 49 : 111 – 123 .
- Saidi , B , Enajjar , S , Bradai , MN and Bouain , A. 2009 . Diet composition of smooth-hound shark, Mustelus mustelus (Linnaeus, 1758), in the Gulf of Gabès, southern Tunisia . Journal of Applied Ichthyology , 25 : 113 – 118 .
- San Martín , MJ , Braccini , JM , Tamini , LL , Chiaramonte , GE and Perez , JE. 2007 . Temporal and sexual effects in the feeding ecology of the marbled sand skate Psammobatis bergi Marini, 1932 . Marine Biology , 151 : 505 – 513 .
- Scenna , LB , García de la Rosa , SB and Díaz de Astarloa , JM. 2006 . Trophic ecology of the Patagonian skate, Bathyraja macloviana, on the Argentine continental shelf . ICES Journal of Marine Science , 63 : 867 – 874 .
- Serena , F , Mancusi , C and Barone , M , eds. 2010 . “ Field identification guide to the skates (Rajidae) of the Mediterranean Sea ” . In Guidelines for data collection and analysis. Biologia Marina Mediterranea Vol. 17:204 ,
- Simpfendorfer , CA , Goodreid , AB and McAuley , RB. 2001 . Diet of three commercially important shark species from Western Australian waters . Marine and Freshwater Research , 52 : 975 – 985 .
- Smale , MJ and Cowley , PD. 1992 . The feeding ecology of skates (Batoidea: Rajidae) off the cape south coast, South Africa . South African Journal of Marine Science , 12 : 823 – 834 .
- Stergiou , KI and Karpouzi , VS. 2002 . Feeding habits and trophic levels of Mediterranean fish . Reviews in Fish Biology and Fisheries , 11 : 217 – 254 .
- Wetherbee , BM and Cortés , E. 2004 . “ Food consumption and feeding habits ” . In Biology of Sharks and their Relatives , Edited by: Carrier , JC , Musick , JA and Heithaus , MR . 225 – 246 . New York : RC Press .
- Yamaguchi , A and Taniuchi , T. 2000 . Food variations and ontogenetic dietary shift of the starspotted-dogfish Mustelus manazo at five locations in Japan and Taiwan . Fisheries Sciences , 66 : 1039 – 1048 .