Abstract
In this paper the larval morphology of the Palaearctic Meloe subgenera Eurymeloe and Coelomeloe is revised. We describe the first instar larva of Meloe (Eurymeloe) apenninicus, endemic to the Italian peninsula and, comparatively, triungulins of three more species of the subgenus Eurymeloe: M. apivorus, M. ganglbaueri, misidentified in literature, and of M. mediterraneus, incompletely described. Descriptions are accompanied by scanning electron microscope (SEM) photos of diagnostic characters and by a table reporting the size of the morphological details of 11 Euro-Mediterranean species and of M. (Coelomeloe) tuccius. The taxonomic position of the subgenus Coelomeloe and its distinction from Eurymeloe are discussed on the basis of larval morphology. Taxonomical problems concerning some Eurymeloe species are also discussed. A key to first instar larvae of all known Eurymeloe and Coelomeloe triungulins is provided.
Keywords:
Introduction
Family Meloidae (Insecta: Coleoptera) is well known in the biological and medical literature due to its unusual larval development related to its parasitic habits on Hymenoptera Aculeata and Orthoptera Caelifera, and the cantharidin production, a natural terpenoid with several pharmacological applications (see Bologna Citation1991; Bologna et al. Citation2010). Phylogenetic relationships of blister beetles were recently clarified by Bologna et al. (Citation2008). The presence of a peculiar modality of larval development, called hypermetamorphosis or hypermetaboly, is common to all Meloidae from subfamilies Nemognathinae and Meloinae, with the probable exception of the basal subfamily Eleticinae, that may have predator rather than parasitoid larvae (Pinto et al. Citation1996; Bologna et al. Citation2001, Citation2008; Bologna & Di Giulio Citation2011). The development of the most derived subfamilies includes seven distinct larval instars, often divided into five morphologically and biologically distinct phases (Paoli Citation1932; Selander & Mathieu Citation1964); the first one, usually called triungulin, locates the host (or eventually the prey), walking actively on the ground (most Meloinae) or is phoretic on wild bees or other insects (most Nemognathinae and some Meloinae) (see Bologna Citation1991; Di Giulio et al. Citation2003; Bologna & Di Giulio Citation2011 for reviews).
The morphology of the first instar larva was largely used in the systematics of this family to evidence phylogenetic relationships at different taxonomic levels (Bologna & Pinto Citation2001; Bologna et al. Citation2008). Phoretic behaviour and relative morpho-functional adaptations are common to both Nemognathinae and Meloinae, and the hypothesis of convergent evolution was advanced and supported (Bologna & Pinto Citation2001; Bologna & Di Giulio Citation2011). In particular, according to molecular and morphological evidence, phoresy evolved multiple times in different tribes of the subfamily Meloinae (Bologna & Pinto Citation2001; Bologna et al. Citation2008).
The meloine genus Meloe Linnaeus, 1758 (tribe Meloini) includes some 150 Holarctic and Afrotropical species. The first larval instars of all of them are adapted to phoresy on bees, with different degrees of specialization at the level of antennal sensorium, head setation, mandibles, femurs, claws, etc. (Bologna & Pinto Citation1992, Citation1995, Citation1998, Citation2001; Pinto & Bologna Citation1993; Di Giulio et al. Citation2002; Bologna & Di Giulio Citation2011). In particular, the antennal sensorium, a chemoreceptive structure that in Coleoptera larvae seems to be involved in prey/host finding (Giglio et al. Citation2008), is reduced in species that are highly adapted to phoresy, since they do not look for the host species (Pinto & Bologna Citation1993).
The first instar larvae of about 30 species belonging to 11 of the 16 subgenera have been described, and the first instar larvae of the subgenera are identifiable (reviews in Bologna & Pinto Citation1998; Di Giulio et al. Citation2002). Larval studies were relevant for the systematics of some genera; e.g., triungulin morphology suggested moving some genera, such as Physomeloe Reitter, 1911, Berberomeloe Bologna, Citation1989, and Trichomeloe Reitter, 1911 out of the genus Meloe, and confirmed the subgeneric division of the genus, in most cases based on adult morphology (Bologna Citation1989; Bologna & Aloisi Citation1994; Bologna & Di Giulio Citation2008).
Eurymeloe Reitter, 1911 is the most speciose subgenus of Meloe, with ca. 50 mostly Palaearctic species (exceptions from northern Somalia and from the Aleutinian islands; Bologna Citation1988). Two distinct groups of species, namely the “brevicollis” and “rugosus” groups, have been identified on adult morphology (Bologna Citation1988), but here we question the validity of these groups and the possible inclusion of the monotypic subgenus Coelomeloe Reitter, 1911 in Eurymeloe, as a third group, based on larval morphology (Selander Citation1985; Bologna et al. Citation1989; Bologna & Pinto Citation2001). Larvae of few species of Eurymeloe have been described so far, some of these after unreliable identification (reviews in Bologna Citation1988, and Bologna et al. Citation1989).
A larval synopsis of the subgenus Eurymeloe is the main object of this contribution. In detail, the aims of this study are to: (i) describe the triungulin of Meloe apenninicus Bologna, Citation1988, endemic to the Italian peninsula; (ii) describe comparatively the triungulin of M. apivorus Reitter, 1895, a controversial taxon, endemic to High Atlas (Morocco) and previously referred as subspecies to M. affinis Lucas, 1847; (iii) compare the triungulin of apivorus Reitter, 1895 against that of affinis Lucas, 1847 (var. setosus Escherich, Citation1890); (iv) describe comparatively the first instar larvae of two Euro-Mediterranean species, M. ganglbaueri Apfelbeck, 1905 and M. mediterraneus G. Müller, 1925, both poorly described; (v) assess the taxonomic status of Coelomeloe and the proposed synonymy with Eurymeloe; (vi) provide a key to all known first instar larvae of Eurymeloe and Coelomeloe (12 Euro-Mediterranean species) and a comparative table with the main measures of body and appendices of these species.
Materials and methods
The description of triungulins and the key of identification are based on the following material preserved in 70% ethanol and/or mounted on slides, if not differently indicated larvae and adults are kept at Roma Tre University, Rome, Italy, in the M. A. Bologna collection. Sampled localities are ordered by country, from north to south and from west to east. The key was built basing both on direct study of specimens and on descriptions available in the literature (Bologna Citation1988; Bologna et al. Citation1989; Lückmann & Kuhlmann Citation1997; Di Giulio et al. Citation2002; Lückmann & Scharf Citation2004); when contrasting characters were found, direct observation was considered more reliable.
| i. | Meloe (Eurymeloe) affinis Lucas, 1847 (var. setosus Escherich, Citation1890)
| ||||||||||||||||||||||||||||||||||||||||||||||
| ii. | Meloe (Eurymeloe) apenninicus Bologna, Citation1988
| ||||||||||||||||||||||||||||||||||||||||||||||
| iii. | Meloe (Eurymeloe) apivorus Reitter, 1895
| ||||||||||||||||||||||||||||||||||||||||||||||
| iv. | Meloe (Eurymeloe) brevicollis Panzer, 1773
| ||||||||||||||||||||||||||||||||||||||||||||||
| v. | Meloe (Eurymeloe) corvinus Marseul, 1876
| ||||||||||||||||||||||||||||||||||||||||||||||
| vi. | Meloe (Eurymeloe) flavicomus Wollaston, 1854
| ||||||||||||||||||||||||||||||||||||||||||||||
| vii. | Meloe (Eurymeloe) ganglbaueri Apfelbeck, 1905
| ||||||||||||||||||||||||||||||||||||||||||||||
| viii. | Meloe (Eurymeloe) glazunovi Pliginskji, 1910
| ||||||||||||||||||||||||||||||||||||||||||||||
| ix. | Meloe (Eurymeloe) mediterraneus G. Müller, 1925
| ||||||||||||||||||||||||||||||||||||||||||||||
| x. | Meloe (Eurymeloe) rugosus Marsham, 1802
| ||||||||||||||||||||||||||||||||||||||||||||||
| xi. | Meloe (Eurymeloe) scabriusculus Brandt and Ericshon, 1832
| ||||||||||||||||||||||||||||||||||||||||||||||
| xii. | Meloe (Coelomeloe) tuccius, Rossi, 1792
| ||||||||||||||||||||||||||||||||||||||||||||||
Adults were reared at 24–25°C in a thermostatic chamber with photoperiod control; females oviposited in sand provided as a substrate in rearing boxes, and their eggs were kept till hatching.
Morphological analysis was performed under an Olympus BX51 light microscope equipped with Colorview II camera (SIS) and Cell*D image analysis software (SIS) to examine and photograph the cleared specimens mounted on slides in Canada balsam, and a FEI dualbeam FIB/SEM Helios Nanolab as SEM for material mounted on stubs after critical point dehydration and gold sputtering.
Terminology of larval structures follows MacSwain (Citation1956), Lawrence (Citation1991), and Bologna & Pinto (Citation2001). For some traits of larval chaetotaxy we followed notational conventions as suggested by Selander (Citation1990).
Results
Description of the first instar larva of Meloe (Eurymeloe) apenninicus. Measurements in Triungulin campodeiform (–a2, 4e); body elongate, fusiform, about 3.8 times as long as maximum width (on mesothorax). Colour light brown; head and legs brown, with distinctly irregular reticulate microsculpture (–a2); mandibles and area surrounding stemmata strongly sclerotized, dark brown (). Tergites and sternites of thorax and abdomen well sclerotized with reticulate microsculpture with transversely imbricated polygonal mesh (–a1); posterior meshes of sterna and laterotergites with short, spine-like backward prolongations that provide a microserrate aspect to posterior margins.
Table I. Main sizes (in mm) of the Meloe (Eurymeloe) first instar larvae mounted on slides and examined with a light microscope. Abbreviations: A1–A3 = length of antennomeres I–III; ABL = length of abdomen; ASL = length of apical antennal seta on A3; BL = body length; ESL = epicranial stem length; HL = head length; HW = head width; N. = number of examined specimens; SD = diameter of stemmata; PL = prothorax length; PW = prothorax width; SPA = abdominal spiracle maximum width; SPM = mesothoracic spiracle maximum width; TSL = length of terminal abdominal setae
Figure 1. First instar larvae of Meloe (Eurymeloe) (a) apenninicus, (b) apivorus, (c) ganglbaueri and (d) mediterraneus: habitus in (a–d) dorsal, (a1–d1) right lateral and (a2–d2) ventral view.

Figure 2. First instar larvae of Meloe (Eurymeloe) (a) apenninicus, (b) apivorus, (c) ganglbaueri and (d) mediterraneus: head in (a–d) dorsal, (a1–d1) right lateral and (a2–d2) ventral view; (a3–d3) right antenna in anterior view.
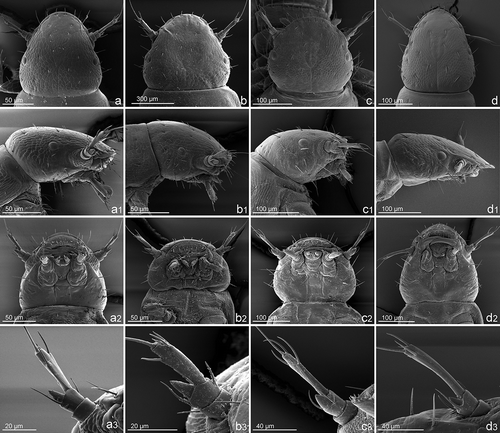
Figure 3. First instar larvae of Meloe (Eurymeloe) (a) apenninicus, (b) apivorus, (c) gangbaueri and (d) mediterraneus: (a–d) pro- and mesonotum (dorsal view), (a1–d1) abdominal tergites I–III (left lateral view) and (a2–d2) claws (midlegs).

Figure 4. Images of slide-mounted first instar larvae of the following Meloe (Eurymeloe) species: (a) brevicollis, (b) corvinus, (c) mediterraneus, (d) affinis, (e) apenninicus, (f) apivorus, (g) flavicomus, (h) ganglbaueri, (i) glazunovi, (j) rugosus, (k) scabriusculus; and (l) Meloe (Coelomeloe) tuccius. Scale bar: 0.2 mm.

Head (–a2) subtriangular, slightly transverse (L/W = 0.9), widest just behind the posterior margin of stemmata; sides broadly curved from stemmata to occipital foramen, straight and convergent anteriorly (); basal elevation absent; anterior margin of head rounded. Epicranial suture Y shaped; frontal arms straight, slightly diverging at base (angle about 35°), markedly curved laterally to antennae at distal third; epicranial stem short (0.04 mm). One stemma on each side ( ); stemmata round, distinctly convex, dorsolaterally placed (, ). Frontoclypeal region with 14 setae (); apex of frontoclypeus with one transverse row (frontoclypeal row, FCR1–3) of three pairs of setae, FCR3 longer than others; one sensory pit between FCR1 and FCR2; one medial sensory pit between pair of setae FCR1; four pairs of setae posterior to FCR, one directly behind FCR2 and three following a curved line paralleling the arms of the epicranial suture (from posterior to anterior setae 1–3); one sensory pit present between setae 1 and 2. Each epicranial plate dorsally with a basal group of four minute setae, eight longer setae mostly placed laterally around the stemma and three sensory pits (one medial, one lateral and one ocular distinctly anterior to stemma). Labrum ( ) directed ventrally, transverse, anteriorly rounded and posteriorly straight, with nine pairs of setae of varying length (two lateral very long), one single medial seta and one pair of pits. Antennae anterolaterally directed (, a3); antennomere I short, ring-like, slightly asymmetrical with one dorsal sensory pit; segment II slightly longer than I, with three long apical setae (one dorsal, one lateral, one ventral), one minute seta (outer side) near sensory appendix and one dorsolateral pit; sensory appendix on antennomere II well developed, conical, lateroventral (outer side), as long as antennomere I ( ); antennomere III slender and long, straight, sub-clavate, compressed laterally, about 1.8 times longer than II, with a long apical seta, three long subapical setae, two lateral (one on outer and one on inner sides) and one dorsal in position, one minute seta near the base of apical seta; one lateral and one sub-basal pit. Mandibles with broad and prominent base, apical half narrowing and abruptly bending inward, ental surface smooth; partially covered by labrum in the resting position ( ); outer margin of mandible with two setae, one sensory pit between them and one pit mesodorsally. Maxillae with stipes rectangular with four setae; mala simple, lobiform, slightly protruding, with four to five spiniform setae, one longer than the others; cardo transverse, small, with minute seta; maxillary palpi with joints as follows: I extremely short, with one ventral pit; II slightly longer than I, with two ventrolateral subequal setae, one on the external and one on the internal side; III with the external side widened from base to apex, about three times the length of I and II together, asymmetrical (lateral side longer than medial side), with one dorsolateral seta (inner side); apex of segment III obliquely truncate, with a large, sensorial area composed of one apical, stick-like sensory appendix and surrounded about 10 shorter papillae; outer side with one slender digitiform sensillum. Gulamentum weakly sclerotized with two elongate anterior setae. Prementum fused with mentum with one pair of basal setae lined by one pair of external sensory pits; one pair of ligular setae present between insertion of palpi; labial palpi ( ) small, with segment I short, II cylindrical, twice longer than I, with one moderately long, stick-like, apical sensory appendix, inserted in a prominent base and surrounded by five to seven shorter subapical papillae.
Thorax () with segments transverse and subrectangular, broader than head; prothorax as wide as meso- and metathorax; margins of pronotum convergent anteriorly, margins of meso- and metanotum slightly rounded. Ecdysial line well marked, complete on pro- and mesonotum, vestigial on metanotum. Pronotum about 2.3 times wider than long, each half of notum with 13 setae and five pits symmetrically placed along three transverse, subparallel rows; anterior row (AR) with five setae and two pits; medial row (MR) with three setae; posterior row (PR) with five setae and two to three pits; prosternum with three pairs of medial setae the two anterior pairs similar in length, posterior pair much longer than others. Mesonotum slightly shorter than pronotum; AR with two setae grouped with one pit mesodorsally, two setae laterally, anterior to the spiracle. MR with three to four setae and one shorter seta near the spiracle; PR with five setae and two pits; three pairs of medial setae on mesosternum, the anterior pair extremely short. Setae of metathorax similar in number, position and relative dimensions to those of mesothorax.
Legs elongate (): coxa short and broad, with four elongate setae, transversally arranged, four to five minute basal setae and one pit; trochanter with two moderately long setae and four to six pits; femur with six setae and one pit, the longest ventral femoral seta, much shorter than femur; tibiae and claws increasing in length from pro- to metathorax; tibiae stout, tapered at apex and with four longitudinal rows of five to six moderately long setae; claw ( ) conicofalcate, thin, acute and slightly curved at apex, with two long setae of slightly different length inserted almost at the same level near base.
Spiracles. Marginal ring (peritreme) of mesothoracic and I abdominal distinctly protruding. Mesothoracic spiracle elliptical, obliquely placed on laterotergite. Abdominal spiracle I ( ) dorsally placed on laterotergite, transversely elliptical; abdominal spiracles II–VIII small, round, subequal and scarcely protruding, laterally positioned, placed on laterotergites.
Abdomen (, ) fusiform with transverse, rectangular tergites; maximum width at segment III; sternites entire ( ). Mediotergites with two transverse rows (MR posteriorly displaced) of setae – their arrangement on each half of tergites as follows: AR with two minute setae (four on tergite I, two medial and two lateral) and one pit, MR + PR with nine elongate setae and one pit; tergite IX with one elongate pair of setae on PR (caudal setae) which are less long then the last four abdominal segments combined. AR of each sternite with one pair of minute seta, MR absent and PR with eight elongate setae, each slightly different in length. Sternite IX with two small seta on AR; six long setae and one pit on PR. Abdominal apex (segment X or pygopod) apically membranous, transversally divided in two parts: dorsal part semicircular with six extremely small setae transversally arranged, ventral part longitudinally divided in two lobes ( ), moderately produced, with two pairs of setae along posterior margin.
Comparative description of the first instar larva of Meloe (Eurymeloe) apivorus. Measurements in –b2, 2b–b3, 3b–b2, 4f. Body slender, about 4.8 times as long as maximum width (on mesonotum), subparallel sided, with abdomen elongate, only slightly fusiform in dorsal view (, ) but very thick in lateral view ( ). Head short, much wider than long (L/W = 0.8) with subbasal maximum width. Antennomere III 2.6 times longer than II. Abdomen maximum width at segment IV.
Comparative description of the first instar larva of Meloe (Eurymeloe) ganglbaueri. Measurements in –c2, 2c–c3, 3c–c2, 4h. Body about four times as long as maximum width (on mesothorax). Head globose, suboval (, c2), much wider than long (W/L = 0.87), with sides protruding just behind the stemmata (, ). Antennomere III about 1.9 times longer than II. Prosternum with three pairs of medial setae increasing in length from anterior to posterior, anterior pair much shorter than medial pair. Legs distinctly short (, c1, 4h). Abdomen maximum width at segment V (, ).
Comparative description of the first instar larva of Meloe (Eurymeloe) mediterraneus. Measurements in –d2, 2d–d3, 3d–d2, 4c. Body about 4.7 times as long as maximum width (on abdominal segments III–IV). Colour yellow, slightly darker on head and legs. Head deeply sculptured (–d1) with subhexagonal meshes, suboval (, ), longer than wide (L/W = 1.14), widest just behind stemmata, at middle head side; sides slightly curved from stemmata to occipital foramen. Frontal arms of the epicranial suture bisinuate, widely diverging at base (angle about 60°); epicranial stem very long (average of four specimens = 0.086 mm), about as long as the antenna. Stemmata extremely convex and protruding (–d1). One pair of sensory pits at the level of setae FCR2; one medial seta between FCR1. Ocular setae at the level of anterior margin of stemmata. Antenna ( ) with sensory appendix on segment II small; antennomere III very long, cylindrical, about 3.6 times longer than II, slightly curved posteriorly. Maxillae with stipes bearing five setae and one pit; maxillary palpomeres I and II short, ring-like, subequal in length, III about two times the length of I and II together. Legs elongate (). Abdominal spiracle I suboval, distinctly protruding (). Abdominal tergites with three transverse rows of setae, their arrangement on each half of tergites as follows: AR with two minute setae (four on tergite I) and one pit, MR with two setae, PR with seven setae. Caudal setae slightly longer than the last two abdominal segments.
Key to the first instar larvae of Eurymeloe and Coelomeloe species.
The following key to the species is based on the careful observation of all available larvae (see material and method), except M. (E.) murinus, for which we refer to the descriptions of Cros (Citation1912, Citation1917, Citation1919, Citation1929).
| 1. | Abdominal spiracle VIII () ca. four times greather than II–VII, slightly wider than I. Periocular area black (), the remaining head surface, prothorax and urites II–V dark brown; meso-methatorax and urites VI–IX light brown.…………… M. (E.) affinis (var. setosus) | ||||
| 2. | Abdominal spiracle VIII small, similar in size (or just slightly wider) to others, except I. Body colouration not as above2 | ||||
| 3. | Head evidently longer than wide (L/W > 1.1); templa subparallel or only slightly widened3 | ||||
| 4. | Head slightly longer or about as long as wide (< 1.1); templa evidently rounded and widened.4 | ||||
| 5. | Abdominal spiracle I transverse; greatest width of head sub-basal.…………… M.(E.) brevicollis and M.(E.) corvinus | ||||
| 6. | Abdominal spiracle I suboval; greatest width of head at middle. M. (E.) mediterraneus | ||||
| 7. | Abdominal spiracle I very large and transverse, about 1/3 the entire tergite width…………… M. (C.) tuccius | ||||
| 8. | Abdominal spiracle I suboval, only slightly transverse.5 | ||||
| 9. | Abdominal spiracle VIII larger than II–VI (), VI just slightly larger.…………… M. (E.) scabriusculus | ||||
| 10. | Abdominal spiracle VIII as large as II–VII.6 | ||||
| 11. | Prothorax campaniform (), sides roundly converging in front, fore width clearly shorter than posterior7 | ||||
| 12. | Prothorax subrectangular, sides subparallel, fore width subequal or slightly shorter than posterior9 | ||||
| 13. | Ratio L/W of head = 0.9. Abdominal spiracle I subequal in size to mesothoracic one. Maximal abdominal width on urite III…………… M. (E.) apenninicus | ||||
| 14. | Ratio L/W of head ≥ 1.0. Abdominal spiracle I similar or wider than mesothoracic one. Abdominal width not as above8 | ||||
| 15. | Maximal abdominal width on urite II. Abdominal spiracle I evidently wider than mesothoracic one M. (E.) flavicomus | ||||
| 16. | Maximal abdominal width on urite IV. Abdominal spiracle I evidently similar to mesothoracic one…………… M. (E.) murinus (from description) | ||||
| 17. | Head very short (L/W ratio: 0.80–0.87)10 | ||||
| 18. | Head more elongate (L/W ratio: 0.90–0.97)11 | ||||
| 19. | Maximal abdominal width on urite V, abdomen swollen in lateral view ( )…………… M. (E.) ganglbaueri | ||||
| 20. | Maximal abdominal width on urite IV, abdomen slender in lateral view ( )…………… M. (E.) apivorus | ||||
| 21. | Maximal width of head at base (). Setae on all body short and thin…………… M. (E.) rugosus | ||||
| 22. | Maximal width of head at middle of templa (). Setae on all body robust and spiniform M. (E.) glazunovi | ||||
A relevant incongruence exists between our description of M. (C.) tuccius first instar larva and that of Cros (Citation1929), both based on specimens hatched from eggs oviposited by reared females. The shape of the abdominal spiracle I was considered as rounded as the others by Cros (Citation1929), while in all triungulins we examined (Canary islands, Sardinia, and Lampedusa, an Italian island part of the Maghrebian platform) it is clearly wide and transverse ().
Remarks concerning M. affinis, M. ganglbaueri and M. mediterraneus were discussed above.
Discussion
The present study, based on the comparison of rich material of different taxa, clarifies some taxonomical problems of this complex subgenus of Meloe.
| a. | Selander (Citation1985) elevated Eurymeloe to genus level and considered Coelomeloe and Meloegonius Reitter, 1911 as synonyms. Bologna (Citation1988), Bologna et al. (Citation1989), and Bologna & Pinto (Citation2001) considered the three taxa as subgenera of Meloe, emphasizing the close similarity between Eurymeloe and Coelomeloe, and questioned their synonymy. According to present analysis of the larvae of all available species of Eurymeloe and Coelomeloe, the differences of Coelomeloe suggest to keep these subgenera separate. Actually, the subgenera are characterized only by plesiomorphic traits of antennae (with conical antennal sensorium), unmodified claws, legs and larvae. In detail, the first instar larvae of M. (C.) tuccius differ from Eurymeloe because of: (i) head strongly sclerotized, large compared to body size; (ii) basal part of frontal sclerite extremely thin, frontal sutures not sinuate, only slightly diverging at base; (iii) stemmata large; (iv) mandibles ventral, subtriangular, extremely short, with wide base; (v) thoracic tergites distinctly decreasing in width and length from pro- to metathorax; (vi) mesothoracic spiracle clearly protruding; (vii) abdomen short compared to body size; (viii) abdominal spiracle I very large and transverse, about as wide as one third of the tergite; (ix) setae of body strong and spiniform. It is worth noting, however, that spiracles modified as in M. (C.) tuccius (vi, viii), though at a much lesser degree, are present also in M. (E.) brevicollis and M. (E.) corvinus, suggesting parallel biological adaptations. | ||||
| b. | Selander (Citation1985) excluded affinis, ganglbaueri and mediterraneus from Eurymeloe as “atypical Meloe”, because of their widened head. Actually, from Bologna and Pinto (Citation2001) and from present data, these three species belong to Eurymeloe and the shape of the head of this subgenus is remarkably variable. In particular, M. (E.) affinis var. setosus appears to be clearly distinct because of the apomorphic condition of abdominal spiracle VIII, extremely widened and transverse (about four times the size of II–VII) and not small as in all Eurymeloe (only in scabriusculus it is slightly wider than II–VII). Moreover, this triungulin is the only Meloe with a bicoloured abdomen, similarly to some apterous and brachypterous Lyttini genera (Berberomeloe Bologna, Citation1989; Trichomeloe Reitter, 1911: see Bologna Citation1989; Bologna & Di Giulio Citation2008), or to several Mylabrini (MacSwain Citation1956; Di Giulio & Bologna, unpublished). According to Cros (Citation1936: 192) this triungulin is not phoretic but probably fossorial, but this hypothesis needs confirmation. The possibility to define the exact identification of the studied species (see below), and to study larval behaviour and molecular traits, could permit to better clarify the taxonomic position of this meloid beetle. The variety setosus was separated because of the presence of light yellow isolated setae along the posterior margin of abdominal tergites, the presence of a constant frontal furrow, the distinct elytral sculpture and the smaller size. Escherich (Citation1890: 90, note 5) considered the possibility to ascribe it to a different species. Actually, this taxon was never revised and it could represent a distinct species, also suggested by Peyehrimhoff (in Cros Citation1934), who observed the specimens from western Algeria (Sidi Bel Abbès, Bonnier) studied by Cros (Citation1934, Citation1936). The possibility to compare triungulins of affinis var. setosus (studied by Cros (Citation1934, Citation1936), with those of apivorus, both obtained by eggs, permits us to resolve partially the taxonomic problem of the affinis-complex. In fact, Bologna (Citation1988) considered affinis as polytypic, including four subspecies: apivorus (High Atlas; described as distinct species), schatzmayri Bologna, Citation1988 (Tripolitania), simillimus Escalera, 1914 (Middle Atlas; described as distinct species), and the nominate subpecies (northern Tunisia, Algeria and Morocco). The differences among subspecies are evident in adults (Bologna Citation1988, Citation1991), except between apivorus and simillimus, which probably represent a single species (Bologna Citation1988). The great differences between the first instar larvae of affinis var. setosus and apivorus do support the specific value of apivorus. The great difference of the spiracle VIII of affinis var. setosus vs. that of all other Eurymeloe probably represents an automorphy of this taxon, versus all Meloe and other meloine larvae. According to Cros (Citation1936: 189–190), this spiracle is adapted to emerge from the liquid honey, similarly to most Nemognathinae first instar larvae with differently conically modified abdominal spiracle VIII (Bologna Citation1991). Like all other Eurymeloe of the rugosus group (sensu Bologna Citation1988), adults of this species are mostly nocturnal and were collected only under stones (Cros Citation1934, Citation1936; Bologna Citation1988). | ||||
| c. | According to larval morphology, taxonomical distinction between rugosus and apenninicus is confirmed. The latter species is endemic to the Italian peninsula and is geographically vicariant to former species. Adults of the two species are very similar and the possibility of co-specificity was supposed (Bologna Citation1988), but triungulins differ primarily because of the head size and by some morphometric features (). | ||||
| d. | In this paper we describe and figure the first instar larva of ganglbaueri for the first time. In fact, the description made by Cros (Citation1943) is based on triungulins from Oujda (eastern Morocco), but ganglbaueri is not distributed in North Africa (Bologna Citation2008). In the Oujda area, the only species of Eurymeloe with well visible dorsal setae is otini Peyerimhoff, 1949, which was recently synonymized with saharensis Chobaut, 1898 (Ruiz et al. Citation2010). The triungulin from Oujda could belong to saharensis or to another undescribed species. Also the re-description of mediterraneus, supported by figures, allows us to recognize definitively this species, previously inadequately described (Cros Citation1922, Citation1935, Citation1943). | ||||
| e. | Another taxonomic inference emerging from the present study concerns the definition of groups of species among Eurymeloe. Bologna (Citation1988), studying adult morphology, tried to divide this subgenus into two groups, the brevicollis and the rugosus groups, the latter split in two subgroups (rugosus and murinus). Actually, this division is not supported by larval morphology. In fact, mediterraneus, previously included in the rugosus group, shows some similarities with species of the brevicollis group such as brevicollis and corvinus (see –c). On the contrary, scabriusculus (), previously ascribed to the brevicollis group, seems more similar to glazunovi () and other species close to rugosus. Of course, affinis, referred to the rugosus group, must be ascribed to a separate group. | ||||
| f. | Finally, from our analysis, the first instar larvae of brevicollis () and corvinus () appear to be almost identical and we suspect a co-specificity of these two taxa. The brevicollis-complex, as evidenced by Bologna (Citation1988, 1991) includes several species, such as corvinus, algiricus Escherich, Citation1890, austrinus Wollaston, 1854, curticornis Escalera, 1914, ibericus Reitter, 1895, and others that could represent only subspecies of brevicollis, distinct by phenetic features of colour and body sculpture. | ||||
Acknowledgements
This research was supported by a fund of the Italian Ministry for Instruction, University and Research (Grant No. PRIN 20085YJMTC). We wish to thank the following entomologists, most Italian, for providing us with specimens or for their help during our field research: Paolo A. Audisio (Roma), Maurizio Biondi (L'Aquila), Giuseppe M. Carpaneto (Roma), Achille Casale (Sassari), Pierfilippo Cerretti (Verona), Pier Mauro Giachino (Torino), Teiji Kifune (Fukuoka, Japan), Johannes Lückmann (Bensheim, Germany), Carla Marangoni (Roma), M. Angeles Marcos-García (Salamanca, Spain), Emanuela Maurizi (Roma), Maurizio Mei (Roma), Pier Francesco Murgia (Quartucciu, Cagliari), B. Giuseppe Osella (Verona), John D. Pinto (Waldport, OR, USA), Augusto Vigna Taglianti (Roma), Marzio Zapparoli (Viterbo). Thanks to the anonymous referee and the Editor for their suggestions.
References
- Bologna , MA. 1988 . Note su Eurymeloe e revisione delle specie euromediterranee del gruppo rugosus (Coleoptera, Meloidae) . Fragmenta Entomologica , 20 : 233 – 301 .
- Bologna , MA. 1989 . Berberomeloe, a new West Mediterranean genus of Lyttini for Meloe majalis Linné (Coleoptera, Meloidae). Systematics and bionomics . Bollettino di Zoologia , 55 ( 1988 ) : 359 – 366 .
- Bologna , MA. 1991 . Coleoptera Meloidae. Fauna d'Italia. XXVIII , Bologna : Calderini .
- Bologna , MA. 2008 . “ New nomenclatorial and taxonomic acts, and comments, Meloidae; family Meloidae Gyllenhal, 1810 ” . In Catalogue of Palaearctic Coleoptera, 5 , Edited by: Löbl , I and Smetana , A . 45 – 49 . 370 – 412 . Apollo Books : Stenstrup .
- Bologna , MA and Aloisi , G. 1994 . Systematics and bionomics of Physomeloe Reitter, with description of the first instar larva (Coleoptera Meloidae) . EOS Revista Española de Entomologia , 69 : 45 – 56 .
- Bologna , MA , Aloisi , G and Marangoni , C. 1989 . Nuove osservazioni su Eurymeloe Reitter e descrizione di larve del I stadio (Coleoptera, Meloidae) . Bulletin et Annales de la Société Royale Belge d'Entomologie , 125 : 67 – 75 .
- Bologna , MA and Di Giulio , A. 2008 . Revision of the genus Trichomeloe Reitter, with the description of new species and first instar larvae (Coleoptera: Meloidae) . Contributions to Zoology , 77 : 227 – 248 .
- Bologna , MA and Di Giulio , A. 2011 . Biological and morphological adaptations in the pre-imaginal phases of the beetle family Meloidae . Atti dell'Accademia Nazionale Italiana di Entomologia , 59 : 141 – 152 .
- Bologna , MA , Fattorini , S and Pinto , JD. 2001 . Review of the primitive blister beetle genus Iselma with description of the first instar larva (Coleoptera: Tenebrionoidea: Meloidae) . African Entomology , 9 : 105 – 129 .
- Bologna , MA , Oliverio , M , Pitzalis , M and Mariottini , P. 2008 . Phylogeny and evolutionary history of the blister beetles (Coleoptera, Meloidae) . Molecular Phylogenetics and Evolution , 48 : 679 – 693 .
- Bologna , MA and Pinto , JD. 1992 . A review of Meloe (Taphromeloe), including a description of the first-instar larva of M. (T.) erythrocnemus and comments on the classification of the tribe Meloini (Coleoptera: Meloidae) . Proceedings of the Entomological Society of Washington , 94 : 299 – 308 .
- Bologna , MA and Pinto , JD. 1995 . The triungulin of two palaearctic Meloe subgenera: Lasiomeloe Reitter and Micromeloe Reitter (Coleoptera, Meloidae), with bionomic and taxonomic notes . Bollettino di Zoologia , 62 : 383 – 393 .
- Bologna , MA and Pinto , JD. 1998 . A review of the Afrotropical species of Meloe Linnaeus 1758 (Coleoptera Meloidae) with descriptions of first instar larvae, a key to species and an annotated catalogue . Tropical Zoology , 11 : 19 – 59 .
- Bologna , MA and Pinto , JD. 2001 . Phylogenetic studies of Meloidae (Coleoptera), with emphasis on the evolution of phoresy . Systematic Entomology , 26 : 33 – 72 .
- Bologna , MA , Turco , F and Pinto , JD. 2010 . “ 11.19. Meloidae Gyllenhal 1810 ” . In Coleoptera, Beetles, Vol. 2: Morphology and Systematics (Elateroidea, Bostrichiformia, Cucujiformia partim) , Edited by: Leschen , R AB , Beutel , RG , Lawrence , JF , Kristensen , NP and Beutel , RG . 681 – 693 . Berlin/New York : De Gruyter . Arthropoda: Insecta. Handbook of Zoology
- Cros , A. 1912 . Contribution à la biologie de Méloides algériens (Note préliminare) . Feuille des Jeunes Naturalistes , 42 : 131 – 132 .
- Cros , A. 1917 . Forme des ongles des larves primaries des Meloidae et valeur du terme “trioungulin” . Annales de la Société Entomologique de France , 86 : 159 – 164 .
- Cros , A. 1919 . Notes sur les larves primaries des Meloidae avec indicatons des larves nouvelles . Annales de la Société Entomologique de France , 88 : 261 – 279 .
- Cros , A. 1922 . Notes sur les larves primaries des Meloidae avec indicatons des larves nouvelles. (2e Série) . Annales de la Société Entomologique de France , 90 ( 1921 ) : 133 – 135 .
- Cros , A. 1929 . Observations nouvelles sur le Meloés . Annales de Sciences Naturelles, Zoologie (10) , 12 : 137 – 191 .
- Cros , A. 1934 . Le Meloe affinis Lucas var. setosus Escherich ses moeurs, sa larve primaire . Bulletin de la Société d'Histoire Naturelle de l'Afrique du Nord , 21 : 88 – 104 .
- Cros , A. 1935 . Description d'une larve primaire de Meloe de provenance marocaine et d'espèce inconnue . Bulletin de la Société de Sciences naturelles du Maroc , 15 : 151 – 156 . 1
- Cros , A. 1936 . Le Meloe affinis Lucas, Var. setosus Escherich. Etude biologique . Bulletin de la Société d'Histoire Naturelle de l'Afrique du Nord , 27 : 185 – 196 .
- Cros , A. 1943 . Considérations genérales sur les espèces de Meloe du group “rugosus” Marsh. Observations biologiques sur le M. ganglbaueri Apfelbeck et le M. mediterraneus Müller . Annales de Sciences Naturelles, Zoologie (11) , 5 : 61 – 77 .
- Di Giulio , A , Aberlenc , HP , Vigna Taglianti , A and Bologna , MA. 2003 . Definition and description of larval types of Cyaneolytta (Coleoptera Meloidae) and new records on their phoretic association with Carabidae (Coleoptera) . Tropical Zoology , 16 : 165 – 187 .
- Di Giulio , A , Bologna , MA and Pinto , JD. 2002 . Larval morphology of Meloe of the subgenus Mesomeloe: Inferences on its phylogenetic position and a first instar larval key to the Meloe subgenera (Coleoptera: Meloidae) . Italian Journal of Zoology , 69 : 339 – 344 .
- Escherich , K. 1890 . Revision der behaarten Meloë-Arten der alten Welt . Wiener Entomologische Zeitung , 9 : 87 – 96 .
- Giglio , A , Brandmayr , P , Ferrero , EA , Giulianini , PG , Perrotta , E , Talarico , FF and Zetto Brandmayr , T. 2008 . Ultrastructure of the antennal sensorial appendage of larvae of Ophonus ardosiacus (Lutshnik, 1922) (Coleoptera, Carabidae) and possible correlations between size and shape and the larval feeding habits . Zoologischer Anzeiger , 247 : 209 – 221 .
- Lawrence , JF. 1991 . “ Order Coleoptera ” . In Immature Insects , Edited by: Stehr , FW . Vol. 2 , 144 – 298 . Dubuque , IA : Kendal/Hunt .
- Lückmann , J and Kuhlmann , M. 1997 . Die Triungulinen von Meloë brevicollis Panz. und Meloë rugosus Marsh. Mit Anmerkungen zur Biologie und Ökologie der Larven (Col., Meloidae) . Entomologische Nachrichten Berichten , 41 : 183 – 189 .
- Lückmann , J and Scharf , S. 2004 . Description of the first instar larvae of three species of Meloe with a key to the triungulins of Central European species of this genus (Coleoptera: Meloidae) . European Journal of Entomology , 101 : 313 – 322 .
- MacSwain , JW. 1956 . A classification of the first instar larvae of the Meloidae (Coleoptera) . University of California Publications in Entomology. , 12 ( i–iv ) : 1 – 182 .
- Marcos-García , MA and Moreno Fresneda , AR. 2012 . Los sírfidos (Diptera, Syrphidae) agentes dispersores de larvas triungulinas (Coleoptera, Meloidae) . Boletin de la Asociación española de Entomologia , 36 : 193 – 196 .
- Paoli , G. 1932 . Osservazioni sulla biologia del Dociostaurus maroccanus Thnb . in Italia nelle fasi gregarie e solitarie e sull'azione di alcuni Insetti parassiti. Nuovi Annali di Agricoltura , 12 : 27 – 639 .
- Pinto , JD and Bologna , MA. 1993 . The first instar larva of Meloe afer and M. occultus, with a clarification of antennal structure in larval Meloe (Coleoptera: Meloidae) . Coleopterist Bulletin , 47 : 340 – 348 .
- Pinto , JD , Bologna , MA. and Bouseman , JK. 1996 . First-instar larvae, courtship and oviposition in Eletica: Amending the definition of the Meloidae (Coleoptera: Tenebrionoidea) . Systematic Entomology , 21 : 63 – 74 .
- Ruiz , JL , Bologna , MA and García-París , M. 2010 . Taxonomia y distribución de Meloe (Eurymeloe) saharensis Chobaut, 1898(Coleoptera, Meloidae), con nuevas synonymias y primeros registros para Europa y Macaronesia . Graellsia , 66 : 85 – 96 .
- Selander , RB. 1985 . Spastomeloe, a new genus of Meloini from Peru (Coleoptera: Meloidae) . Journal of the Kansas Entomological Society , 58 : 668 – 685 .
- Selander , RB. 1990 . Blister beetles of the genus Pseudopyrota (Coleoptera: Meloidae) . Journal of the Kansas Entomological Society , 63 : 46 – 79 .
- Selander , RB and Mathieu , JM. 1964 . The ontogeny of blister beetles (Coleoptera, Meloidae) . I. A study of three species of the genus Pyrota. Annals of the Entomological Society of America , 57 : 711 – 732 .