
Abstract
The Centaureo-Portenschlagiellion describes the cliff vegetation of the Southeastern Adriatic. This habitat type harbours many endemic taxa and has a high value for biodiversity conservation. Notwithstanding its importance, knowledge of the syntaxonomy of this alliance is still poor. This article aims at revising the synchorological, coenological and floristic relationships of the associations of the Centaureo-Portenschlagiellion. The revision is based on a data set of 103 relevés of Mediterranean xerothermic cliffs from Croatia, Bosnia i Herzegovina and Montenegro. The relevés were clustered by using the Flexible beta method. An indicator species analysis was used to identify the diagnostic taxa of the main clusters of relevés and non-metric multidimensional scaling ordination was undertaken to visualize the floristic relationships among them. Results revealed that only seven associations belong to the Centaureo-Portenschlagiellion, whereas the others already described in phytosociological literature were invalidly described or should be treated as syntaxonomic synonyms of the previous ones. Two associations (Inulo-Centaureetum cuspidatae and Portenschlagiello-Campanuletum portenschlagianae) were reduced to the rank of subassociations of the Moltkio-Inuletum verbascifoliae. Two other subassociations were described for the first time. Data provided with this revision may be considered as essential base-line information that should aid in evaluating the state of this vegetation type in the future.
Introduction
Limestone cliffs harbour a high number of endemic and rare plant and animal species. The European Council Directive 92/43/EEC, the so-called “Habitat Directive”, lists “Calcareous rocky slopes with chasmophytic vegetation” (habitat code: 8210) among the habitat types constituting the European protected area within the Natura 2000 network. Although the Habitat Directive refers to the member states of the European Union, many neighbouring countries are going to align their protected area systems with the Natura 2000 criteria (e.g. Petrović et al. Citation2012 for Montenegro; Milanović et al. Citation2015 for Bosnia and Herzegovina).
The Interpretation Manual of the Habitat Directive (European Commission Citation2013) provides a general description for each habitat type, including also syntaxonomical references. Thus, syntaxonomy is an important source of information for identifying habitats and coordinating conservation measures within and beween countries.
The syntaxonomy of xerothermic limestone chasmophytic vegetation of the Adriatic area was reviewed by Trinajstić (Citation1980) who described the order “Centaureo-Campanuletalia”, including two alliances, “Centaureo-Campanulion” and “Centaureo-Portenschlagiellion”. The latter alliance covers an elongated area in the central and southern part of the eastern Adriatic pertaining to Croatia, Bosnia and Herzegovina; Montenegro and Albania (Jovanović et al. Citation1986; Petrović et al. Citation2012; Terzi et al. Citation2018). Trinajstić (Citation1980, Citation2008) classified five associations in the Centaureo-Portenschlagiellion: Portenschlagiello ramosissimae-Campanuletum portenschlagianae, Campanulo pyramidalis-Moltkietum petraeae, Inulo verbascifoliae-Centaureetum cuspidatae, Centaureetum ragusinae and Seslerio robustae-Putorietum calabricae. The distribution areas of this alliance include some protected and narrowly endemic taxa, such as Centaurea cuspidata, C. ragusina, C. radichii, Fibigia triquetra and Campanula portenschlagiana.
Before this syntaxonomic scheme, Lakušić (Citation1968, Citation1970) had classified the limestone cliff vegetation of the eastern Adriatic in another order, Moltkietalia petraeae, consisting of two alliances, the Centaureo dalmaticae-Campanulion and the Edraianthion tenuifolii, including coastal and inland vegetation, respectively (see also Terzi and Di Pietro Citation2016). Thus, several other chasmophytic associations (Fibigio triquetrae-Cerinthetum tristis, Micromerio kerneri-Onosmetum dalmaticae, Moltkio petraeae-Campanuletum lepidae, Moltkio petraeae-Centaureetum voraginicolae nom. inval. (Art. 3 l of ICPN), Moltkio petraeae-Inuletum verbascifoliae, Puccinellio-Centaureetum crithmifoliae nom. inval. (Art. 2b) and Teucrio arduinii-Seselietum globiferi) have been described for xerothermic limestone cliffs occurring in an area potentially falling within the distribution range of the Centaureo-Portenschlagiellion, but they were classified in the Moltkietalia petraeae (Lovrić and Rac Citation1987; Lovrić and Bedalov Citation1987; Lovrić and Rac Citation1989, Lovrić et al. Citation2002).
The two orders, Centaureo-Campanuletalia and Moltkietalia petraeae, at least in their original descriptions, show some floristic, coenological and geographical overlaps, especially along the more thermophilous fringe of the Moltkietalia petraeae (see Terzi and Di Pietro Citation2016). The authors of the abovementioned associations followed one of these two syntaxonomic concepts, but did not compare or discuss their findings in the light of the two different possible interpretations, leaving unresolved the coenological overlaps between the syntaxonomic schemes proposed by Lakušić (Citation1968) and Trinajstić (1980).
More recently, Mucina et al. (Citation2016) solved this issue, differentiating the two orders and restricting the Centaureo-Campanuletalia to the thermo-mesomediterranean chasmophytic vegetation of limestone cliffs of the Adriatic coastal regions and leaving the Moltkietalia petraeae for the montane and alpine belts of the Central and Southern Dinarides.
Within this overall pattern, the coenological and floristic relationships among the associations of the Centaureo-Portenschlagiellion and those originally classified in the Moltkietalia petraeae but clearly belonging to the thermo-mesomediterranean belt, remain unclear, as well as the diagnostic species of each of them. For example, the steno-endemic Campanula portenschlagiana is considered to be a character species of the Moltkio petraeae-Inuletum verbascifoliae (Lovrić and Bedalov Citation1987), Campanulo-Moltkietum petraeae (Horvatić Citation1963) and also of the Portenschlagiello ramosissimae-Campanuletum portenschlagianae (Trinajstić Citation1980). Moreover, this species was also recorded with high frequency in the Inulo verbascifoliae-Centaureetum cuspidatae and in the Campanulo pyramidalis-Moltkietum petraeae in the Biokovo Mountains (compare Trinajstić Citation1987).
Notwithstanding the importance of this habitat type, knowledge about the syntaxonomic differentiation of the associations of the Centaureo-Portenschlagiellion is still poor.
The nomenclature of the two orders, Centaureo-Campanuletalia and Moltkietalia petraeae, was reviewed in some recent articles through the validation and/or typification of some syntaxon names (Terzi and Di Pietro Citation2016, Jasprica and Terzi Citation2017, Terzi et al. Citation2017). Moreover, in a syntaxonomic revision of the xerothermic chasmophytic vegetation of the central part of the Mediterranean Basin, Terzi et al. (Citation2018) defined the syntaxonomic position of the Centaureo-Portenschlagiellion compared with the other alliances occurring in the area. The aim of this article is to revise the synchorological, coenological and floristic features of the associations of the Centaureo-Portenschlagiellion.
Materials and methods
This revision is based on relevés already published in the phytosociological literature and classified in the syntaxa listed in Supplementary Appendix S1 (). Three relevés on Koločep Island, originally published by Horvatić (Citation1971), and assigned to the Seslerio robustae-Putorietum calabricae were excluded from the data set. In fact, the occurrences of Anthyllis barba-jovis and Helichrysum italicum indicate a transition towards the Anthyllidion barbae-jovis (sensu Mucina et al. Citation2016).
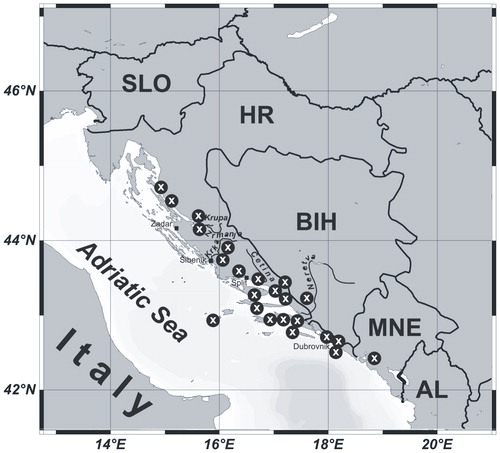
Figure 1. The study area with distribution of relevés. Abbreviations: SLO: Slovenia; HR: Croatia; BIH: Bosnia and Herzegovina; MNE: Montenegro; AL: Albania.
Two other associations of the Centaureo-Portenschlagiellion (“Puccinellio-Aurinietum leucadae” and “Aurinio-Brassicetum frutescentis”) from Vis Archipelago and Palagruža (Middle Adriatic, Croatia) were also excluded from this revision because they were invalidly and briefly described without any suppporting phytosociological table (Art. 2b, Lovrić and Bedalov Citation1987, Lovrić and Rac Citation1991).
A data matrix (relevés x species) was prepared replacing the original Braun-Blanquet scores with the ordinal value of the combined scale proposed by van der Maarel (Citation1979). Taxa indicated only at the genus level were omitted from the data set as well as mosses and lichens that were not always recorded.
The plot size is indicated for only 68% of the relevés. The average plot size of relevés was 43 m2 with minimum and maximum values of 1 m2 and 200 m2, respectively. Since excessively small or excessively large plot sizes may affect the outcomes of statistical analyses (Otýpková and Chytrý Citation2006), relevés recorded on plots outside the range 10 m2 to 100 m2 were removed. Eventually the data matrix consisted of 93 relevés and 188 taxa. To reduce noise in the dataset (compare McCune and Grace Citation2002), rare taxa occurring in only 1 or 2 relevés were removed before analyses.
The relevés were clustered by using the Flexible beta method, with the Bray-Curtis coefficient. The β value was set at –0.25 because with this value the Flexible beta is a space-conserving sorting strategy (McCune and Grace Citation2002). The resulting dendrogram was pruned at the level yielding the highest number of indicator species (IndSp, compare McCune and Grace Citation2002). Indicator species analysis (ISA, Dufrêne and Legendre Citation1997) was run for the first 20 partitioning levels of the dendrogram, further partitions dealing with minor variations. A taxon was considered as IndSp of a cluster if its indicator value (IndVal) turned out to be higher for that cluster than for the others of the same partitioning level, and if it turned out to be significant (p < .05) in a Monte Carlo test using 10,000 permutations.
IndSp were also used to characterize floristically the clusters of relevés. To increase interpretability of results, each IndSp was assigned to only one cluster along the descending hierarchical typology of the dendrogram. In accord with Dufrêne and Legendre (Citation1997), the best clustering level for a species was considered as the one for which the IndVal first reached its maximum value.
Results were interpreted from a syntaxonomic standpoint on the basis of the occurrences of the association type-relevés within the clusters and the set of IndSp associated to each cluster. Diagnostic taxa of the higher syntaxonomic ranks (alliance, order and class) were deduced from the scientific literature (compare Terzi et al. Citation2018).
A non-metric multidimensional scaling (NMDS) ordination was undertaken to visualize the floristic relationships among the main clusters of relevés. The NMDS was run in the “slow and thorough” autopilot mode of the PC-ORD (McCune and Mefford Citation2011), selecting the Bray-Curtis coefficient as dissimilarity measure.
All the statistical analyses described above were performed using the software PC-ORD, version 6.22 (McCune and Mefford Citation2011).
For each association, a brief description of its syntaxonomic and nomenclatural vicissitudes, the holotypus, locus classicus, distribution range, diagnostic, frequent and dominant taxa were reported. The locus classicus of a syntaxon is here defined as the locality from which the original diagnosis of that syntaxon (or at least its nomenclatural type) was described. To identify the “dominant taxa,” (i) first we selected those taxa occurring in at least one relevé of the phytosociological table with a score (on the ordinal scale) of 5 or more and then (ii) we calculated for each of these taxa the sum of the scores for all the relevés included in the phytosociological table. Those taxa yielding the highest values were considered as “dominant taxa.” The diagnostic taxa were subjectively selected from the IndSp list (Supplementary Appendix S2).
Results
The dendrogram was pruned at the 10th partitioning level that being the partition yielding the highest number of IndSp. The occurrence of the type-relevés of the associations in the clusters allowed us to assign each of the 10 main clusters of the dendrogram () to one or more association.
Figure 2. Clustering of relevés. Partitioning levels are indicated by numbers; the two clusters originated at each partition are labelled as “a” and “b”. The main clusters represent the following associations: CaM: Campanulo austroadriaticae-Moltkietum petraeae; MkO: Micromerio kerneri-Onosmetum dalmaticae and Moltkio petraeae-Campanuletum lepidae; TaS: Teucrio arduinii-Seselietum globiferi; CpM1/2: Campanulo pyramidalis-Moltkietum petraeae; IvC: Inulo verbascifoliae-Centaureetum cuspidatae; SrP: Seslerio robustae-Putorietum calabricae; Crl1: Centaureetum ragusinae limonietosum anfracti; Crt: Centaureetum ragusinae typicum; Crl2: Centaureetum ragusinae limonietosum cancellati; FtC: Fibigio triquetrae-Cerinthetum tristis; MpI: Moltkio petraeae-Inuletum verbascifoliae.
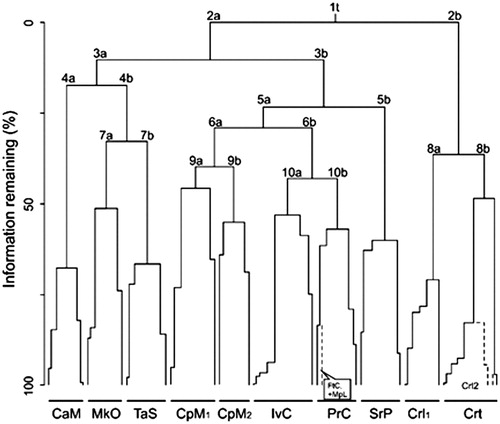
The first dendrogram division separates the relevés of the Centaureetum ragusinae (: cluster 2b) which were further arranged into two clusters. The first cluster (Crl1) represents the Centaureetum ragusinae subass. limonietosum anfracti. The second one (Crt) includes the relevés of the Centaureetum ragusinae typicum and another sub-group, namely Crl2, which includes relevés from Biševo Island (HR) representing a third coastal subassociation, C.r. subass. limonietosum cancellati. Several IndSp are associated with these clusters, some of them having high IndVal, such as Centaurea ragusina, Crithmum maritimum, Lotus cytisoides, Limonium dictyophorum and L. cancellatum. Most of these taxa are ingressive from the Crithmo-Staticetea class and indicate that most of the sampled sites were salt-sprayed coastal cliffs.
Four other clusters of relevés were characterized by many IndSp, with high IndVal, and they are floristically well distinguishable from the rest of the relevés (Supplementary Appendix S2). Three of them (cluster 4a, 4 b and 5b) represent the following associations: Campanulo austroadriaticae-Moltkietum petraeae (CaM), Seslerio robustae-Putorietum calabricae (SrP) and Teucrio arduinii-Seselietum globiferi (TaS). The fourth one (MkO) includes the type relevés of two associations, namely, Micromerio kerneri-Onosmetum dalmaticae and Moltkio petraeae-Campanuletum lepidae, together with another relevé of the Moltkio petraeae-Centaureetum voraginicolae. The latter association was invalidly described since one of the name-giving taxa (Centaurea voraginicola) had not been validly published before (or simultaneously with) the description of the new association (Art. 3l, Weber et al. Citation2000). These three associations are represented in the data set by very few relevés so that a definitive interpretation of their floristic composition cannot be expressed. Despite being grouped in the same cluster, some phytosociologically important species distinguish the Micromerio kerneri-Onosmetum dalmaticae (e.g. Micromeria kerneri, Seseli tomentosum) from the Moltkio petraeae-Campanuletum lepidae (e.g. Campanula fenestrellata subsp. fenestrellata, Arenaria orbicularis, Iris adriatica). Therefore, these associations are provisionally maintained as independent associations and further phytosociological data are needed to clarify their syntaxonomic relationship.
The cluster 5a is differentiated by only one IndSp (Portenschlagiella ramosissima) and its further subdivisions highlight two main clusters representing the Campanulo pyramidalis-Moltkietum petraeae from one side (cluster 6a, CpM1-2) and four associations (cluster 6b, IvC and PrC) scarcely differentiated on the other: Inulo verbascifoliae-Centaureetum cuspidatae, Portenschlagiello ramosissimae-Campanuletum portenschlagianae, Fibigio triquetrae-Cerinthetum tristis and Moltkio petraeae-Inuletum verbascifoliae. The last two associations are represented in the data-set by only one relevé, i.e. their holotypes.
The NMDS ordination resulted in a two axis solution with a final stress of 19.4, a fairly high value (). The ordination explained only 64.5% of the total proportion of variance, axes 1 and 2 accounting for 52.0% and 12.5% of variance, respectively.
Figure 3. NMDS ordination of relevés. Abbreviations are the same used in the dendrogram ().
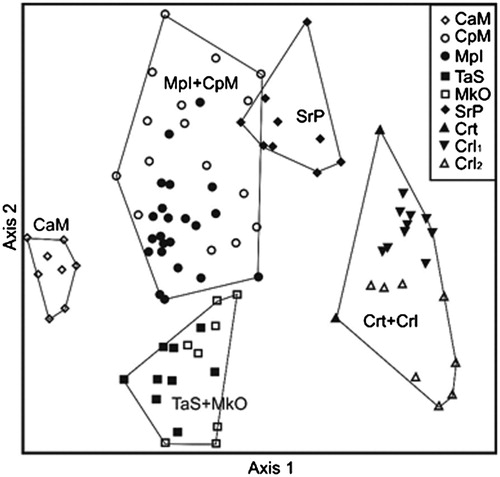
Axis 1 roughly represents an altitudinal gradient, with the Centaureetum ragusinae and the Seslerio robustae-Putorietum calabricae developing near the coastline at one end, and the Campanulo austroadriaticae-Moltkietum petraeae, of the Lovćen Mt. in Montenegro, at the other end. The other associations lie in the middle part of the diagram with those recorded along the eastern Adriatic canyons by Lovrić and Rac (Citation1987, Citation1989) and Lovrić et al. (Citation2002) in the lower part of the diagram. The cluster 6a, representing the Campanulo pyramidalis-Moltkietum petraeae, shows some overlaps with the cluster 6b, that is, Inulo verbascifoliae-Centaureetum cuspidatae (IvC) and Portenschlagiello ramosissimae-Campanuletum portenschlagianae (PrC). The results of the NMDS ordination confirm the general relationships among the associations as they are revealed by the cluster analysis. Online Supplementary Table SI shows the percentage frequencies of the diagnostic species for their corresponding vegetation units.
Discussion
Based on the results, the following syntaxonomic scheme is here proposed:
Class Asplenietea trichomanis (Braun-Blanquet in Meier et Braun-Blanquet 1934) Oberdorfer 1977
Order Centaureo dalmaticae-Campanuletalia pyramidalis Trinajstić ex Terzi et Di Pietro 2016
Alliance Centaureo cuspidatae-Portenschlagiellion ramosissimae Trinajstić ex Terzi et Di Pietro 2016
– Campanulo pyramidalis-Moltkietum petraeae Horvatić ex Trinajstić Citation1964 | |||||
– Centaureetum ragusinae Horvat ex Terzi, Jasprica et Caković 2017 | |||||
C.r. typicum Terzi, Jasprica et Caković 2017
C.r. limonietosum anfracti Hećimović ex Terzi, Jasprica et Caković 2017
C.r. limonietosum cancellati subass. nov. hoc loco
– Micromerio kerneri-Onosmetum dalmaticae Lovrić ex Terzi, Jasprica et Caković 2017 [syn. Moltkio petraeae-Centaureetum voraginicolae Lovrić nom. inval. (3l)] | |||||
– Moltkio petraeae-Campanuletum lepidae Lovrić ex Terzi, Jasprica et Caković 2017 | |||||
– Moltkio petraeae-Inuletum verbascifoliae Wendelberger in Lovrić et Rac 1987 [syn. Fibigio triquetrae-Cerinthetum tristis Lovrić in Lovrić et Rac 1987, Inulo verbascifoliae-Centaureetum cuspidatae Trinajstić ex Terzi et Di Pietro 2016, Portenschlagiello ramosissimae-Campanuletum portenschlagianae Jasprica et Terzi Citation2017] | |||||
M.p.-In.v. typicum subass. nov. hoc loco [holotypus rel. J1, tab. 6, p. 126, Lovrić and Rac (Citation1987)]
M.p.-In.v. centaureetosum cuspidatae (Terzi et Di Pietro 2016) stat. nov. hoc loco [=Inulo verbascifoliae-Centaureetum cuspidatae Trinajstić ex Terzi et Di Pietro 2016: 1367]
M.p.-In.v. fibigietosum triquetrae (Jasprica et Terzi Citation2017) stat. nov. hoc loco [=Portenschlagiello ramosissimae-Campanuletum portenschlagianae Trinajstić ex Jasprica et Terzi Citation2017: 332]
– Seslerio robustae-Putorietum calabricae Horvatić ex Birač Citation1973 | |||||
– Teucrio arduinii-Seselietum globiferi Lovrić, Rac et Šolić ex Terzi, Jasprica et Caković 2017 | |||||
Order Moltkietalia petraeae Lakušić Citation1968
Alliance Edraianthion tenuifolii Lakušić Citation1968
– Campanulo austroadriaticae-Moltkietum petraeae (Tomić-Stanković ex Terzi, Jasprica et Caković 2017) Terzi, Jasprica, Caković et Di Pietro 2018 | |||||
Campanulo pyramidalis-Moltkietum petraeae
This association was invalidly described by Horvatić (Citation1963) for the Mediterranean mountain belt (see also Trinajstić Citation1998) of Central and Southern Dalmatia, Croatia. It was validated by Trinajstić (Citation1964) with an original diagnosis consisting of only one relevé recorded on Korčula Island at nearly 300 m a.s.l. in the eu-Mediterranean belt. Our results showed that the Campanulo pyramidalis-Moltkietum petraeae is widely distributed, from the Southern Dalmatian islands, such as Korčula and Mljet, where the association turned out to be distinguished by some xerothermic species (i.e. Erica manipuliflora, Brachypodium retusum), up to Mt. Biokovo where a Mediterranean mountain variant is identified. No character taxa were found for this association which is distinguished only by three transgressive taxa from alliance and order (Campanula pyramidalis, Moltkia petraea and Portenschlagiella ramosissima).
Holotypus: relevé p. 158, in Trinajstić (Citation1964); Locus classicus: Korčula Island; Distribution area: Central and Southern Dalmatia; Differential taxa of the eu-mediterranean variant: Erica manipuliflora, Brachypodium retusum; Frequent and dominant taxa: Inula verbascifolia, Moltkia petraea, Brachypodium retusum, Campanula pyramidalis and Asplenium ceterach subsp. ceterach (the latter is frequent but not dominant).
Centaureetum ragusinae
This association was invalidly described by Horvat (Citation1942: 464) and subsequently renamed as Phagnalo-Centaureetum ragusinae by Horvatić (Citation1963). Hećimović (Citation1984) described the halophytic subassociations (C.r. limonietosum anfracti) distinguished by the steno-endemic Limonium dictyophorum. Our results show a floristic differentiation between the relevés from this subassociation and those carried out on Biševo Island and containing Limonium cancellatum (cluster Crl2). These two groups of relevés represent the halophytic aspects of the Centaureetum ragusinae association, with some ingressive taxa from the Crithmo-Staticetea class. The relevés from Biševo Island are here included in the new subassociation C.r. limonietosum cancellati (holotypus, rel. 7, tab. 5 in Pavletić Citation1973: Centaurea ragusina 3, Crithmum maritimum 1, Inula verbascifolia 1, Limonium cancellatum +, Capparis orientalis +, Phagnalon graecum +, Juniperus phoenicea +, Allium ampeloprasum +, Reichardia picroides +, Convolvulus cneorum +, Daucus carota subsp. hispanicus +).
The Centaureetum ragusinae is usually classified within the Centaureo cuspidatae-Portenschlagiellion (e.g. Trinajstić Citation1980, Citation2008). However, Terzi et al. (Citation2018) showed floristic similarities with the associations of the Centaureo-Campanulion alliance, in the Northern Adriatic. These similarities are due to the occurrence of the halophytic taxa recorded in several other cliff associations of the Centaureo-Campanulion. On the other hand, the Centaureetum ragusinae lacks almost completely the character species of both of the abovementioned alliances. Therefore, we provisionally classify the Centaureetum ragusinae within the Centaureo-Portenschlagiellion for geographical reasons, the distribution area of the main character taxon of the association, Centaurea ragusina, extending over the central and southern parts of Dalmatia.
Holotypus: rel. 2, tab. of “Phagnalo-Centaureetum ragusinae,” p. 258, in Trinajstić (Citation1995); Locus classicus: cliffs in the northwestern part of Mljet Island, Southern Dalmatia; Distribution area: Dalmatia; Diagnostic taxa: Centaurea ragusina, Convolvulus cneorum, Phagnalon rupestre subsp. graecum (locally). Frequent taxa: Centaurea ragusina, Inula verbascifolia, Crithmum maritimum, Allium ampeloprasum, Silene vulgaris, Convolvulus cneorum; Dominant taxon: Centaurea ragusina.
Micromerio kerneri-Onosmetum dalmaticae
The Micromerio kerneri-Onosmetum dalmaticae was invalidly described by Lovrić and Rac (Citation1987) for the canyons of Zrmanja and Čikola (Northern Dalmatia) and classified in the Edraianthion tenuifolii and Moltkietalia petraeae. This association has been recently moved to the Centaureo-Portenschlagiellion (Terzi et al. Citation2018).
The relevés of the Micromerio kerneri-Onosmetum dalmaticae were grouped in the same dendrogram cluster together with those of the Moltkio petraeae-Campanuletum lepidae (). Despite the floristic similarities between these two associations, they cannot be reunited in only one association because their character taxa are mutually exclusive. However, their syntaxonomic relationships need to be reassessed when more relevés are available.
On the other hand, the Moltkio petraeae-Centaureetum voraginicolae, invalidly described by Lovrić (in Lovrić and Rac Citation1987), is here considered as a synonym of the Micromerio kerneri-Onosmetum dalmaticae because of the high frequency of two character taxa, Micromeria kerneri and Asplenium ceterach subsp. bivalens (tab. 1).
Holotypus: rel. 5, tab. 2, p. 113, in Lovrić and Rac (Citation1989); Locus classicus: Krupa canyon, Northern Dalmatia; Distribution area: Northeastern Adriatic, Northern Dalmatia; Diagnostic taxa: Onosma echioides subsp. dalmatica (d), Micromeria kerneri, Seseli tomentosum, Asplenium ceterach subsp. bivalens; Frequent taxa: Onosma echioides subsp. dalmatica, Inula verbascifolia, Seseli tomentosum, Galium corrudifolium, Iris illyrica, Sesleria tenuifolia subsp. tenuifolia, Sedum hispanicum, Micromeria kerneri, Asplenium trichomanes, As. ceterach subsp. bivalens, Campanula pyramidalis, Sedum dasyphyllum; Dominant taxa: Onosma echioides subsp. dalmatica, Inula verbascifolia, Seseli tomentosum.
Moltkio petraeae-Campanuletum lepidae
This association is here classified in the Centaureo-Portenschlagiellion due to the presence of Moltkia petraea, Portenschlagiella ramosissima and Tanacetum cinerariifolium. However, lying in the northern part of the alliance distribution area, the Moltkio petraeae-Campanuletum lepidae includes also a character taxon of the Centaureo-Campanulion (Campanula fenestrellata subsp. fenestrellata) among its most frequent taxa.
Holotypus: rel. 2, tab. 2, p. 113, in Lovrić and Rac (Citation1989); Locus classicus: Krupa Canyon, Northern Dalmatia; Distribution area: Northern Dalmatian inland; Diagnostic taxa: Campanula fenestrellata subsp. fenestrellata, Arenaria orbicularis, Iris adriatica (d); Frequent taxa: Campanula fenestrellata subsp. fenestrellata, C. pyramidalis, Moltkia petraea, Galium corrudifolium, Asperula aristata, Asplenium trichomanes; Dominant taxa: Campanula fenestrellata subsp. fenestrellata, Ficus carica.
Moltkio petraeae-Inuletum verbascifoliae
The “Inula candida-Moltkea petraea-Ass. (Horv. 41) Wendelb 63” was invalidly described by Wendelberger (Citation1963) for the Biokovo mountain ridge, and classified in the Micromerion croaticae (Wendelberger Citation1963, Citation1965). This association was accidentally validated by Lovrić and Rac (Citation1987) through the publication of only one relevé, and classified in the Edraianthion tenuifolii. Subsequently, Lovrić et al. (Citation2002) moved this association to the Centaureo-Campanulion.
According to Lovrić and Rac (Citation1987), the Moltkio petraeae-Inuletum verbascifoliae is characterized by Campanula portenschlagiana and corresponds pro parte to the Campanulo-Moltkietum petraeae sensu Horvatić Citation1963, replacing at higher altitude the Fibigio triquetrae-Cerinthetum tristis along the Mt. Biokovo coastal cliffs. However, they did not mention or discuss the syntaxonomic proposals of Trinajstić (Citation1980, Citation1987) who described two other associations with Campanula portenschlagiana, namely Portenschlagiello ramosissimae-Campanuletum portenschlagianae and Inulo verbascifoliae-Centaureetum cuspidatae, characterized by Fibigia triquetra and Centaurea cuspidata, respectively. In the cluster analysis, the four associations mentioned above turned out to be grouped in the same cluster. Moreover, this cluster showed some floristic overlaps with the Campanulo pyramidalis-Moltkietum petraeae.
This result is biased by the low number of relevés for the Moltkio petraeae-Inuletum verbascifoliae and Fibigio triquetrae-Cerinthetum tristis, which were represented in our data set by only their holotypes. Nonetheless, these two associations were validly published and their names have priority over Portenschlagiello ramosissimae-Campanuletum portenschlagianae and Inulo verbascifoliae-Centaureetum cuspidatae (Terzi and Di Pietro Citation2016; Jasprica and Terzi Citation2017). Therefore, we consider only one association – whose correct name is Moltkio petraeae-Inuletum verbascifoliae, characterized by Campanula portenschlagiana.
The Portenschlagiello ramosissimae-Campanuletum portenschlagianae and Inulo verbascifoliae-Centaureetum cuspidatae are here lowered to the rank of subassociations of the Moltkio petraeae-Inuletum verbascifoliae as M.p.-In.v. fibigietosum triquetrae and M.p.-In.v. centaureetosum cuspidatae, respectively. Actually, the distribution range of the stenoendemic Centaurea cuspidata, restricted to the southern part of the Biokovo ridge, is included within the distribution area of Fibigia triquetra. However, these two stenoendemics species seem to be mutually exclusive, at least in the relevés included in the data set. In fact, Jasprica and Terzi (Citation2017) collected 10 relevés of the Portenschlagiello ramosissimae-Campanuletum portenschlagianae with no record of Centaurea cuspidata, whereas the relevés of the Inulo verbascifoliae-Centaureetum cuspidatae carried out by Trinajstić (Citation1980, Citation1987) include no record of Fibigia triquetra. For this reason, we have considered three subassociations of the Moltkio petraeae-Inuletum verbascifoliae: the typical one (M.p.-In.v. typicum), the M.p.-In.v. centaureetosum cuspidatae characterized by Centaurea cuspidata and the M.p.-In.v. fibigietosum triquetrae, characterized by Fibigia triquetra.
Holotypus: rel. J1, tab. 6, p. 126, in Lovrić and Rac (Citation1987); Locus classicus: Mt. Biokovo; Distribution area: Central and Southern Dalmatia; Diagnostic taxa: Campanula portenschlagiana, Centaurea cuspidata, Fibigia triquetra and locally Cerinthe minor subsp. auriculata. Frequent and dominant taxa: Campanula pyramidalis, Inula verbascifolia, Portenschlagiella ramosissima, C. portenschlagiana.
Seslerio robustae-Putorietum calabricae
This association is represented in our dataset by the relevés of the Birač’s thesis (1971) from which the original diagnosis of the association was derived and effectively published 2 years later (Birač Citation1973). The lectotypus hoc loco of the Seslerio robustae-Putorietum calabricae is relevé 6 of tab. II in Birač (Citation1971), as here reported: Putoria calabrica 3, Inula verbascifolia 2, Parietaria judaica 1, Centaurea glaberrima 1, Sesleria robusta +, Campanula pyramidalis +, Asplenium ceterach +, Micromeria juliana +, Brachypodium retusum +, Helichrysum italicum +, Dittrichia viscosa +, Aurinia leucadea +, Euphorbia dendroides +, Lavandula latifolia +, Picris hispidissima +, Sonchus asper subsp. glaucescens +, Smyrnium olusatrum +.
Locus classicus: The Ombla Estuary near the city of Dubrovnik; Distribution area: Southern Dalmatia, Montenegro and Northwestern Albania; Diagnostic taxa: Putoria calabrica, Sesleria robusta, Aurinia leucadea, Lavandula latifolia. Frequent and dominant taxa: Putoria calabrica, Inula verbascifolia, Campanula pyramidalis s.l., Sesleria robusta.
Teucrio arduinii-Seselietum globiferi
This vegetation exhibits a wide altitudinal range, from sea level (e.g. in the Kotor Bay, Montenegro) up to 1,000 m a.s.l., in Bosnian and Herzegovinian canyons (Lovrić et al. Citation2002, Milović Citation2015). Additionally, stands of this community show some similarities with the thermophilous chasmophytic vegetation of walls (Jasprica et al. Citation2017).
Holotypus: rel. 7, tab. 1, p. 45, in Lovrić et al. (Citation2002); Locus classicus: The Ombla Estuary, near the city of Dubrovnik; Distribution area: Dalmatia, Southern Bosnia and Herzegovina, Montenegro; Diagnostic taxa: Seseli globiferum, Hieracium waldsteinii subsp. plumulosum (d), Teucrium arduinii; Frequent taxa: Seseli globiferum, Campanula pyramidalis s.l., Alyssoides utriculata, Asplenium ceterach subsp. ceterach; Dominant taxa: Seseli globiferum.
Campanulo austroadriaticae-Moltkietum petraeae
This association was originally described as a subassociation (campanuletosum pyramidalis) of the “Campanulo-Moltkietum petraeae Horvatić Citation1963“ and classified in the Centaureo-Campanulion (Tomić-Stanković Citation1970). Subsequently, the subassociation name was corrected (by using Campanula austroadriatica instead of C. pyramidalis) and the syntaxon was raised to the rank of association (Terzi et al. Citation2017, Citation2018). The Campanulo austroadriaticae-Moltkietum petraeae, is in an intermediate syntaxonomic position between the Centaureo cuspidatae-Portenschlagiellion and the Edraianthion tenuifolii (Terzi et al. Citation2017, Citation2018). The association describes the thermophilous communities of the Mt. Lovćen towards Kotor Bay and is substituted by the Saxifrago crustatae-Moltkietum petraeae (Tomić-Stanković ex Terzi et al. Citation2017) Terzi et al. Citation2018 in colder habitats. The latter association contains some typical taxa of less xerothermic conditions, and it clearly belongs to the Edraianthion tenuifolii (compare Terzi et al. Citation2018). For this reason, the Saxifrago crustatae-Moltkietum petraeae has not been considered in this revision.
Holotypus: rel. 11, tab. II, in Tomić-Stanković (Citation1970; see: Terzi et al. Citation2018); Locus classicus: Tisove ploče, Mt. Lovćen; Distribution area: Southeastern Adriatic coast (Croatia, Montenegro); Diagnostic taxa: Campanula austroadriatica, Silene tommasinii, Peucedanum longifolium (d), Achnatherum calamagrostis (d); Frequent taxa: Moltkia petraea, Campanula austroadriatica, Asplenium trichomanes, Cephalaria leucantha, Tanacetum cinerariifolium; Dominant taxa: Moltkia petraea, Campanula austroadriatica, Achnatherum calamagrostis.
Conclusion
The phytocoenological diversity of the limestone cliffs of Central and Southern Dalmatia has been hidden behind different syntaxonomic interpretations, involving two overlapping orders, Moltkietalia petraeae and Centaureo-Campanuletalia. The proposal to restrict the distribution range of the Moltkietalia petraeae to the montane and alpine belts of the Central and Southern Dinarides, as provided with the EVC, has resolved the problem of the ecological overlapping of this order with the Centaureo-Campanuletalia, which was restricted to the coastal and subcoastal Adriatic area.
However, the syntaxonomic and synchorological relationships between the subcoastal associations already classified in the Moltkietalia petraeae and in the Centaureo-Portenschlagiellion (Centaureo-Campanuletalia) have remained unclear.
This revision has shown that four associations (Moltkio petraeae-Inuletum verbascifoliae, Fibigio triquetrae-Cerinthetum tristis, Inulo verbascifoliae-Centaureetum cuspidatae, and Portenschlagiello ramosissimae-Campanuletum portenschlagianae) are not sufficiently distinguishable to be treated as autonomous associations. As a consequence they have been reunited as a sole association whose name is the earliest validly published one, Moltkio petraeae-Inuletum verbascifoliae.
The Inulo verbascifoliae-Centaureetum cuspidatae (i.e. the nomenclatural type of the Centaureo-Portenschlagiellion) and Portenschlagiello ramosissimae-Campanuletum portenschlagianae have been here treated as subassociations of the Moltkio petraeae-Inuletum verbascifoliae.
Moreover, the Campanulo austroadriaticae-Moltkietum petraeae – which is intermediate between the Centaureo-Portenschlagiellion and the Edraianthion tenuifolii (Terzi et al. Citation2018) – has to be classified in the latter alliance for the occurrences of several taxa typical of cool conditions.
This revision has clarified the syntaxonomic diversity of the Centaureo-Portenschlagiellion, which includes only seven associations, whereas the others already described in phytosociological literature are invalidly described (e.g. Moltkio petraeae-Centaureetum voraginicolae, Puccinellio-Aurinietum leucadae, Aurinio-Brassicetum frutescentis) or are here treated as syntaxonomic synonyms of the previous ones.
It is well known that the Centaureo-Campanuletalia order is characterized by numerous endemic and rare plant taxa and communities on both sides of the Adriatic Sea (e.g. Di Pietro and Wagensommer Citation2008; Nikolić et al. Citation2015) and this revision represents a contribution to the knowledge of coenological diversity of the xerothermic cliffs vegetation in the eastern part of the Adriatic Basin.
Nonetheless, this study also highlights the need for further investigations in the less known areas, such as the Vis Archipelago (Middle Adriatic, Croatia) – an area rich in many stenoendemic taxa of exceptional phytogeographical importance, such as Puccinellia teyberi, Asperula visianii, Centaurea friderici, C. crithmifolia – or the coastal and subcoastal areas of Montenegro and Albania which are still poorly known from a phytosociological point of view and of great importance to identify the southern boundary of the distribution area of the Centaureo-Portenschlagiellion.
Nomenclature
Taxonomic nomenclature follows Flora Croatica Database (https://hirc.botanic.hr/fcd/, accessed on March 2018), and The Plant List (http://www.theplantlist.org/, accessed on March 2018) for taxa not included there. Syntaxonomic nomenclature follows Terzi et al. (Citation2017), and Škvorc et al. (Citation2017) for syntaxa not included in Terzi et al. (Citation2017).
Supplemental Material
Download PDF (675.4 KB)Acknowledgements
The authors thank Dr Nikša Glavić for preparation of the map of the study area and Steve Latham (UK) for improving the English and two anonymous referees for their useful suggestions.
Disclosure statement
No potential conflict of interest was reported by the authors.
Related Research Data
References
- Birač V. 1971. Biljni pokrov Srđa i okolice Dubrovačke rijeke [Vegetation of the Srđ Hill and the Rijeka Dubrovačka area] [M.Sc. thesis]. Zagreb, HR: Faculty of Sciences, University of Zagreb.
- Birač V. 1973. Vegetacija Srđa i okolice Dubrovačke rijeke [Vegetation of the Srđ Hill and the area of the Rijeka Dubrovačka]. Acta Bot Croat. 22:135–170.
- Di Pietro R, Wagensommer RP. 2008. Analisi fitosociologica su alcune specie rare e/o minacciate del Parco Nazionale del Gargano (Italia centro-meridionale) e considerazioni sintassonomiche sulle comunità casmofitiche della Puglia [Phytosociological analysis of some rare and/or threatened species of the Gargano National Park (Central-Southern Italy) and syntaxonomic considerations on the chasmophytic communities of Apulia]. Fitosociologia. 45:177–200.
- Dufrêne M, Legendre P. 1997. Species assemblages and indicator species: the need for a flexible asymmetrical approach. Ecol Monogr. 67:345–366.
- European Commission. 2013. Interpretation Manual of European Union Habitats – Eur 28. Brussels, BE: European Commission DG Environment, Nature ENV B.3.
- Hećimović S. 1984. Vegetation der inseln Bobara und Mrkan [Vegetation of the islets of Bobara and Mrkan. Acta Bot Croat. 43:109–118.
- Horvat I. 1942. Biljni svijet Hrvatske [The plant world of Croatia]. Zemljopis Hrvatske. 13:83–481.
- Horvatić S. 1963. Vegetacijska karta otoka Paga s općim pregledom vegetacijskih jedinica Hrvatskog primorja [Vegetation map of the island of Pag with survey of the plant communities of Hrvatsko Primorje]. Prirodosl istraž. 33(IV):3–187.
- Horvatić S. 1971. Osnovne vegetacijske jedinice primorskog krša i pitanje njihove pojačane zaštite [Basic vegetation units of the coastal karst areas and their protection]. In: Varićak T, editor. Simpozij o zaštiti prirode u našem kršu. Zagreb, HR: Jugoslavenska akademija znanosti i umjetnosti; p. 109–135.
- Jasprica N, Pandža M, Milović M, Dolina K. 2017. The wall vegetation (Cymbalario-Parietarietea diffusae Oberdorfer 1969) of the Croatian historical coastal cities (NE Mediterranean). In: Millaku F, Berisha N, Krasniqi E, editors. Book of abstract of the 37th meeting of the Eastern Alpine and Dinaric Society for vegetation ecology. Peć, XK: University of Haxhi Zeka. p. 34.
- Jasprica N, Terzi M. 2017. Limestone cliff vegetation of Portenschlagiello ramosissimae-Campanuletum portenschlagianae, Croatia. Nat Croat. 26(2):331–337.
- Jovanović B, Lakušić R, Rizovski R, Trinajstić I, Zupančič M. 1986. Prodromus Phytocoenosum Jugoslaviae: ad mappam vegetationis m 1:200 000. Bribir-Ilok, HR: Naučno veće vegetacijske karte Jugoslavije.
- Lakušić R. 1968. Planinska vegetacija jugoistočnih Dinarida [The mountain vegetation of the south-eastern Dinaric Alps]. Glasn Republ Zavoda Zastitu Prir Prirodnjačkog Muzeja Titograd. 1:9–75.
- Lakušić R. 1970. Die Vegetation der sudostlichen Dinariden [The vegetation of the south-eastern Dinaric Alps]. Vegetatio. 21:321–373.
- Lovrić AŽ, Bedalov M. 1987. Ekološko-geobotanički pregled primorskih i Kanjonskih endema zapadnih Dinarida i Jadranskih otoka [Ecological and geobotanical revision of coastal and canyon endemics in West Dinaric karst and Adriatic islands]. In: Berberović L, Hadžibeganović, Vuković, T, editors. Symposium on: Protection of endemics in the living world of Yugoslavia, May 15–16, 1986. Sarajevo, YU: Akademia nauka i umenosti Bosne i Hercegovine; p. 167–198.
- Lovrić AŽ, Rac M. 1987. Fitocenološka analiza vegetacije biokovskog područja [Phytosociological analysis of the vegetation of Mt. Biokovo region]. Acta Biokov. 4:97–142.
- Lovrić AŽ, Rac M. 1989. Florističke osobitosti i zaštita fitocenoza u rječnim kanjonima Dalmacije (Cetina, Krka i Zrmanja) [Floristic peculiarities and the phytocoenoses protection in the Dalmatian canyons]. Acta Biokov. 5:105–120.
- Lovrić AŽ, Rac M. 1991. A comparison of the coastal vegetation of Greece, Yugoslavia, Bulgaria, and Turkey. Bot Chron. 10:315–324.
- Lovrić AŽ, Rac M, Šolić ME. 2002. Biljnogeografska sličnost kanjona Dalmacije i Hercegovine [Phytogeographical similarity among canyons in Dalmatia and Herzegovina]. Ekološke Monografije. 5:39–49.
- McCune B, Grace JB. 2002. Analysis of ecological communities. Oregon: MjM Software, Gleneden Beach.
- McCune B, Mefford MJ. 2011. PC-ORD. Multivariate analysis of ecological data. Oregon: MjM Software, Gleneden Beach.
- Meier H, Braun-Blanquet J. 1934. Prodrome Des Groupements Vegetaux Fasc. 2 (Classe des Asplenietales rupestres. Groupements rupicoles) [The Asplenietales rupestres class. Rupicolous groups]. Montpellier: Mari-Lavit.
- Milanović Đ, Brujić J, Đug S, Muratović S, Lukić Bilela L. 2015. Field guide to Natura 2000 habitat types in Bosnia and Herzegovina. Brussels, B: Prospect C&S s.a.
- Milović M. 2015. Seseli L. In: Nikolić T, Milović M, Bogdanović S, Jasprica N, editors. Endemi u hrvatskoj flori [Endemics in flora of Croatia]. Zagreb, HR: Alfa d.d. pp. 418–431.
- Mucina L, Bültmann H, Dierßen K, Theurillat JP, Raus T, Čarni A, Šumberová K, Willner W, Dengler J, Garcia RG, et al. 2016. Vegetation of Europe: hierarchical floristic classification system of vascular plant, bryophyte, lichen, and algal communities. Appl Veg Sci. 19(Suppl. 1):3–264.
- Nikolić T, Milović M, Bogdanović S, Jasprica N, editors. 2015. Endemi u hrvatskoj flori [Endemics in flora of Croatia]. Zagreb, HR: Alfa d.d.
- Otýpková Z, Chytrý M. 2006. Effects of plot size on the ordination of vegetation samples. J Veg Sci. 17:465–472.
- Pavletić Z. 1973. Flora i vegetacija Biševa s posebnim obzirom na biljnogeografski položaj otoka [Flora and vegetation of Biševo with special regard to the biogeographic position of the island]. [Ph.D. thesis]. Zagreb, HR: Faculty of Sciences, University of Zagreb.
- Petrović D, Hadžiablahović S, Vuksanović S, Mačić V, Lakušić D. 2012. Katalog tipova staništa Crne Gore značajnih za Evropsku uniju [A list of habitat types of Montenegro important for the European Union]. Podgorica-Beograd, MNE-SRB: Montenegrin Regional Environmental Center.
- Škvorc Ž, Jasprica N, Alegro A, Kovačić S, Franjić J, Krstonošić D, Vraneša A, Čarni A. 2017. Vegetation of Croatia: phytosociological classification of the high-rank syntaxa. Acta Bot Croat. 76(2):200–224.
- Terzi M, Di Pietro R. 2016. Nomenclatural remarks on the chasmophytic vegetation of the Centaureo-Campanuletalia. Plant Biosyst. 150(6):1364–1369.
- Terzi M, Jasprica N, Caković D. 2017. Xerothermic chasmophytic vegetation of the central Mediterranean Basin: a nomenclatural revision. Phytocoenologia. 47(4):365–383.
- Terzi M, Jasprica N, Caković D, Di Pietro R. 2018. Revision of the central Mediterranean xerothermic cliff vegetation. Appl Veg Sci. 21(3):514–532.
- Tomić-Stanković K. 1970. Vegetacija Lovćena u Crnoj Gori [Vegetation of Mt. Lovćen in Montenegro]. Zajednica Naučnih Ustanova Kosova, Studije, Knj. 1:77–93.
- Trinajstić I. 1964. Moltkea petraea (Tratt.) Gris. u biljnom pokrovu otoka Korčule [Moltkea petraea (Tratt.) Gris. in the vegetation cover of the island of Korčula]. Acta Bot Croat. 23:157–160.
- Trinajstić I. 1980. Apercu syntaxonomique de la vegetation des rochers de l’espace Adriatique [Syntaxonomic overview of the vegetation of rocks in the Adriatic Basin]. Studia Geobot. 1:203–213.
- Trinajstić I. 1987. Sintaksonomski pregled biljnih zajednica planine Biokovo [Syntaxonomic survey of the plant communities of the Mt. Biokovo]. Acta Biokov. 4:143–174.
- Trinajstić I. 1995. Vegetacijske značajke otoka Mljeta [Vegetational characteristics of the island of Mljet]. In: Benović A, Durbešić P, editors. Simpozij Prirodne značajke i društvena valorizacija otoka Mljeta. Pomena, Mljet, HR: Hrvatsko ekološko društvo. p. 247–269.
- Trinajstić I. 1998. Fitogeografsko raščlanjenje klimazonalne šumske vegetacije Hrvatske [Plantgeographycal division of climazonal forest vegetation of Croatia]. Šumar List. 122:407–421.
- Trinajstić I. 2008. Biljne zajednice Republike Hrvatske [Plant communities of Croatia]. Zagreb, HR: Akademija šumarskih znanosti.
- van der Maarel E. 1979. Transformation of cover-abundance values in phytosociology and its effects on community similarity. Vegetatio. 39:97–114.
- Weber HE, Moravec J, Theurillat J-P. 2000. International code of phytosociological nomenclature. 3rd edition. J Veg Sci. 11(5):739–768.
- Wendelberger G. 1963. Die Schwarzföhrenwälder Südosteuropas [The black pine forests of the southeastern Europe]. Mitt Florist Soziol Arb gem NF. 10:130–148.
- Wendelberger G. 1965. Zur Vegetationsgliederung Südosteuropas [The vegetation classifcation of the Southeastern Europe]. Mitt Naturwiss Ver Steiermark. 95:245–286.