Abstract
We evaluated the vertical use of the water column for feeding by young of the year (YOY) of brown trout (Salmo trutta) and Atlantic salmon (S. salar) in a natural river during the summer. Prey trait analysis and diet composition demonstrated that brown trout showed a higher ability to feed on prey in the surface of the water column than Atlantic salmon. Hence, the differences in the ability to feed at different depths of the water column (surface vs. bottom) are important adaptive features that may reduce interspecific competition between the two sympatric fish species. Finally, results derived from the prey trait analysis provide useful information on resource partitioning between sympatric salmonid species.
Introduction
Ecological interaction in streams between Atlantic salmon Salmo salar Linnaeus, 1758 and brown trout Salmo trutta Linnaeus, 1758 is well documented. A consequence of this knowledge is the great amount of literature on the habitat utilization of these species (e.g., Heggenes et al. Citation1999; Crisp Citation2000; Armstrong et al. Citation2003; Jonsson & Jonsson Citation2011). Previously and based on stream habitat selection studies, several researchers have stated that spatial niche overlap is considerable where the two species co-occur, although young Atlantic salmon tend to occupy faster flowing and shallower habitats (e.g., Baglinière & Arribe-Moutounet Citation1985; Heggenes et al. Citation1999). Moreover, when both fish species co-occur, habitat use by Atlantic salmon is restricted through interspecific competition with the more aggressive brown trout (Heggenes et al. Citation1999 and references therein). Indeed, Höjesjö et al. (Citation2005) demonstrated in a controlled stream channel environment that trout held station by swimming actively in the central regions of food patches, whereas salmon occupied the margins.
The diet composition of brown trout and Atlantic salmon, according to field studies based on number, volume and occurrence methods, is similar (e.g., Maitland Citation1965; Pedley & Jones Citation1978; Gibson & Cunjak Citation1986; Suárez et al. Citation1988). In the literature, there is an increasing number of studies under controlled laboratory conditions about the foraging competition between different salmonids species (e.g., Höjesjö et al. Citation2005 and references therein). These controlled studies provide invaluable preliminary information on feeding behaviour, but further studies under natural conditions are needed to increase ecological realism. In this context, prey trait analysis has been proposed as a functional approach to understand mechanisms involved in predator-prey relationships (de Crespin de Billy & Usseglio-Polatera Citation2002), and consequently it may be useful for understanding inter- and intra-species interactions and the mechanisms that determine food partitioning between them (Sánchez-Hernández & Cobo Citation2011, Citation2012; Sánchez-Hernández et al. Citation2011). So, assuming that age-0 Atlantic salmon and brown trout overlap for food and habitats, the most territorial or the most adaptive species will have prior access to the most profitable resources. Hence, the objective of this study was to analyse whether young of the year (YOY) of S. salar and S. trutta show similar habitat requirements for feeding or if vertical segregation is possible between both sympatric species using prey trait analysis.
Material and methods
The study area was located in the River Ulla, NW Spain (catchment area = 2803 km2, total length = 132 km). The sampling site is located in a protected area called “Islas de Gres” in Ponte Ledesma (UTM: 29T 554283 4738221), and it is an important spawning and refuge area for YOY. Protocols used in this study conform to the ethical laws of the regional government (Xunta de Galicia).
The study was conducted in a riffle section (length = 13.5 m and width = 13 m) where Atlantic salmon and brown trout juveniles were captured. We established a transect perpendicular to the direction of flow at the middle of the study site and, at five equally spaced intervals across the transect, water depth (cm) and current velocity (cm/s) were measured. A water current meter (JDC Electronic; Flowatch®) was used to measure velocity. Depths were measured with a meter stick. Water depth varied between 20 and 60 cm and mean water velocity was ∼68 cm/s. At the moment of the survey (10:00 a.m.), water temperature was 18.2°C, conductivity 98.1 μS/cm and pH 7.82. Dissolved oxygen levels were high (92.4% and 8.66 mg/L).
Fishes were collected in one survey (29 August 2011) using pulsed DC backpack electrofishing equipment (Hans Grassl GmbH, ELT60II). For the purpose of the study 44 fishes of age 0+ were selected and killed immediately by an overdose of anaesthetic (benzocaine): 0+ S. salar (n = 23, mean fork length = 6.8 cm ± 0.17 SE) and 0+ S. trutta (n = 21, mean fork length = 6.6 cm ± 0.18 SE). In the laboratory, specimens were dissected and stomach contents were removed. Prey items were identified to the lowest taxonomic level possible and data are offered on relative abundance and frequency of occurrence of preys (Sánchez-Hernández et al. Citation2011). Diet overlap between species was assessed using the Schoener's overlap index (Schoener Citation1970).
By means of the software R (version 2.11.1), a fuzzy principal component analysis (FPCA) was used to analyse the habitat requirements for feeding according to the prey items consumed by fishes. We used the same trait database and trait analyses as de Crespin de Billy (Citation2001), and de Crespin de Billy & Usseglio-Polatera (Citation2002). The complete list of taxa and scores used in the prey trait analysis are provided as supplementary material; see Appendix A. To evaluate the potential vulnerability of invertebrates to fish predation, de Crespin de Billy & Usseglio-Polatera (Citation2002) created a total of 71 different categories for 17 invertebrate traits. Information was structured using a “fuzzy coding” procedure (Chevenet et al. Citation1994). A score was assigned to every taxon, describing its affinity for each trait category, from “0” indicating “no affinity” to “5” indicating “high affinity”. The taxonomic resolution (order, family or genus) used in the classification process corresponded to the lowest possible level of determination of taxa in fish gut contents. When identification to genus was not possible, or in the case of missing information for a certain genus, the value assigned for a trait was that of the family level, using the average profile of all other genera of the same family (de Crespin de Billy & Usseglio-Polatera Citation2002; Rodrígues-Capítulo et al. Citation2009). In this study, Hydroptilidae and Leptoceridae were not included in the analysis because trait values are still not available. Advantages and disadvantages of prey trait analysis and its application in different freshwater species to study feeding behaviour have been published elsewhere (e.g., Sánchez-Hernández et al. Citation2011), suggesting that this methodology provides extremely valuable ecological information on the mechanisms involved in predator-prey relationships, and complements traditional diet analysis (Sánchez-Hernández & Cobo Citation2011; Sánchez-Hernández et al. Citation2011, Citation2012). In the present study, we used four ecological traits of macroinvertebrate prey to demonstrate that partitioning of feeding habitat between species is possible; “current velocity”, “macrohabitat”, “surface drift tendency” and “water drift behaviour” traits were chosen for the analysis of the feeding habitat requirements of these co-occurring species. Differences between species were tested with Mann–Whitney U test in PASW Statistics 18.0 software for Windows (SPSS Inc.).
Results
No empty stomachs were observed. The diets of both fish species consisted primarily of aquatic invertebrates, with Baetidae dominating, although in different proportions (see ). In the case of brown trout, chironomid larvae were also an important item, showing Baetidae and Chironomidae with the same abundance (25.42%). Regarding the input of allochthonous preys, significant differences in the abundance of terrestrial invertebrates were found between species (Mann-Whitney U test, p < 0.001) with higher values in brown trout (5.3% ± 12.66 SE) than in Atlantic salmon (0.2% ± 0.87 SE). Schoener's index showed high values of diet overlap (value = 1).
Table I. Diet composition for each fish species in the Ulla River. Abundance (A i %) and frequency of occurrence (F i %)
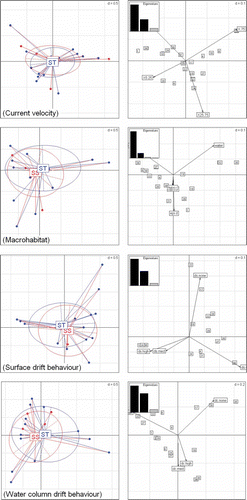
Diagrams of prey trait analyses are shown in . The “current velocity” trait shows a more ample distribution of values in the FPCA for Atlantic salmon, although overlap between both species is high (). Opposite, in (“macrohabitat”) the ellipse of brown trout is displaced to the right, which reveals a higher contribution in specific abundance of terrestrial invertebrates to the diet. The trait “tendency to drift at the water surface” () and diet analysis () suggest that S. trutta tends to feed on prey with high surface drift tendency due to the presence of terrestrial invertebrates such as aerial imagoes of Trichoptera, Chironomidae and Simuliidae. Finally, as can be seen in , high overlap was found between species in the ability to feed on preys with tendency to drift in the water column. However, the ellipse of brown trout is displaced to the right, which reveals a higher contribution in specific abundance of prey items with medium and high water drift tendency.
Discussion
In salmonids, as in many other fish species, there is normally a shift in the composition of the diet during ontogeny, the diet of YOY being dominated by aquatic macroinvertebrates (e.g., Suárez et al. Citation1988; Sánchez-Hernández & Cobo Citation2012) such as in the present study. For example, in our study mayflies of the family Baetidae constituted the most important prey both in abundance and frequency terms for Atlantic salmon, results that are broadly in accordance with previous studies on other freshwater fish species (e.g., Sánchez-Hernández & Cobo Citation2011).
Several authors have found that diet overlap is high between Atlantic salmon and other salmonids species in sympatry (Gibson & Cunjak Citation1986; Coghlan et al. Citation2007). In a previous study in Spain, Suárez et al. (Citation1988) found significant interspecific differences in the diet between S. salar and S. trutta in May and November, although they did not find differences in April and July. On the other hand, Pedley and Jones (Citation1978) stated that interspecific competition between S. salar and S. trutta was only significant during the summer months when food was abundant. Our results, using the Schoener's index, suggest a remarkable similarity in prey utilization patterns between species as previously stated by other researchers (Gibson & Cunjak Citation1986; Coghlan et al. Citation2007). However, the high overlap values found in the present study might not indicate competition, for two reasons. First, as no data on potential preys (benthos and drift) are available, food could be so abundant that fishes might not compete for it. Second, fishes usually can adopt different strategies that overcome the effects of competition (Sánchez-Hernández & Cobo Citation2011; Sánchez-Hernández et al. Citation2011); in our case prey trait analysis and diet composition shows higher preference to feed on surface (terrestrial) drift for YOY brown trout.
Höjesjö et al. (Citation2005) found in a controlled stream channel experiment that brown trout held station by swimming actively in the central regions of food patches, whereas Atlantic salmon occupied the margins, generally remaining stationary on the stream bed. Coghlan et al. (Citation2007) found in subyearling fish that Atlantic salmon consumed a greater proportion of baetid mayflies than rainbow trout Oncorhynchus mykiss (Walbaum, 1792), which fed more frequently on chironomid larvae and aerial-terrestrial invertebrates. In our study, differences in the number of terrestrial invertebrates consumed by both species on the stream surface are evident; observations are in agreement with Suárez et al. (Citation1988), as during the summer season they found that terrestrial prey were more abundant in S. trutta than in S. salar. Hence, terrestrial prey constitute an important food item that may contribute greatly to reduce fish interactions between YOY of sympatric S. trutta and S. salar, as found in other fish species (e.g., Sánchez-Hernández et al. Citation2011 and references therein). Accordingly, prey trait analysis demonstrates that differences in habitat utilization between both fish species are related to the ability to feed at different depths of the water column (surface vs. bottom) due to the higher contribution in specific abundance of terrestrial invertebrates in the diet of brown trout.
In conclusion, our study supports the hypothesis that differences in the ability to feed at different depths of the water column in the same riffle sections between S. salar and S. trutta may reduce trophic competition, and may be seen as important adaptations in feeding behaviour when these fish species co-occur. Additionally, we show that prey trait analysis provides extremely valuable ecological information on the mechanisms involved in predator-prey relationships, and complements traditional diet analysis (Sánchez-Hernández & Cobo Citation2011; Sánchez-Hernández et al. Citation2011, Citation2012). However, future studies might extend the present work and investigate the uses of microhabitat feeding patches by snorkel observations to corroborate if the vertical distribution between species for feeding is evident.
Appendix.xls
Download MS Excel (126.5 KB)Acknowledgements
This work has been partially supported by the project 10PXIB2111059PR of the Xunta de Galicia, and the project MIGRANET of the Interreg IV B SUDOE (South-West Europe) Territorial Cooperation Programme (SOE2/P2/E288). The authors are also grateful to two anonymous referees for their helpful comments.
References
- Armstrong , JD , Kemp , PS , Kennedy , GJA , Ladle , M and Milner , NJ. 2003 . Habitat requirements of Atlantic salmon and brown trout in rivers and streams . Fisheries Research , 62 : 143 – 170 .
- Baglinière , JL and Arribe-Moutounet , D. 1985 . Microrépartition des populations de truite commune (Salmo trutta L.) de juvenile de saumon atlantique (Salmo salar L.) et des autres espèces présentes dans la partie haute du Scorff (Bretagne) . Hydrobiologia , 120 : 229 – 239 .
- Chevenet , F , Dolédec , S and Chessel , D. 1994 . A fuzzy coding approach for the analysis of long-term ecological data . Freshwater Biology , 31 : 295 – 309 .
- Coghlan , SM Jr , Cain , GR and Ringler , NH. 2007 . Prey selection of subyearling Atlantic salmon and rainbow trout coexisting in a natural stream . Journal of Freshwater Ecology , 22 : 591 – 608 .
- Crisp , DT. 2000 . Trout and salmon: Ecology, conservation and rehabilitation , 224 Oxford , , UK : Wiley-Blackwell .
- de Crespin de Billy , V. 2001 . Régime alimentaire de la truite (Salmo trutta L.) en eaux courantes: Rôles de l'habitat physique des traits des macroinvertébrés , Lyon , , France : L'université Claude Bernard . Ph.D. thesis
- de Crespin de Billy , V and Usseglio-Polatera , P. 2002 . Traits of brown trout prey in relation to habitat characteristics and benthic invertebrate communities . Journal of Fish Biology , 60 : 687 – 714 .
- Gibson , RJ and Cunjak , RA. 1986 . An investigation of competitive interactions between brown trout (Salmo trutta L.) and juvenile Atlantic salmon (Salmo salar L.) in rivers of the Avalon Peninsula, Newfoundland . Canadian Technical Report of Fisheries and Aquatic Sciences , 1472 : 1 – 82 .
- Heggenes , J , Baglinière , JL and Cunjak , RA. 1999 . Spatial niche variability for young Atlantic salmon (Salmo salar) and brown trout (S. trutta) in heterogeneous streams . Ecology of Freshwater Fish , 8 : 1 – 21 .
- Höjesjö , J , Armstrong , JD and Griffiths , SW. 2005 . Sneaky feeding by salmon in sympatry with dominant brown trout . Animal Behaviour , 69 : 1037 – 1041 .
- Jonsson , B and Jonsson , N. 2011 . “ Ecology of Atlantic salmon and brown trout. Habitat as a template for life histories ” . In Fish & Fisheries Series 33 , Dordrecht : Springer .
- Maitland , PS. 1965 . The feeding relationships of salmon, trout, minnows, stone loach and three-spined stickle-backs in the River Endrick, Scotland . Journal of Animal Ecology , 34 : 109 – 133 .
- Pedley , RB and Jones , JW. 1978 . The comparative feeding behavior of brown trout, Salmo trutta L. and Atlantic salmon, Salmo salar L. in Llyn Dwythwch, Wales . Journal of Fish Biology , 12 : 239 – 256 .
- Rodrígues-Capítulo , A , Muñoz , I , Bonada , N , Gaudés , A and Tomanova , S. 2009 . “ La biota de los ríos: Los invertebrados ” . In Conceptos y técnicas en ecología fluvial , Edited by: Elosegi , A and Sabater , S . 253 – 270 . Bilbao : Fundación BBVA .
- Sánchez-Hernández , J and Cobo , F. 2011 . Summer food resource partitioning between four sympatric fish species in Central Spain (River Tormes) . Folia Zoologica , 60 : 189 – 202 .
- Sánchez-Hernández , J and Cobo , F. 2012 . Summer differences in behavioural feeding habits and use of feeding habitat among brown trout (Pisces) age classes in a temperate area . Italian Journal of Zoology , 79 : 468 – 478 .
- Sánchez-Hernández , J , Servia , MJ , Vieira-Lanero , R and Cobo , F. 2012 . Application of the analysis of prey ecological characteristics (traits) for the study of the feeding behaviour of bottom-feeder fishes: The example of the stickleback (Gasterosteus gymnurus Cuvier, 1829) . Limnetica , 31 : 59 – 76 . In Spanish
- Sánchez-Hernández , J , Vieira-Lanero , R , Servia , MJ and Cobo , F. 2011 . Feeding habits of four sympatric fish species in the Iberian Peninsula: Keys to understanding coexistence using prey trais . Hydrobiologia , 667 : 119 – 132 .
- Schoener , TW. 1970 . Nonsynchronous spatial overlap of lizards in patchy habitats . Ecology , 51 : 408 – 418 .
- Suárez , JL , Reiriz , L and Anadón , R. 1988 . Feeding relationships between two salmonid species and the benthic community . Polish Archives of Hydrobiology , 35 : 341 – 359 .
Supplementary Material
Supplementary material for this article is available from: http://www.aix.cemagref.fr/htmlpub/divisions/Hyax/titres–publication.htm (doi: 10.1111/j.1095-8649.2002.tb01694.x).