
Abstract
Calathus cinctus Motschulsky, 1850 was only recently separated from the group Calathus melanocephalus (Linnaeus, 1758). C. cinctus has now been recorded for the first time in Croatia, along the temporary Krčić River situated beneath Mt. Dinara. However, revision of museum collections revealed that this species also occurs at other localities in Croatia, in the Alpine region and along the Adriatic coast. Its sister species C. melanocephalus, as the type species of the group, is widely distributed in Croatia, whereas C. mollis Marsham, 1802 is mostly restricted to the coastal region, with a few records from Alpine and Continental Croatia. The distribution range of C. cinctus overlaps with the distribution ranges of its sister species. Life-history traits were studied using pitfall traps in a karst region of Croatia, including eu-Mediterranean and sub-Mediterranean zones, in riparian and karst habitats, and on agricultural land. Based on goodness-of-fit criteria, the zero-inflated negative binomial model and negative binomial model best described the distribution of C. cinctus. For both models, there was a statistically significant difference between habitat types for activity density, indicating that C. cinctus has a preference for open habitats. It was recorded above a range of soil types (e.g. red, brown, sandy, cultivated), with sparse vegetation, suggesting that sufficient sun exposure is a more important requirement than soil type. The seasonal dynamics of C. cinctus differed between eu-Mediterranean and sub-Mediterranean zones, despite the rather close proximity of the studied localities. Geographical variability of seasonal activity was most likely due to differences in climate, altitude and the vicinity of the high mountain. Calathus cinctus populations in both zones were wing-dimorphic, with brachypterous specimens prevailing in both populations, indicating population stability.
Introduction
Calathus cinctus (synonyms: Calathus ruficollis Gautier des Cottes, 1868 and Calathus erythroderus Gemminger & Harold, 1868) belongs to the genus Calathus Bonelli, 1810 and subgenus Neocalathus Ball & Nègre, 1972 (Löbl & Smetana Citation2003). The genus has mainly a Holarctic distribution, and is species rich including several endemic species (Ruiz et al. Citation2010). Calathus cinctus was recently separated from the C. melanocephalus (Linnaeus, 1758) group that was comprised of three species: C. cinctus, C. mollis Marsham, 1802 and C. melanocephalus, the latter being the type species (Aukema Citation1990). Previously, C. cinctus was considered a subspecies of C. mollis (Lindroth Citation1945). The taxonomic status of this group was questioned several times in the past due to the frequent occurrence of morphologically intermediate specimens (see Aukema Citation1990). However, cross-breeding results showed that these species are reproductively isolated (Aukema Citation1990).
All three species are distributed in the Palaearctic region, having a mainly European distribution (Löbl & Smetana Citation2003). Calathus melanocephalus has the widest distribution range of all three species and occurs not only in Europe, but also in North Africa and Southwest Asia (up to the Caucasus and east to Trans-Baikal in Siberia) (Aukema Citation1990; Löbl & Smetana Citation2003). According to the present knowledge, C. mollis has a wider distribution range than previously documented (Aukema Citation1990; Löbl & Smetana Citation2003). Although it occurs in most European countries, it is mainly confined to coastal areas of the Atlantic and the Mediterranean (Aukema Citation1990; Anderson et al. Citation2000; Löbl & Smetana Citation2003). It also occurs along the Mediterranean coast of North Africa and the Middle East (Löbl & Smetana Citation2003). Calathus cinctus has a more restricted distribution range than its two sister species. It has been recorded in several European countries, ranging from Great Britain and Portugal in the west to Latvia and Greece in the east. It appears that C. cinctus does not occur as far north the other two species, but in the Mediterranean area it extends farther south, from Morocco to Syria (Aukema Citation1990; Löbl & Smetana Citation2003).
In northern and central Europe, C. cinctus and both its sister species prefer open habitats with sparse vegetation exposed to sun (Thiele Citation1977; Aukema Citation1990; Lindroth Citation1992a). They occur in dry grasslands, heaths, dunes, meadows, pastures, gardens and arable lands (e.g. Thiele Citation1977; Lindroth Citation1992a; Anderson et al. Citation2000; Luff Citation2002) and often achieve their greatest abundance on sandy or gravel-mixed soil (Lindroth Citation1992a).
Life-history traits are appropriate indicators of habitat changes (e.g. fragmentation, habitat loss) and as such are useful tools in applied conservation biology (Henle et al. Citation2004; Barbaro & Van Halder Citation2009). Calathus cinctus life-history traits, including daily and annual rhythms, reproduction, feeding and dispersal ability, are well known in northern Europe (Aukema Citation1990), while there are no data from the Mediterranean area. As previously documented for other carabid beetle species, species phenology is often regionally specific (e.g. Dülge Citation1994; Matalin Citation2006; Kádár et al. Citation2015); thus, regional data are required in order to validate generalisations based on life-history traits.
The objectives of this paper are: (1) to present the first finding of C. cinctus and its distribution in Croatia, together with the distribution of its sister species; and (2) to present further data on C. cinctus life-history traits (e.g. habitat preferences, seasonal dynamics, sex ratio, dispersal abilities). Distribution data of the C. melanocephalus group are based on literature data, and museum and private collections. Life-history traits of C. cinctus are documented from wet and karst habitats located in the eu-Mediterranean and sub-Mediterranean zones of Croatia. These regions are geographically close though they differ in terms of climate and vegetation (Zaninović et al. Citation2008; Vukelić Citation2012).
Materials and methods
Study area
The study was conducted in the karst region of Croatia, in three different riparian and adjacent habitats: around the Krčić River (44°02ʹ20.2“N; 16°15ʹ53.9”E), around Vrana Lake (43°57ʹ35.4“N; 15°33ʹ34.8”E) and in the Nin saltpan (44°14ʹ34.49“N; 15°11ʹ25.4”E). All localities are situated in the Mediterranean region, Krčić River in the sub-Mediterranean zone of deciduous vegetation and the two others in the eu-Mediterranean zone of evergreen vegetation. The climate around Vrana Lake and the Nin saltpan is a typical Mediterranean climate with hot summers, while the area of Krčić River has a temperate humid climate with hot summers (Šegota & Filipčić Citation2003).
The Krčić River is situated beneath Mt. Dinara, at an altitude of 370 m. It is a temporary river that typically dries up during the hot summer and early autumn months. The catchment area of the Krčić River covers 157 km2, while the length of its surface flow is 10.5 km. It is characterised by both surface and subsurface flow dynamics and ends with a 40-m-high waterfall falling into the Krka River (Bonacci et al. Citation2006).
Vrana Lake, with an area of 30 km2, is the largest coastal lake in Dalmatia, protected as a nature park. The lake is a karst cryptodepression that interacts with the sea and ground waters. The elevation of its surface is only 0.1 m. Although Vrana Lake is separated from the Adriatic Sea by a limestone ridge (800–2500 m wide), it is specifically influenced by the sea since it is directly connected to it by the 800 m long Prosika canal (Katalinić et al. Citation2008).
The Nin saltpan is located in the shallow lagoon of Nin Bay, covering an area of 0.55 km2. This area was previously a salt marsh, though saltpans, or salt evaporation ponds, were created over time for the extraction of salt from seawater. These salt ponds are separated by earthen levees to regulate water levels that vary during the year, depending on the phase of salt production, and are productive resting and feeding grounds for many water bird species.
Sampling and identification
Calathus cinctus was collected using pitfall traps, together with other soil arthropods. Soil fauna was sampled along the course of the Krčić River in two main habitat types: riparian and karst (). Riparian habitats were located parallel to the river course, while karst habitats were located at least 30 m away from riparian habitats. Within each habitat type, 36 sites were selected; each was sampled with three pitfall traps, placed 5 m apart in a linear transect. In total, 216 pitfall traps were set. The traps were partially filled with a 3:2 solution of ethylene glycol and water. To reduce the surface tension of the liquid, a drop of unscented detergent was added. A dark roof was placed above each pitfall trap to protect them from rain and litter. Samples were collected monthly from April to late November in 2014. The collected specimens were kept in 75% ethyl alcohol.
Near Vrana Lake, C. cinctus was sampled by pitfall traps in riparian (reed beds and wet meadows) and karst habitats (Mediterranean shrublands, agricultural lands and Aleppo pine forests; ). In each habitat type, three sites were selected, with the exception of reed beds and wet meadows, which were sampled with four sites (in total 17 sites). At each site, five pitfall traps were arranged in a transect line at 5-m intervals. In total, 85 pitfall traps were set. The traps were filled with a solution of wine vinegar, salt and water. A roof was placed above each trap. Soil fauna was sampled monthly from January to December in 2007. The collected specimens were pinned or kept dry.
Table I. Vegetation composition and habitat description of the studied habitats along the Krčić River, Vrana Lake and the Nin saltpan (Croatia).
In the Nin saltpan, soil fauna was collected using pitfall traps in three habitats: dikes, levees and dry salt pools (). Within each habitat type, four sites were selected, each sampled with five pitfall traps, placed 5 m apart in a transect line. In total, 60 pitfall traps were set. The traps were filled with a solution of wine vinegar, salt and water. Samples were collected monthly from April to August 2011. At this time, sampling was stopped as the saltpan is an important nesting area for the black-winged stilt [Himantopus himantopus (Linnaeus, 1758)] and adults began to feed on the soil invertebrates from the pitfall traps. All collected specimens were pinned or kept dry.
Calathus cinctus was identified using Aukema (Citation1990) and Lompe (Citation2015). Nomenclature follows Löbl and Smetana (Citation2003). Voucher specimens are deposited in the first author’s collection (Department of Biology, Faculty of Science, Zagreb, Croatia) and the Coleopteran collection (National Museum Zadar, Croatia).
Vegetation analysis
Vegetation of the studied habitats was represented with phytosociological relevés, made in accordance with standard Central European methodology (Braun-Blanquet Citation1964). Plant nomenclature follows Flora Europaea (Tutin et al. Citation1964–1980, Citation1993).
Data analyses
Distribution data analyses
Data on the distribution of C. cinctus and its sister species were obtained from field studies, the literature and entomological collections. For each record, the following data were provided: locality and in brackets the name of the geographical region (e.g. Slavonia, Gorski Kotar, Istria, Primorje, Lika, Dalmatia and Dalmatian Zagora), date, year and data source. For certain records exact dates are missing, since they were not provided in the cited literature. In these cases, the year of publication was used as the date. Data sources include the literature citation and name of the entomological collection. Entomological collections of the Zadar National Museum (NMZ, Coleopteran collection), Split Natural History Museum (NHM, Novak Collection), and first author’s coleopteran collection were reviewed. The distribution data are presented and discussed in terms of three biogeographical regions: Alpine, Continental and Mediterranean (according to EEA Citation2015). Additionally, the data are classified in two categories: old and new records, the latter including records following the publication of the most recent catalogue of the carabid beetles of Croatia (Drovenik & Peks Citation1999). All records were georeferenced and distribution maps created in ArcGIS 10.2 (ESRI Citation2012).
Life-history trait analyses
Dominant species analysis was determined according to Tietze (Citation1973). Species represented by > 5% in the assemblage are considered dominant. Seasonal activity was calculated on the number of specimens collected during one year. Since the number of pitfall traps and the sampling length varied between studied sites, the catch was standardised as the number of specimens per trap and per day. This was presented for two localities (Krčić River and Vrana Lake), while at the Nin saltpan it could not be shown due to the low number of sampled individuals. Activity density was compared between habitats, separately for each locality. This comparison was complicated due to the high number of samples with zero C. cinctus individuals, which made routine statistical tests inappropriate. To overcome excess-zero problems, we modelled observed count data using generalised linear models. A few models were tested to find the best fit for C. cinctus data, as there is no general model that would fit overdispersed soil animal data (Sileshi Citation2008). We used models assumed to be applicable for count data with excess zeros: Poisson model, zero-inflated Poisson model (ZIP; Lambert Citation1992), zero-inflated negative binomial model (ZINB; Martin et al. Citation2005) and negative binomial model (NBD; Lawless Citation1987). Goodness of fit was tested by Akaike’s information criterion (AIC; Johnson & Omland Citation2004). The activity density between habitats was then compared using likelihood ratio statistics. Pairwise comparisons were done for least squares means, using adjustment for multiple comparisons. Models were run using the SAS University Edition (SAS Institute Inc Citation2015) with the GLM procedure.
Sex of C. cinctus was determined on the basis of tarsus shape and genitalia. Sex ratio was compared using the nonparametric sign test run in Statistica 12 (StatSoft Citation2013). Ageing was based on elytral hardness and colour (van Dijk Citation1979). In this study, only two age categories were distinguished: tenerals and old adults. Teneral adults were recently hatched beetles, easily recognised by weak chitinisation and yellow-brownish elytra. Old adults were characterised by brown elytra and hard chitinisation. Flight ability was determined by hind wing development and all collected specimens were checked for hind wings. Two groups were recognised: short-winged (brachypterous) and long-winged (macropterous) specimens. Additionally, we measured elytron length (EL) and greatest wing length (WL). All proportions were measured using an eyepiece micrometer on a Zeiss Stemi 2000 microscope. Measurements were done according to Aukema (Citation1990). A total of 50 specimens (40 brachypterous and 10 macropterous) from the Krčić area and 46 specimens (40 brachypterous and six macropterous) from Vrana Lake were measured. Differences in wing length (elytron and hind wings) between locations and sexes were tested using Mann–Whitney U test. The selection of the test depended on the normality data distribution, tested with the Shapiro–Wilk W test. These tests were performed in Statistica 12 (Statsoft Inc Citation2013).
Results and discussion
Distribution of Calathus cinctus and its sister species C. melanocephalus and C. mollis in Croatia
Calathus cinctus was recorded for the first time in Croatia along the river bed of the temporary Krčić River in 2014. It was not listed in the previous carabid beetle checklist (Drovenik & Peks Citation1999) or Catalogue of Palaearctic Coleoptera (Löbl & Smetana Citation2003) for Croatia. Additionally, revision of the Coleopteran collection of the Zadar National Museum revealed the occurrence of this species in other parts of Croatia (; Appendix 1). It was found in two biogeographical regions of Croatia: Alpine and Mediterranean (Appendix 1). In total, it was recorded at nine localities, with most records confined to the Mediterranean region. It was mainly found at lower altitudes up to 400 m, with the exception of the Alpine region, where it was recorded at 1060 m.
Figure 1. Distribution of the Calathus melanocephalus group in Croatia: (a) Calathus cinctus, (b) Calathus melanocephalus and (c) Calathus mollis. New records include data since 1999.
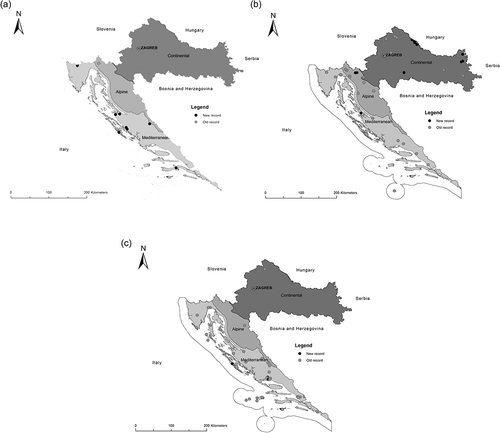
Calathus melanocephalus is present in all three biogeographical regions of Croatia: Alpine, Continental and Mediterranean (; Appendix 1). In total, it has been recorded at 31 localities and most findings were confined to the Continental biogeographical region. According to old literature data, this species was widespread through Dalmatia (Schlosser Klekovski Citation1875-77; Apfelbeck Citation1904; Sahlberg Citation1912-1913) and Slavonia (Schlosser Klekovski Citation1875-77). Additionally, it has been found on several mountains (e.g. Risnjak, Snježnik, Učka, Velebit; Appendix 1). However, in the previously cited literature, precise localities were not given. Interestingly, it was also recorded on the peak of Mt. Biokovo, (peak Sveti Jure, 1762 m; Vujčić-Karlo & Durbešić Citation2002). According to Aukema (Citation1990), it is the only species of this group to occur at higher altitudes in the mountains.
Calathus mollis is present in all three biogeographical regions of Croatia: Alpine, Continental and Mediterranean (; Appendix 1). Most records have been confined to the Mediterranean region (35 localities), predominantly in southern Dalmatia, along the Adriatic coast and islands (Appendix 1). However, findings from other biogeographical regions were scarce. Similarly, this species is often abundant along the Atlantic coast in northern Europe (e.g. Lindroth Citation1992a; Anderson et al. Citation2000; Luff Citation2007; Desender et al. Citation2008) and along the Mediterranean coast (Ganglbauer Citation1892; du Chatenet Citation2005). It has primarily been found at lower altitudes, with the exception of the Plitvice Lakes (Alpine biogeographical region) at 560 m.
In Croatia, the distribution range of C. cinctus overlaps with the distribution ranges of its sister species, particularly in the Mediterranean and Alpine regions. Sympatry and mixed populations of C. cinctus and C. mollis were observed at the Vrana Lake sites. These findings are in accordance with observations from northern Europe where mixed populations were also observed (Aukema Citation1990).
Life history traits of Calathus cinctus
Habitat preferences
In total, 119 specimens of C. cinctus were sampled along the river bed of the temporary Krčić River. The majority of specimens were recorded in the karst habitats (90 specimens), while only 29 specimens were caught in the riparian habitats (). Dominant species analysis showed that in karst habitats, C. cinctus belonged to the class of dominant species (8.63% of the total catch), while in riparian habitats it was rare with a frequency of ≤ 1% (A. Brigić, unpub. data). Based on goodness-of-fit criteria (I), the ZINB and NBD model best described the C. cinctus distribution. For both models, there was a statistically significant difference between two habitat types (karst and riparian habitats) in activity density (likelihood ratio test, NBD model: χ2(1) = 4.01, p < 0.05; ZINB model: χ2(1) = 4.11, p < 0.05). The Poisson model gave the worst fit to the data (), corroborating Sileshi (Citation2008).
Table II. Akaike’s information criterion (AIC) and Bayesian information criterion (BIC) for the tested models with statistical significance of differences between karst and riparian habitats (p value) of the Krčić River and Vrana Lake. For the description of habitat types see . AIC and BIC are in the form “smaller is better”. Legend: ZIP: zero inflated Poisson model; ZINB: zero-inflated negative binomial model; NBD: negative binomial model.
In the surroundings of Vrana Lake, a total of 200 specimens of C. cinctus were caught. Most specimens were collected in arable land, e.g. 131 specimens in olive groves and 40 specimens in vineyards, followed by 17 specimens recorded in the Mediterranean shrublands. Similarly to riparian habitats of the Krčić River, C. cinctus was sporadically found in riparian habitats of Vrana Lake, e.g. six specimens in wet meadows and two specimens in reed beds. Goodness-of-fit criteria () showed that the ZINB and NBD models best described the C. cinctus distribution, similarly to the data from the Krčić River. Habitat type was a statistically significant predictor for activity densities (likelihood ratio test, NBD model: χ2(4) = 53.27, p < 0.001; ZINB model: χ2(4) = 53.27, p < 0.001). Pairwise comparisons (done for ZINB and NBD models) revealed significant differences in activity density between agricultural land and other habitats: wet meadows (p < 0.0001), Aleppo pine forest (p < 0.001), reed beds (p < 0.0001) and Mediterranean shrublands (p < 0.0001). Therefore, comparison of activity density via models for both locations clearly showed that C. cinctus is mainly a xerophilous species, preferring open thermophilous shrublands, pastures and arable land. These results are in accordance with Aukema (Citation1990) and Luka et al. (Citation2009).
In the Nin saltpan, only 23 specimens of C. cinctus were recorded. Most specimens were found on higher dikes (21 specimens), while a single specimen was recorded from the leeves directly above the salt pool during inundation. There were insufficient data to present the seasonal dynamic in the saltpan (catches: 14 specimens in May, eight in June and one in July) due to the large population of black-winged stilt nesting on the leeves, since adults, after some time, began to feed on soil invertebrates from the pitfall traps. Therefore, the trapping period ended before the end of summer and the expected autumn peak of this species’ activity was not recorded.
Overall, C. cinctus was recorded on different soil types along the Krčić River and around Vrana Lake (). Most individuals were collected above terra rosa and dry brown soil forming shallow cover over limestone bedrock. Occasionally, it was also found above sandy soils. Similarly, C. cinctus was recorded above different soil types in northern Europe (Aukema Citation1990). Summarising the data on habitat properties (), the most important predictor of C. cinctus habitat selection could be sun exposure, making the soil type a less relevant factor.
Seasonal activity
In both study areas, C. cinctus showed two activity peaks: in spring and in late autumn (). In the sub-Mediterranean zone (Krčić River), the highest surface activity of beetles was observed in June and was associated with mature beetle activity and young beetle emergence. Generally, mature beetles were recorded throughout the season, while tenerals appeared in May (five individuals) and in June (11 individuals), the latter in accordance with Aukema (Citation1990). After the spring peak, the number of beetles strongly decreased, with no beetles collected in August. The second peak of activity was detected in November and was determined by the activity of old beetles. Females dominated the catches along the Krčić River (76 females: 43 males) and the sex ratio differed significantly (sign test, p < 0.001). Overall, females were more active than males during the whole vegetation season.
Figure 2. Seasonal activity of Calathus cinctus in the sub-Mediterranean (Krčić River) and in the eu-Mediterranean (Vrana Lake) zones. Monthly catches in pitfall traps were pooled in both studied regions.
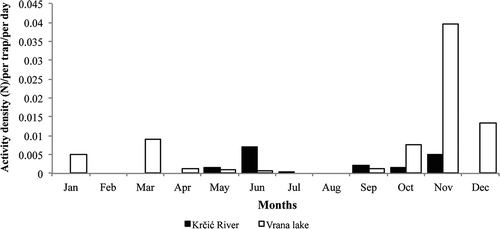
In the eu-Mediterranean zone (Vrana Lake), two peaks of activity were observed: at the end of winter and in autumn (). The first peak of activity was already observed in March and was determined by mature beetle activity. After this peak, the number of beetles strongly decreased. Teneral adults were observed in April (two individuals) and in May (two individuals). Aestivation is likely to begin in July, since no beetles were collected during July and August. The highest surface activity of beetles was observed in November and was exclusively associated with the activity of mature beetles, while in Switzerland the autumn maximum was observed in October (Luka et al. Citation2009). The sex ratio did not differ significantly (102 females: 98 males; sign test, p > 0.05). Males were more active in January, May, September and November, while females were more active in March, April, June and October, implying that C. cinctus is an autumn breeder, as previously documented by Aukema (Citation1990).
The present study showed that C. cinctus seasonal activity differed between the sub-Mediterranean and eu-Mediterranean zones, despite the close geographical proximity of the localities (less than 55 km air distance). Differences in seasonal patterns are highly likely to be due to the type of climate, the altitude and the vicinity of Mt. Dinara. Both climates are characterised by hot summers, with average maximum air temperatures as high as 41°C (Šegota & Filipčić Citation2003; Zaninović et al. Citation2008). In the sub-Mediterranean zone, the winters are cold, with a minimum average air temperature as low as −15°C, while in the eu-Mediterranean zone the air temperatures rarely fall below 0°C (Zaninović et al. Citation2008). Additionally, weather conditions in the sub-Mediterranean zone are also highly influenced by the vicinity of Mt. Dinara (Prelovšek Citation2010). Therefore, climatic conditions are more harsh and extreme in the sub-Mediterranean than in the eu-Mediterranean zone, causing a strong decrease in surface activity of C. cinctus and other carabid beetles in late autumn (A. Brigić, unpub. data), corroborating findings of Filippov (Citation2006) for C. melanocephalus. Contrary to that, mild winter weather conditions enable surface activity and overwintering of C. cinctus mature adults in the eu-Mediterranean zone. Furthermore, our study showed that seasonal activity patterns of C. cinctus differ between northern (Aukema Citation1990) and southern populations, with seasonal activity beginning earlier in southern Europe. Similar patterns have been observed for other carabid beetle species, e.g. Carabus coriaceus Linné, 1758 (Dülge Citation1994; Kádár et al. Citation2015), Carabus germarii Sturm, 1815 (Paill Citation2000; Kádár et al. Citation2015) and Pterostichus strenuus Panzer, 1796 (Dülge Citation1994). The life cycle of some carabid beetle species may last 2 years (Matalin Citation2007), indicating that our data sets have some limitations. Therefore, in future studies, it would be interesting to focus on activity patterns of C. cinctus in a few consecutive years and to rear adults and larvae under different laboratory conditions, in order to estimate the duration and occurrence of individual development phases.
Flight abilities
Calathus cinctus populations both in the eu-Mediterranean and in the sub-Mediterranean zones are wing-dimorphic, having short- (brachypterous) and long- (macropterous) winged specimens (I), which corroborates results from northern Europe (Aukema Citation1990). In total, 111 specimens of C. cinctus recorded in the sub-Mediterranean zone (Krčić River) were brachypterous, while only eight specimens were macropterous (two males and six females). Similarly, brachypterous specimens (194 individuals) prevailed over macropterous (six individuals) in the eu-Mediterranean zone (Vrana Lake). Aukema (Citation1990) demonstrated that C. cinctus shows a simple Mendelian inheritance pattern of wing morphology, with brachyptery dominant over macroptery. The relatively low proportion of macropterous specimens in both studied populations (6.72% in the Krčić area and 3.00% around Vrana Lake) suggests that these populations are stable and long established (Den Boer Citation1977; Lindroth Citation1992b).
Wing length in short-winged specimens was about 0.45 times the length of the elytron, while in long-winged specimens it was about 1.5 times the elytron length. Elytron and wing length differed significantly in size between the sexes (Mann–Whitney U test, elytron: U = 500.00, p < 0.001; wing length: U = 613.00, p < 0.001), while differences between locations were not significant (elytron: U = 940.00, p > 0.05; wing length: U = 1017.50, p > 0.05). Generally, females had longer wings than males and macropterous specimens had longer elytra than brachypterous specimens (), which is in accordance with the findings of Aukema (Citation1990). Hence, in comparison with this cited study, it was observed that both brachypterous and macropterous specimens (of both sexes) had longer wings in southern populations compared to populations in northern Europe, implying that C. cinctus individuals are longer in the Western Balkans than in northern Europe. A similar pattern has been observed for other carabid beetle species, e.g. Pterostichus rhaeticus (Brigić et al. Citation2014). In northern Europe, C. cinctus is considered a good disperser (Den Boer Citation1977). According to the present knowledge, its dispersal ability is unclear in the Mediterranean area, due to the high frequency of brachypterous specimens and no actual flight data, and remains to be tested in future studies, using other trapping methods (e.g. window traps or Malaise traps).
Table III. Measurements (mm) of short- and long-winged males and females of Calathus cinctus from the sub-Mediterranean (Krčić River) and eu-Mediterranean (Vrana Lake) area. Legend: SW: short winged; LW: longed winged; n: number of measured individuals; EL: elytron length; WL: maximum wing length; SD: standard deviation.
In conclusion, our study shows that C. cinctus is a common species in the Mediterranean area. It prefers open habitats, with sparse vegetation and sufficient insolation. The seasonal activity patterns are region-specific, governed by both a north–south gradient and regional climate conditions. Future studies should focus on dispersal abilities of C. cinctus, and the importance of wing morphology in (re-)establishment of new populations, particularly in unstable temporary river environments.
Acknowledgements
The authors would like to thank three anonymous reviewers for improving the manuscript, Bože Kokan for providing access to the Novak Collection (Split National History Museum), Zorana Sedlar for her assistance with ArcGIS and Linda Zanella for proofreading the English. Additionally, we would like to thank Vladimir Bartovsky, Gordan Lukač and Morana Bačić for assistance in field sampling. This study was supported by the University of Zagreb (Nos 202310, 202675 to Ivančica Ternjej) and by Zadar County and the Vrana Lake Nature Park (Snježana Vujčić-Karlo).
Additional information
Funding
References
- Anderson R., McFerran D., Cameron A. 2000. The Ground Beetles of Northern Ireland. Atlases of the Northern Ireland Flora and Fauna. Vol. 1. Ulster Museum. 246 pp.
- Apfelbeck V. 1904. Die Käferfauna der Balkanhalbinseln, mit Berücksichtigung Klein-Asiens und der Insel Kreta. I Band. Berlin: Friedländer und Sohn Verlag. pp. 1–422.
- Aukema B. 1990. Taxonomy, life history and distribution of three closely related species of the genus Calathus (Coleoptera: Carabidae). Tijdschift Voor Entomologie 133:121–141.
- Barbaro L, Van Halder I. 2009. Linking bird, carabid beetle and butterfly life-history traits to habitat fragmentation in mosaic landscapes. Ecography 32:321–333. DOI:10.1111/eco.2009.32.issue-2.
- Bonacci O, Jukić D, Ljubenkov I. 2006. Definition of catchment area in karst: Case of the rivers Krčić and Krka, Croatia. Hydrological Sciences Journal 51:682–699. DOI:10.1623/hysj.51.4.682.
- Braun-Blanquet J. 1964. Pflanzensoziologie: Grundzüge der Vegetationskunde. Wien: Verlag Eugen Ulmer. pp. 1–865.
- Bregović A. 1985. Zbirka karabida Entomološkog odjela Gradskog muzeja Varaždin. Godišnjak Gradskog Muzeja Varaždin 7:221–248.
- Brigić A. 2012. Carabid beetle (Coleoptera, Carabidae) assemblages across beech-fir forest - open habitat ecotones in Croatian Dinaric Alps. Doctoral thesis. Zagreb: Faculty of Science. pp. 1–209.
- Brigić A, Vujčić-Karlo S, Alegro A, Šegota V, Ternjej I. 2014. Ecology, biology and conservation of Pterostichus rhaeticus Heer, 1837 (Coleoptera: Carabidae) at the edge of its distribution range, in the Western Balkans. Italian Journal of Zoology 81:517–529. DOI:10.1080/11250003.2014.947338.
- Bušnja I. 2008. Trčci (Carabidae, Coleoptera) Zbirka Košćec iz Gradskog muzeja Varaždin. In: Šicel M, Spevec B, editors. Franjo Košćec i njegovo djelo 1882–1968. Varaždin: Hrvatska akademija znanosti i umjetnosti. pp. 173–193.
- Den Boer PJ. 1977. Dispersal power and survival. Carabids in cultivated countryside. Miscellaneous Papers Landbouwhogeschool Wageningen 14:1–190.
- Depoli G. 1929. I Coleopteridella liburnia, Parte I: Adephaga – Palpicornia. Fiume: Rivistadella Societta di Studifiumani. pp. 1–338.
- Desender K, Dekoninck W, Maes D, Crevecoeur L, Dufrêne M, Jacobs M, Lambrechts J, Pollet M, Stassen E, Thys N. 2008. Een nieuwe verspreidingsatlas van de Loopkevers en Zandloopkevers (Carabidae) in Belgiё. Brussels: Institut voor Natuur-en Bosonderzoek and Koninklijk Belgisch Instituut voor Natuurwetenschappen. pp. 1–184.
- Domić D. 2009. Seasonal activity of ground beetles (Coleoptera, Carabidae) in Kopački rit with particular overview on the bioindicator species of contamination. Master Thesis. Osijek: University Josip Juraj Strossmayer Osijek. pp. 1–103.
- Drovenik B, Peks H. 1999. Catalogus faune. Carabiden der Balkanländer (Coleoptera, Carabidae). Schwanfelder Coleoterologische Mitteilungen 1:1–103.
- du Chatenet G. 2005. Coléoptéres d’Europe. Carabes, Carabiques et Dytiques. Volume I Adephaga. N.A.P. Verriěres le Buisson: Editions Verriěres le Buisson. pp. 1–40.
- Dülge R. 1994. Seasonal activity of carabid beetles in wooded habitats in northwest Germany. In: Desender K, Dufrêne M, Loreau M, Luff ML, Maelfait JP, editors. Carabid Beetles: Ecology and Evolution. Dordrecht: Kluwer Academic Publishers. pp. 125–131.
- Durbešić P, Vujčić-Karlo S, Šerić-Jelaska L, Pintarić K. 2006. Abundance and seasonal dynamics of arthropods in the meadow community Arrhenatheretum elatioris near Varaždin, Croatia. Periodicum Biologorum 108:3–10.
- EEA. 2015. Biogeographical regions dataset of European environment agency. Available: http://www.eea.europa.eu/data-and-maps/data/biogeographical-regions-europe-2. Accessed Nov 2015.
- ESRI. 2012. ArcGIS desktop: Release 10.2. Redlands, CA: Environmental Systems Research Institute.
- Filippov BY. 2006. Seasonal aspects of life cycles in Calathus melanochephalus and C. micropterus (Coleoptera, Carabidae) in the Northern Taiga. Entomological Review 86:901–909. DOI:10.1134/S0013873806080057.
- Ganglbauer L. 1892. Die Käfer von Mitteleuropa I. Wien: Carl Gerold’s Sohn Verlag. pp. 1–557.
- Gjurašin B. 2001. Katalog tvrdokrilaca (Insecta, Coleoptera) zbirke „Koča“. Natura Croatica 10:1–187.
- Henle K, Davies KF, Kleyer M, Margules C, Settele J. 2004. Predictors of species sensitivity to fragmentation. Biodiversity and Conservation 13:207–251. DOI:10.1023/B:BIOC.0000004319.91643.9e.
- Hoffmann A. 1926. Coleopterologisches aus dem Dalmatiner Karst. Entomologischer Anzeiger (1921-1936) 6:76–78.
- Johnson JB, Omland KS. 2004. Model selection in ecology and evolution. Trends in Ecology & Evolution 19:101–108. DOI:10.1016/j.tree.2003.10.013.
- Kádár F, Fazekas JP, Sárospataki M, Lövei GL. 2015. Seasonal dynamics, age structure and reproduction of four Carabus species (Coleoptera: Carabidae) living in forested landscapes in Hungary. Acta Zollogica Academiae Scientiarum Hungaricae 61:57–72. DOI:10.17109/AZH.61.1.57.2015.
- Katalinić A, Rubinić J, Bušelić G. 2008. Hydrology of two coastal karst cryptodepressions in Croatia: Vrana Lake vs Vrana Lake. In: Sengupta M, Dalwani R, editors. Proceedings of the 12th World Lake Conference (Taal 2007). Jaipur: Ministry of Environment & Forests, Government of India. 732–743.
- Koča GJ. 1905. Popis tvrdokrilaca (Kornjaša) Vinkovačke okolice. Prilog fauni Hrvatske i Slavonije. Zagreb: Glasnik Hrvatskog naravoslovnog društva. pp. 1–96.
- Kovačević Ž. 1958. Entomološka istraživanja u Nacionalnom parku „Plitvička jezera“. Zagreb: Nacionalni park „Plitvička jezera“. pp. 401–408.
- Kučinić M, Perović F, Vujčić-Karlo S. 1996. Entomološka istraživanja donjeg toka rijeke Neretve. Elaborat, preliminarno izvješće. Zagreb: Hrvatski prirodoslovni muzej. pp. 1–70.
- Lambert D. 1992. Zero-inflated poisson regression, with an application to defects in manufacturing. Technometrics 34:1–14. DOI:10.2307/1269547.
- Langhoffer A. 1886-1897. Prilozi entomološkoj fauni Hrvatske. Kornjaši Hrvatske. Rad Jugoslavenske Akademije 141:50–81.
- Lawless JF. 1987. Negative binomial and mixed poisson regression. Canadian Journal of Statistics 15:209–225. DOI:10.2307/3314912.
- Lindroth CH. 1945. Die Fennoskandischen Carabidea. Ein tiergeografische Studie. 1. Eine tiergeografische Studie. 1. Spezieller Teil. - Göteborgs Kungliga Vetenskaps-och Vitterhetssamhälles Handlingar 6:1–709.
- Lindroth CH. 1992a. Ground beetles (Carabidae) of Fennoscandia: A zoogeographic study, Specific knowledge regarding the species, Part I. Washington D.C: Smithsonian Institution Libraries and the National Science Foundation. pp. 1–630.
- Lindroth CH. 1992b. Ground beetles (Carabidae) of Fennoscandia: a zoogeographic study, General analysis with a discussion on biogeographic principles, Part III. Washington D.C: Smithsonian Institution Libraries and the National Science Foundation. pp. 1–814.
- Löbl I, Smetana A. 2003. Catalogue of Palaearctic Coleoptera, Vol. 1. Stenstrup: Apollo Books. 819 pp.
- Lompe A. 2015. Gattung Calathus. Available: http://www.coleo-net.de/coleo/texte/calathus.htm. Accessed Nov 2015.
- Luff ML. 2002. Carabid assemblage organization and species composition. In: Holland JM, editor. The agroecology of carabid beetles. Andover: Intercept Ltd. pp. 41–80.
- Luff ML. 2007. The Carabidae (ground beetles) of Britain and Ireland, Handbooks for the identification of British insects. St. Albans: Royal Entomological Society. pp. 1–247.
- Luka H, Marggi W, Huber C, Goneseth Y, Nagel P. 2009. Coleoptera, Carabidae: Ecology, atlas, Fauna Helvetica 24. Neuchâtel: Schweizerische Entomologische Gesellschaft. 677 pp.
- Marcuzzi G. 1986. Contributions to the knowledge of coleopterous fauna of Dalmatia. Rad Jugoslavenske Akademije Znanosti I Umjetnosti 424:191–237.
- Martin TG, Wintel BA, Rhodes JR, Kuhnert PM, Field SA, Low-Choy SJ, Tyre A, Possingham HP. 2005. Zero tolerance ecology: Improving ecological inference by modelling the source of zero observations. Ecology Letters 8:1235–1246. DOI:10.1111/j.1461-0248.2005.00826.x.
- Matalin AV. 2006. Geographic variability of the life cycle in Pterostichus melanarius (Coleoptera, Carabidae). Entomological Review 86:409–422. DOI:10.1134/S0013873806040051.
- Matalin AV. 2007. Typology of life cycles of ground beetles (Coleoptera, Carabidae) in Western Palaearctic. Entomological Review 87:947–972. DOI:10.1134/S0013873807080027.
- Müller G. 1922. Materiali per una fauna colleotterologica della isole e degli scogli dell’Adriatico. Liburnia Rivista trimestrale della Sezione di Fiume del Club Alpino Italiano 16:56–75.
- Müller J. 1914. Berücksichtigung der Fauna von Lissa, Curzola und Pelagosa. In: Ginzberger A, editor. Beiträge zur Naturgeschichte der Scoglien und kleineren Inseln Süddalmatiens. Ergebnisse von zwei im Mai und Juni 1911 und im Juli 1914 unter-Stützung aus der Erbschaft Trietl ausgeführten Reisen. Wien: Kaiserlich-Koniglichen Hof- und Staatsdruckerei. pp. 370–392.
- Novak P. 1952. Kornjaši Jadranskog primorja. Zagreb: Jugoslavenska akademija znanosti i umjetnosti. pp. 1–251.
- Novak P. 1964. I Coleotteridella Dalmazia. Trieste: Atti del Museo Civico di Storia Naturale 24. pp. 53–132.
- Novak P. 1970. Rezultati istraživanja kornjaša našeg otočja. Jugoslavenska akademija znanosti i umjetnosti 38:5–58.
- Paill W. 2000. Slugs as prey for larvae and imagines of Carabus violaceus (Coleoptera: Carabidae). In: Brandmayr P, Lövei GL, Zetto Brandmayr T, Casale A, Vigna Taglianti A, editors. Natural History and Applied Ecology of carabid Beetles. Sofia–Moscow: Pensoft. pp. 221–227.
- Prelovšek M. 2010. Climate. In: Mihevc A, Prelovšek M, Zupan Hajna N, editors. Introduction to the Dinaric Karst. Postojna: Karst Research Institute at ZRC SAZU. pp. 20–24.
- Ruiz C, Jordal BH, Emerson BC, Will KW, Serrano J. 2010. Molecular phylogeny and Holartic diversification of the subtribe Calathina (Coleoptera: Carabidae: Sphodrini). Molecular Phylogenetics and Evolution 55:358–371. DOI:10.1016/j.ympev.2009.10.026.
- Sahlberg J 1912-1913. Coleoptera Balcanica. Finska Vetenskaps-Societetens Főrhandligarrhandligar, Bd. LV, No. 15. pp. 1–108.
- SAS Institute Inc. 2015. Cary, NC: SAS University Edition.
- Schlosser Klekovski K. 1875-77. Fauna kornjaša trojedne kraljevine. Zagreb: Jugoslavenska akademija znanosti i umjetnosti. pp. 1–342.
- Šegota T, Filipčić A. 2003. Köpenova podjela klima i hrvatsko nazivlje. Geoadria 8:17–37.
- Sileshi G. 2008. The excess-zero problem in soil animal count data and choice of appropriate models for statistical inference. Pedobiologia 52:1–17. DOI:10.1016/j.pedobi.2007.11.003.
- Statsoft Inc. 2013. Statistica (Date Analysis Software System), Version 12. Available: http://www.statsoft.com/. Accessed Nov 2015.
- Tallósi B. 2008. Population-level baseline surveying and preparative investigations for the monitoring of carabid beetles (Coleoptera, Carabidae) in areas along the Drava river and in Baranja (Croatia). In: Purger JJ, editor. Biodiversity studies along the Drava river. Pecs, Hungary: University of Pecs. pp. 165–220.
- Thiele HU. 1977. Carabid beetles in their environments. Berlin: Springer-Verlag. 369 pp.
- Tietze F. 1973. Zur ökologie, Soziologie und Phänologie der Laufkäfer (Coleoptera, Carabidae) des Grünlandes im Süden der DDR II, 2. Teil: Die diagnostisch wichtigen Carabidenarten des untersuchten Grünlandes und ihre Verbreitungsschwerpunkte. Hercynia 10:111–126.
- Tutin TG, Burges NA, Chater AO, Edmondson JR, Heywood VH, Moore DM, Valentine DH, Walters SM, Webb DA, editors. 1993. Flora Europaea 1. Cambridge: Cambridge University Press.
- Tutin TG, Heywood VH, Burges NA, Moore DM, Valentine DH, Walters SM, Webb DA, editors. 1964–1980. Flora Europaea 1–5. Cambridge: Cambridge University Press.
- van Dijk T. 1979. On the relationship between egg production, age and survival in two carabid beetles: Calathus melanocephalus L. and Pterostichus coerulescens L. (Coleoptera, Carabidae). Oecologia 40:63–80. DOI:10.1007/BF00388811.
- Vujčić-Karlo S, Durbešić P. 2002. Visinska distribucija trčaka (Carabidae) na Biokovu. In: Kerovec M, Durbešić P, editors. Prirodoslovna istraživanja Biokovskog područja, II. Zagreb: Hrvatsko ekološko društvo. pp. 255–265.
- Vukelić J. 2012. Šumska vegetacija Hrvatske. Zagreb: Šumarski fakultet Sveučilišta u Zagrebu i Državni zavod za zaštitu prirode. pp. 1–398.
- Zaninović K, Gajić-Čapka M, Perčec Tadić M, Vučetić M, Milković J, Bajić A, Cindrić K, Cvitan L, Katušin Z, Kaučić D, Likso T, Lončar E, Lončar Ž, Mihajlović D, Pandžić K, Patarčić M, Srnec L, Vučetić V. 2008. Climate Atlas of Croatia: 1961-1990, 1971-2000. Zagreb: Meteorological and Hydrological Service of Croatia.
Appendix 1
Distribution of Calathus cinctus and its sister species Calathus melanocephalus and Calathus mollis in Croatia. Legend: AB Collection: Andreja Brigić Collection; CNHM: Croatian Natural History Museum (Zagreb); NMZ: Zadar National Museum; VCM: Varaždin City Museum; NHM: Natural History Museum (Split).