
Abstract
Squirrels have been traded in the pet market for several decades, and numerous species have established in the wild. The Asiatic species Callosciurus erythraeus and Callosciurus finlaysonii have been introduced into other parts of Asia, in Europe and South America. In this study, (1) we conducted a genetic characterization of C. erythraeus introduced into Argentina and compared them with native and introduced populations in Asia, and (2) we analyzed genetic variation among the four invasion foci in Argentina in order to corroborate that the pathway of invasion was a single introduction event in the country and subsequent translocations. We analyzed mitochondrial (cytochrome b, Cyt b; cytochrome oxidase c subunit I, COI and D-loop) and nuclear (recombination activating gene I, RAGI) DNA markers using the classical method (DNA barcoding gap analysis) and also the Automatic Barcode Gap Discovery method (ABGD). The markers D-loop, COI, and RAG1 indicated that the introduced squirrels from the different invasion foci formed a monophyletic group that, together with only one haplotype for the D-loop and COI markers, supported the hypothesis of one introduction event into Argentina followed by subsequent translocations. Unexpectedly, sequences from squirrels captured in Argentina were more related to C. finlaysonii than to C. erythraeus for D-loop and Cyt b markers. However, intraspecific variation among sequences of C. erythraeus belonging to different subspecies or collected in different regions was large and comparable with the distance to the sequences from Argentina. The ABGD method also indicated large genetic variability within C. erythraeus and close proximity between squirrels from Argentina and C. finlaysonii. The complex taxonomy of Callosciurus, as occurs with the sister species C. erythraeus and C. finlaysonii, requires a thorough systematic revision. A simultaneous analysis of diagnostic morphological characters and genetic markers is needed and will provide new insight regarding the worldwide invasion of Asiatic squirrels.
Introduction
The pet trade is a common pathway of introduction of several species worldwide and it is a constant source of individuals that may establish wild populations as a consequence of either accidental escapes or deliberate releases (Hulme et al. Citation2008; Keller et al. Citation2011). Squirrels have been traded in both the legal and illegal pet markets for several decades, and numerous species have now been established in the wild, some of which are considered invasive species (Palmer et al. Citation2008; Bertolino Citation2009). The most frequent vectors of squirrel introductions are the pet market, private citizens and zoos (Bertolino Citation2009). Several mammalian species are intensively hunted and traded in Asia within and between nations, where squirrels are sold in fresh food and pet markets (Timmins & Duckworth Citation2008). The tropics, particularly the forests of South and Southeast Asia, are hotspots of squirrel diversity; however, most studies have focused on temperate squirrel species, raising the need for further research to understand the ecological and taxonomical complexity of tropical squirrels (Kropowski & Nandini Citation2008).
Squirrels of the subfamily Callosciurinae are native to the Indomalayan Region. The genus Callosciurus (Sciuridae: Rodentia) occurs over a large portion of the Indochinese and Malaysian subregions and it is classified into 15 species (Corbet & Hill Citation1992). The species Callosciurus erythraeus (Pallas, 1779) and Callosciurus finlaysonii (Horsfield, 1824) have been introduced into other parts of Asia, Europe and South America as a result of the pet trade (Bertolino Citation2009). Wild populations of C. finlaysonii have only been reported in Italy (Bertolino et al. Citation1999), while C. erythraeus has been recorded in Argentina, Belgium, France, Japan and the Netherlands (Lurz et al. Citation2013). However, a recent study conducted in Japan described some individuals that were morphologically identified as C. erythraeus but genetically closer to C. finlaysonii (Oshida et al. Citation2007). The authors assumed that both species have been introduced into Japan (Oshida et al. Citation2007), where a new haplotype of C. finlaysonii was described in a subsequent study (Kuramoto et al. Citation2012). Recently, Oshida et al. (Citation2013) found that C. erythraeus griseimanus was more closely related to C. finlaysonii than to C. erythraeus cf. hendeei, and suggested that C. e. griseimanus could be considered a distinct species; all of which reinforces the need for further genetic studies.
In 1970, 10 squirrels were introduced close to Luján city, in the province of Buenos Aires (Argentina), and founded the first population of exotic squirrels in South America (Guichón & Doncaster Citation2008). These arboreal squirrels were morphologically identified as C. erythraeus thai (Aprile & Chicco Citation1999); however, the subspecies could also correspond to C. erythraeus atrodorsalis (Cassini & Guichón Citation2009). Variation in pelage colour has been described in the original site of introduction, in Luján, where individuals had the typical olive brown agouti dorsal pelage with a black stripe on the back and reddish ventral coloration. However, squirrels with yellow-creamy underparts and no black stripe on their backs were also found (Cassini & Guichón Citation2009). Other populations of C. erythraeus have been introduced in three Argentinian provinces due to its charismatic appearance (Benitez et al. Citation2013). Interviews with local residents indicated that squirrels were subsequently translocated from the invasion focus in Luján to other locations within Argentina, where they were established as a consequence of intentional releases (Guichón et al. Citation2005; Benitez et al. Citation2013).
In this study, we aimed to analyze genetic variation among the invasion foci of C. erythraeus established in Argentina and their similarity with native and introduced populations in Asia. Therefore, the objectives of this study were (1) to conduct a genetic characterization of the squirrels introduced in Argentina using both mitochondrial and nuclear DNA markers, and (2) to analyze genetic variation among the invasion foci in Argentina in order to corroborate the pathway of invasion as a single introduction event and subsequent translocations into new areas.
Materials and methods
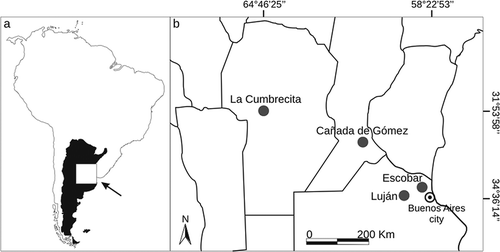
Between February 2007 and June 2010, we obtained tissue samples for DNA analysis of 30 individuals captured in all known invasion foci of C. erythraeus in Argentina: (1) 13 from Luján, Buenos Aires province (34°33'S, 59°07'W), (2) seven from Escobar (34°21'S, 58°48'W), Buenos Aires province, 35 km from Luján, (3) five from Cañada de Gómez (32°48'S, 61°23'W), Santa Fe province, 290 km from Luján, and (4) five from La Cumbrecita (31°53'S, 64°46'W), Córdoba province, 610 km from Luján (Benitez et al. Citation2013) (). Squirrels inhabit rural and urbanized areas where they use highly fragmented woodland patches mainly composed of introduced tree species. In each study site, 30–50 cage traps were set at fixed points 40 m apart on tree branches during four consecutive days. Captured squirrels were anaesthetized subcutaneously using a ketamine-acepromazine mixture, pelage coloration was described, tissue samples from one ear were preserved in 96% ethanol, and individuals were released at the same site of capture.
Figure 1. (a) Location of the area of interest in Argentina (white square) and (b) study areas in the four invasion foci located next to the cities (black dots) of Luján and Escobar in Buenos Aires province, Cañada de Gómez in Santa Fé province, and La Cumbrecita in Córdoba province (lines divide provinces).
We used the mitochondrial gene cytochrome oxidase c subunit I (COI, 650 bp) and a 500-bp fragment of the control region (D-loop) to compare among individuals from the different invasion foci in Argentina and with Callosciurus sequences available in GenBank, from both their native and introduced ranges. These two molecular markers were chosen for their high intraspecific variability, which allows evaluation of the genetic variation of the squirrels in Argentina. To complement the genetic characterization of these squirrels within the genus Callosciurus, some samples were sequenced using the mitochondrial gene cytochrome b (Cyt b) and the nuclear autosomic locus of the recombination activating gene 1 (RAG1). These markers are less variable than COI and D-loop but useful for phylogenetic approaches, and have been previously used for Callosciurus, allowing the comparison with sequences available in GenBank. DNA was extracted using the salt-extraction protocol (Aljanabi & Martínez Citation1997). D-loop sequences were amplified using the universal primers Thr-L15926: 5´-CAATTCCCCGGTCTTGTAAACC-3´ located in the neighbouring tRNA-pro gene and DL-H16340: 5´-CCTGAAGTAGGAACCAGATG-3´ (Vila et al. Citation1999). The Cyt b gene was amplified using a combination of the primers LMVZ055´-CGAAGCTTGATATGAAAAACCATCGTTG-3´ (Smith & Patton Citation1993) and H15910 5´-GATTTTTGGTTTACAAGACCGAG-3´ (Oshida et al. Citation2000). The COI gene was amplified and sequenced in the Canadian Centre for DNA Barcoding, Biodiversity Institute of Ontario (University of Guelph, Guelph, Ontario) using the cocktail primers described in Ivanova et al. (Citation2007) as part of a collaboration between Consejo Nacional de Investigaciones Científicas y Técnicas, Argentina (CONICET) and International Barcode of Life (IBOL). Finally, the RAG1 gene was amplified using primers designed by Steppan et al. (Citation2004). Thermal profile for all markers are detailed in the Appendix; polymerase chain reaction (PCR) products were examined on 1% agarose gels and both chains (forward and reverse) were automatically sequenced in an ABI3100 sequencer (MACROGEN, Inc. Korea). Chromatograms were edited using the program BIOEDIT 7.0 (Hall Citation1999). The sequences were aligned using Clustal X (Thompson et al. Citation1997).
A molecular phylogenetic approach was used to describe the genetic similarity of the squirrels captured in Argentina to available sequences of the four markers of C. erythraeus, its sister species C. finlaysonii and the congenerics C. caniceps, C. prevostii, C. orestes, C. notatus, C. nigrovittatus and C. inornatus (see Appendix). We also used sequences of other squirrel genera as outgroup species. For the D-loop marker, we sequenced 23 samples obtained in the four invasion foci of Argentina (Luján, Escobar, Cañada de Gómez and La Cumbrecita) and used 74 available sequences of the genus Callosciurus, including sequences of C. erythraeus from native (China and Taiwan) and introduced (Japan) ranges, C. finlaysonii from native (Laos and Thailand) and introduced (Japan) ranges, C. prevostii and three species of the genus Petaurista. For the Cyt b marker, we sequenced samples obtained in one invasion focus of Argentina (Cañada de Gómez) and used 30 sequences of Callosciurus that included C. erythraeus from native (Vietnam and Taiwan) and introduced (Japan) ranges, C. finlaysonii from its native range, C. prevostii, C. nigrovittatus, C. inornatus, C. notatus and C. caniceps. We also included two sequences of the species Lariscus insignis and Sciurus lis. For the COI marker, we sequenced samples from 23 squirrels captured in the four invasion foci of Argentina, and used 11 sequences available of C. erythraeus from its native range (Vietnam and China), C. prevostii, C. orestes, C. notatus and the outgroup Dremomys rufigenis. For RAG1, we sequenced four samples obtained in three invasion foci of Argentina, and used four sequences of C. erythraeus from native ranges (China and Taiwan), one of C. prevostii and the outgroup Sundasciurus philippinensis. We found no available sequences of C. finlaysonii for the markers COI and RAG1.
We conducted separate analyses for each molecular marker and estimated appropriate substitution models using the Akaike Information Criterion as implemented in MrAIC (Nylander Citation2004). The model of sequence evolution that best fitted our sequences data for D-loop and COI markers was HKY+G+I. The best model of sequence evolution for Cyt b marker was TN+G and for RAG1 marker the best evolution model was HKY+G+I. We analysed three phylogenetic reconstruction methods. First, we performed maximum likelihood (ML) phylogenetic reconstruction using TreeFinder (Jobb et al. Citation2004) and calculated confidence intervals for the edges of the ML tree using bootstrapping (Felsenstein Citation1985), based on 1000 repetitions. Second, we conducted Bayesian inference (BI) analysis using MrBayes 3.1.2 (Huelsenbeck & Ronquist Citation2001; Ronquist & Huelsenbeck Citation2003). Four chains were run simultaneously (three heated, one cold) for 20,000,000 generations, with tree space sampled every 100th generation. After a graphical analysis of the evolution of the likelihood scores, we discarded the first 300,000 generations as burn-in. We used the remaining trees to calculate the consensus tree. Third, we performed a maximum parsimony (MP) analysis using TNT 1.1 (Goloboff et al. Citation2008) and a traditional search using tree bisection reconnection (TBR) with 1000 random sequence additions, and jacknife support values based on 1000 replicates. For each marker, we computed a distance matrix of the Kimura two-parameter (K2P) model (Kimura Citation1980). In addition to the classical method (DNA barcoding gaps analysis, based on K2P distances) we explored species limits using the Automatic Barcode Gap Discovery method (ABGD) (Puillandre et al. Citation2012a). This method of DNA taxonomy automatically finds the distance at which a barcode gap occurs and sorts the sequences into putative species based on this distance (Puillandre et al. Citation2012a). Therefore, it is applicable as an independent tool without an a priori species hypothesis, and it provides insight into whether the taxonomic identification based on morphological features has any genetic support. This method is also advantageous in terms of computation time and does not require an ultrametric tree that relies heavily on the correctness of the speciation model (Puillandre et al. Citation2012a). We used several mitochondrial markers (D-loop, Cyt b and COI) to avoid problems such as the presence of pseudogenes, incomplete lineage sorting or introgression (Puillandre et al. Citation2012b). The alignments were uploaded to http://wwwabi.snv.jussieu.fr/public/abgd/abgdweb.html and ABGD was run with the default initial settings (Pmin = 0.001, Pmax = 0.1, Steps = 10, X relative gap width = 1.5, Nb bins = 20) and both distance models (Jukes-Cantor and Kimura).
Results
Squirrels analyzed in this study showed high variability in pelage colour and included individuals with a reddish ventral pelage and a black stripe on the back, others with yellow-creamy underparts and no black stripe on their backs, and intermediate variations such as reddish belly with yellow-creamy or orange chest, groin and/or genital area, and with or without a black stripe on the back (Appendix).
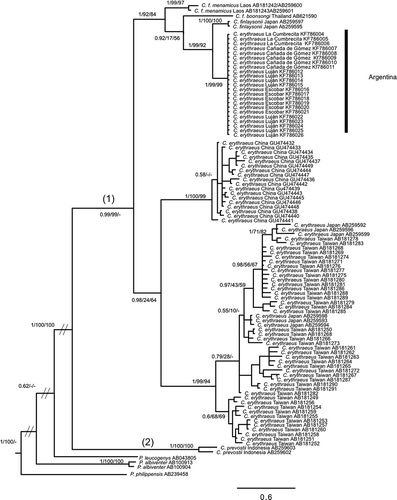
We found only one haplotype by sequence alignment of the D-loop marker in squirrels captured in Argentina. The ML, BI and MP phylogenetic trees obtained had similar topologies and we therefore only show the BI tree with the support values of the three analyses (). The haplotype found in Argentina has not been previously sampled in other populations and was phylogenetically closer to sequences of C. finlaysonii, particularly to the two haplotypes sampled in Japan that had been morphologically identified as C. erythraeus (Oshida et al. Citation2007).
Figure 2. Bayesian inference trees of Callosciurus genus. Tree derived from D-loop. Numbers next to branches are Bayesian posterior probabilities, jackknife support values followed by bootstrap values, respectively. Numbers between brackets on a branch that defines a clade correspond to the groups obtained with the Automatic Barcode Gap Discovery (ABGD) method for the marker.
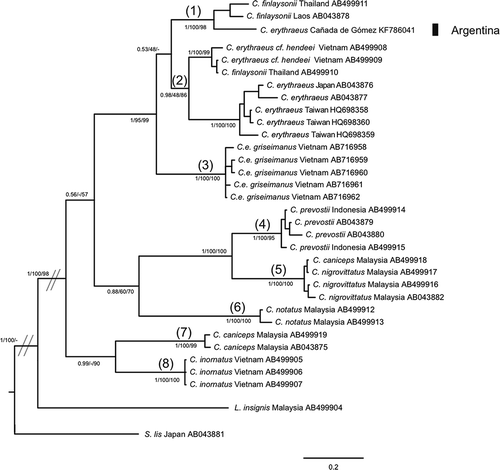
Sequence alignment of the Cyt b marker comprised 700 bp positions. The ML, BI and MP phylogenetic trees obtained had similar topologies (). The haplotype found in Cañada de Gómez, Argentina, has not been described in other populations though it was highly related to C. finlaysonii. C. erythraeus appeared as sister group. However, one sequence described as C. finlaysonii from Thailand was also included in this group. This sequence was strangely identical to one C. erythraeus cf. hendeei from Vietnam. A similar misclassification seemed to occur with a sequence of C. caniceps from Malaysia that was grouped together with C. nigrovittatus instead of its conspecifics (). It must be noted that sequences of C. erythraeus griseimanus separated from the other sequences of C. erythraeus that grouped with C. finlaysonii.
Figure 3. Bayesian inference trees of Callosciurus genus. Tree derived from cytochrome b (Cyt b). Numbers next to branches are Bayesian posterior probabilities, jackknife support values followed by bootstrap values, respectively. Numbers between brackets on a branch that defines a clade correspond to the groups obtained with the Automatic Barcode Gap Discovery (ABGD) method for the marker.
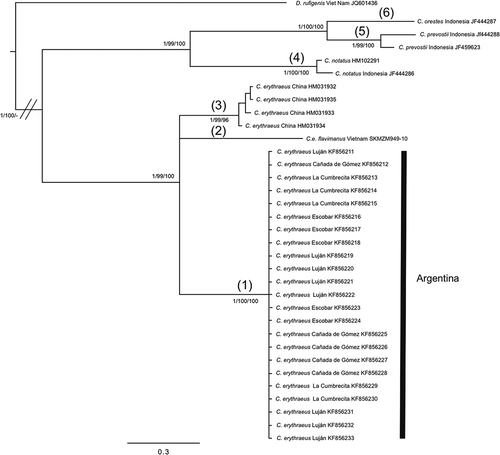
For the COI marker we found only one haplotype in the four invasion foci of Argentina, as we expected given the lower evolution rate of the COI marker compared with D-loop analysis. The ML, BI and MP phylogenetic trees obtained had similar topologies (). We obtained a non-resolved polytomy among the individuals captured in Argentina in the monophyletic group with C. erythraeus from China, and the sequence of C. erythraeus flavimanus from Vietnam.
Figure 4. Bayesian inference trees of Callosciurus genus. Tree derived from citochrome oxidase c subunit I (COI). Numbers next to branches are Bayesian posterior probabilities, jackknife support values followed by bootstrap values, respectively. Numbers between brackets on a branch that defines a clade correspond to the groups obtained with the Automatic Barcode Gap Discovery (ABGD) method for the marker.
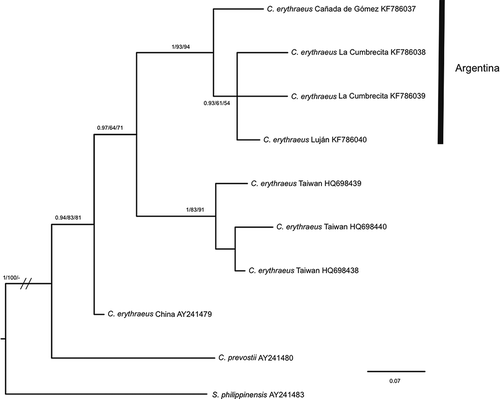
For the RAG1 marker, the sequence alignment comprised 887 bp and we found eight variable sites in the squirrels captured in Argentina, resulting in four haplotypes. The ML, BI and MP phylogenetic trees obtained had similar topologies (). Squirrels captured in Argentina formed a monophyletic group; however, C. erythraeus did not form a monophyletic group and the sequences obtained in Taiwan were closer to the sequences from Argentina than those from China ().
Figure 5. Bayesian inference trees of Callosciurus genus. Tree derived from recombination activating gene I (RAGI). Numbers next to branches are Bayesian posterior probabilities, jackknife support values followed by bootstrap values, respectively.
We estimated the percent sequence divergence amongst taxa for each marker separately and calculated the sequence divergence within and between each species of Callosciurus. Pairwise comparisons of Callosciurus species for D-loop () and Cyt b markers () indicated that the smallest percent sequence divergence amongst species occurred between the specimens identified as C. finlaysonii and squirrels captured in Argentina (5.88% for D-loop; 5.39% for Cyt b). These values were two times smaller than the divergence between C. erythraeus and C. finlaysonii; however, divergence between Cyt b sequences from Argentina and C. erythraeus (10.96%) and C. erythraeus griseimanus (10.89%) was similar to the divergence between C. erythraeus and C. erythraeus griseimanus (10.24%) ().
Table I. Percentage of sequence divergence (Kimura Citation1980) of Callosciurus species for the D-loop marker.
Table II. Percentage of sequence divergence (Kimura Citation1980) of Callosciurus species for the cytochrome b (Cyt b) marker.
For the COI gene (), the smaller divergence (7.34%) was between the squirrels captured in Argentina and C. erythraeus from China, which diverged by 8.8% from C. erythraeus flavimanus from Vietnam. For RAG1 (), we found that the smaller sequence divergence (1.53%) was between the squirrels captured in Argentina and C. erythraeus, which diverged by 2.01% from C. prevostii.
Table III. Percentage of sequence divergence (Kimura Citation1980) of Callosciurus species for the cytochrome oxidase c subunit I (COI) marker.
Table IV. Percentage of sequence divergence (Kimura Citation1980) of Callosciurus species for the recombination activating gene I (RAGI) marker.
The ABGD method for the D-loop data set resulted in seven groups for the recursive partition with prior of 0.001, 0.002, 0.003, 0.005, 0.008, 0.013, 0.022; six groups with 0.036, and two with 0.6 and 0.1. The primary partition was stable on the range of prior values with two groups (). When lowering the relative width of the barcoding gap (X-value) and increasing the prior intraspecific limit, we obtained more groups than the number of species defined by taxonomy. We were not able to identify a barcode gap for the D-loop data set. Using the Cyt b dataset, we obtained one partition (i.e., no barcoding gap) using the standard settings, though eight groups separated when lowering the X-value to 1.1 (see ), with a prior of intraspecific divergence up to 0.0215, that were similar to the number of species defined by taxonomy. For the COI dataset, the number of groups for the recursive partition was six with the entire prior (0.1–0.001). The primary partition was stable on the whole range of prior values, and in all cases we obtained the same six groups indicated in , that were similar to the number of species defined by the taxonomy.
Discussion
The markers, D-loop, COI and RAG1, used in this study indicated that the introduced squirrels from the different invasion foci in Argentina formed a monophyletic group. Together with the description of only one haplotype for D-loop and COI markers obtained in the four foci, our results support the information collected in interviews with local residents that indicated a single introduction event into Argentina followed by translocations into new areas within the country. Management decisions should focus on these invasion pathways to prevent further expansion (Borgnia et al. Citation2013). Variation in pelage colour among squirrels captured in Argentina showed no genetic support. Unexpectedly, squirrels captured in Argentina were more related to C. finlaysonii than to C. erythraeus for D-loop and Cyt b markers. However, intraspecific variation among sequences of C. erythraeus belonging to different subspecies or collected in different regions was large and comparable with the distance to the sequences from Argentina. We could not resolve the relationship between the sequences from Argentina and C. erythraeus using the COI and RAG1 markers.
The classification of the 15 species of the genus Callosciurus is based on morphological features, and also several subspecies have been described on the basis of pelage variations (Moore Citation1961; Moore & Tate Citation1965; Chakraborty Citation1985; Corbet & Hill Citation1992). However, identification of individuals may be difficult given the large number of species and subspecies, many of which closely resemble each other and often show considerable colour variation (Moore & Tate Citation1965; Chakraborty Citation1985; Corbet & Hill Citation1992). The species C. erythraeus and C. finlaysonii have been referred to by Timmins and Duckworth (Citation2008) as the C. erythraeus-C. finlaysonii complex. High sequence divergence within C. erythraeus supports the idea that at least this species could represent a species complex (Clare et al. Citation2007). Regional variation in C. finlaysonii is so extreme that it is doubtful that all these forms constitute discrete species (Corbet & Hill Citation1992), while hybrids may occur in certain regions where both species occur (Moore & Tate Citation1965; Timmins & Duckworth Citation2008). Genetic introgression between subspecies, and even between species, has been reported in other squirrels given the hybridization that frequently occurs in pet markets and stores (Wettstein et al. Citation1995; Good et al. Citation2008; Hird & Sullivan Citation2009; Chang et al. Citation2011). Oshida et al. (Citation2007) and Kuramoto et al. (Citation2012) concluded that the squirrels introduced into Japan that had been identified as C. erythraeus but were genetically closer to C. finlaysonii could be regarded either as C. finlaysonii or as hybrids between both species. However, no studies have yet evaluated whether the present classification of Callosciurus has any genetic support.
Phylogenetic relationships among Callosciurus species indicate three major lineages, one of which is characterized by mainland species (Oshida et al. Citation2011). The Indochina Peninsula lineage contains two sublineages: caniceps-inornatus and erythraeus-finlaysonii, which is composed by C. e. griseimanus, C. erythraeus cf. hendeei and C. finlaysonii (Oshida et al. Citation2013). Interestingly, C. e. griseimanus proved to be more closely related to C. finlaysonii; however, genetic distances among the three forms were very similar, suggesting a polytomic phylogenetic relationship which remained unresolved (Oshida et al. Citation2013). Using the same Cyt b marker, we could not resolve the relationship between these species, as in the previous study by Oshida et al. (Citation2013). However, the monophyletic group formed by C. e. grisemaneus from Vietnam was supported by our analyses (molecular phylogenetic approach and ABGD method) and was used as an independent group to estimate genetic distances. These results indicate that certain subspecies of C. erythraeus could be more closely related to C. finlaysonii than to other conspecifics.
Sequence divergence indicated that the squirrels captured in Argentina were more closely related to C. finlaysonii, when this comparison was possible (D-loop and Cyt b markers), than to C. erythraeus. However, divergence between squirrels captured in Argentina and C. erythraeus was similar to the divergence between subspecies of Callosciurus (e.g. between C. erythraeus and C. erythraeus griseimanus using the Cyt b marker). Sequence divergence using the COI marker also resulted in a similar genetic distance between the squirrels captured in Argentina and C. erythraeus (7%) and between C. erythraeus from China and C. erythraeus flavimanus from Vietnam (9%).
The ABGD method was efficient when intraspecific diversity for the molecular marker was lower than the interspecific diversity, i.e. when sequences sampled within the same species were more similar than sequences sampled from different species. That was the case for COI and Cyt b markers, although with a shorter barcode gap for the latter. Squirrels from Argentina were grouped with C. finlaysonii using the Cyt b marker and were separated from C. erythraeus using the COI marker. C. erythraeus was separated into several groups with both markers, which is consistent with the other analyses. For the D-loop marker, the ABGD method indicated multiple species hypotheses (e.g. one species split into two, or several species merged into a single one). Therefore, the D-loop marker was not entirely consistent in defining species limits using this method due to intra- and interspecific distance overlap. This is a consequence of the high mutation rate for the D-loop marker and a theoretical limitation of the ABGD method and, more generally, of the methods that identify species limits based on the barcode gap, because these methods use the population mutation rate, estimated from the data set, as the main parameter (Puillandre et al. Citation2012a).
The complex taxonomy of certain Callosciurus species and subspecies, such as described for C. erythraeus and C. finlaysonii, and the genetic and morphological data reported in these last decades require a thorough systematic revision. Misidentification in the field cannot be discarded in certain cases. However, identification based on genetic data cannot be fully applied until a simultaneous analysis of diagnostic morphological characters and genetic markers is conducted. These studies will provide new insight regarding Callosciurus’ status in its native area and will aid the study of worldwide invasion pathways of introduction.
Acknowledgements
This study was supported by grants from CONICET (Argentina), International Development Research Centre (Canada), Biodiversity Institute of Ontario/Canadian Centre DNA Barcoding (BIO/CCDB, Canada) and Genome Canada to ML; ANPCyT (Argentina, PICT 32195) and Universidad Nacional de Luján (UNLU, Argentina) to MLG. We thank students of the UNLU for fieldwork assistance, especially Laura Messeta, Sandra Almada and Marina Hertzriken, and Guillermo Cassini and Ignacio Túnez for fruitful discussion and comments. We are especially grateful to Mariana Fasanella and Cecilia Bruno of Centro Regional de Estudios Genómicos (CREG) for assistance in laboratory work.
References
- Aljanabi SM, Martínez I. 1997. Universal and rapid salt-extraction of high quality genomic DNA for PCR based techniques. Nucleic Acids Research 25:4692–4693. doi:10.1093/nar/25.22.4692.
- Aprile G, Chicco D. 1999. Nueva especie exótica de mamífero en la Argentina: la ardilla de vientre rojo (Callosciurus erythraeus). Mastozoología Neotropical 6:7–14.
- Benitez VV, Almada Chávez S, Gozzi AC, Messetta LM, Guichón ML. 2013. Invasion status of Asiatic red-bellied squirrels in Argentina. Mammal Biology 78:164–170.
- Bertolino S. 2009. Animal trade and non-indigenous species introduction: The world-wide spread of squirrels. Diversity and Distributions 15:701–708. doi:10.1111/j.1472-4642.2009.00574.x.
- Bertolino S, Currado I, Mazzoglio PJ. 1999. Finlayson’s (variable) squirrel Callosciurus finlaysonii in Italy. Mammalia 63:522–525.
- Borgnia M, Benitez VV, Gozzi AC, Guichón ML. 2013. La ardilla de vientre rojo en Argentina y el manejo de especies introducidas como un problema biológico y social. Ecología Austral 23:147–155.
- Cassini GH, Guichón ML. 2009. Variaciones morfológicas y diagnosis de la ardilla de vientre rojo Callosciurus erythraeus (Pallas, 1779) en Argentina. Mastozoolgía Neotropical 16:39–47.
- Chakraborty S. 1985. Studies on the genus Callosciurus Gray (Rodentia: Sciuridae). Records of the Zoological Survey of India, Miscellaneous Publications, Occasional Papers, No 63:1–93.
- Chang SW, Oshida T, Endo H, Nguyen ST, Dang CN, Nguyen DX, Jiang X, Li ZJ, Lin LK. 2011. Ancient hybridization and underestimated species diversity in Asian striped squirrels (genus Tamiops): Inference from paternal, maternal and biparental markers. Journal of Zoology 285:128–138. doi:10.1111/j.1469-7998.2011.00822.x.
- Clare EL, Lim BK, Engstrom MD, Eger JL, Hebert PDN. 2007. DNA barcoding of Neotropical bats: Species identification and discovery within Guyana. Molecular Ecology Notes 7:184–190. doi:10.1111/j.1471-8286.2006.01657.x.
- Cooper JK, Sykes G, King S, Cottrill K, Ivanova NV, Hanner R, Ikonomi P. 2007. Species identification in cell culture: A two-pronged molecular approach. In Vitro Cellular and Developmental Biology - Animal 43:344–351. doi:10.1007/s11626-007-9060-2.
- Corbet W, Hill D. 1992. Mammals of the Indomalayan region: A systematic review. London: Oxford University Press.
- Felsenstein J. 1985. Confidence limits on phylogenies: An approach using the bootstrap. Evolution 39:783–791. doi:10.2307/2408678.
- Goloboff PA, Farris JS, Nixon KC. 2008. TNT, a free program for phylogenetic analysis. Cladistics 24:774–786. doi:10.1111/j.1096-0031.2008.00217.x.
- Good JM, Hird S, Reid N, Demboski JR, Steppan SJ, Martin-Nims TR, Sullivan J. 2008. Ancient hybridization and mitochondrial capture between two species of chipmunks. Molecular Ecology 17:1313–1327. doi:10.1111/j.1365-294X.2007.03640.x.
- Guichón ML, Doncaster CP. 2008. Invasion dynamics of an introduced squirrel in Argentina. Ecography 31:211–220. doi:10.1111/j.0906-7590.2008.5308.x.
- Guichón ML, Fasola L, Bello M. 2005. Expansión poblacional de una especie introducida en la Argentina: la ardilla de vientre rojo Callosciurus erythraeus. Mastozoología Neotropical 12:189–197.
- Guo Z, Wang Y, Ran J, Guo C, Li B, Zhang M, Song P. 2011. Genetic structure of Pallas’s squirrel (Callosciurus erythraeus) populations from artificial forests in Hongya County, Sichuan, China. Acta Ecologica Sinica 31:71–77. doi:10.1016/j.chnaes.2010.11.012.
- Hall T. 1999. BioEdit: A user-friendly biological sequence alignment editor and analysis program for Windows 95/98/NT. Nucleic Acid Symposium Series 41:95–98.
- Hird S, Sullivan J. 2009. Assessment of gene flow across a hybrid zone in red-tailed chipmunks (Tamias ruficaudus). Molecular Ecology 18:3097–3109. doi:10.1111/j.1365-294X.2009.04196.x.
- Huelsenbeck JP, Ronquist F. 2001. MRBAYES: Bayesian inference of phylogenetic trees. Bioinformatics 17:754–755. doi:10.1093/bioinformatics/17.8.754.
- Hulme PE, Bacher S, Kenis M, Klotz S, Kühn I, Minchin D, Nentwig W, Olenin S, Panov V, Pergl J, Pyšek P, Roques A, Sol D, Solarz W, Vilà M. 2008. Grasping at the routes of biological invasions: A framework for integrating pathways into policy. Journal of Applied Ecology 45:403–414. doi:10.1111/j.1365-2664.2007.01442.x.
- Ivanova NV, Zemlak TS, Hanner RH, Hebert PDN. 2007. Universal primer cocktails for fish DNA barcoding. Molecular Ecology Notes 7:544–548. doi:10.1111/j.1471-8286.2007.01748.x.
- Jobb G, von Haeseler A, Strimmer K. 2004. TREEFINDER: A powerful graphical analysis environment for molecular phylogenetics. Bio Med Central Evolutionary Biolgy 4:18.
- Keller R, Geist J, Jeschke J, Kühn I. 2011. Invasive species in Europe: Ecology, status, and policy. Environmental Sciences Europe 23:23–17. doi:10.1186/2190-4715-23-23.
- Kimura M. 1980. A simple method for estimating evolutionary rates of base substitutions through comparative studies of nucleotide sequences. Journal of Molecular Evolution 16:111–120. doi:10.1007/BF01731581.
- Kropowski JL, Nandini R. 2008. Global hotspots and knowledge gaps for tree and flying squirrels. Journal of Current Science 95:851–856.
- Kuramoto T, Torii H, Ikeda H, Endo H, Rerkamnuaychoke W, Oshida T. 2012. Mitochondria DNA Sequences of Finlayson’s squirrel found in Hamamatsu, Shizuoka Prefecture, Japan. Mammal Study 37:63–67. doi:10.3106/041.037.0108.
- Lu L, Chesters D, Zhang W, Li G, Ma Y, Ma H, Song X, Wu H, Meng F, Zhu C, Liu Q. 2012. Small mammal investigation in spotted fever focus with DNA-barcoding and taxonomic implications on rodents species from Hainan of China. PLoS One 7:E43479. doi:10.1371/journal.pone.0043479.
- Lurz PWW, Hayssen V, Geissler K, Bertolino S. 2013. Callosciurus erythraeus (Rodentia: Sciuridae). Mammalian Species 902:60–74. doi:10.1644/902.1.
- Moore JC. 1961. Geographical variation in some reproductive characteristics of diurnal squirrels. Bulletin of the American Museum of Natural History 122:1–32.
- Moore JC, Tate GHH. 1965. A study of the diurnal squirrels, Sciurinae, of the Indian and Indo-Chinese subregions. Fieldiana Zoology 48:1–351.
- Nylander, JAA. 2004. MrAIC.pl. Program distributed by the author. Evolutionary Biology Centre, Uppsala University, Sweden.
- Oshida T, Dang CN, Nguyen ST, Nguyen NX, Endo H, Kimura J, Sasaki M, Hayashida A, Takano A, Koyabu D, Hayashi Y. 2013. Phylogenetic position of Callosciurus erythraeus griseimanus from Vietnam in the genus Callosciurus. Mammal Study 38:41–17. doi:10.3106/041.038.0105.
- Oshida T, Dang CN, Nguyen ST, Nguyen NX, Endo H, Kimura J, Sasaki M, Hayashida A, Takano A, Yasuda M, Hayashi Y. 2011. Phylogenetic relationship between Callosciurus caniceps and C. inornatus (Rodentia, Sciuridae): Implications for zoogeographical isolation by the Mekong River. Italian Journal of Zoology 78:328–335. doi:10.1080/11250003.2010.490566.
- Oshida T, Ikeda K, Yamada K, Masuda R. 2001a. Phylogeography of the Japanese giant flying squirrel, Petaurista leucogenys, based on mitochondrial DNA control region sequences. Zoological Science 18:107–114. doi:10.2108/zsj.18.107.
- Oshida T, Lee JK, Lin LK, Chen YJ. 2006. Phylogeography of Pallas’s squirrel in Taiwan: Geographical isolation in an arboreal small mammal. Journal of Mammalogy 87:247–254. doi:10.1644/05-MAMM-A-123R1.1.
- Oshida T, Lin LK, Masuda R, Yoshida MC. 2000. Phylogenetic relationships among Asian species of Petaurista (Rodentia, Sciuridae), inferred from mitochondrial cytochrome b gene sequences. Journal Zoological Science 17:123–128. doi:10.2108/zsj.17.123.
- Oshida T, Shafique MC, Barkati S, Masuda R. 2004. A preliminary study on the genetic diversity of small populations of two flying squirrel species from Pakistan (Hylopetes fimbriatus and Petaurista petaurista albiventer), based on mitochondrial DNA control region sequences. Mammal Study 29:21–26. doi:10.3106/mammalstudy.29.21.
- Oshida T, Torii H, Lin L-K, Lee J-K, Chen Y-J, Endo H, Sasaki M. 2007. A preliminary study on origin of Callosciurus squirrels introduced into Japan. Mammal Study 32:75–82. doi:10.3106/1348-6160(2007)32[75:APSOOO]2.0.CO;2.
- Oshida T, Yasuda M, Endo H, Hussein NA, Masuda R. 2001b. Molecular phylogeny of five squirrel species of the genus Callosciurus (Mammalia, Rodentia) inferred from cytochrome b gene sequences. Mammalia 65:473–482. doi:10.1515/mamm.2001.65.4.473.
- Palmer GH, Koprowski JL, Pernas T. 2008. Tree squirrels as invasive species: Conservation and management implications. In: Witmer GW, Pitt WC, Fagerstone KA, editors. Managing vertebrate invasive species: Proceedings of an international symposium, USDA/APHIS/Wildlife Services. Fort Collins, CO: National Wildlife Research Center. pp. 273–282.
- Puillandre N, Lambert A, Brouillet S, Achaz G. 2012a. ABGD, Automatic Barcode Gap Discovery for primary species delimitation. Molecular Ecology 21:1864–1877. doi:10.1111/j.1365-294X.2011.05239.x.
- Puillandre N, Modica MV, Zhang Y, Sirovich L, Boisselier MC, Cruaud C, Holford M, Samadi S. 2012b. Large-scale species delimitation method for hyperdiverse groups. Molecular Ecology 21:2671–2691. doi:10.1111/j.1365-294X.2012.05559.x.
- Ronquist F, Huelsenbeck JP. 2003. MRBAYES 3: Bayesian phylogenetic inference under mixed models. Bioinformatics 19:1572–1574. doi:10.1093/bioinformatics/btg180.
- Smith MF, Patton JL. 1993. The diversification of South American murid rodents: Evidence from mitochondrial DNA sequence data for the akodontine tribe. Biological Journal of the Linnean Society 50:149–177. doi:10.1111/j.1095-8312.1993.tb00924.x.
- Steppan SJ, Storz BL, Hoffmann RS. 2004. Nuclear DNA phylogeny of the squirrels (Mammalia: Rodentia) and the evolution of arboreality from c-myc and RAG1. Molecular Phylogenetics and Evolution 30:703–719. doi:10.1016/S1055-7903(03)00204-5.
- Thompson JD, Gibson TJ, Plewniak F, Jeanmougin F, Higgins DG. 1997. The CLUSTAL_X windows interface: Flexible strategies for multiple sequence alignment aided by quality analysis tools. Nucleic Acids Research 25:4876–4882. doi:10.1093/nar/25.24.4876.
- Timmins RJ, Duckworth JW. 2008. Diurnal squirrels (Mammalia Rodentia Sciuridae) in Lao PDR: Distribution, status and conservation. Tropical Zoology 21:11–56.
- Vila C, Amorim IR, Leonard JA, Posada D, Castroviejo J, Petrucci-Fonseca F, Crandall KA, Ellegren H, Wayne RK. 1999. Mitochondrial DNA phylogeography and population history of the grey wolf Canis lupus. Molecular Ecology 8:2089–2103. doi:10.1046/j.1365-294x.1999.00825.x.
- Wettstein PJ, Strausbauch M, Lamb T, States J, Chakraborty R, Jin L, Riblet R. 1995. Phylogeny of 6 Sciurus aberti subspecies based on nucleotide sequences of cytochrome b. Molecular Phylogenetics and Evolution 4:150–162. doi:10.1006/mpev.1995.1015.
Appendix
Polymerase chain reaction thermal cycling amplification profiles.a
Species name; locality of collection; GenBank accession numbers of markers; pelage colouration: of the belly (red, orange or creamy), other underparts such as chest, groins or genital area (red, orange or creamy), and the back (black stripe, faint black stripe or no stripe) of the squirrels captured in Argentina, and references of samples used in this study.