
Abstract
Lytta zubovi is a poorly known sexually dimorphic species of blister beetles recently described from Crimea. The female, until now known only from a photograph, is described and the first instar larva obtained from eggs is described and figured using scanning electron microscopy (SEM). Larval morphology greatly supports the previous tentative placement in the genus Lytta.
Introduction
An enigmatic new species of blister beetles, endemic to Crimea, was recently described by Bologna and Nikitsky (Citation2010) as Lytta zubovi and tentatively referred to the Holarctic genus Lytta Fabricius, 1775. Only two males were examined and the female was only known from a photograph, in which it appeared brachyelytrous and possibly apterous.
The placement of this new species in the tribe Lyttini, genus Lytta, was based on the male genitalia and mesosternal structure. Some primitive characters (see Bologna & Nikitsky Citation2010 for details) are shared with the subgenus Poreospasta, an amphi-Pacific element from western North America and central Asia (Selander Citation1960), but the flattened head, pronotum and elytra, the transverse shape of the pronotum and the brachyelytrous condition of the female greatly distinguish this species from all other known Lytta.
The recent collection of two new specimens, including a female, adds to our knowledge of species distribution, ecology and morphology. Additionally, eggs and first instar larvae were obtained from the female. First instar larval morphology is of considerable importance in estimating phylogenetic relationships in Meloidae (Bologna & Pinto Citation2001), and is useful here in helping to determine the tribal and generic placement of L. zubovi.
The primary aims of this paper are to: (a) describe the female morphology; (b) describe the first instar larval morphology; (c) infer phylogenetic relationships of this species.
Materials and methods
One male and one female of L. zubovi were collected on 26 April 2012 (see Results) and both maintained in the laboratory. They were observed during copulation in the field; the female was held in a terrarium and, after oviposition, was killed in 95% alcohol. The mesothoracic muscle of this female was dissected and processed for DNA analysis. Afterwards the specimen was pinned for morphological study and deposited in the Marco A. Bologna collection, Department of Sciences, University Roma Tre. The male specimen is preserved at the Zoological Museum of Moscow, Lomonosov State University.
About 200 eggs were laid (27 April 2012) on leaves of Senecio positioned at the bottom of the rearing terrarium (), and after 12 days (9 May 2012), about 30 first instar larvae hatched and were fixed in 70% alcohol.
Figure 1. Eggs of Lytta zubovi laid on leaves.

Adult morphological analysis was performed using an Olympus SZX12 stereomicroscope equipped with a drawing tube; larval morphology was studied with an Olympus BX51 light Colorview II camera (SIS) and Cell*D image 285 analysis software (SIS) for examining and photographing the cleared specimens mounted on slides in Canada balsam, and an FEI dualbeam FIB/SEM Helios Nanolab for material mounted on stubs after critical point dehydration and gold sputtering. Terminology of larval structures follows MacSwain (Citation1956), Lawrence (Citation1991) and Bologna and Pinto (Citation2001). For some traits of larval chaetotaxy, we followed notational conventions suggested by Selander (Citation1990). Quantitative data presented in the larval description represent means based on three measured, randomly selected specimens.
Results
Faunistics
The first record of this species refers to eastern Crimea, Arabatskaya Strelka (near the Arabatskaya Fortress); the following two new specimens were collected by V. V. Savchuk at Peschanaya Ravine, 3.5 km northeast from Primorskyi settlement (45°07’45.44”N–35°31’43.81”E), which is located ca. 12 km northeast of Feodosia.
Ethology
In our previous paper (Bologna & Nikitsky Citation2010), we described the linear phase of the sexual behaviour. This is confirmed by our new observation (), and is similar to that of other genera of the subfamily Meloinae (see Bologna Citation1991 for a review). During the study of the new pair, a dorsal male posterior phase was observed () but not recorded in detail.
Figure 2. Linear phase of sexual behaviour of Lytta zubovi.

Figure 3. Dorsal phase of sexual behaviour of Lytta zubovi.

Adults are very agile and were observed to move rapidly both in the field and the laboratory. When disturbed, the beetles try to dig rapidly into the litter, which is close to the steppe type. Thanatosis () were also observed. Adults fed () on leaves of herbaceous plants of the genera Senecio (Asteraceae) and Salvia (Lamiaceae) provided in the terrarium by the observers. No information is available on host plants in the field.
Figure 4. Thanatosis behaviour of Lytta zubovi.

Figure 5. Feeding activity of Lytta zubovi.

The female oviposited on the bottom of the rearing terrarium and on the plants positioned in the terrarium (), as is typical in the laboratory. However, we assume that in the field eggs are laid in a hole excavated in the soil, as in other Meloinae. Courtship sporadically took place after oviposition, but no additional oviposition occurred.
Egg and first instar larva description
Egg
Length = 0.85–0.90 mm. Colour pale yellow, elongate, cylindrical, rounded at both ends, enlarged antero-apically, maximum width 250 μm. As in other meloid species (Bologna & Di Giulio Citation2002, Citation2003; Di Giulio et al. Citation2002), a micropylar area is present at the anterior pole of the egg, consisting of a crown of about 20 small, subcircularly arranged micropyles (diameter 30 μm). Each micropyle consists of a sloping V-shaped chorionic flap overhanging a central cavity.
Habitus
Triungulin campodeiform (); body poorly sclerotized, thin and elongate, subparallel sided, with abdomen slightly fusiform. Body length 1.7 mm (from abdominal apex to labrum); head length 0.22 mm (from occipital foramen to clypeolabral suture), maximum width 0.25 mm; diameter of stemmata 16 μm; epicranial stem 53 μm; antennal length 75 μm, antennal seta length 125 μm; prothorax length 0.2 mm, maximum width 0.28 mm; abdominal length 1 mm, maximum width (segments III–IV) 0.27 mm, terminal setae length 0.28 mm; diameter of spiracles: mesothoracic 125 μm, abdominal I 120 μm, abdominal II–VIII respectively from 100 μm to 120 μm. Colour of membranous areas whitish, head, legs and sclerites light brown, distal half of mandibles dark brown. Terga of thorax and abdomen slightly sclerotized and entire; epipleura of thorax and abdomen completely fused with terga displacing spiracles in a dorsal position; sterna of thorax and abdomen membranous, with the exception of abdominal sternum IX and small areas around sternal setae. Cuticle reticulate with transverse polygonal meshes, more evident around terga, discal areas of terga smooth. Tergal setae short.
Figure 6. Lytta zubovi, habitus of first instar larva. (a) Dorsal view; (b) left lateral view; (c) ventral view.

Head
(). Rounded, slightly broader than long, greatest width at the level of the stemmata; sides subparallel, broadly curved in posterior half; basal elevation absent; anterior margin subtruncate, slightly rounded laterally. Epicranial suture Y-shaped, complete to antennal base; basal half of frontal arms parallel, widely diverging apically to antennal insertions. Stemmata small, circular, slightly convex, placed dorsolaterally. Frontoclypeal region with 16 setae; apex of frontoclypeus with one transverse row (frontoclypeal row, FCR) of three pairs of setae: FCR1 similar in length to FCR3, FCR2 longer than others; one additional pair of shorter setae present medially between FCR1; one sensory pit between FCR2 and FCR3. Four pairs of setae posterior to FCR along a curved line paralleling arms of epicranial suture (setae 1–4 from posterior to anterior); one sensory pit present between setae 1 and 2; setae 2–4 subequal, 1 shorter. Basal part of each epicranial plate dorsally with four minute setae and one posterolateral pit arranged in a longitudinal row (basal row, BR) parallel to basal stem of epicranial suture; nine pairs of setae and four pairs of sensory pits anterior to BR: one pair of setae close to epicranial stem with one pair of pits placed anteriorly; one pair of shorter setae anterior to BR; seven pairs of variously sized setae (including ocular seta) and two pits encircling stemmata. Ocular sensory pit slightly anterior to stemma; ocular seta anterior to stemma and to ocular pit. Posterior half of ventral surface of each epicranial plate with one medial sensory pit, anterior half with four setae and four sensory pits arranged as follows: two pairs of setae transversally lined at the level of the cardo and two pairs of setae and three pits (one lateral and two medial, near mandibular acetabulum) placed anteriorly; one large pit placed medially, close to base of maxillae. Labrum (, ) transverse, subrectangular with curved sides, bearing 11 pairs of setae of varying length and one pair of sensory pits.
Figure 7. Lytta zubovi, head of first instar larva. (a) Dorsal view; (b) left lateral view; (c) ventral view; (d) frontal view.
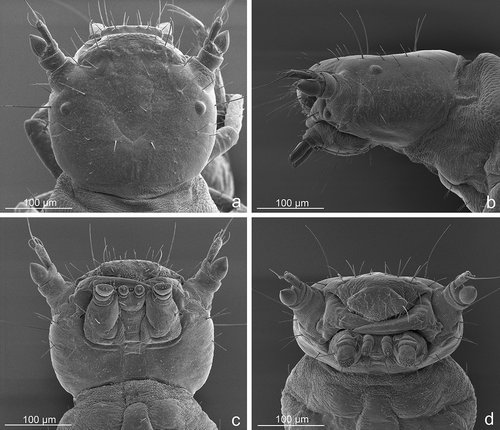
Figure 8. Lytta zubovi, first instar larva. (a) Labrum and mouthparts in frontal view; (b) labrum and mouthparts in ventral view; (c) left antenna in dorsal view; (d) left hind tibiotarsus in dorsal view; (e) metanotum and first abdominal tergum; (f) apex of the abdomen in posterodorsal view.

Antennae () short, directed anterolaterally; antennomere I short, ring-like with one dorsal sensory pit; II slightly shorter than I, distinctly asymmetrical, longer on anterior side, with three elongate setae (two dorsal and one ventral), one minute dorsal seta and one dorsal pit medial to others, near the sensory appendix; sensory appendix well developed, large, bulbous, hyaline and pointed at tip, positioned on outer side of antennomere II; III slender and elongate, cylindrical, slightly clavate, about as long as I and II together, with a long apical seta (less than three times as long as entire antenna), three elongate, subequal, subapical setae, two lateral (one on outer and one on inner sides) and one dorsal, curved towards apical one; one minute seta near base of apical seta. Mandibles (, ) robust, conical-falcate, basal half broad and slightly sclerotized, apical half strongly sclerotized, narrowing and abruptly bending inward, ental margin medially keeled and with two edges: ventral edge smooth; dorsal edge with eight triangular, upcurved, toothlike emarginations; outer margin of mandible with two setae, one sensory pit between them and one pit mesodorsally. Maxillae with stipes about as long as wide and bearing two rows of setae: anterior row with two long setae and one medial pit; posterior row with two shorter setae and one pit between them; mala simple, lobiform, slightly protruding, with 7–8 spiniform setae, none of which is obviously stouter than others; cardo transverse, subrectangular, with one short lateral seta; maxillary palpomeres I and II () short, I only slightly longer and wider; I with one ventral sensory pit; II with two subequal setae, one dorsal on outer side and one small ventral; III subrectangular, subparallel-sided, slightly narrower than II, dorso-ventrally flattened and spoon-like (slightly concave dorsally), 1.8 times the length of I and II together, with one basal and dorsolateral seta (inner side) and one ventrolateral pit (outer side); apex of palpomere III convex and membranous, with a sensorial area composed of about 30 conical and subequal sensilla, one medial, larger and cylindrical with ring-like base, and one very small close to medial; outer side of palpomere III with one slender digitiform sensillum. Gula without setae; submentum, mentum and prementum poorly sclerotized; submentum with two setae; mentum with two shorter setae and four pits; prementum with four setae, two short and basal and two longer and apical. Labial palpomeres () narrow and cylindrical, palpomere I slightly asymmetric (outer side longer) and one small ventral seta; palpomere II about twice as long as I, with a lateral pit and an apical circular slightly swollen sensorial area composed of 10 conical and subequal sensilla, one larger medially placed and cylindrical with large ring-like base, and one very small close to medial.
Thorax
Segments transverse, slightly broader than head; prothorax slightly wider than meso- and metathorax; margins of each thoracic segment rounded. Ecdysial line complete on pronotum, incomplete on mesonotum and absent on metanotum. Anterior margin of pronotum membranous; each half of pronotum with 12 short setae and five pits symmetrically placed along three transverse, subparallel rows; anterior row (AR) with four setae and four pits; medial row (MR) irregular, with four setae and one pit; posterior row (PR) with four setae and two pits; prosternum with three pairs of medial setae arranged longitudinally and one pair anterior. Mesonotum subrectangular, slightly narrower and shorter than pronotum, with AR composed of four pairs of setae; MR with two lateral setae and one pit; PR with four setae and two pits; three pairs of medial setae on mesosternum, anterior pair extremely short. Metanotum () subequal to mesonotum, more rounded on sides; setae of metathorax similar in number, position and relative dimensions to those of mesothorax.
Legs
Slender, without lanceolate setae: coxa conical and elongate, with one small apical seta, four elongate medial setae, transversally arranged, gradually decreasing in length from base to apex, three minute basal setae and one pit; trochanter with four apical setae and five pits; femur not enlarged in the middle, slightly shorter than tibiotarsus, with six setae and one pit, longest ventral femoral seta much shorter than femur; fore femora slightly more robust and shorter than others; tibiotarsi and claws increasing in length from pro- to metathorax; tibiotarsi slightly tapered at apex and with five longitudinal rows of 5–7 spinelike, moderately long setae; claw () conical-falcate, thin, acute and slightly curved at apex, with two setae of different length inserted at different levels near base (apical distinctly longer than basal).
Spiracles
(, ). Round, internally papillate, with a small opening. Peritreme round and flat, slightly elevated and with about 10 projections encircling opening. Mesothoracic spiracle anterolateral in position, abdominal spiracles medial in position, all spiracles distinctly dorsal and subequal in diameter.
Abdomen
Slightly fusiform, approximately 1.8 times as long as thorax; segments II–VI subequal, maximum width at III; segment IX smallest and suboval; sternal abdominal area membranous, except for sternum IX. Tergites distinctly transverse, with rounded sides, with three transverse rows of setae on each half of tergum as follows: AR with three minute setae (four on tergum I, ) and one pit; MR with three setae at level of spiracle, two medial and one lateral (long in segments II–VIII); PR with seven setae of various size and one pit. AR and MR of tergum IX with same setation as segments II–VIII, PR composed of five elongate setae, longer than those of segments II–VIII, and one pair of very long setae (caudal setae), about as long as the last two abdominal segments combined. AR and MR of sterna I–IX with one pair of minute medial setae each, PR with four pairs of setae (three pairs on segment I), two outer pairs (one pair on segment I) very long (except on segment IX). Pleurites completely fused with tergites, displacing spiracles dorsally. Segment X membranous (), divided by a transverse anal fold in two parts: dorsal part semicircular with six extremely small setae transversally arranged, ventral part (pygopod) longitudinally divided into two lobes, moderately produced.
Female morphology
The description of L. zubovi (Bologna & Nikitsky Citation2010) included only mention of the sexual dimorphism of this species, without a detailed account of female morphology.
The habitus of the female is represented in and . The female is similar to the male except for the following morphological details: (1) antennomere III visibly longer than IV; (2) median line of pronotum indistinct; (3) base of pronotum with very indistinct sinuosity medially; (4) scutellar setae brownish; (5) metasternum shorter; (6) elytra short, covering less than the anterior half of the abdomen, last four abdominal tergites not covered, terga distinctly rugose; (7) metathoracic wings absent; (8) foretarsi with ventral pad of longer and denser setae; (9) metatibiae slightly curved as mesotibiae; (10) posterior margin of penultimate sternite greatly curved and emarginated; (11) apical sternite with rounded posterior margin.
Discussion
The placement of the Crimean species in the tribe Lyttini, genus Lytta, is well supported by larval morphology; thus, any suspicion about a possible assignment to Meloini is resolved.
The larva definitely belongs to the tribe Lyttini based on the combination of the following morphological characters, considered diagnostic by MacSwain (Citation1956): (1) epicranial suture complete to the antennal base; (2) very short antennomere II; (3) sensory appendix well developed on antennomere II; (4) terminal antennal seta more than twice as long as the entire antenna; (5) absence of modified seta at apex of mala; (6) maxillary palpomere III subparallel sided; (7) fusion between pleurites and tergites dorsally displacing the abdominal spiracles.
In particular, this larva has in common with other Lytta several characters such as: (1) head rounded, slightly wider than long; (2) weakly sclerotized gula; (3) boundary between clypeus and frons marked by a row of eight setae; (4) antennal sensory appendix bulbous, large, apically pointed; (5) tergites with posterior marginal row of 10 setae; (6) spiracle on abdominal segment I subequal to mesothoracic spiracle; (7) abdominal spiracles II–VIII subequal to I. However, it is worth noting the following characters that differ from the known Lytta larvae and that possibly support a distinct subgeneric assignment: (1) first antennal segment longer than the second (it is shorter than the second in Lytta); (2) antennal seta less than 3 times longer than the entire antenna (in Lytta 3–6 times longer than entire antenna); (3) ecdysial line complete on pronotum, incomplete on mesonotum and absent on metanotum (it is complete on all thoracic segments and on first abdominal tergite in Lytta, except L. vescicatoria, see MacSwain Citation1956); (4) abdominal sterna VIII and IX membranous, unsclerotized (sclerotized in Lytta).
Preliminary molecular data also support the placement of L. zubovi among Lyttini. In fact, in the framework of a molecular phylogenetic study of the tribe Lyttini (Pitzalis et al., in preparation), we compared the sequence of 16S mtDNA of L. zubovi with that of 65 other taxa (Bologna Citation1991; Bologna & Pinto Citation2001; Bologna et al. Citation2008). In all trees, L. zubovi is included in the clade of the genus Lytta, as the sister of L. (Poreospasta) auriculata Horn, 1870.
The form of sexual dimorphism in L. zubovi is characterized by brachyelytrous and apterous females, and holelytrous, fully winged males. Additionally, the female is considerably larger than the male. Although uncommon in the Lyttini, brachyelytry and complete absence of hind wings, believed to be independently evolved, do characterize a single other species of Lytta, one Lydomorphus Fairmaire, 1882, the genera Berberomeloe Bologna, 1989, Trichomeloe Reitter, 1911 and Parameloe Denier, 1933, as well as several additional genera in other lineages of Meloidae (see e.g. Bologna Citation1991; Bologna & Pinto 2002; Bologna & Nikitsky Citation2010 for a wider discussion). In the last three genera, however, both sexes share these reduced features. The only other meloid group with the same degree of sexual dimorphism in elytral and hind wing development as occurs in L. zubovi is in the North American genus Cordylospasta.
Due to its distinct adult morphology, we suspect that L. zubovi should eventually be referred to a new subgenus of Lytta. However, such action will require the study of a large number of species from all subgenera, particularly those from central and eastern Asia.
Acknowledgements
We thank V. V. Savchuk, Primorskyi settlement, Crimea, who collected the new specimens of Lytta zubovi and sent them to one of us (NBN), and N. M. Kaygorodova from Krym, who first discovered these beetles. We are indebted to our colleague J. D. Pinto (Riverside, CA, USA) who carefully revised the English and provided many significant suggestions. The electron microscopy analyses were performed at the Laboratorio Interdipartimentale di Microscopia Elettronica (LIME) of the University “Roma Tre”, Rome, Italy.
References
- Bologna MA. 1991. Coleoptera Meloidae. Fauna d’Italia. XXVIII. Bologna: Calderini.
- Bologna MA, Di Giulio A. 2002. Review of the Southern Africa genus Prolytta Kaszab, with a description of the first instar larva, and bionomic and taxonomic remarks (Coleoptera, Meloidae). Invertebrate Systematics 16:177–194. doi:10.1071/IT01017.
- Bologna MA, Di Giulio A. 2003. Egg and first-instar larval morphology of Prionotolytta binotata (Péringuey, 1888), an endemic southern African species (Coleoptera, Meloidae). African Entomology 11:213–219.
- Bologna MA, Nikitsky NB. 2010. A new enigmatic dimorphic Lytta from Crimea, Ukraine (Coleoptera: Meloidae). Zoosystematica Rossica 19:70–76.
- Bologna MA, Oliverio M, Pitzalis M, Mariottini P. 2008. Phylogeny and evolutionary history of the blister beetles (Coleoptera, Meloidae). Molecular Phylogenetics and Evolution 48:679–693. doi:10.1016/j.ympev.2008.04.019.
- Bologna MA, Pinto JD. 2001. Phylogenetic studies of Meloidae (Coleoptera), with emphasis on the evolution of phoresy. Systematic Entomology 26:33–72. doi:10.1046/j.1365-3113.2001.00132.x.
- Bologna MA, Pinto JD. 2002. The Old World genera of Meloidae (Coleoptera): a key and synopsis. Journal of Natural History 36:2013–2102.
- Di Giulio A, Bologna MA, Pinto JD. 2002. Larval morphology of Meloe subgenus Mesomeloe: Inferences on its phylogenetic position and a first instar larval key to the Meloe subgenera (Coleoptera, Meloidae). The Italian Journal of Zoology 69:339–344. doi:10.1080/11250000209356479.
- Lawrence JF. 1991. Order Coleoptera. In: Stehr FW, editor. Immature insects. Vol. 2. Dubuque, Iowa: Kendall/Hunt. pp. 144–298.
- MacSwain JW. 1956. A classification of the first instar larvae of the Meloidae (Coleoptera). University of California Publications in Entomology 12:i–iv, 1–182.
- Selander RB. 1960. Bionomics, systematics, and phylogeny of Lytta, a genus of blister beetles (Coleoptera, Meloidae). Illinois Biological Monographs 28:i–vi, 1–295.
- Selander RB. 1990. Blister beetles of the genus Pseudopyrota (Coleoptera: Meloidae). Journal of the Kansas Entomological Society 63:46–79.