
Abstract
The aim of this study was to analyse and compare the cerebral arterial circle, its anastomoses, its branches and the manner of blood supply in 20 individuals of the Camelidae family. The Camelidae family was represented by Camelus bactrianus (n = 8), Camelus dromedarius (n = 2), Lama guanicoe (n = 5) and Lama glama (n = 5). We performed the analyses on post-mortem material obtained from Polish zoological gardens. Arteries of the heads were injected with pigmented acetone solution of vinyl superchloride. Arteries of the brain base in Camelus and Lama species are similar. In contrast, the pattern of brain base arteries in Camelidae differs from the patterns described in other species of the Artiodactyla order. Specific traits of the base brain arterial pattern in Camelidae may prove useful as a taxonomic exponent of the animals within the Artiodactyla order.
Introduction
Subsequent to studies on the vascular supply of the human brain, initiated at the beginning of the 17th century by Willis (Citation1664) and continued at present using contemporary techniques, analogous analyses were conducted in animals. Jenke (Citation1919) described patterns of arterial supply to the brain in domestic animals, including horses, dogs, cattle and pigs, and compared the results of these observations with the arterial pattern in the base of the human brain. Currently, the list of publications related to cerebral arteries in animals has been significantly extended, including several species: dog (Ueshima & Suenaga Citation1972; Habermebl Citation1973; Wiland Citation1973; Tanuma Citation1981), horse (Rösslein Citation1987), cattle and small ruminants (König Citation1979), yak (Ding et al. Citation2007), European bison (Węgrzyn et al. Citation1983), giraffe (Frąckowiak & Jakubowski Citation2008), fallow deer (Brudnicki Citation2011), goat (Längle Citation1973; Brudnicki Citation2000), cat (Klein Citation1980), various species of Felidae (Frąckowiak & Godynicki Citation2003) and also rat (Jabłoński Citation1975), coypu (Roskosz et al. Citation1986), guinea pig (Ocal & Ozer Citation1992; Majewska-Michalska Citation1994, Citation1995), European beaver (Frąckowiak & Śmiełowski Citation1998) and otter (Brudnicki Citation2012).
In Camelidae, such analyses were infrequent and included description of the blood vessels only for the domesticated form of dromedary (Kanan Citation1970; Smuts & Bezuidenhout Citation1987; Ocal et al. Citation1999). This study aimed to describe a comparative anatomical analysis of the cerebral arterial circle, its branches, its anastomoses and the manner of its blood supply in various species of the Camelidae family.
Material and methods
The studies were conducted on 20 preparations of head arteries, including the cerebral arteries, prepared from post-mortem material of animals belonging to the Camelidae family, represented by Camelus bactrianus (Linnaeus, 1758; n = 8), Camelus dromedarius (Linnaeus, 1758; n = 2), Lama guanicoe (Müller, 1776; n = 5) and Lama glama (Linnaeus, 1758; n = 5). The post-mortem material originated from Polish national zoological gardens. The course and anastomoses of arteries and their variations and asymmetry were examined on arterial cast preparations. The cast preparations were obtained using bilateral injections of pigmented acetone solution of vinyl superchloride into the common carotid arteries introduced at the pressure of 60–80 kPa. Following hardening of the filling material, the specimens were subjected to biological maceration in tap water at 45°C for 1 month, yielding arterial casts on the osseous scaffold. The applied nomenclature followed Nomina Anatomica Veterinaria (Citation2012), as illustrated by Simoens et al. (Citation1978–79).
Results
The cerebral arterial circle (Circulus arteriosus cerebri) is situated within the cranium at the base of the brain ( and ). In the studied Camelidae the cerebral arterial circle was delimited bilaterally at the caudal quadrant by the caudal communicating arteries (Aa. communicantes caudales), and at the rostral quadrant by bilateral rostral cerebral arteries (Aa. cerebri rostrales).
Figure 1. Dorsal view of the arterial circle of the brain and its connections and branches of the Bactrian camel (Camelus bactrianus). 1: A. basilaris; 2: Aa. cerebelli rostrales; 3: A. cerebri media; 4: A. choroidea rostralis; 5: Rete mirabile epidurale rostrale; 6: A. communicans caudalis; 7: A. cerebri caudalis; 8. A. cerebri rostralis; 9. A. cerebelli caudalis.
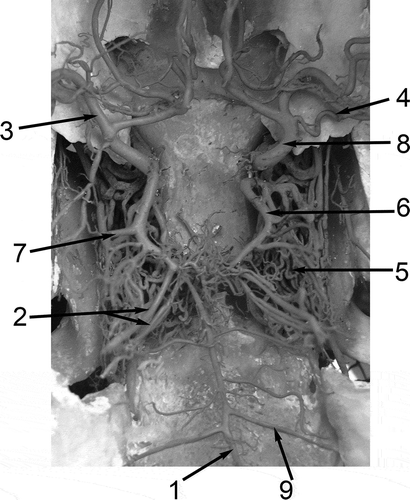
Figure 2. Dorsal view of the arterial circle of the brain and its connections and branches of the llama (Lama glama). 1: A. basilaris; 2: Aa. cerebelli rostrales; 3: A. cerebri media; 4: A. choroidea rostralis; 5. Rete mirabile epidurale rostrale; 6. A. communicans caudalis; 7. A. cerebri caudalis; 8. A. cerebri rostralis; 9. A. cerebelli caudalis.
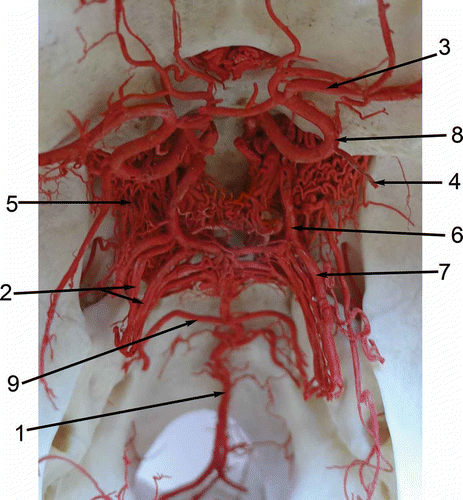
The rostral cerebral artery and the caudal communicating artery stem directly from blood vessels of the rostral epidural rete mirabile, providing the main source of blood for the cerebral arterial circle. A manifestly lower input to the cerebral arterial blood supply was provided by the basilar artery, which is formed by the vertebral arteries and anastomoses with the cerebral arterial circle. The rostral cerebral artery (A. cerebri rostralis) arises from the rostral epidural rete mirabile and delimits the rostro-lateral segment of the cerebral arterial circle, whereafter it extends to the region of the olfactory bulbs.
The rostral cerebral artery yields the rostral choroidal artery and a large middle cerebral artery, and then it forms an anastomosis with the rete mirabile at the optic chiasm (Rete chiasmaticum).
In three Bactrian camels, both dromedaries and all llamas, the bilateral rostral cerebral arteries anastomosed with each other at the level of the Chiasma opticum through a short rostral communicating artery, in this way completing the cerebral arterial circle. In the other five Bactrian camels, the rostral cerebral arteries were not connected with each other.
The rostral cerebral artery was connected with the caudal communicating artery through additional anastomoses, which were demonstrated unilaterally in one guanaco and in two Bactrian camels, and bilaterally in one guanaco, one llama and five Bactrian camels.
In three Bactrian camels, the rostral cerebral artery and the caudal communicating artery originated from an intracranial segment of the internal carotid artery, while in one llama, such an intracranial segment of the internal carotid artery extended only into the left caudal communicating artery.
Following its exit from the rostral epidural rete mirabile, the caudal communicating artery (A. communicans caudalis) forms the caudolateral segment of cerebral arterial circle.
The bilateral caudal communicating arteries are connected with each other through an anastomosis, which was a single artery in four Bactrian camels, one llama, one guanaco and one dromedary, and a double artery in only one guanaco and one dromedary.
The basilar artery (A. basilaris) was formed by the junction of the bilateral vertebral arteries and in most animals it fused rostrally with both caudal communicating arteries in the median plane. However, in two Bactrian camels, the basilar artery manifested asymmetry in the manner of fusion with the caudal communicating artery as it was shifted to the right caudal communicating artery in one of the Bactrian camels, and to the left one in the other. In one guanaco, one dromedary and three Bactrian camels, the basilar artery was additionally connected with the right caudal communicating artery through a few anastomoses.
Before joining the caudal communicating arteries, the basilar artery formed a rhomboid arterial ring in all llamas and in both dromedaries. In another vascular variation in one llama, the basilar artery was partially double.
The basilar artery gave off caudal and rostral cerebellar arteries and arteries supplying the pons.
The rostral choroidal artery (A. choroidea rostralis) originated from the dorsal side of the rostral cerebral artery. The vascular varieties involved also a variable number of rostral choroidal arteries. Two rostral choroidal arteries were unilaterally present in one dromedary and in one Bactrian camel, while they were bilaterally present in three Bactrian camels and in the other dromedary.
The middle cerebral artery (A. cerebri media) branched away from the rostral cerebral artery halfway down its length, forming its principal branch. In some of the studied animals, the middle cerebral artery was represented by multiple blood vessels and manifested asymmetry of its origin. A double middle cerebral artery was demonstrated in two Bactrian camels, two llamas and one guanaco on both sides, and on a single side in three Bactrian camels and two dromedaries. A triple middle cerebral artery was detected in three Bactrian camels and a single dromedary on one side, and in a single guanaco on both sides.
The caudal cerebral artery (A. cerebri caudalis) represented one of the most pronounced blood vessels given off by the cerebral arterial circle and stemmed from the rostral segment of the caudal communicating artery.
As a vascular variety, a double caudal cerebral artery was detected in one llama and two Bactrian camels on one side, and in one dromedary on both sides.
In some animals, the caudal cerebral artery anastomosed with the rostral cerebral artery through additional blood vessels. Such a junction of the two segments of the cerebral arterial circle was noted in a llama and two dromedaries.
The rostral cerebellar artery (A. cerebelli rostralis) may be represented by a single or a multiple blood vessel (involving two, three or even four blood vessels). Due to the significant variability in the number of branches representing the rostral cerebellar artery, the corresponding branches stemmed not only from the caudal communicating artery (most frequently two or three in number), but also from the rostral part of the basilar artery (usually represented by a double blood vessel).
The caudal cerebellar artery (A. cerebelli caudalis) originated from the basilar artery, perpendicular to its long axis and arising halfway down its length, close to the caudal edge of the pons.
As a vascular variety, a double caudal cerebellar artery was present on a single side in one Bactrian camel, one llama and three guanaco, and on both sides in two llamas. Its bilateral single manifestation was detected in five Bactrian camels, a single dromedary, two llamas and a single guanaco.
The internal carotid artery (A. carotis interna) stems from the common carotid artery, passes through the jugular foramen and joins the rostral epidural rete mirabile.
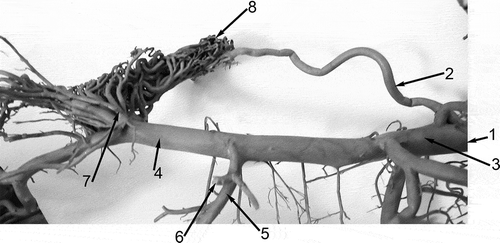
The internal carotid artery, together with branches of the maxillary artery and the external ophthalmic artery, provides the blood supply for the rostral epidural rete mirabile ().
Figure 3. Left lateral view of the arteries supplying the rostral epidural rete mirabile and some of the head arteries of the Bactrian camel (Camelus bactrianus). 1: A. carotis communis; 2. A. carotis interna; 3. A. carotis externa; 4. A. maxillaris; 5. A. alveolaris inferior; 6. A. temporalis profunda caudalis (cut off); 7. Rami rostrales ad rete mirabile epidurale rostrale; 8. Rete mirabile epidurale rostrale.
The internal ethmoidal artery (A. ethmoidalis interna) stems from the maxillary artery and joins the rostral cerebral artery in the ethmoidal fossa.
The chiasmatic rete (Rete chiasmaticum) at the optic chiasm is joined to the external ethmoidal artery, and within the cranium it anastomoses with the rostral cerebral artery.
The paired rostral epidural rete mirabile (Rete mirabile epidurale rostrale) is formed by numerous dense, reciprocally anastomosed arteries rests on the floor of the cranial cavity.
In all examined Camelidae, the rostral epidural rete mirabile was found to be connected to the fully persistent internal carotid artery. Moreover, the rostral cerebral artery and caudal communicating artery, which together form the cerebral arterial circle, arose separately from the rostral epidural rete mirabile.
Discussion
The conducted observations of arteries at the cerebral base of animals belonging to the Camelidae family have demonstrated a significant similarity of the blood vessels in camels and llamas, i.e. the camels of the Old and the New World.
The only sources of information on this vascular region in Camelidae include the pattern of cerebral blood supply in a dromedary published by Kanan (Citation1970), the description of the blood vessels presented in the textbook of Smuts and Bezuidenhout (Citation1987), and morphometric studies of the dromedary by Ocal et al. (Citation1999) or selected even-toed ungulates by Godynicki (Citation1972).
In principle, the results of the observations in this study are consistent with the literature data, while divergences involve nomenclature used by the above quoted authors, as compared to the current official anatomic nomenclature.
The cerebral arterial circle in Camelidae is supplied directly by blood vessels originating from the rostral epidural rete mirabile and by the basilar artery, formed by fusion of the vertebral arteries. Moreover, the chiasmatic rete, situated in the examined animals between the external ethmoidal artery and cerebral arterial circle, supports the blood supply to the brain, and such a rete was demonstrated in cattle by Simoens et al. (Citation1978–79). In Camelidae, the rostral epidural rete mirabile anastomoses with branches of the maxillary artery and with the internal carotid artery, which distinguishes this vascular pattern in Camelidae among other artiodactylous animals (Godynicki Citation1972; Godynicki & Frąckowiak Citation1979).
In the studied Camelidae, the rostral cerebral artery and the caudal communicating artery usually do not stem from a terminal branching of the internal carotid artery, but each of them stems directly from blood vessels of the rostral epidural rete mirabile. Such an independent origin of the rostral cerebral artery and the caudal communicating artery from the rostral epidural rete mirabile was demonstrated in a dromedary only by Kanan (Citation1970), and in various species of Camelidae by Godynicki (Citation1972). A distinct opinion on the matter was presented by Smuts and Bezuidenhout (Citation1987), who stated that in the camel, the rostral cerebral and caudal communicating arteries arose from the terminal division of the internal carotid artery, which in the study of Ocal et al. (Citation1999) was designated as the cerebral carotid artery.
Nomenclature related to the rostral cerebral artery has not been made uniform and in various publications it differs from the nomenclature sanctioned by Nomina Anatomica Veterinaria (Citation2012).
According to Simoens et al. (Citation1978–79), the rostral cerebral artery consists of three segments, the initial fragment contained between the terminal division of the internal carotid artery and the origin of the middle cerebral artery, the subsequent segment which reaches the chiasma opticum, and the third fragment which runs rostrally in the longitudinal fissure of the brain to bend caudally on the genu of the corpus callosum and continues its course on the medial surface of the cerebral hemisphere. In species in which the third segment of the bilateral rostral cerebral arteries do not form a common trunk but are arranged in parallel to each other in the longitudinal fissure of the brain, the arterial cerebral circle becomes closed by the anterior communicating artery (Jenke Citation1919; Wiland Citation1967; Gillilan Citation1974). In the present study, such a rostral communicating artery has been detected in all examined Camelidae, except for five Bactrian camels in which the rostral cerebral arteries were neither connected nor fused.
The course of the caudal communicating artery in Camelidae is different as compared to other animals since it does not connect the internal carotid artery with the basilar artery (Simoens et al. Citation1978–79).
In the study of Jenke (Citation1919), the caudal communicating artery was marked as the caudal communicating branch of the internal carotid artery, while Rösslein (Citation1987) stated that it represented a caudal branch of the arterial cerebral circle. In this study, we have followed the Nomina Anatomica Veterinaria (Citation2012) and recommendations enclosed in the study of Simoens et al. (Citation1978–79) and, consequently, the caudal communicating artery has been termed in this way, beginning at its origin in the rostral epidural rete mirabile to its fusion with the basilar artery.
The branches stemming from the arterial circle of the brain, which supply various regions of the brain, manifest variability both in their number and in their course. Their vascular varieties in the studied Camelidae include a variability in the number of branches forming the rostral choroidal artery, middle cerebral arteries and caudal cerebral artery.
In the studied animals, the rostral cerebellar arteries formed numerous branches, of which two or three stemmed from the caudal communicating artery while the other two or three branches originated from the basilar artery.
In the studied Camelidae, the caudal cerebellar artery stemmed from the basilar artery close to the caudal edge of the pons, and such an origin of this artery was described earlier in a dromedary by Kanan (Citation1970) and by Smuts and Bezuidenhout (Citation1987). Vascular varieties in Camelidae included a double caudal cerebellar artery, while multiple caudal cerebellar arteries were earlier described in Equidae by Frąckowiak and Godynicki (Citation1999).
Variability of the basilar artery included the manner of its junctions with the caudal communicating arteries and, less frequently, with segmental manifestation of a double vessel.
Traits of the vascular system also supply data of general biological importance. Shoshani and McKenna (Citation1998) used certain properties of the vascular system for drafting a cladogram of contemporary mammals. It is suggested that the specificity of the brain base arterial pattern in Camelidae may provide a taxonomic exponent of the animals within the order of Artiodactyla.
Conclusions
Arteries at the brain base in Camelidae of the Old and the New World resemble each other, whereas the pattern of brain base arteries in Camelidae differs from the patterns described in other species of the Artiodactyla order.
It is suggested that specific traits of the brain base arterial pattern in Camelidae may prove to be useful as a taxonomic characteristic of the animals within the Artiodactyla order.
References
- Brudnicki W. 2000. Basilar arteries of the brain in domestic goat (Capra hircus L). Electronic Journal of Polish Agricultural University Veterinary Medicine 3:1–6.
- Brudnicki W. 2011. Morphometric analysis of the brain base arteries in fallow deer (Dama dama). Original Paper Veterinarni Medicina 56:462–468.
- Brudnicki W. 2012. Brain base arteries: Pattern and variation in the European otter (Lutra Lutra). Anatomia, Histologia, Embryologia 41:358–361. doi:10.1111/j.1439-0264.2012.01145.x.
- Ding Y, Shao B, Wang J. 2007. The arterial supply to the brain of the yak (Bos grunniens). Annals of Anatomy - Anatomischer Anzeiger 189:31–38. doi:10.1016/j.aanat.2006.07.011.
- Frąckowiak H, Godynicki S. 1999. The caudal intercarotid artery in species of order Perissodactyla. Folia Morphologica (Praha) 58:27–30.
- Frąckowiak H, Godynicki S. 2003. Brain basal arteries in various species of Felidae. Polish Journal of Veterinary Sciences 6:195–200.
- Frąckowiak H, Jakubowski H. 2008. Arterial vascularization in the giraffe brain. Annales Zoologici Fennici 45:353–359. doi:10.5735/086.045.0418.
- Frąckowiak H, Śmiełowski J. 1998. Cephalic arteries in the European beaver (Castor fiber). Acta theriologica (Warsz) 43:219–224.
- Gillilan LA. 1974. Blood supply to brains of ungulates with and without a Rete Mirabile Caroticum. The Journal of Comparative Neurology 153:275–290. doi:10.1002/cne.901530305.
- Godynicki S. 1972. Morfologia porównawcza układu tętnic głowy u niektórych zwierząt parzystokopytnych (Artiodactyla). Roczniki Akademii Rolniczej w Poznaniu 36:5–60.
- Godynicki S, Frąckowiak H. 1979. Arterial branches supplying the rostral and caudal retia mirabilia in artiodactyls. Folia Morphologica (Warsz) 4:505–510.
- Habermebl KH. 1973. Zur Topographie der Gehirngefäße des Hundes. Anatomia, Histologia, Embryologia: Journal of Veterinary Medicine Series C 2:327–353. doi:10.1111/j.1439-0264.1973.tb00413.x.
- Jabłoński R. 1975. Zmienność tętnic podstawy mózgowia u szczura laboratoryjnego. Prace Wydz. Nauk Przyrodn. Seria B 23:59–67.
- Jenke TW. 1919. Die Gehirnarterien des Pferdes, Hundes, Rindes und Schweines verglichen mit denen des Menschen. Leipzig: Diss. Dresden.
- Kanan CV. 1970. The cerebral arteries of Camelus dromedarius. Acta Anatomica 7:605–616.
- Klein T. 1980. Korrosionsanatomische Untersuchungen am Blutgefäßsystem des Encephalon und der Meninges bei Felis domestica. Anatomia, Histologia, Embryologia 9:236–279. doi:10.1111/j.1439-0264.1980.tb00863.x.
- König H. 1979. Anatomie und Entwicklung der Blutgefässe in der Schädelhohle der Hauswiederkäuer (Rind, Schaf und Ziege). Stuttgart: Ferdinand Enke Verl.
- Längle D. 1973. Korrosionsanatomische Untersuchungen am Blutgefässsystem des Encephalon und der Meninges bei Capra hircus. München: Diss. med. vet.
- Majewska-Michalska E. 1994. Vascularization of the brain in guinea pig. I. Gross anatomy of the arteries and veins. Folia Morphologica (Warsz) 53:242–268.
- Majewska-Michalska E. 1995. Vascularization of the brain in guinea pig. II. Regions of vascular supply and spatial topography of the arteries in particular parts of the brain. Folia Morphologica (Warsz) 54:33–40.
- Nomina Anatomica Veterinaria. 2012. International Committee on Veterinary Gross Anatomical Nomenclature. 5th ed. Hannover, Germany, Columbia, MO, Ghent, Belgium, Sapporo, Japan: Editorial Committee.
- Ocal MK, Erden H, Ogut I, Kara ME. 1999. A Quantitative Study of the Circulus Arteriosus Cerebri of the Camel (Camelus dromedarius). Anatomia, Histologia, Embryologia 28:271–272. doi:10.1046/j.1439-0264.1999.00199.x.
- Ocal MK, Ozer M. 1992. The circulus arteriosus cerebri in the guinea pig. Annals of Anatomy - Anatomischer Anzeiger 174:259–260. doi:10.1016/S0940-9602(11)80365-1.
- Roskosz T, Wiland C, Maliński J. 1986. The arteries of the base of the brain in Coypu (Myocastor coypus) Molina. Annals of Warsaw Agriculture University 13:11–16.
- Rösslein C. 1987. Angioarchitektonische Untersuchungen an den Arterien des Encephalon und der Meninges beim Pferd. München: Inaug-Diss. Ludwig Maximilians Universität.
- Shoshani J, McKenna MC. 1998. Higher taxonomic relationships among extant mammals based on morphology, with selected comparisons of results from molecular data. Molecular Phylogenetics and Evolution 9:572–584. doi:10.1006/mpev.1998.0520.
- Simoens P, De Vos NR, Lauwers H. 1978–79. Illustrated anatomical nomenclature of the heart and the arteries of head and neck in the domestic mammals. Mededelingen van de Fac. Diergeneeskunde Rijksuniv. Gent.
- Smuts MMS, Bezuidenhout AJ. 1987. Anatomy of the dromedary. Oxford: Cladendon Press.
- Tanuma K. 1981. A morphological study on the circle of Willis in the dog. Okajimas Folia Anatomica Japonica 58:155–176.
- Ueshima T, Suenaga Y. 1972. Arteries of the basal region of the brain in the dog. I. Origins of main arteries. Tottori University Journal of the Faculty of Agriculture 7:38–46.
- Węgrzyn M, Roskosz T, Makowiecka M. 1983. Brain arteries of the European bison, Bison bonasus L. 1758. Annals of Warsaw Agriculture University SGGW – AR – Veterinary Medicine 11:9–16.
- Wiland C. 1967. Krąg tętniczy u lisów. PTPN Prace Kom. Nauk Rol. i Kom. Nauk Leśn 23:305–324.
- Wiland C. 1973. Variation of the basal arteries of the brain in dogs. Folia Morphologica (Warsz) 32:63–70.
- Willis T. 1664. Cerebri Anatome: cui accessit nervorum descriptio et usus. Londini: Martyn & Alleftry.