
Abstract
Macrofungi play important roles in forest ecology as wood decayers, symbionts, and pathogens of living trees. For the effective forest management, it is imperative to have a comprehensive overview of macrofungi diversity in specific areas. As a part of the National Institute of Biological Resources projects for discovering indigenous fungi in Korea, we collected macrofungi in Gayasan National Park from 2017 to 2018. These specimens were identified based on morphological characteristics and sequence analysis of internal transcribed spacer (ITS) or the nuclear large subunit rRNA (LSU) region. We discovered 17 macrofungi new to Korea: Butyrea japonica, Ceriporia nanlingensis, Coltricia weii, Coltriciella subglobosa, Crepidotus crocophyllus, Cylindrobasidium laeve, Fulvoderma scaurum, Laetiporus cremeiporus, Lentinellus castoreus, Leucogyrophana mollusca, Marasmius insolitus, Nidularia deformis, Phaeophlebiopsis peniophoroides, Phanerochaete angustocystidiata, Phlebiopsis pilatii, Postia coeruleivirens, and Tengioboletus fujianensis. We described their detailed morphological characteristics.
1. Introduction
Macrofungi are fungal species that form large fruiting bodies visible by the naked eye [Citation1]. They commonly include Basidiomycota and Ascomycota with large spore bearing structures, but also a few Zygomycota. Macrofungal species have crucial roles in ecosystems for nutrient cycles and wood decomposition. Depending on the nutritional modes, they can be classified into three groups: saprotrophs, symbiotrophs, and pathotrophs [Citation2]. Saprotrophs decompose organic matters and contribute to cycling of various elements, such as carbon, nitrogen, and oxygen [Citation3]. Symbiotrophs mobilize nutrients from soil to plants with mycorrhiza, helping the survival of plants [Citation4]. Pathotrophs cause disease in living plants, leading to economic loss, but at the same time, play an essential role of maintaining the ecological balance in forest [Citation5]. Some wild macrofungi are useful to human and animal for their nutritional and medicinal properties [Citation6].
Due to their significant ecological and economic importance, it is necessary to understand macrofungal diversity based on trustworthy and comprehensive information for their practical conservation [Citation7,Citation8]. Macrofungi have strong correlation with plant species diversity, and thus act as important indicators of forest community dynamics [Citation9]. In addition, fungal community studies often reveal crucial information, including of rare and edible fungi [Citation10]. An exhaustive fungal survey of an interested area, often achieved by repeated visits, can be useful for discovering new or unrecorded species.
Gayasan is located in Gyeongsangnam-do and Gyeongsangbuk-do at the eastern part of South Korea. For the preservation of the ecosystem and protection of biodiversity, Gayasan was designated as the National Park of Korea in 1972. Gayasan National Park has a well-preserved natural ecosystem which has been often surveyed for its composition and diversity of flora, fauna, and macrofungi [Citation11–13]. Two previous fungal investigations of this region in 2007 and 2016 [Citation14] confirmed that Gayasan has high levels of macrofungal diversity. As a part of the projects by the National Institute of Biological Resources, we surveyed the macrofungi in Gayasan National Park from 2017 to 2018, under the objective of better understanding fungal diversity of the area and finding new or unrecorded fungal species.
For the identification of fungal species, we employed both morphological observation and molecular analysis. Many macrofungi cannot be identified at the species level using only morphological characters due to their morphological similarities. Sequence-based identification of fungi is now commonly used to overcome the limitation of morphology-based identification [Citation15,Citation16]. The collected samples were initially identified by the analysis of the ribosomal RNA sequence, then re-confirmed and described morphologically by microscopic observation. In this study, we identified 17 macrofungal species new to Korea based on morphological and molecular analyses and provided the detailed morphological descriptions.
2. Materials and methods
2.1. Sampling
Gayasan National Park (35°45′00″∼35°49′30″N, 128°02′30″∼128°09′30″E) is located across Hapcheon-gun and Geochang-gun of Gyeongsangnam-do and Goryeong-gun, Sungju-gun, and Gimcheon-si of Gyeongsangbuk-do, South Korea. Specimens were collected from 2017 to 2018 in Gayasan National Park. Dried specimens were deposited in the Seoul National University Fungus Collection (SFC).
Figure 1. Fruiting bodies of the 17 unrecorded species in Korea. (A) Crepidotus crocophyllus; (B and C) Cylindrobasidium laeve; (D) Marasmius insolitus; (E) Nidularia deformis; (F) Leucogyrophana mollusca; (G) Tengioboletus reticulatus; (H) Coltricia weii; (I) Coltriciella subglobosa; (J) Fulvoderma scaurum; (K) Butyrea japonica; (L) Ceriporia nanlingensis; (M) Laetiporus cremeiporus; (N) Phaeophlebiopsis peniophoroides; (O) Phanerochaete angustocystidiata; (P) Phlebiopsis pilatii; (Q) Postia coeruleivirens; (R) Lentinellus castoreus.

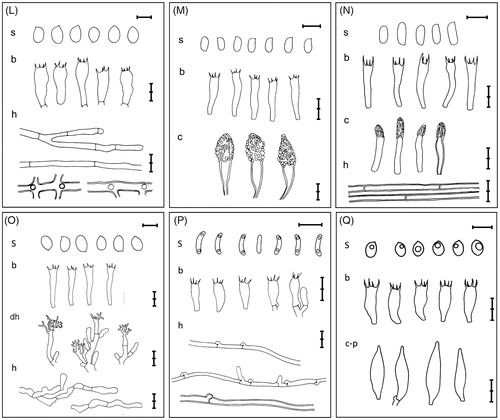
Figure 2. Microscopic feature of the 17 unrecorded species in Korea. (A) Crepidotus crocophyllus; (B) Cylindrobasidium laeve; (C) Marasmius insolitus; (D) Nidularia deformis; (E) Leucogyrophana mollusca; (F) Tengioboletus reticulatus; (G) Coltricia weii; (H) Coltriciella subglobosa; (I) Fulvoderma scaurum; (J) Butyrea japonica; (K) Ceriporia nanlingensis; (L) Laetiporus cremeiporus; (M) Phaeophlebiopsis peniophoroides; (N) Phanerochaete angustocystidiata; (O) Phlebiopsis pilatii; (P) Postia coeruleivirens; (Q) Lentinellus castoreus. (s: basidiospores, b: basidia, c: cystidia, c-c:cheilocystidia, c-p:pleurocystidia, cy: cystidioles, dh: dendrohyphidia h: hyphae, p: peridioles; a scale bar = 5µm).

2.2. DNA sequence-based identification
Genomic DNA was extracted using a modified CTAB extraction protocol [Citation17]. The internal transcribed spacer (ITS) region, a fungal barcode sequence [Citation18], was amplified in a C1000 thermal cycler (Bio-Rad, Richmond, CA) with primer sets of ITS1F/ITS4 [Citation19] or ITS1F/ITS4B [Citation20], as described in previous studies [Citation21]. For Tengioboletus fujianensis, we used primer sets of LR0R/LR5 to amplify the nuclear large subunit rRNA (LSU) region [Citation22,Citation23]. The PCR products were purified with ExpinTM PCR Purification Kit (GeneAll Biotechnology, Seoul, Korea) according to the manufacturer’s instructions. DNA sequencing was performed with corresponding PCR primer sets at Macrogen (Seoul, Korea) using ABI Prism 3700 Genetic Analyzer (Life Technologies, Gaithersburg, MD). Prior to the molecular identification, each sequence was compared with reference sequences in GenBank database using BLAST. Sequences were edited and aligned using MAFFT v7 [Citation24]. Neighbor-joining phylogenetic analyses were performed in MEGA6 using the Kimura 2-parameter model and 1000 bootstrap replicates for tree inference. All the sequences of 17 species have been deposited in GenBank ().
Table 1. Molecular and morphological identification of unrecorded species obtained from Gayasan National Park.
2.3. Morphological observation
Identifications of the specimens were confirmed by their macro- and microscopic features, following the published descriptions. Microscopic observation was performed with 5% (w/v) KOH, 1% (w/v) phloxine, Congo red, and Melzer’s reagent (IKI), using a Nikon SMZ1500 dissecting microscope and a Nikon Eclipse 80i optical microscope (Nikon, Tokyo, Japan). We measured basidia (n = 20/specimen), cystidia (n = 20/specimen), and basidiospores (n = 20/specimen).
3. Results and discussion
A total of 17 species were identified as unrecorded species in Korea using phylogenetic analyses of ITS or LSU sequences (Supporting Information Figure 1 ∼ 6). Most species were identified by ITS phylogenetic trees, but T. fujianensis was identified based on LSU phylogenetic analysis [Citation25]. All species showed 98.8%−100% sequence similarity with previously reported species and formed distinct clades (92–100 in Neighbor-joining bootstrap values). Subsequently, we observed morphology of each specimen and confirmed the identification by phylogenetic analyses. Among 17 unrecorded species, seven species were included in order Polyporales and four in order Agaricales. Other species belonged to order Hymenochaetales (3 species), Boletales (2 spp.), and Russulales (1 sp.). Eight unrecorded species had resupinate basidiocarps. The reverse taxonomy approach was appropriate for these species due to their subtle macro-morphological differences [Citation26].
Five genera were new to Korea: Butyrea Miettinen, Fulvoderma L.W. Zhou & Y.C. Dai, Nidularia Fr. & Nordholm, Phaeophlebiopsis Floudas & Hibbett, and Tengioboletus G. Wu & Zhu L. Yang. Butyrea was segregated from Junghuhnia [Citation27], Fulvoderma from Pyrrhoderma [Citation28], and Phaeophlebiopsis from Phlebiopsis [Citation29]. Morphologically, Butyrea has a straw colored basidiocarp and cylindrical basidiospores, whereas Junghuhnia bears a whitish basidiocarp and ellipsoid basidiospores [Citation27]. The basidiocarp of Fulvoderma is yellowish brown, in contrast to that of Pyrrhoderma which is reddish brown to almost black [Citation28]. In the case of Phaeophlebiopsis and Phlebiopsis which form distinct clades in the molecular phylogenetic analysis, there is no apparent character to distinguish two genera [Citation29]. Therefore, phylogenetic analyses were vital to correctly identify species belonging to these genera.
Some species have long been misidentified as other species or genus because of the similarities in morphological features. These species are difficult to identify correctly without molecular phylogenetic analyses. To date, Cylindrobasidium laeve has been misidentified as C. evolvens [Citation30–33] because of the shared morphological characters, such as almost resupinate and grayish basidiocarp. In the case of Laetiporus cremeiporus, the accurate identification was difficult due to overlapping micro- and macro-morphological characteristics with L. surphureus, of which are orange pileus and cream to white pores [Citation30,Citation31,Citation34,Citation35]. Postia coeruleivirens also has been incorrectly identified as P. caesia [Citation30,Citation34–36] because of the shared characters between two species: conchate and whitish basidiocarp sometimes with bluish flecks. Nidularia deformis and T. fujianensis have long been identified as members of Cyathus and Xerocomus, respectively, based on morphological observations [Citation33,Citation37]. Overlooking the presence of funicular cords under peridioles caused misidentification of Nidularia as Cyathus [Citation38]. In the past, boletes with tomentose, dry, and brownish pileus were treated as Xerocomus in Korea [Citation30,Citation31,Citation34,Citation35,Citation39]. However, some distinctive characters of Tengioboletus, such as white to yellow hymenophore and context without color change when broken, make it possible to distinguish Tengioboletus from Xerocomus [Citation25].
Numerous misidentifications found in the past reports suggest that much fungal diversity of Korea is yet to be discovered. In order to identify macrofungi precisely, molecular analysis must be performed with a well-curated macrofungal sequence database and coupled with morphological observation [Citation16]. Backed by exhaustive and continuous fungal surveys, the combined approach will increase the possibility of discovering more unrecorded or novel macrofungi in Korea.
4. Taxonomy
4.1. Order Agaricales
4.1.1. Crepidotus crocophyllus (Berk.) Sacc., Syll. fung. (Abellini) 5: 886 (1887)
Pileus 10–65 mm in diam., semicircular to fan-shaped, broadly convex, sometimes applanate, whitish to ocherous brown or pale orange brown, with reddish brown fibrils or small scales, sometimes with whitish to pale orange fuzz around the attachment part to the substrate. Lamellae crowded, whitish when young, becoming brown when mature. Cheilocystidia subcylindrical to clavate, sometimes subutriform, 20–35 × 5–10 µm. Basidia clavate to narrowly clavate, 25–47 × 6–9 µm, with a basal clamp and mainly 2-spored, often 4-spored, rarely 1-spored. Basidiospores globose, occasionally subglobose, 5.1–7.2 × 4.9–7.0 µm, finely punctate ( and ).
Specimen examined: Korea. Gyeongsangnam-do Hapcheon-gun, Gayasan National Park, 35°47′41″N 128°05′02″E, 618 m, 7 Sep 2018, H. Lee, N. H. Kim & A. Lupala, SFC20180907-112.
Remarks: This species is characterized by having an orange brown pileus with fibrils or small scales. Unlike the previous description of C. crocophyllus given by Bandala et al. [Citation40], 2-spored basidia were frequently observed in the specimen observed.
4.1.2. Cylindrobasidium laeve (Pers.) Chamuris, Mycotaxon 20(2): 587 (1984)
Basidiocarps resupinate, sometimes semipileate, easily detached from the substrate, whitish spots when young which expand to radial lines and then form cracks, on vertical substrates with pileus-like edges or pileoli; upper surface of exposed edge tomentose, often zonate. Hymenophore uneven to tuberculate, cream to reddish ocher when young, grayish when old, margin white and finely redented when young. Hyphal system monomitic, hyphae thin- to thick-walled, 3–5 μm in diam., septa with clamp connections. Leptocystidia cylindrical to slenderly subclavate, sometimes tortuous, 40–75 × 3–6 μm. Basidia cylindrical to slenderly subclavate, 45–60 × 5–8 μm, 4-spored, with a basal clamp. Basidiospores oval, smooth, hyaline, 7.5–10.3 × 5.2–6.1 μm (, , and ).
Specimen examined: Korea. Gyeongsangnam-do Hapcheon-gun, Gayasan National Park, 35°47′50″N 128°05′44″E, 585 m, 8 Feb 2017, Y. W. Lim, N. K. Kim & H. J. Cho, SFC20170208-07.
Remarks: This species has been misidentified as Cylindrobasidium evolvens in Korea. C. laeve differs from C. evolvens by the shape and dimension of leptocystidia. While the leptocystidia of C. laeve is cylindrical to slenderly subclavate, that of C. evolvens is fusiform and wider (45–70 × 5–7 μm) [Citation41].
4.1.3. Marasmius insolitus Kiyashko & E.F. Malysheva, Phytotaxa 186(1): 14 (2014)
Pileus 8–20 mm in diam., hemispherical to convex, sometimes indistinctly umbonate, slightly pruinose when young, pale yellow with slightly darker center; margin slightly inflexed. Lamellae adnate, crowded, with 2–3 series of lamellulae, whitish to pale cream; lamellar edge serrulate. Stipe 30–60 × 2–3 mm, cylindrical, hollow, pruinose, pale cream in upper part, grayish brown at base, with whitish basal tomentum. Cheilocystidia numerous, thick-walled, broom-shaped, main body subcylindrical to subclavate, sometimes irregular in outline, 12.3–28.8 × 3.8–7.1 μm, thick-walled apical projections up to 15 μm long. Pleurocystidia numerous, similar to cheilocystidia in shape and size. Basidia clavate, 22.7–29.4 × 5.5–7.2 µm, 4-spored. Basidiospores ellipsoid, 9.4–12.5 × 3.7–4.4 µm, smooth, thin-walled, hyaline ( and ).
Specimen examined: Korea. Gyeongsangnam-do Hapcheon-gun, Gayasan National Park, 35°47′11″N 128°05′09″E, 653 m, 7 Sep 2018, H. Lee, N. H. Kim & A. Lupala, SFC20180907-112.
Remarks: M. insolitus is characterized by having a pale yellowish pileus, a pruinose stipe, and broom-like cystidia. The original description of this species [Citation42] states that the pileus and the stripe are similar in color. In this specimen, however, the color of the stipe is darker than the pileus.
4.1.4. Nidularia deformis (Willd.) Fr., in Fries & Nordholm, Symb. gasteromyc. (Lund) 1: 3 (1817)
Basidiocarp small, 3–10 mm in diam., globose to subglobose, often pea-shaped, tuberculate; sessile and broadly attached at the base. Peridium creamy white to grayish cinnamon, floccose, inner side smooth, completely enclosing when young, cover falling away when mature. Peridioles 1–1.8 mm in diam., lenticular, brown to coffee brown, smooth and glossy, immersed in mucus. Peridial hyphae 2–7 µm in diam., with many spinose branches, pale brown, with slightly thickened wall. Basidia not observed. Basidiospores 6.4–8.1 × 3.8–4.6 µm, broadly ellipsoid to ovoid, hyaline, with slightly thickened wall ( and ).
Specimen examined: Korea. Gyeongsangbuk-do Seongju-gun, Gayasan National Park, 35°48′25″N 128°08′24″E, 622 m, 8 Sep 2017, H. J. Cho, K. H. Park & N. H. Kim, SFC20170908-34.
Remarks: The genus Nidularia is first to be reported in Korea. In the past, a species having peridioles with funicular cords was identified as Cyathus, but the specimen in this study has spiny hyphae and peridioles without funicular cords, which are typical characteristics of the genus Nidularia [Citation38].
4.2. Order Boletales
4.2.1. Leucogyrophana mollusca (Fr.) Pouzar, Česká Mykol. 12(1): 33 (1958)
Basidiocarps annual, resupinate, effused, thin and delicate. Hymenophore merulioid to smooth, bright orange when fresh, buff to pale yellow when dry; margin whitish and floccose to fimbriate when fresh. Subiculum monomitic, subicular hyphae with clamp connections, thin walled, 2–4.5 µm in diam., occasionally branching. Basidia clavate to subclavate, 8–10 × 23–35 µm, 4-spored, with a basal clamp. Basidiospores broadly ellipsoid to ellipsoid, 5.2–6.3 × 4–5.5 µm, smooth, dextrinoid ( and ).
Specimen examined: Korea. Gyeongsangnam-do Hapcheon-gun, Gayasan National Park, 35°47′53″N 128°05′55″E, 640 m, 22 Aug 2017, H. J. Cho & K. H. Park, SFC20170822-61.
Remarks: L. mollusca is morphologically similar to L. pseudomollusca, a species previously reported in Korea. L. molluca is distinguished by the basidiospore size which is smaller than that of L. pseudomollusca (6–7.5 (–9) × 4–5 µm) [Citation41].
4.2.2. Tengioboletus reticulatus G. Wu & Zhu L. Yang, in Wu, Li, Zhu, Zhao, Han, Cui, Li, Xu & Yang, Fungal Diversity 81: 146 (2016)
Pileus 50–120 mm in diam., convex when young, applanate when mature, olive brown to dark brown, smooth, dry. Context pale yellow, non-discoloration when cut. Hymenophore adnate to slightly sinuate, yellow, up to 10 mm thick; pores ca. 1/mm; tubes up to 10 mm long, yellow. Stipe 70–140 × 10–20 mm, subcylindrical to subclavate, light yellow to yellowish brown, with distinct olive brown reticulations, non-discoloration when cut; basal mycelium cream colored. Cheilocystidia scattered, 15–32 × 6–9 μm, cylindrical to subclavate, thin-walled. Pleurocystidia scattered, 30–55 × 8–10 μm, fusiform, thin-walled. Basidia subclavate to clavate, 31–38 × 8–13 μm, 4-spored. Basidiospores subfusiform, 11–14 × 3.5–5.7 μm, yellowish brown, smooth ( and ).
Specimen examined: Korea. Gyeongsangnam-do Hapcheon-gun, Gayasan National Park, 35°47′43″N 128°08′24″E, 622 m, 25 Jul 2018, H. Lee, H. J. Cho & N. H. Kim, SFC20180725-31.
Remarks: Tengioboletus is the genus introduced to Korea by this study. The specimen collected in Korea is generally consistent with the original description of T. reticulatus [Citation25], but the reticulation of stipe is rather dark.
4.3. Order Hymenochaetales
4.3.1. Coltricia weii Y.C. Dai, in Dai, Yuan & Cui, Sydowia 62(1): 16 (2010)
Basidiocarps annual, stipitate, leathery, corky. Pilei circular, applanate to slightly infundibuliform, 25–40 mm in diam. Upper surface orange brown to dark reddish brown, distinctly zonate, velutinous; margin entire, sometimes crenate, often wavy. Pore surface cream to pale buff when young, becoming grayish brown; pores round to angular, 2–4/mm; dissepiments entire, often slightly lacerate. Stipe dark brown, corky, velutinous, 10–17 × 2–3 mm. Hyphal system monomitic, generative hyphae simple septate, yellowish brown, slightly thick-walled, occasionally branched, 5–8 μm in diam. Basidia clavate to subclavate, 15–22 × 7–9.5 μm, 4-spored. Basidiospores mostly broadly ellipsoid, 5.5–6.3 × 4.5–5.2 μm, thick-walled, smooth ( and ).
Specimen examined: Korea. Gyeongsangnam-do Hapcheon-gun, Gayasan National Park, 35°47′41″N 128°05′02″E, 604 m, 25 Jul 2017, H. J. Cho, K. H. Park & N. H. Kim, SFC20170725-23.
Remarks: C. weii is morphologically similar to C. cinnamomea, but the size of basidiospore is smaller than that of C. cinnamomea (5.6–6.8 × 4.3–5.0 μm vs 6.9–8.1 × 5.5–6.4 μm) [Citation43].
4.3.2. Coltriciella subglobosa Y.C. Dai, Fungal Diversity 45: 160 (2010)
Basidiocarps annual, resupinate to reflexed, soft, filaceous when fresh, cottony when dry. Pilei often fused, sometimes imbricate. Upper surface velutinous, grayish brown. Pore surface reddish brown, margin cream to pale yellow; pores round, 3–4/mm; dissepiments slightly lacerate. Hyphal system monomitic, simple septate, yellowish brown, frequently branched, 6–9 µm in diam. Cystidioles present, subcylindrical to slightly ventricose, 5–22 × 6–8 µm. Basidia clavate to subclavate, 19–24 × 8–11 µm, thin-walled, 4-spored. Basidiospores subglobose to broadly ellipsoid, 6.4–8.1 × 5.0–6.2 µm, yellowish brown, verrucose ( and ).
Specimen examined: Korea. Gyeongsangnam-do Hapcheon-gun, Gayasan National Park, 35°48′20″N 128°06′05″E, 692 m, 16 Mar 2017, Y. W. Lim, SFC20170316-01.
Remarks: The specimen in this study morphologically fits the original description of C. subglobosa [Citation44]. C. subglobosa is easily distinguished from other species by effused-reflexed basidiocarp and subglobose basdiospore.
4.3.3. Fulvoderma scaurum (Lloyd) L.W. Zhou & Y.C. Dai, in Zhou, Ji, Vlasák & Dai, Mycologia 110(5): 879 (2018)
Basidiocarps semi-stipitate to stipitate, on dead woods. Pilei applanate, sometimes convex, depressed at the center, up to 75 mm in diam., margin entire. Upper surface with a yellow to brown, becoming blackish brown, glabrous, zonate, rugose, and sulcate, margin white when young; if present, stipe lateral, 60 × 10 mm, glabrous, same or darker color with pilei. Pore surface yellowish brown to brown; pores round, 6–8/mm. Hyphal system monomitic, generative hyphae up to 5 µm in diam., parallel. Basidia clavate, 11–16 × 6–10 µm, 4-spored. Basidiospores globose to subglobose, 5–6.5 × 5–6 µm, smooth, hyaline, sometimes guttulate ( and ).
Specimen examined: Korea. Gyeongsangnam-do Hapcheon-gun, Gayasan National Park, 35°47′51″N 128°05′43″E, 584 m, 8 Sep 2017, H. J. Cho, K. H. Park & N. H. Kim, SFC20170908-69.
Remarks: Fulvoderma is first to be reported in Korea. F. scaurum was used to be identified as Pyrrhoderma scaurum, but Zhou et al. [Citation28] recently introduced a new genus Fulvoderma. P. scaurum was subsequently transferred to the genus Fulvoderma. While the genus Pyrrhoderma has resupinate basidiocarp, the genus Fulvoderma has sessile or stipitate basidiocarp.
4.4. Order Polyporales
4.4.1. Butyrea japonica (Núñez & Ryvarden) Miettinen & Ryvarden, Ann. bot. fenn. 53(3–4): 161 (2016)
Basidiocarps annual, resupinate, up to 80 mm in diam. Pore surface cream to pale yellow; pores circular to angular, 5–7/mm. Hyphal system dimitic, generative hyphae thin-walled, with clamp connections, 2–3 μm in diam., skeletal hyphae thick-walled, 2–4 μm in diam. Cystidia cylindrical to subclavate, 25–38 × 4–7 μm. Basidia clavate, 12–19 × 3.5–5 μm, 4-spored, with a basal clamp. Basidiospores cylindrical, straight to suballantoid, 4–5 × 2–2.4 μm, smooth, inamyloid ( and ).
Specimen examined: Korea. Gyeongsangbuk-do Seongju-gun, Gayasan National Park, 35°48′28″N 128°08′21″E, 654 m, 23 Aug 2017, H. J. Cho & K. H. Park, SFC20170823-29.
Remarks: Butyrea is the genus first reported in Korea from this study. Miettinen & Ryvarden [Citation27] established a new genus Butyrea, which included only two species, B. japonica and B. luteoalba. B. japonica is distinguished from B. luteoalba by the presence of gloeocystidia. In addition, B. japonica is distributed in Asia, while B. luteoalba is distributed in North Europe.
4.4.2. Ceriporia nanlingensis B.K. Cui & B.S. Jia, in Jia & Cui, Mycotaxon 116: 458 (2011)
Basidiocarps annual, resupinate, corky, fragile when dry, up to 0.6 mm thick. Pore surface white to pale pink when fresh, brownish when dry; pores irregular, 3–6/mm; dissepiments mostly thin, entire. Hyphal system monomitic, generative hyphae thin-walled, sometimes thick-walled, often branched, 3–7 µm in diam. Basidia clavate, 18–28 × 3.5–5 µm, 4-spored. Basidiospores ellipsoid to oblong, 3.5–5 × 1.5–2 µm, mostly with guttules ( and ).
Specimen examined: Korea. Gyeongsangnam-do Hapcheon-gun, Gayasan National Park, 35°47′07″N 128°04′59″E, 721 m, 26 Jun 2018, H. Lee, H. J. Cho & N. H. Kim, SFC20180626-06.
Remarks: C. nanlingensis is characterized by having cystidia and a lavender colored pore surface. C. viridans is morphologically similar to C. nanlingensis, but the former does not have cystidia [Citation45].
4.4.3. Laetiporus cremeiporus Y. Ota & T. Hatt., Mycol. Res. 113(11): 1289 (2009)
Basidiocarps annual, imbricate, imbricate clusters up to 400 mm long. Pileus flabelliform, up to 300 mm wide. Upper surface bright orange to reddish, often zonate, radially rugose. Pore surface white to pale cream; pores nearly circular to angular, 2–3/mm, with thin dissepiments. Hyphal system dimitic, generative hyphae thin-walled, rarely branched, 4–11 µm in diam., binding hyphae thick-walled, non-septate, up to 20 µm in diam. Basidia clavate, 16–28 × 5–8 µm, 2- or 4-spored. Basidiospores ellipsoid to oblong, 5–7.5 × 3.5–5 µm ( and ).
Specimen examined: Korea. Gyeongsangnam-do Hapcheon-gun, Gayasan National Park, 35°47′29″N 128°06′01″E, 515 m, 30 Aug 2018, H. Lee, N. H. Kim & A. Lupala, SFC20180830-40; ibid, 35°47′32″N 128°05′52″E, 535 m, 8 Sep 2017, H. J. Cho, K. H. Park & N. H. Kim, SFC20170908-42.
Remarks: Due to the light orange to reddish orange pileus surface and yellowish white to cream pore surface, L. cremeiporus has often been misidentified as L. sulphureus. However, Ota et al. [Citation46] confirmed that L. sulphureus s. lat. was geographically divided into three phylogenetic species (East Asia, Europe, and North America) based on ITS sequences. Although it is difficult to distinguish three species based on morphological characteristics, L. sulphureus in East Asia is identified as L. cremeiporus, in accordance with the phylogenetic analyses and geographical distribution [Citation46].
4.4.4. Phaeophlebiopsis peniophoroides (Gilb. & Adask.) Floudas & Hibbett, Fungal Biology 119(7): 710 (2015)
Basidiocarps annual, entirely resupinate, with cracks. Hymenophore pale gray to yellowish gray, smooth, margin whitish. Hyphal system monomitic, hyphae with frequent branching, 2–6 µm in diam. Cystidia conical, incrusted with crystals, 10–25 × 7–15 µm. Basidia subcylindrical to subclavate, 17–28 × 3–5.5 µm, 4-spored. Basidiospores ellipsoid to oblong, 3.5–5 × 2–2.5 µm, smooth, thin-walled, inamyloid ( and ).
Specimen examined: Korea. Gyeongsangnam-do Hapcheon-gun, Gayasan National Park, 35°48′25″N 128°06′18″E, 743 m, 8 Sep 2017, H. J. Cho, K. H. Park & N. H. Kim, SFC20170908-86.
Remarks: Phaeophlebiopsis is first reported in Korea. Floudas and Hibbett [Citation29] introduced the new genus Phaeophlebiopsis to accommodate several Phlebiopsis-like species. The specimen collected in Korea morphologically corresponds to the original description [Citation47] and is distinguished from other Phlebiopsis species recorded in Korea by having a grayish hymenophore.
4.4.5. Phanerochaete angustocystidiata Sheng H. Wu, Bot. Bull. Acad. Sin., Taipei 41(2): 166 (2000)
Basidiocarps annual, entirely resupinate, membranaceous. Hymenophore whitish gray to light cream, cracked; margin white. Hyphal system monomitic, generative hyphae 2–6 µm in diam., thin-walled to thick-walled. Lamprocystidia cylindrical, subulate, incrusted with crystals at upper part, 25–50 × 4–8 µm. Basidia clavate to subclavate, 17–33 × 3.5–5 µm, 2- or 4-spored. Basidiospores cylindrical, 5.5–7.5 × 2–3 µm, inamyloid ( and ).
Specimen examined: Korea. Gyeongsangnam-do Hapcheon-gun, Gayasan National Park, 35°48′23″N 128°05′41″E, 684 m, 16 Mar 2017, Y. W. Lim, SFC20170316-16.
Remarks: This species is characterized by having narrow cystidia, basidiospore, and lamprocystidia with encrustation. Morphological characteristics of this specimen generally correspond to the original description given by Wu [Citation48], but the color of the hymenophore is more whitish.
4.4.6. Phlebiopsis pilatii (Parmasto) Spirin & Miettinen, in Miettinen, Spirin, Vlasák, Rivoire, Stenroos & Hibbett, MycoKeys 17: 25 (2016)
Basidiocarps annual, phleboid, fragile, cracked. Hymenophore light brown to yellowish brown, smooth to merulioid. Hyphal system monomitic, generative hyphae 2–4 µm in diam., thick-walled, clamp connections lacking. Dendrohyphidia multiple branched, up to 40 µm long. Basidia subcylindrical to subclavate, 22–35 × 5.5–7 µm, 4-spored. Basidiospores ellipsoid to oblong, 5–8 × 4–5 µm, inamyloid ( and ).
Specimen examined: Korea. Gyeongsangnam-do Hapcheon-gun, Gayasan National Park, 35°47′23″N 128°06′05″E, 497 m, 8 Feb 2017, Y. W. Lim, N. K. Kim & H. J. Cho, SFC20170208-01; ibid, 35°47′33″N 128°05′54″E, 538 m, 8 Feb 2017, Y. W. Lim, N. K. Kim & H. J. Cho, SFC20170208-05.
Remarks: P. pilatii has a phlebioid basidiocarp and simple-septate hyphae with thick-walled [Citation49]. In addition, this species is easily distinguished from other species by having a yellowish brown hymenophore and multiple branched dendrohyphidia.
4.4.7. Postia coeruleivirens (Corner) V. Papp, Mycotaxon 129(2): 411 (2015)
Basidiocarps conchate to flabellate, fragile. Upper surface white to cream, often with bluish flecks, pubescent. Pore surface white to pale cream, sometimes with grayish tint; pores 6–8/mm. Hyphal syetem monomitic, thin-walled to slightly thick-walled, 3-6 μm in diam. Basidia clavate to subclavate, 8.5–15 × 3.5–5 μm, 4-spored. Basidiospores cylindrical, 4–6 × 1–1.5 µm ( and ).
Specimen examined: Korea. Gyeongsangnam-do Hapcheon-gun, Gayasan National Park, 35°47′13″N 128°05′06″E, 659 m, 25 Jul 2018, H. Lee, H. J. Cho & N. H. Kim, SFC20180725-73.
Remarks: This species has been misidentified as Postia caesia in Korea. The pore size of P. caesia is larger (4–5/mm) than that of P. coeruleivirens (6–8/mm) [Citation50]. Moreover, the basidiospore width of P. caesia is broader than that of P. coeruleivirens (1.42–1.64 vs 1–1.5 μm in width).
4.5. Order Russulales
4.5.1. Lentinellus castoreus (Fr.) Kühner & Maire, Bull. Trimest. Soc. Mycol. Fr. 50: 16 (1934)
Basidiocarps conchate to flabellate, 15–50 mm in diam., subimbricate. Pileus yellowish brown to reddish brown, tomentose. Lamellae subdistant, up to 4 mm broad, cream to pale brown, margin serrate. Hyphal system trimitic, generative hyphae thin-walled, 3–8 μm in diam., with clamp connections. Pleurocystidia fusiform, 22–30 × 4–7 µm, lanceolate, with basal clamps. Gloeocystidia 20–45 × 5–7 µm, subcylindricla to subclavate, sometimes slightly capitate. Basidia subclavate to clavate, 13–22 × 5–7 µm, 4-spored. Basidiospores subglobose to broadly ellipsoid, 3–4.5 × 3–4 µm, thin-walled, amyloid ( and ).
Specimen examined: Korea. Gyeongsangnam-do Hapcheon-gun, Gayasan National Park, 35°47′53″N 128°05′60″E, 652 m, 26 Jul 2017, H. J. Cho, K. H. Park & N. H. Kim, SFC20170726-44.
Remarks: Because the macromorphology of L. castoreus is very diverse, it is not easy to distinguish between L. castoreus and its closely related species, L. ursinus. However, the former has bigger basidiospores than those of L. ursinus (3–4.6 × 2–3.5 µm) [Citation51].
Supplemental Material
Download PNG Image (55.6 KB){kind=link}
Supplemental Material
Download PNG Image (63.8 KB){kind=link}
Supplemental Material
Download PNG Image (98 KB){kind=link}
Supplemental Material
Download PNG Image (163.7 KB){kind=link}
Supplemental Material
Download PNG Image (117.7 KB){kind=link}
Supplemental Material
Download PNG Image (147.4 KB){kind=link}
Disclosure statement
No potential conflict of interest was reported by the author(s).
Additional information
Funding
References
- Kirk PM, Cannon PF, Stalpers JA. Dictionary of the fungi. 10th ed. Wallingford (UK): CAB International; 2008.
- Mueller GM, Schmit JP, Leacock PR, et al. Global diversity and distribution of macrofungi. Biodivers Conserv. 2007;16(1):37–48.
- Thormann MN. Diversity and function of fungi in peatlands: a carbon cycling perspective. Can J Soil Sci. 2006;86(Special Issue):281–293.
- Read DJ, Perez-Moreno J. Mycorrhizas and nutrient cycling in ecosystems – a journey towards relevance? New Phytol. 2003;157(3):475–492.
- Lapitan PG, Fernando ES, Suh MH, et al. Biodiversity and natural resources conservation in protected areas of Korea and the Philippines. In: Hwang YS, editor. ASEAN-Korea environmental cooperation unit. Seoul: GeoBook Publishing Co.; 2010. p. 248.
- Wani BA, Bodha RH, Wani AH. Nutritional and medicinal importance of mushrooms. J Med Plants Res. 2010;4(24):2598–2604.
- Zotti M, Persiani AM, Ambrosio E, et al. Macrofungi as ecosystem resources: conservation versus exploitation. Plant Biosyst. 2013;147(1):219–225.
- Halme P, Heilmann-Clausen J, Rämä T, et al. Monitoring fungal biodiversity-towards an integrated approach. Fungal Ecol. 2012;5(6):750–758.
- Gabel A, Gabel M. Comparison of diversity of macrofungi and vascular plants at seven sites in the black hills of South Dakota. Am Midl Nat. 2007;157(2):258–296.
- Molina R, Pilz D, Smith J, et al. Conservation and management of forest fungi in the Pacific Northwestern United States: an integrated ecosystem approach. In: Moore D, Nauta NN, Evans SE, Rotheroe M, editors. Fungal conversation issues and solutions. British mycological society symposium series. Vol. 22. Cambridge (UK): Cambridge University Press; 2001. p. 19–63.
- Song MJ, Kim H, Lee BY, et al. Analysis of traditional knowledge of medicinal plants from residents in Gayasan National Park (Korea). J Ethnobiol Ethnomed. 2014;10(1):74.
- You JH, Jeon SK, Seol JW. Flora and conservation plant of Gayasan National Park. J Korea Soc Environ Restor Technol. 2013;16(1):109–130.
- Moon TY, An MS, Kim HG, et al. Distribution of functional feeding and habitat trait groups of benthic macroinvertebrates and biological evaluation of water quality in Gayasan national park. J Environ Sci Int. 2018;27(6):383–399.
- Ko PY, Hong KS, Choe SY, et al. Distribution of spontaneously growing mushrooms in Gayasan national park. J Natl Park Res. 2018;9(1):1–15.
- Hebert PD, Ratnasingham S, De Waard JR. Barcoding animal life: cytochrome c oxidase subunit 1 divergences among closely related species. Proc R Soc Lond B. 2003;270(suppl_1):S96–S99.
- Hibbett D, Abarenkov K, Kõljalg U, et al. Sequence-based classification and identification of fungi. Mycologia. 2016;108(6):1049–1068.
- Rogers SO, Bendich AJ. Extraction of total cellular DNA from plants, algae and fungi. In: Gelvin SB, Schilperoort RA, editors. Plant molecular biology manual. Boston (MA): Kluwer Academic Publishers; 1994. p. 183–190.
- Schoch CL, Seifert KA, Huhndorf S, et al. Nuclear ribosomal internal transcribed spacer (ITS) region as a universal DNA barcode marker for fungi. Proc Natl Acad Sci USA. 2012;109(16):6241–6246.
- White TJ, Bruns T, Lee S, et al. Amplification and direct sequencing of fungal ribosomal RNA genes for phylogenetics. In: Innis MA, Gelfand DH, Sninsky JJ, White TJ, editors. PCR protocols: a guide to methods and applications. San Diego (CA): Academic Press; 1990. p. 315–322.
- Gardes M, Bruns TD. ITS primers with enhanced specificity for basidiomycetes-application to the identification of mycorrhizae and rusts. Mol Ecol. 1993;2(2):113–118.
- Park MS, Fong JJ, Lee H, et al. Delimitation of Russula subgenus Amoenula in Korea using three molecular markers. Mycobiology. 2013;41(4):191–201.
- Hopple JS, Vilgalys R. Phylogenetic relationships among coprinoid taxa and allies based on data from restriction site mapping of nuclear rDNA. Mycologia. 1994;86(1):96–107.
- Wu G, Feng B, Xu J, et al. Molecular phylogenetic analyses redefine seven major clades and reveal 22 new generic clades in the fungal family Boletaceae. Fungal Divers. 2014;69(1):93–115.
- Katoh K, Standley DM. MAFFT multiple sequence alignment software version 7: improvements in performance and usability. Mol Biol Evol. 2013;30(4):772–780.
- Wu G, Li YC, Zhu XT, et al.. One hundred noteworthy boletes from China. Fungal Divers. 2016;81(1):25–188.
- Binder M, Hibbett DS, Larsson KH, et al. The phylogenetic distribution of resupinate forms across the major clades of mushroom‐forming fungi (Homobasidiomycetes). Syst Biodivers. 2005;3(2):113–157.
- Miettinen O, Ryvarden L. Polypore genera Antella, Austeria, Butyrea, Citripora, Metuloidea and Trulla (Steccherinaceae, Polyporales). Ann Bot Fenn 2016;53:157–173.
- Zhou LW, Ji XH, Vlasák J, et al. Taxonomy and phylogeny of Pyrrhoderma: a redefinition, the segregation of Fulvoderma, gen. nov., and identifying four new species. Mycologia. 2018;110(5):872–889.
- Floudas D, Hibbett DS. Revisiting the taxonomy of Phanerochaete (Polyporales, Basidiomycota) using a four gene dataset and extensive ITS sampling. Fungal Biol. 2015;119(8):679–719.
- Jung YS. Natural resource survey of Songnisan National Park. Seoul: Korea National Park Service; 2003. (In Korean).
- Kim JK. Natural resource survey of Deogyusan National Park. Seoul: Korea National Park Service; 2004. (In Korean).
- Lim YW, Lee JS, Jung HS. Fungal Flora of Korea. In: Shin S, editor. Wood rotting fungi. Vol 1. Incheon: Junghaengsa, Inc; 2010. p.164.
- Choi HP. The illustrated big guidebook of Korean mushroom. Seoul: Academybook Publishing Co.; 2015. (In Korean).
- Lee JK, Lim YW, Jeon YC, et al. Research project on the survey of forest biota/Reginal monitoring of the higher fungi in the forests of Gyenggi-do, western Gangwon-do, and western Jeju-do provinces. Pocheon-si: Korea National Arboretum; 2015. (In Korean).
- Lee JK, Lim YW, Ko PY, et al. Research project on the survey of forest biota/Monitoring of the higher fungi in the forests of eastern Jeju Island, southern and western sea islands of Korean Peninsula. Pocheon-si: Korea National Arboretum; 2016. (In Korean).
- Ko BJ. Natural resources monitoring of Jirisan National Park (third year). Gyeongsangnam-do: Garam Publishing Co.; 2004. (In Korean).
- Park WH, Lee JH. New wild fungi of Korea. Seoul: Kyo-Hak Publishing Co.; 2011. (In Korean).
- Sermenli Baş H, Işıloğlu M. A new gasteromycete genus record for Turkey. Turk J Bot. 2009;33(4):323–324.
- Park YD. Natural resources monitoring of Jirisan National Park (fourth year). Gyeongsangnam-do: Garam Publishing Co.; 2005. (In Korean).
- Bandala VM, Montoya L, Mata M. Crepidotus crocophyllus found in Costa Rica and Mexico and revision of related species in subsection Fulvifibrillosi. Mycologia. 2008;100(2):335–346.
- Breitenbach J, Kränzlin F. Fungi of Switzerland. Vol 2. Non-gilled Fungi. Luzern: Verlag Mykologia; 1986. 528 p.
- Kiyashko AA, Malysheva EF, Antonín V, et al. Fungi of the Russian Far East 2. New species and new records of Marasmius and Cryptomarasmius (Basidiomycota). Phytotaxa. 2014;186(1):1–28.
- Niemelä T. Polypores, lignicolous fungi. Norrlinia. 2005;13:1–320.
- Dai YC. Hymenochaetaceae (Basidiomycota) in China. Fungal Divers. 2010;45(1):131–343.
- Jia BS, Cui BK. Notes on Ceriporia (Basidiomycota, Polyporales) in China. Mycotaxon. 2011;116(1):457–468.
- Ota Y, Hattori T, Banik MT, et al. The genus Laetiporus (Basidiomycota, Polyporales) in East Asia. Mycol Res. 2009;113(Pt 11):1283–1300.
- Gilbertson RL, Adaskaveg JE. Studies on wood-rotting basidiomycetes of Hawaii. Mycotaxon. 1993;49:369–397.
- Wu SH. Six new species of Phanerochaete from Taiwan. Bot Bull Acad Sin. 2000;41:165–174.
- Miettinen O, Spirin V, Vlasák J, et al. Polypores and genus concepts in Phanerochaetaceae (Polyporales, Basidiomycota). MycoKeys. 2016;17:1–46.
- Miettinen O, Vlasák J, Rivoire B, et al. Postia caesia complex (Polyporales, Basidiomycota) in temperate Northern Hemisphere. Fungal Syst Evol. 2018;1(1):101–129.
- Miller OK, Stewart L. The genus Lentinellus. Mycologia. 1971;63(2):333–369.