Abstract
Fine roots (<2 mm) are very dynamic and play a key role in forest ecosystem carbon and nutrient cycling and accumulation. We reviewed root biomass data of three main European tree species European beech, (Fagus sylvatica L.), Norway spruce (Picea abies L. Karst.) and Scots pine (Pinus sylvestris L.), in order to identify the differences between species, and within and between vegetation zones, and to show the relationships between root biomass and the climatic, site and stand factors. The collected literature consisted of data from 36 beech, 71 spruce and 43 pine stands. The mean fine root biomass of beech was 389 g m−2, and that of spruce and pine 297 g m−2 and 277 g m−2, respectively. Data from pine stands supported the hypothesis that root biomass is higher in the temperate than in the boreal zone. The results indicated that the root biomass of deciduous trees is higher than that of conifers. The correlations between root biomass and site fertility characteristics seemed to be species specific. There was no correlation between soil acidity and root biomass. Beech fine root biomass decreased with stand age whereas pine root biomass increased with stand age. Fine root biomass at tree level correlated better than stand level root biomass with stand characteristics. The results showed that there exists a strong relationship between the fine root biomass and the above-ground biomass.
Introduction
In forest ecosystems the below-ground carbon pool often exceeds the above-ground pool. Root systems account for 10 – 45% of the total tree stand carbon pool, and most of it is allocated to stumps and coarse roots (Santantonio et al., Citation1977; Fogel, Citation1983; Helmisaari et al., Citation2002). Fine root (<2 mm) biomass is relatively small (Keyes & Grier, Citation1981; Vogt et al., Citation1996); nevertheless fine roots are very dynamic and play a key role in forest ecosystem carbon and nutrient cycling and accumulation (Berg, Citation1984; Joslin & Henderson, Citation1987; Hendrick & Pregitzer, Citation1993; Helmisaari et al., Citation2002). There is less information about root system carbon pools and it is more difficult to generalize than that of the above-ground pools, mainly due to methodological difficulties in determining the fine root biomass, labor-intensive nature of such studies, and variety of internal and external factors affecting stand level root biomass (e.g. Persson, Citation1983; Vogt & Persson, Citation1991).
In previous reviews fine root biomass has been related to several abiotic and biotic factors such as soil properties, tree species, stand characteristics, and climatic factors (Jackson et al., Citation1996; Vogt et al., Citation1986, Citation1996; Cairns et al., Citation1997; Leuschner & Hertel, Citation2003; Chen et al., Citation2004). In most cases, fine root biomass and the abiotic or biotic factors have not been correlated or only weakly correlated when datasets from different sources have been included. The relationships have been often clearer and stronger in studies carried out with the same species, within a limited geographical area, or in controlled experiments. Based on the earlier findings the following hypotheses can be presented.
-
Tree fine root biomass is higher in the temperate than in the boreal zone (Vogt et al., Citation1986; Jackson et al., Citation1996).
-
Deciduous trees have a higher fine root biomass than conifer trees in the temperate zone (Vogt et al., Citation1986, Citation1996; Leuschner & Hertel, Citation2003).
-
The higher the soil fertility the lower the fine root biomass (Keyes & Grier, Citation1981; Vogt et al., Citation1983, Citation1986, Citation1987; Jackson et al., Citation1996; Finér & Laine, Citation1998; Helmisaari et al., Citation2007), even though there can be species-specific differences (Leuschner & Hertel, Citation2003).
-
The higher the soil acidity the lower the fine root biomass, but there can be species-specific differences and differences related to the length of exposure to the acid deposition (Jentschke et al., Citation2001; Godbold et al., Citation2003; Leuschner & Hertel, Citation2003; Leuschner et al., Citation2004).
-
The fine root biomass increases until canopy closure, thereafter it does not increase or decrease (Vogt et al., Citation1983; Vanninen et al., Citation1996; Helmisaari et al., Citation2002; Claus & George, Citation2005), but there can be species specific variation from this pattern (Leuschner & Hertel, Citation2003).
-
Fine root biomass at tree level correlates better than the stand level fine root biomass with the stand characteristics (Chen et al., Citation2004; Helmisaari et al., Citation2007).
Material and methods
Collection of root biomass data
The data collected for this review had to fulfill the following criteria:
-
Only published data were used.
-
We limited our data set to common European tree species, for which large number of published information on fine root biomass exists. These species were European beech (Fagus sylvatica L.), Norway spruce (Picea abies L. Karst.) and Scots pine (Pinus sylvestris L.). Data from very young stands (age < 10 years) were not included. The data sets included stands of Norway spruce and Scots pine from temperate to boreal zones and beech stands only from the temperate zone.
-
Fine root biomass was determined with the coring method. The data had been collected at one sampling event or represented means of several seasonal or annual samplings. The number of core samples and the sampling depth varied between studies. It was assumed that the sampling covered the depth of majority of fine root biomass.
-
Only fine root biomass < 2 mm in diameter was included. Living (biomass) and dead roots (necromass) were treated separately.
-
Fine root data from stands or plots where soil manipulations had been done were excluded.
-
Sites were classified into three fertility classes: poor, medium, rich. The classification was in some cases done already in original papers (e.g. Leuschner & Hertel, Citation2003) or it was done by the authors using the information found in the original papers.
-
The following site characteristics were collected from the original papers: the location of the site, elevation, mean annual precipitation, mean annual air temperature, topsoil pH, and topsoil C/N ratio. The following tree stand characteristics were recorded: tree stand age, height, mean diameter, density, volume and basal area.
The data basis
The collected material consisted of data from 36 beech, 71 Norway spruce and 43 Scots pine stands ( – , ). In addition, 18 of the beech stands, 20 of the spruce, and 8 pine stands were included from the review of Leuschner & Hertel (Citation2003) concerning tree stands in the temperate zone. All beech stands were located in the temperate zone within a narrow longitudinal gradient, but there was substantial variation in the climatic, tree, and soil characteristics between these stands. There were spruce and pine stands from both temperate and boreal zones with large variation in the climatic, tree, and soil characteristics between the stands. There were spruce stands also at high elevations whereas the pine stands were mainly located in lowlands. All site characteristics were not available from all stands.
Figure 1. The location of reviewed stands. The stands located in Finland, Estonia, Norway and Sweden were classified in the boreal zone.
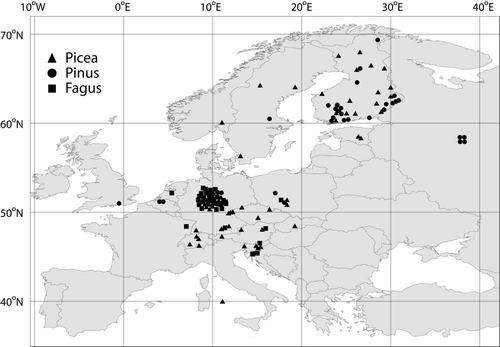
Table I. Sources of data
Table II. Descriptive statistics for the beech, spruce and pine stands in the temperate zone
Table III. Descriptive statistics for the spruce and pine stands in the boreal zone
Statistical tests
One way Anova was used to test the differences between species and vegetation zones and the paired comparisons were done with Tukey's test. Pearson correlation coefficients were calculated between fine root biomass and climatic, tree stand and soil characteristics of the stand, and linear regressions were fit to the data. The analyses were done with the Anova, Correlation, and Regression procedures of the SPSS® Base for Windows 14.0 statistical package.
Results
Stand level fine root biomass
The mean living beech fine root biomass was 389 ± 206 g m−2, that of spruce 297 ± 143 g m−2, and that of pine 277 ± 150 g m−2. The necromasses were 920 ± 948 g m−2, 190 ± 95 g m−2, and 133 ± 100 g, respectively. In the temperate zone the fine root biomass of beech was significantly higher than that of spruce (p = 0.015; ). The fine root biomass of spruce and pine did not differ from each other either in the temperate zone or in the boreal zone ( and ). For spruce the fine root biomass did not differ between the temperate and boreal zones, but for pine it was significantly (p = 0.002) higher in the temperate zone than in the boreal zone.
There was a significant positive correlation between beech and pine fine root biomass and latitude in the temperate zone (). Among pine stands the variation in latitude was only 1° (), and most of the beech data were from a narrow latitude range without any latitudinal trend in fine root biomass. There were no significant correlations between the fine root biomass and the elevation of the stand in either of the vegetation zones.
Table IV. The Pearson correlation coefficients between the fine root biomass (g m2) and the climatic, site and stand factors.a
There was a significant positive correlation between the mean annual precipitation and the fine root biomass of spruce in the boreal zone (), but most of the data were from a narrow precipitation range without any relationship between the amount of precipitation and the fine root biomass.
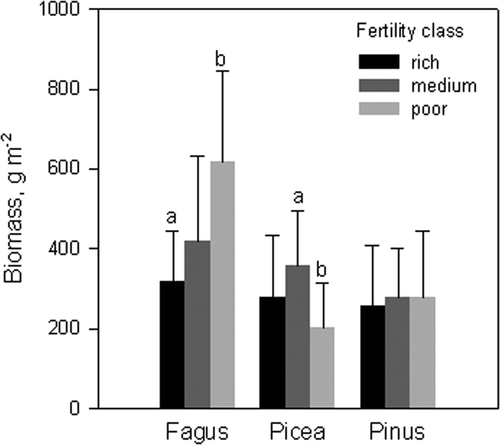
In beech (p = 0.08, N = 35) and spruce (p = 0.007, N = 70) stands there was a significant difference in the fine root biomasses between different fertility classes, but no such differences existed in pine stands (). The fine root biomass of spruce stands was lowest in the poorest sites, whereas in beech stands it was the highest in the poorest sites.
Figure 2. Mean fine root biomass in different site fertility classes for beech, spruce and pine stands. Error bars indicate standard deviations, and in each species group letters show the fertility classes which differ significantly from each other according to the Tukey's test (p < 0.05).
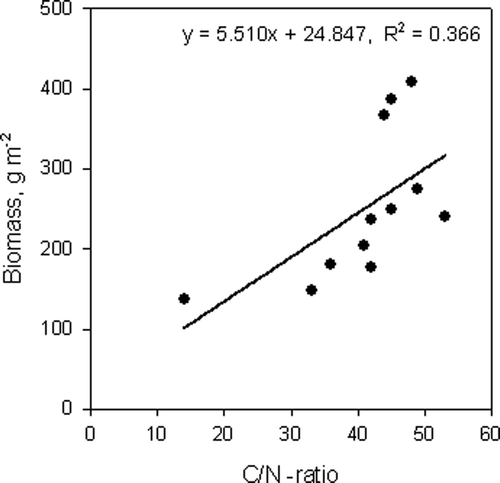
The mineral soil acidity did not correlate with the fine root biomass (). However, there was a significant relationship between pine fine root biomass and the C/N ratio in the soil organic layer ().
Figure 3. Relationship between C/N ratio of organic soil layer and fine root biomass for Scots pine stands.
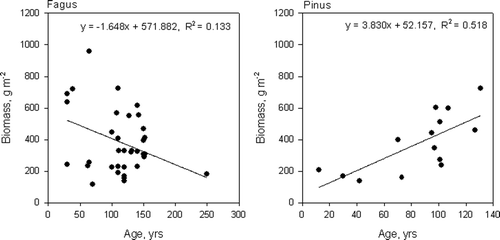
The fine root biomass of beech decreased with stand age and that of pine increased with stand age in the temperate zone (). There was one exceptionally old beech stand in the data. However, the relationship between beech stand age and fine root biomass changed only slightly if that stand was excluded from the analysis (y = −1.705 + 577.416, p = 0.053).
Figure 4. Relationship between stand age and fine root biomass in the temperate zone for beech and Scots pine stands.
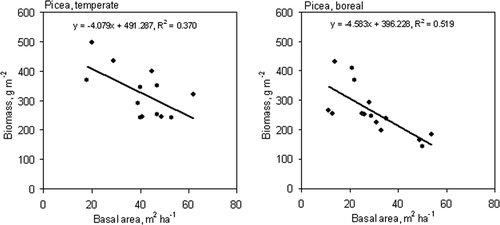
The fine root biomass of spruce decreased with the increase in the basal area of the stand in both vegetation zones (). At the stand level, there were no other significant correlations between the fine root biomass and the stand characteristics.
Figure 5. Relationship between stand basal area and fine root biomass for spruce in the temperate and boreal zones.
Root biomass at tree level
The fine root biomass per tree was calculated by dividing the fine root biomass per hectare with the stem number per hectare. It was significantly higher for beech than for spruce (p = 0.001) or pine (p = 0.013) ( and ). The fine root biomass per tree did not differ between spruce or pine in either of the zones. The ratio of the stand basal area to the fine root biomass did not differ significantly between studied tree species (p = 0.681).
The root biomass per tree increased with stand age in beech and in spruce stands in the temperate zone and in pine stands in both vegetation zones (, ). The fine root biomass per tree increased with basal area per tree in beech and in spruce stands in the temperate zone and in pine stands in the boreal zone ().
Figure 6. Relationship between stand age and fine root biomass per tree for beech and spruce in the temperate zone and for Scots pine in the temperate and the boreal zone.
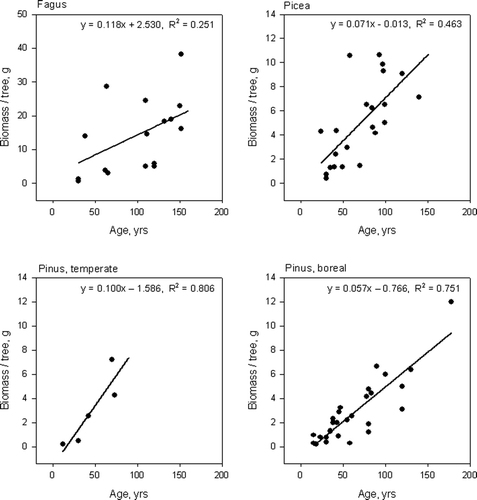
Figure 7. Relationship between stand basal area per tree and fine root biomass per tree for beech and spruce in the temperate zone and for Scots pine in the boreal zone.
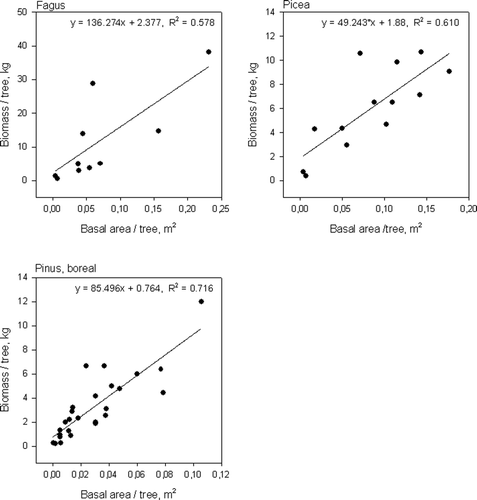
Table V. The Pearson correlation coefficients between biomass per tree (g) and the stand age and basal area per tree.a
Discussion
The reviewed papers showed that the average fine root biomasses for the temperate zone species were within the range reported by Noguchi et al. (Citation2007) for other temperate forests (49 – 749 g m−2). The fraction of roots <5 mm in diameter was larger than that of <2 mm roots. However, the biomasses reported by Vogt et al. (Citation1996) for <5 mm diameter roots in boreal forests (60 – 165 g m−2) were smaller than those reviewed in this study. The root collection methods could differ between studies.
We could reach our aim to find out the differences in fine root biomass between the temperate vs. boreal zones only partly, because for the deciduous trees distributed both in the boreal and the temperate zone the fine root data were too scarce. Silver birch (Betula pendula Roth.) is economically the most important deciduous tree species in the boreal zone, and it is also widely distributed in the temperate zone of Europe (Evans, Citation1984; Krüssmann, Citation1976). Fine root biomass (<2 mm in diameter) varied from 154 to 538 g m−2 in three young and middle-aged silver birch stands (Hobbie et al., Ecosystems, 2006, submitted) (Mamayev, Citation1977; Uri et al., Citation2007).
Fine root biomass had a large variation between sites. Despite of the scatter caused by this variation, there were trends supporting or rejecting our hypotheses on the relationships between fine root biomass, and climatic, tree stand and soil characteristics.
The results supported only partly the first hypothesis that the fine root biomass is higher in the temperate than in the boreal zone. However, this was true for pine but not for spruce. It is unlikely that these differences are related to variation in the sampling depth, which was less than 40 cm mineral soil in most of the studies from the boreal Scots pine forests but up to 60 – 100 cm mineral soil in most of the temperate Scots pine forests. Jackson et al. (Citation1996) reported in their review that the rooting depth of the boreal forests is shallower than that of temperate forests. In boreal forests 80 – 90% of roots are in the top 30 cm, or even top 20 cm, since Helmisaari et al. (Citation2007) reported that only 7.5% of Scots pine and Norway spruce fine roots were in the mineral soil layer 20 – 30 cm deep, while in temperate forests 78% of roots are found in the top 50 cm of soil (Jackson et al., Citation1996). Thus, the underestimate in boreal Scots pine fine root biomass caused by not sampling deeper than 30 or 40 cm in the mineral soil is relatively small, and the minor error related to different sampling depths is not likely to have caused the observed difference between the zones. Spruce sampling depths did not systematically differ between the zones.
The differing age of the Scots pine stands between zones may have been another source of error, as Scots pine fine root biomass correlated positively with stand age in the temperate zone. In the data on temperate Scots pine forests only 14% of the stands were younger than 40 years old whereas the share of young stands was over two times higher (32%) in the data from the boreal region. The greatest difference between Scots pine stands from temperate and boreal zones was, however, stand density. Temperate Scots pine stands were much denser than boreal stands. When the fine root biomass was calculated on the basis of stocking, there were no differences between zones in the mean fine root biomass per pine tree.
Contrary to our results, Jackson et al. (Citation1996) concluded that fine root biomass was generally higher in temperate deciduous and coniferous forests than in the boreal forests. Their data were, however, based only on a few data points from these regions, and European forests were poorly represented. Based on data presented here from European forests we cannot generally conclude that fine root biomass is higher in the temperate zone. This is in agreement with the findings of Vogt et al. (Citation1996).
Some of the observed significant relationships were based on clustered data. For instance, the significant correlation between beech and pine fine root biomass and latitude in the temperate zone was based on clustered data. For beech the cluster consisting of a few southernmost stands (latitude < 46°N) had low beech fine root biomasses which may be partly related to the low sampling depth (down to 20 cm mineral soil). Thus, especially for deeper-rooted beech (Bolte et al., Citation2004; Claus & George, Citation2005), the differences in the sampling depth may have been an additional source of error. Most of the beech data was from a narrow latitude range without any latitudinal trend in the beech fine root biomass. Among the temperate pine stands the variation in latitude was only 1° and the data could not be used for analyzing the relationship between the fine root biomass and latitude.
The results supported the second hypothesis that the fine root biomass of deciduous trees is higher than that of conifers. Vogt et al. (Citation1996), Leuschner & Hertel (Citation2003), and Noguchi et al. (Citation2007) reported similar results, whereas those of Jackson et al. (Citation1996) are contradictory to ours. In our collected data beech stands were less dense than spruce or pine stands but beech trees were taller, and growing on fertile site types. Especially the fine root biomass per tree was several times larger for beech than for conifers in the temperate zone. The high fine root necromass of beech indicates that beech fine roots may also have a high turnover rate. This conclusion is supported by fine root life span estimates using minirhizotron images of individual roots in a 30-year-old common-garden experiment with Fagus sylvatica, Pinus sylvestris, and Picea abies in central Poland (Withington et al., Citation2006). In that study median life span of two smallest root orders (with max. diameter 0.3 – 0.4 mm) of beech was 0.57 years, and was by 15% shorter than that of pine (0.67 years), and by 19% shorter than that of spruce (0.70 years).
The correlations between site fertility characteristics and fine root biomass seemed to be species specific. There was a higher fine root biomass of beech on the poorest sites, and of spruce on medium fertility sites. Also, the positive correlation between the organic layer C/N ratio and pine fine root biomass indicated that the low release of mineral nitrogen increased fine root biomass, thus partly supporting the third hypothesis. The data which reported C/N ratios were solely from Finland. Eight of the pine sites were from the study by Helmisaari et al. (Citation2007), who reported a stronger relationship between the fine root biomass of both Norway spruce and Scots pine and the C/N ratio in Finland, where nitrogen is the main growth-limiting factor on upland soils. Only a few papers reported soil C/N ratios from the temperate region. It may also be that this ratio is ecologically less important in the European temperate region where nitrogen deposition is high, and in many areas nitrogen is no more a growth-limiting factor.
The data did not support the fourth hypothesis – there was no correlation between fine root biomass and soil acidity. Godbold et al. (Citation2003) studied the effect of soil acidity on Norway spruce fine root density in the organic layer, and reported that on the most acidic sites fine root density was higher than on the least acidic sites. As in the beech stands studied by Leuschner et al. (Citation2004) no change in fine root biomass was found with increasing soil acidity.
Precipitation correlated positively only with spruce fine root biomass in the boreal zone. The data was highly scattered and mostly from areas with relatively low annual rainfall (460 – 709 mm year−1), where there was no relationship between fine root biomass and the amount of annual precipitation. The data points (2) with high rainfall (>1100 mm year−1) in boreal forests were from maritime, western Sweden from young stands (24 and 31 years old), which may have partly contributed to data scatter. We did not find significant correlations between fine root biomass and precipitation in the temperate zone, whereas this relationship was reported in several earlier papers. Leuschner & Hertel (Citation2003) found significant correlations between fine root biomass and precipitation for spruce and beech also in the temperate forests. According to Leuschner et al. (Citation2004) beech can grow in regions with both high and low rainfall (500 to 2000 mm year−1), even if it is a drought-sensitive species (Backes & Leuschner, Citation2000). However, not only the total annual rainfall but its seasonal distribution is important for soil water availability and its effects on fine roots. Along a precipitation gradient in Germany, Leuschner et al. (Citation2004) concluded that beech fine root biomass was highly sensitive to summer droughts resulting in a reduction at sites with low precipitation amounts.
The significant correlation between the fine root biomass of beech and pine with stand age in the temperate zone only partly supported the fifth hypothesis that fine root biomass increases by canopy closure, and thereafter it does not increase or decrease. Many studies have reported that fine root biomass peaks at canopy closure (Helmisaari et al., Citation2002; Claus & George, Citation2005), but the stand level data gathered for this study largely varied in this respect. The tree level correlations were better in agreement: fine root biomass increased with stand age, except for spruce in the boreal zone. For beech there was an increasing trend on tree level, but a decreasing trend on stand level. There were less data on the tree level as stand densities were reported in only half of the reviewed beech stands. Many of the highest stand level beech fine root biomasses (>600 g m−2) were in stands younger than 40 years old. Our stand level results confirm the results by Claus & George (Citation2005) on beech stands having the highest fine root biomass at younger age. On the contrary, temperate Scots pine forests had clearly lowest fine root biomasses at young age. Scots pine is a light-demanding early-successional species, and reaches canopy closure later than late-successional beech.
The tree level correlations between fine root biomass and tree age and basal area were better than respective correlations on stand level. Only the correlation between spruce fine root biomass and stand basal area in the boreal zone was better on stand level than on tree level. These higher correlations on tree level compared to stand level supported the sixth hypothesis. When tree level characteristics are observed, stand density variation can be omitted. This approach has earlier been used by, for example, Chen et al. (Citation2004) for boreal and temperate forests, and Helmisaari et al. (Citation2007) for boreal Scots pine and Norway spruce forests. We agree with Chen et al. (Citation2004) who concluded that above-ground variables such as tree size distribution should be included to improve usefulness of fine root biomass data for large-scale studies. Also, fine root biomass studies reporting needle biomass would be ecologically valuable as needle: fine root biomass ratio is reported to vary according to, for example, site fertility and stand characteristics and population origin (Oleksyn et al., Citation1999; Helmisaari et al., Citation2007).
Conclusions
The average fine root biomass of common European tree species falls within the same range as that in the corresponding vegetation zone in the other parts of the world. Many sources of errors and methodological issues are involved in the estimation of fine root biomass at stand level, and the variation observed in this study between sites may partly originate from these factors. However, here, as in some earlier reviews significant relationships between fine root biomass, and climatic, site and stand characteristics could be found. Our results show clearly that at tree level there exists a strong relationship between the fine root biomass and the above-ground biomass. There is a need to standardize the fine root biomass estimation methods and to collect more information on the relationships between fine root biomass and easily measurable or available climatic, site and stand factors to be better able to include the fine roots in ecosystem carbon pool and flux analyses in Europe and elsewhere.
Acknowledgements
Financial support for this work by European COST Action E38 ‘Woody Root processes’ is gratefully acknowledged. The authors thank Dr. Ain Kull for help in designing the map of the location of stands and Dr. Ivan Janssens for providing data.
Related Research Data
References
- Al Sayegh-Petkovšek , S . 2000 . “ Types of ectomycorrhizae from the forest research plots in Kočevska Reka and Zavodnje ” . In The rhizosphere: Studies of forest soils and the rhizosphere and their influences on chosen physiological parameters of forest trees in selected forest ecosystems, forest types and developmental phases of the forest. Studia Forestalica Slovenica (Strokovna in znanstvena dela) 118 , Edited by: Kraigher , H and Smolej , I . 119 – 153 . Ljubljana : Slovenian Forestry Institute .
- Ammer , C and Wagner , S . 2005 . An approach for modelling the mean fine-root biomass of Norway spruce stands . Trees , 19 : 145 – 153 .
- Backes , K and Leuschner , C . 2000 . Leaf water relations of competitive Fagus sylvatica and Quercus petraea trees during four years differing in soil drought . Can J Forest Res , 30 : 335 – 346 .
- Berg , B . 1984 . Decomposition of root litter and some factors regulating the process: Long-term root litter decomposition in a Scots pine forest . Soil Biol Biochem , 16 : 609 – 617 .
- Bolte , A , Rahmann , T , Kuhr , M , Pogoda , P , Murach , D and Gadow , K . 2004 . Relationships between tree dimension and coarse root biomass in mixed stands of European beech (Fagus sylvatica L.) and Norway spruce (Picea abies (L.) Karst.) . Plant Soil , 264 : 1 – 11 .
- Bolte , A and Villanueva , I . 2006 . Interspecific competition impacts on the morphology and distribution of fine roots in European beech (Fagus sylvatica L.) and Norway spruce (Picea abies (L.) Karst.) . Eur J Forest Res , 125 : 15 – 26 .
- Borken , W , Kossmann , G and Matzner , E . 2007 . Biomass, morphology and nutrient contents of fine roots in four Norway spruce stands . Plant Soil , 292 : 79 – 93 .
- Brunner , I , Brodbeck , S and Walthert , L . 2002 . Fine root chemistry, starch concentration, and ‘vitality’ of subalpine conifer forests in relation to soil pH . Forest Ecol Manag , 165 : 75 – 84 .
- Cairns , M A , Brown , S , Helmer , E H and Baumgardner , G A . 1997 . Root biomass allocation in the world's upland forests . Oecologia , 111 : 1 – 11 .
- Chen , W J , Zhang , Q F , Cihlar , J , Bauhus , J and Price , D T . 2004 . Estimating fine-root biomass and production of boreal and cool temperate forests using above-ground measurements: A new approach . Plant Soil , 265 : 31 – 46 .
- Claus , A and George , E . 2005 . Effect of stand age on fine-root biomass and biomass distribution in three European forest chronosequences . Can J Forest Res , 35 : 1617 – 1625 .
- Eldhuset , T D , Lange , H and de Wit , H A . 2006 . Fine root biomass, necromass and chemistry during seven years of elevated aluminium concentrations in the soil solution of a middle-aged Picea abies stand . Sci Total Environ , 369 : 344 – 356 .
- Epron , D , Farque , L , Lucot , E and Badot , P M . 1999 . Soil CO2-efflux in a beech forest: The contribution of root respiration . Ann Forest Sci , 56 : 289 – 295 .
- Evans , J . 1984 . Silviculture of broadleaved woodland . Forest Comm Great Br Bull , 62 : 187 – 191 .
- Finér , L and Laine , J . 1998 . Root dynamics at drained peatland sites of different fertility in southern Finland . Plant Soil , 201 : 27 – 36 .
- Finér , L , Mannerkoski , H , Piirainen , S and Starr , M . 2003 . Carbon and nitrogen pools in an old-growth, Norway spruce mixed forest in eastern Finland and changes associated with clear-cutting . Forest Ecol Manag , 174 : 51 – 63 .
- Fogel , R . 1983 . Root turnover and productivity of coniferous forests . Plant Soil , 71 : 75 – 85 .
- Genenger , M , Zimmermann , S , Hallenbarter , D , Landolt , W , Frossard , E and Brunner , I . 2003 . Fine root growth and element concentrations of Norway spruce as affected by wood ash and liquid fertilisation . Plant Soil , 255 : 253 – 264 .
- Godbold , D L , Fritz , H-W , Jentschke , G , Meesenburg , H and Rademacher , P . 2003 . Root turnover and root necromass accumulation of Norway spuce (Picea abies) are affected by soil acidity . Tree Physiol , 23 : 915 – 921 .
- Grebenc , T . 2005 . Types of ectomycorrhizae on beech (Fagus sylvatica L.) in natural and managed forest , Ljubljana : University of Ljubljana . Doctoral dissertation Available at: http://www.digitalna-knjiznica.bf.uni-lj.si/dd_grebenc_tine.pdf. Accessed 30 Mar 2007
- Grebenc , T and Kraigher , H . 2007a . Types of ectomycorrhiza of mature beech and spruce at ozone-fumigated and control forest plots Available at: http://dx.doi.org/10.1007/s10661-006-9414-3. Accessed 30 Mar 2007
- Grebenc , T and Kraigher , H . 2007b . Changes in community of ectomycorrhizal fungi and increased root turnover under adult beech trees chronically fumigated with double-ambient ozone concentration . Plant Biol , 9 : 279 – 287 .
- Grebenc , T , Štupar , B and Kraigher , H . 2007 . “ Pomen korenin in mikorize za ponor ogljika (The role of roots and mycorrhizae in carbon sequestration) ” . In Podnebne spremembe: vpliv na gozd in gozdarstvo (Climate change: impact on forest and forestry). Studia forestalia Slovenica 130 , Edited by: Jurac , M . 399 – 413 . Ljubljana : Biotechnical Faculty, Department of Forestry and Renewable Forest Resources Slovenia. (in Slovenian) .
- Helmisaari , H S , Derome , J , Nöjd , P and Kukkola , M . 2007 . Fine root biomass in relation to site and stand characteristics in Norway spruce and Scots pine stands . Tree Physiol , 27 : 1493 – 1504 .
- Helmisaari , H S and Hallbäcken , L . 1999 . Fine-root biomass and necromass in limed and fertilized Norway spruce (Picea abies (L.) Karst.) stands . Forest Ecol Manag , 119 : 99 – 110 .
- Helmisaari , H S , Makkonen , K , Kellomäki , S , Valtonen , E and Mälkönen , E . 2002 . Below- and above-ground biomass, production and nitrogen use in Scots pine stand in eastern Finland . Forest Ecol Manag , 165 : 317 – 326 .
- Hendrick , R L and Pregitzer , K S . 1993 . The dynamics of fine root length, biomass, and nitrogen content in two northern hardwood ecosystems . Can J Forest Res , 23 : 2507 – 2520 .
- Hendriks , C MA and Bianchi , F JJA . 1995 . Root density and root biomass in pure and mixed stands of Douglas-fir and Beech . Neth J Agr Sci , 43 : 321 – 331 .
- Jackson , R B , Canadell , J , Ehleringer , J R , Mooney , H A , Sala , O E and Schulze , E D . 1996 . A global analysis of root distributions for terrestrial biomes . Oecologia , 108 : 389 – 411 .
- Janssens , I , Sampson , D A , Curiel-Yuste , J , Carrara , A and Ceulemans , R . 2002 . The carbon cost of fine root turnover in a Scots pine forest . Forest Ecol Manag , 168 : 231 – 240 .
- Jentschke , G , Drexhage , M , Fritz , H W , Fritz , E , Schella , B Lee , D H . 2001 . Does soil acidity reduce subsoil rooting in Norway spruce (Picea abies)? . Plant Soil , 237 : 91 – 108 .
- Joslin , J D and Henderson , G S . 1987 . Organic matter and nutrients associated with fine root turnover in a white oak stand . Forest Sci , 33 : 330 – 346 .
- Keyes , M R and Grier , C C . 1981 . Above- and below-ground net production in 40-year-old Douglas-fir stands on low and high productivity sites . Can J Forest Res , 11 : 599 – 605 .
- Konŏpka , B and Pavlenda . 2004 . Liming and fertilization effects on rhizosphere and soil properties in mountain spruce stand . Ecologia/Bratislava , 23 : 14 – 23 .
- Konŏpka , B , Yuste , J C , Janssens , I A and Ceulemans , R . 2005 . Comparison of fine root dynamics in Scots pine and pendunculate oak in sandy soil . Plant Soil , 276 : 33 – 45 .
- Košir , Ž . 1976 . Zasnova uporabe prostora – Gozdarstvo [The basis for land use – Forestry] , 145 Ljubljana : IGLG . [in Slovenian]
- Kraigher , H . 1997 . Mycobioindication of pollution of two forest sites . Zbornik gozdarstva in lesarstva , 52 : 279 – 322 .
- Kraigher , H . 1999 . Diversity of types of ectomycorrhizae on Norway spruce in Slovenia . Phyton , 39 : 199 – 202 .
- Kraigher , H , Al Sayegh-Petkovšek , S , Grebenc , T and Simončič , P . 2007 . Types of ectomycorrhiza as pollution stress indicators: Case studies in Slovenia . Available at: http://dx.doi.org/10.1007/s10661-006-9413-4. Accessed 30 Mar 2007
- Kraigher , H , Batič , F and Agerer , R . 1996 . Types of ectomycorrhizae and mycobioindication of forest site pollution . Phyton , 36 : 115 – 120 .
- Krüssmann , G . 1976 . Handbuch der Laubgehöltze. 2. neugearbeitete und erweiterte Auflage in drei Bänden und einem Registerband. Band I A-D , 486 Berlin : Paul Parey Verlag . [in German]
- Kutnar , L , Zupančič , M , Robič , D , Zupančič , N , Žitnik , S Kralj , T . 2002 . The delimitation of the regions of provenance of forest tree species in Slovenia based on ecological regions . Zbornik Gozdarstva in Lesarstva , 67 : 73 – 117 .
- Leuschner , C and Hertel , D . 2003 . Fine root biomass of temperate forests in relation to soil acidity and fertility, climate, age and species . Prog Bot , 64 : 405 – 438 .
- Leuschner , C , Hertel , D , Schmid , I , Koch , O , Muhs , A and Hölscher , D . 2004 . Stand fine root biomass and fine root morphology in old-growth beech forests as a function of precipitation and soil fertility . Plant Soil , 258 : 43 – 56 .
- Lõhmus , K and Oja , T . 1983 . On research methods of belowground parts of forests . Soviet Forest Sci , 4 : 56 – 62 . [in Russian]
- Majdi , H . 2001 . Changes in fine root production and longevity in relation to water and nutrient availability in a Norway spruce stand in northern Sweden . Tree Physiol , 21 : 1057 – 1061 .
- Majdi , H and Persson , H . 1995 . Effect of ammonium sulphate application on the chemistry of bulk soil, rhizosphere, fine roots and fine-root distribution in a Picea abies (L.) Karst. stand . Plant Soil , 168 – 169 : 151 – 160 .
- Mälkönen , E . 1974 . Annual primary production and nutrient cycle in some Scots pine stands . Comm Inst For Fenn , 84 : 1 – 87 .
- Mamayev , V . 1977 . “ Root mass in the pine and the birch stand in Oxalis-myrtillus site ” . In Silvicultural research in Southern taiga , Edited by: Orlov , A . 61 – 66 . Moscow : Nauka Publishers . [in Russian]
- Mamayev , V . 1986 . Ecological characteristics of root systems of pine and birch in stands of high productivity in Southern taiga , 129 Academy of Sciences, USSR: Dnepropetrovsk . [in Russian]
- Möttönen , M , Lehto , T , Aphalo , P , Kukkola , M and Mälkönen , E . 2003 . Response of mature stands of Norway spruce (Picea abies) to boron fertilization . Forest Ecol Manag , 180 : 401 – 412 .
- Murach , D . 1984 . Die Reaktion der Feinwurzeln von Fichten (Picea abies Karst.) auf zunehmende Bodenversauerung , 126 Universität Göttingen, Bodenkundliche Berichte 77 . [in German]
- Noguchi , K , Konôpka , B , Satomura , T , Kaneko , S and Takahashi , M . 2007 . Biomass and production of fine roots in Japanese forests . J Forest Res , 12 : 83 – 95 .
- Ohashi , M , Kilpeläinen , J , Finér , L , Risch , A C , Domisch , T Neuvonen , S . 2007 . The effect of red wood ant (Formica rufa group) moundes on root biomass, density, and nutrient concentrations in boreal managed forests . J Forest Res , 12 : 113 – 119 .
- Oleksyn , J , Reich , P B , Chalupka , W and Tjoelker , M G . 1999 . Differential above- and below-ground biomass accumulation of European Pinus sylvestris populations in a 12-yr-old provenance experiment . Scand J Forest Res , 14 : 7 – 17 .
- Ostonen , I , Lõhmus , K and Pajuste , K . 2005 . Fine root biomass, production and its proportion of NPP in a fertile middle-aged Norway spruce forest: Comparison of soil core and ingrowth core methods . Forest Ecol Manag , 212 : 264 – 277 .
- Persson , H . 1979 . Fine-root production, mortality and decomposition in forest ecosystems . Plant Ecol , 41 : 101 – 109 .
- Persson , H . 1983 . The distribution and productivity of fine roots in boreal forests . Plant Soil , 71 : 87 – 101 .
- Püttsepp , Ü , Lõhmus , K , Persson , H A and Ahlstrom , K . 2006 . Fine-root distribution and morphology in an acidic Norway spruce (Picea abies (L.) Karst.) stand in SW Sweden in relation to granulated wood ash application . Forest Ecol Manag , 221 : 291 – 298 .
- Santantonio , D , Hermann , R K and Overton , W S . 1977 . Root biomass studies in forest ecosystems . Pedobiologia , 17 : 1 – 31 .
- Schmid , I . 2002 . The influence of soil type and interspecific competition on the fine root system of Norway spruce and European beech . Basic Appl Ecol , 3 : 339 – 346 .
- Uri , V , Lõhmus , K , Ostonen , I , Tullus , H , Lastik , R and Vildo , M . 2007 . Biomass production, foliar and root characteristics and nutrient accumulation in young silver birch (Betula pendula Roth.) stand growing on abandoned agricultural land . Eur J Forest Res , Available at: http://dx.doi.org/10.1007/s10342-007-0171-9
- Vanguelova , E I , Nortcliff , S , Moffat , A J and Kennedy , F . 2005 . Morphology, biomass and nutrient status of fine roots of Scots pine (Pinus sylvestris) as influenced by seasonal fluctuations in soil moisture and soil solution chemistry . Plant Soil , 270 : 233 – 247 .
- Vanninen , P and Mäkelä , A . 1999 . Fine root biomass of Scots pine stands differing in age and soil fertility in southern Finland . Tree Physiol , 19 : 823 – 830 .
- Vanninen , P H , Ylitalo , H , Sievänen , R and Mäkelä , A . 1996 . Effects of age and site quality on the distribution of biomass in Scots pine (Pinus sylvestris L.) . Trees , 10 : 231 – 238 .
- Vilhar , U and Kraigher , H . 1999 . Types of ectomycorrhizae on Pohorje . Phyton , 39 : 203 – 206 .
- Vogt , K A , Grier , C C and Vogt , D J . 1986 . Production, turnover, and nutrient dynamics of above- and belowground detritus of world forests . Adv Ecol Res , 15 : 303 – 378 .
- Vogt , K A , Moore , E E , Vogt , D J , Redlin , M J and Edmonds , R L . 1983 . Conifer fine root and mycorrhizal root biomass within the forest floors of Douglas-fir stands of different ages and site productivities . Can J Forest Res , 13 : 429 – 437 .
- Vogt , K A and Persson , H . 1991 . “ Measuring growth and development of roots ” . In Techniques and approaches in Forest Tree Ecophysiology , Edited by: Lassoie , J P and Hinckley , T M . 447 – 501 . Boca Raton, FL : CRC Press .
- Vogt , K A , Vogt , D J , Moore , E E , Fatuga , B A , Redlin , M R and Edmonds , R L . 1987 . Conifer and angiosperm fine-root biomass in relation to stand age and site productivity in Douglas-fir forests . J Ecol , 75 : 857 – 870 .
- Vogt , K A , Vogt , D J , Palmiotto , P A , Boon , B , O'Hara , J and Asbjornsen , H . 1996 . Review of root dynamics in forest ecosystems grouped by climate, climatic forest type and species . Plant Soil , 187 : 159 – 219 .
- Withington , J M , Reich , P B , Oleksyn , J and Eissenstat , D M . 2006 . Comparisons of structure and life span in roots and leaves among temperate trees . Ecol Monogr , 76 : 381 – 397 .
- Xiao , C W , Curiel Yuste , J , Janssens , I A , Roskams , P , Nachtergale , L Carrara , A . 2002 . Above- and belowground biomass and net primary production in a 73-yr-old Scots pine forest . Tree Physiol , 23 : 505 – 516 .