
Abstract
Endophytic fungi occupy inner plant tissues, which results in various interactions between the fungus and host. Studies on endophytic fungi have been conducted in Korea for over 30 years. This paper summarizes the published results of those studies. The endophytic fungi of approximately 132 plant species in Korea have been studied since the 1990s, resulting in over 118 publications. The host plants featured in these studies comprised 3 species of mosses, 34 species of woody plants, and 95 species of herbaceous plants. At the family level, the most studied plants were members of the Poaceae family, covering 18 species. Regionally, these studies were conducted throughout Korea, but over half of the studies were conducted in Gyeongsangbuk-do, Gangwon-do, and Chungcheongnam-do. Relatively few studies have been conducted in a metropolis such as Seoul. We confirmed 5 phyla, 16 classes, 49 orders, 135 families, 305 genera, and 855 taxa of endophytic fungi, excluding Incertae sedis, whose relationship with others are unknown. Most of the endophytic fungi belonged to Ascomycota (93.2%), and a few belonged to Basidiomycota (3.6%). Since the diversity of endophytic fungi differs depending on the host plant, plant tissue, and distribution region, future studies should be conducted on multiple host plants and in various regions. Future studies on endophytic fungi are expected to broaden, including genomics and taxonomic and ecological studies of secondary metabolites.
1. Introduction
Plants provide various ecological niches to endophytic fungi, where they colonize internal plant tissues [Citation1]. Since studies on endophytic fungi began in earnest in the 1980s, and in Korea specifically, in the 1990s, several studies have been conducted covering various aspects such as diversity, function, pharmacological substances, and so on [Citation2–4]. The Korean Peninsula has four relatively distinct seasons and is located in Northeast Asia between approximately 32°–43° north and 125°–132° east [Citation5]. The Baekdu-daegan mountain range intersects the central part of the Korean Peninsula from the north at Mt Baekdu (∼2700 m), terminating in the south at Jeju Island, which was historically geologically connected to the peninsula [Citation6]. Climatically, the Korean Peninsula is subtropical in the south (e.g., Jejudo Island) to subarctic in the north (e.g., the Gaemagowon highlands). In terms of floristic regions, the Korean Peninsula is subdivided into eight subregions, including the Kwanbuk district subregion in the northernmost part of the country [Citation7]. Forests cover approximately 63% of the Korean Peninsula and can be classified as coniferous, broad-leaved, or mixed forests [Citation8]. A total of 4606 species of vascular plants are reported: 337 species of ferns, 55 species of gymnosperms, and 4214 species of angiosperms [Citation9]. The distribution of plants according to diverse altitudes, climates, and forest types provides suitable conditions for exploring the biodiversity and ecological phenomena of endophytic fungi.
This review summarizes the results of studies on endophytic fungi in Korea from 1991 to 2021. To prepare this review, the date range was set from 1991 to 2021 in Google Scholar, DataBase Periodical Information Academic (DBPIA), Research Information Sharing Service (RISS), ScienceON, and the Korean Journal of Mycology and Mycobiology. A total of 118 papers were analyzed. Considering that a variety of endophytic fungi are found in various host plants, we identify the plants and regions where studies on endophytic fungi have been conducted and any new relationships that have been discovered. We also discuss the suggested direction of future studies on endophytic fungi and the application of new methods in Korea.
2. Studies on endophytic fungi in Korea
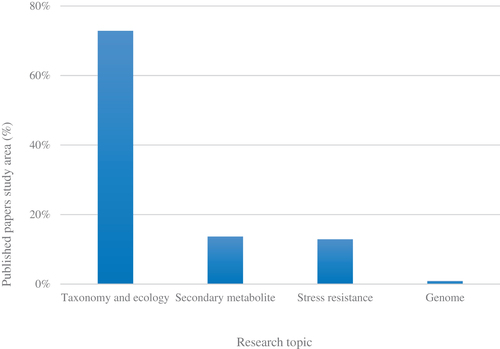
We examined 118 published papers to analyze the biodiversity of endophytic fungi in Korea over the last 30 years. The number of studies on endophytic fungi in Korea is continuously increasing. Initially, there were only one or two publications per year, but now an average of 10 papers are published yearly (). Studies on endophytic fungi in Korea are largely confined to four fields, but this is also diversifying. Most (71.9%) comprise classification and ecological studies, followed by secondary metabolite-related studies (14.8%), such as antifungal detection. This is followed by research related to stress resistance of host plants (12.5%), and finally, genome-related studies (0.8%; ). The reason for the primary focus on taxonomy and ecological studies may be owing to the nascent stage of endophytic fungi studies in Korea. Steady attempts are being made to accurately identify endophytic fungi taxonomically and to isolate and culture them from new host plants and plant tissues. Similar to studies in other fields in the natural sciences, studies on endophytic fungi are expected to significantly broaden from basic studies, such as classification and ecology, to fields comprising secondary metabolites and stress resistance as taxonomic study results accumulate.
Figure 1. The number of study papers on endophytic fungi published in Korea since the 1990s. Publication of studies related to endophytic fungi increased each year. Recently, on average, ∼10 studies on endophytic fungi have been published per year.
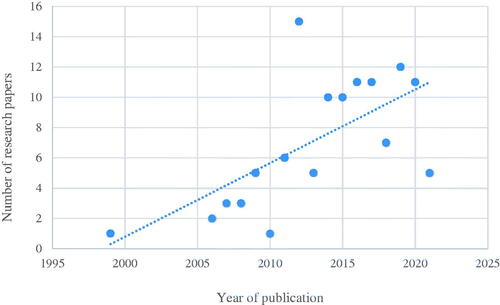
Figure 2. Study areas of published papers related to endophytic fungi in Korea by percentage. These publications were checked based on keywords and the papers were classified into four major topics: taxonomy and ecology (72.8%), secondary metabolites (13.6%), stress resistance (12.8%), and genome (0.8%).
3. Diversity of host plants
The review confirms the inclusion of 50 families, 108 genera, and 132 species of host plants in the study of endophytic fungi to date, with 3 types of mosses, 34 types of woody plants, and 95 types of herbaceous plants being confirmed as host plants. Among the woody plant hosts, 18 species of broad-leaved and 16 species of conifer trees were confirmed, whereas 22 annual and 73 perennial species were recognized among the herbaceous plants (). Additionally, most of the host plants used in the studies were native to the Korean Peninsula. However, four species—Pinus rigida Mill., Aronia sp., Larix kaempferi (Lamb.) Carriere, and Acer buergerianum Miq.—were confirmed as introduced species.
Table 1. Families and number of species of host plants included in studies of endophytic fungi in Koreaa.
The most studied host plants in Korea belong to the Poaceae family, of which 18 species were included in the study of endophytic fungi. Together with Poaceae, Pinaceae (11 species), Amaranthaceae, and Asteraceae (8 species each) constitute the top four families studied. Fewer than five species were studied within most other plant families. Among the studied host plants, ∼56% were the only species studied in a particular family. It is, therefore, necessary to diversify host plants in future studies of endophytic fungi, particularly as studies were conducted by isolating and culturing endophytic fungi from only ∼3.5% of the plant taxa distributed in the Korean Peninsula [Citation9]. Studies on endophytes have been conducted on only a very small number of host plants to date. Among them, research results of crops and conifers as host plants occupy about 47.4%. The reason is that crops such as chili pepper [Citation10] are easy to collect as host plants and research results are linked directly practical aspects such as improving productivity. In case of conifer, some researchers are intensively studying things such as Khingan fir [Citation11] which are feared to be greatly affected by global warming. There is still a large scope for taxonomic and ecological studies and potential for applied studies as well.
Among the target host plants, endophytic fungal studies have been conducted on seven species considered to be relatively vulnerable to environmental changes, including endangered species distributed in Korea [Citation12], endemic species, and climate-sensitive biological indicator species [Citation13]. Among them, two of Korea’s 459 endemic species have been studied: Abies koreana E.H. Wilson and Sedum takesimense Nakai. [Citation14]. In terms of endangered species, Cypripedium japonicum Thunb. and C. macranthos Sw., which accounted for 2.3% of the selected 88 species (11 and 77 species in endangered wildlife classes I and II, respectively) of vascular plants, were studied. Three climate-sensitive biological indicator species (7.6% of 39) have been studied: Cyrtomium falcatum (L.f.) C. Presl., Ilex crenata Thunb., and Camellia japonica L. [Citation9] ().
Figure 3. Characteristic cones (left) and dead trees (right) of Abies koreana E.H. Wilson in Yeongsil of Mt. Halla. This is an endemic and climate-sensitive biological indicator species in Korea. Seven studies have been reported on endophytic fungi related to fir trees, and 12 taxa in 1 division (Ascomycota), 3 classes, 8 orders, 11 families, and 12 genera were reported. The study results are insufficient, considering the urgency to better understand host plant-endophytic fungi associations in fir trees and other plants.

Climate change triggered by global warming has increased the average global temperature by approximately 0.85 °C, and it continues to increase [Citation15]. In addition, irregular occurrences of abnormal meteorological phenomena, such as excessive droughts, high temperatures, and intensified rainfall [Citation16], have been observed and are significantly affecting the ecosystem of the Korean Peninsula. This has a particularly negative effect on plants inhabiting the forest ecosystem. Environmental changes are known to have a significant impact on endemic or endangered species; thus, it is necessary to study ecologically endangered plant species to monitor their adaptation to environmental changes and for their conservation [Citation17].
Roots and leaves served as the primary host plant tissue source in the studies of endophytic fungi in Korea (). Barring Incertae sedis, 3 phyla, 8 classes, 32 orders, 73 families, 140 genera, and 269 taxa of endophytic fungi were confirmed in leaves. By contrast, 5 phyla, 16 classes, 44 orders, 99 families, 216 genera, and 621 taxa were confirmed in roots. Furthermore, 76 genera (35.0%) were identified only from leaves, and 82 genera (37.8%) only from roots. Fifty-nine genera (27.2%) were discovered in the tissues of both organs (). Therefore, the endophytic fungal community differed between the organs of the host plant. However, it cannot be deduced that a certain taxon is found only in specific organ tissues, considering that most fungi can spread through the air and also opportunistically infect plants via random methods, such as spores falling on the soil or leaves. Since the degree of isolation of endophytic fungi may vary depending on the time and medium, it is difficult to accurately identify the distribution of endophytic fungi at the current level. However, the approximate pattern of endophytic fungi in each tissue may be explained [Citation18]. Additionally, endophytic fungi have been isolated from bark (one taxon), flower stalks (four taxa), and fruit (three taxa). Considering that these plant parts have fewer reported results than leaves and roots, the scope of future studies should be expanded to include more diverse plant organs and tissues [Citation18,Citation19].
Figure 4. Endophytic fungi studies categorized according to host plant organ/tissue. Endophytic fungi found in Korea have been isolated mainly from leaves and roots. The top three classes of endophytic fungi isolated from leaves include the Sordariomycetes (50.1%), Dothideomycetes (25.8%), and Leotiomycetes (13.3%); those isolated from roots include the Eurotiomycetes (34.0%), Sordariomycetes (31.2%), and Dothideomycetes (21.5%). Unknown means that no plant tissue was specified in the paper analyzed.
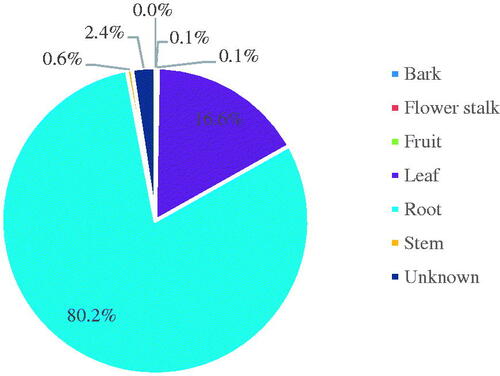
Figure 5. Endophytic fungi isolated from the leaf and root of the host plant. A total of 217 genera were identified: 82 in the leaf, 76 in the root, and 59 in both organ tissues. Genera found in the tissue of both organs likely represent opportunistic infections.

4. Distribution of endophytic fungi in Korea
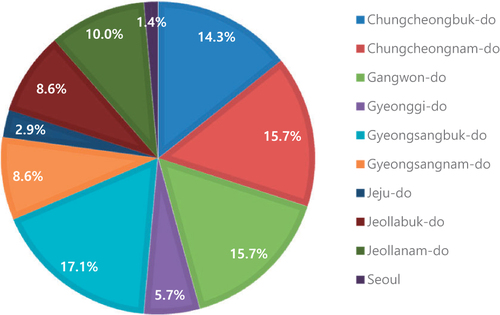
Endophytic fungi have been isolated and studied in all regions of Korea (). By province and administrative district, more than half of the studies were conducted in Chungcheongbuk-do (17.1%), Chungcheongnam-do (15.7%), Gangwon-do (15.7%), and Gyeongsakbuk-do (14.3%). Seoul was the most poorly represented (approximately 1.4%) (). Other metropolitan areas such as Daejeon, Jeollanam-do, Jeollabuk-do, and Gyeongsangnam-do were also poorly represented. This highlights the need for field surveys to confirm the biodiversity of endophytic fungi at the national level. Host plants were collected from forests (34.1%), arable land (17.9%), the coast (21.1%), wetlands (8.9%), cities (4.9%), and bare ground (4.1%). This is because the study of endophytic fungi is based on the host plant; therefore, site selections in most studies were based on the health of the host plants or the presence of suitable host plant habitat.
Figure 6. Study sites (black dots) of endophytic fungi reported in articles published in Korea from 1990 to 2020.
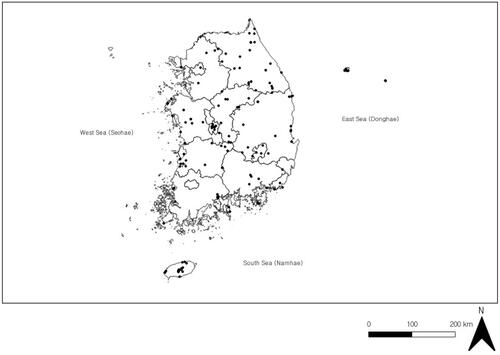
Figure 7. Regional distribution of endophytic fungi studies conducted in Korea from 1990 to 2020. Gyeongsangbuk-do (17.1%), Chungcheongnam-do (15.7%), Gangwon-do (15.7%), and Chungcheongbuk-do (14.3%) together represent more than 50% of the total study areas. Only 1.4% of the studies were performed in Seoul.
From the point of view of ecosystem conservation, studies on endophytic fungi are increasingly important in restoring the health of urban ecosystems. Climate change is altering the vegetation of the Korean Peninsula, and it is necessary to create ecosystems suitable for cities by introducing plant species that can adapt to climate change in the long term. Furthermore, the expansion of green space is inevitable for reducing carbon generated in cities and ecologically improving the environment. Thus, in addition to planting more trees, we need studies on the ecological circulation between plants and functional endophytic fungi. For example, inoculating plants with functional endophytic fungi that can resist urban pollutants and using plant species with high carbon reduction and oxygen generation increases the likelihood of the plants surviving and resisting urban environmental stress.
Studies on endophytic fungi in Korea have been relatively limited in scope, having been conducted mostly in forests based on the presence of host plants. More challenging goals need to be set, considering that the field is now out of the nascent stage. In other words, it is necessary to conduct a nationwide study on the interaction of endophytic fungi with host plants by approaching it from a floristic regional and geographical point of view that can affect the distribution of host plants.
5. Diversity of endophytic fungi in Korea
This review confirmed 5 phyla, 16 classes, 49 orders, 135 families, 305 genera, and 855 taxa among 132 host plants, barring Incertae sedis. At the phylum level, Ascomycota accounted for 93.2%, followed by Basidiomycota (3.6%) and Glomeromycota, Mucoromycota, and Oomycota (2.6%). Sordariomycetes (36.9%), Eurotiomycetes (30.3%), and Dothideomycetes (24.6%) made up 91.8% of the Ascomycota, while Agaricomycetes accounted for 79.6% of the Basidiomycota. Ascomycota comprised 93.5% and Basidiomycota 3.2% of the endophytic fungi isolated from the root, and 93.0% and 4.3%, respectively, from the leaves.
The 2021 National Species List of Korea contained 4683 species of fungi, of which 2020 species (43.1%) are ascomycetes and 2311 species (49.3%) are basidiomycetes [Citation9]. Although these fungi are found in similar proportions in nature in the Korean Peninsula, this is not the case among their endophytic counterparts in plants. The reasons for the asymmetrical dominance of ascomycetes among endophytic fungi cannot be clearly explained. This pattern was consistently observed in most of the studies we considered and was present in both the root and leaf tissues. Mycorrhizal fungi, which are distributed in the root and form a symbiotic relationship with it, belong to the basidiomycetes. Therefore, the overwhelming dominance of ascomycetes in both the root and leaf tissue is unexpected. The dominance of ascomycetes is believed to have a significant influence on plant habitat preference. However, a more detailed explanation for the observed dominance may be required.
Studies on endophytic fungi in Korea have been conducted using primarily isolation and culture methods, which can be described as follows: (1) The tissue of the host plant is sterilized, and the pure fungal strain is isolated using water, (2) the isolated strain is cultured on a slide, and its microscopic shape is observed, (3) DNA is extracted from the isolated strain, (4) the corresponding nucleotide sequence is amplified using fungal-specific primers, and (5) species candidates are selected through phylogenetic analysis and the similarity between nucleotide sequences based on nucleotide sequence databases such as NCBI. For molecular biological identification, molecular markers targeting a specific fungal barcode region are used, such as the internal transcribed spacer (ITS) 1 and ITS2 region that includes the 5.8S ribosomal rDNA. For the molecular identification of endophytic fungi, β-tubulin, a region of the translation elongation factor (TEF), and a region of the ribosomal large subunit (LSU), including domains 1 and 2 of the 28S rDNA region, are used. The choice of molecular marker to be used requires careful consideration, and molecular identification of fungi in Korea is conducted as it is globally. Initially, the ITS barcode region was used for molecular identification, followed by the TEF1α region as a cross-assay [Citation20,Citation21]. In most Korean endophytic fungi studies, the ITS region was used as a molecular marker, accounting for approximately 93.3% of all studies. The LSU marker was used in 3.8% of the studies, β-tubulin in 1.3%, and TEF in 0.7%. Molecular markers such as glyceraldehyde 3-phosphate dehydrogenase (GAPDH) and calmodulin (CaM) were rarely used (0.8%). Although the use of the ITS region alone is not sufficient, it is possible to compare the nucleotide sequence by extending sequencing of the rDNA to the LSU region. The sequences in the TEF region is different from those in the ITS region and, therefore, a new gene could be applied to the same taxon. Furthermore, it has been difficult to identify only the TEF region to date because most of the nucleotide sequences in the database are primarily of the ITS region. The TEF region also has a different scale distance than the rDNA region, so caution should be applied in its interpretation. The ITS marker region used as a DNA barcode comprises approximately 500–800 bp, and while it is somewhat conservative, it has the advantages of accumulating mutations and having a well-established nucleotide sequence database allowing comparisons among related species [Citation22]. However, considering it is impossible to identify the entire fungal taxon using only one barcode gene, there is a constant demand for alternatives.
Experimental methods affect the resulting observed endophytic fungal biodiversity [Citation13]. The culturing of endophytic fungi may vary depending not only on the host plant used for isolation but also on the experimental conditions [Citation18]. When isolating endophytic fungi, the growth rate of each strain varies depending on the medium and temperature. Slow-growing endophytic fungi strains are often lost to fast-growing strains. The number of isolated endophytic fungi strains may also differ depending on the size of the host plant tissue [Citation12] placed on a medium. Experimental methods also affect the observed endophytic fungal biodiversity [Citation13]. Therefore, the biodiversity of endophytic fungi should be assessed keeping in mind the potential impact of isolation and culture methods. Next-generation sequencing (NGS) is preferred for whole endophytic fungi present in a host plant. The method used in NGS cuts the genome into countless fragments, which are then amplified in large quantities. An amplified genome is useful for confirming the existence of microorganisms that cannot be detected by isolation–culture methods and decrypted using bioinformatics. Performing metagenomics using the ITS region can identify the fungal community in a sample collected from a specific environment rather than an individual fungal species. This approach can solve problems inherent in the isolation–culture method and allow for the discovery of new fungi and the distribution and density of endophytic fungi in plants. Considering the advantages of cost, time, and ability to process several samples simultaneously, NGS is an important means to study the endophytic community structure. It is also an alternative approach that can compensate for inaccurate information about endophytic fungi strains that have not been isolated from host plants [Citation14]. Despite these advantages, NGS has some limitations. For example, since the length of the nucleotide sequence used for NGS is approximately 350–500 bp shorter than that of the ITS barcode region, there is a limitation in the taxonomic resolution and the possibility that the actual strain cannot be studied. Therefore, the isolation–culture method and NGS should be performed simultaneously to produce complementary results.
6. Taxonomy of endophytic fungi
It is estimated that there are 1,500,000 to 10,000,000 species of fungi globally [Citation23], including 1,000,000 to 1,250,000 species of endophytic fungi [Citation24]. Most endophytic fungi are of the division Ascomycota, with some Basidiomycota, Mucoromycota, and others [Citation25]. The fungal classification has undergone considerable changes based on molecular phylogeny, and the kingdom Fungi is now composed of 5 subkingdoms and 16 divisions. Most endophytic fungi belong to Ascomycota and Basidiomycota in the subkingdom Dikarya [Citation26].
Using molecular markers yields effective results in the identifying microbial species that are difficult to identify using key physical characteristics owing to very few morphological differences between species. Therefore, using molecular markers supplements morphological species identification, demonstrating effective results even in animals and plants [Citation27], and is essential in evaluating endophytic fungi with great biodiversity.
At this point, a reverse taxonomic approach is required. It is possible to fix the limit of species using available genetic data, such as taxa distributed in various habitats, not cultured or relatively difficult to classify based solely on morphological characteristics, to focus on microscopic morphological analyses. Establishing this approach will enable efficient discovery of fungal biodiversity. However, despite these advantages limiting the species to be analyzed, the relatively low accuracy of sequencing and the construction of databases are often considered obstacles. In particular, the irregularity of interspecies and intraspecies variation poses difficulties in species analysis, creating the requirement of more diverse genetic markers, consequently increased time and cost. These factors explain the increase in studies at the genomic level compared to molecular markers. Thus, it may be necessary to evaluate biodiversity via the development of standard procedures for reverse taxonomy.
Endophytic fungi are closely related to their host plant. Based on this, they are divided into two groups [Citation28]. Group I comprises one cluster (Class 1) and includes endophytic fungi belonging to the family Clavicipitaceae. Compared to other clusters, in Class 1, the range of hosts is limited, and most are specific herbaceous species such as Lolium spp. Group II comprises endophytic fungi other than the Clavicipitaceae family and can be divided into three clusters (Classes 2–4). Most Group II fungi are classified into Dikarya Ascomycota and Basidiomycota and are associated with a wide range of hosts. Class 3 comprises fungi found in the aboveground part of the plant. Class 4 comprises pseudomycorrhizal fungi symbiotic with the underground roots of angiosperms, commonly referred to as dark septate endophytic fungi. Of the 855 taxa studied in Korea, none fall within Class 1, and one falls within Class 4. All the other studied species in Korea fall into Classes 2 and 3. According to previous studies, variations in infected tissue, infection patterns, and diversity in the plant and modes of transmission are important criteria for classification into Classes 2 and 3. We propose that these criteria distinguishing Class 2 and Class 3 represent a kind of ecological scale, and it is questionable whether they can be realistically applied to studies. The ecological scale that divides Class 2 and Class 3 appears to be related to host plant and fungi preferences; fungi belonging to Class 2 are friendlier to host plants than fungi in Class 3. Class 2 has a narrow range of host plants and demonstrates more diverse infected tissues and broader infection patterns than Class 3 because the tissue in the host plant may be infected in several ways, from the root to the aboveground parts. Furthermore, the fungal diversity is inevitably lowered because infection within plants can occur widely. Finally, the transfer method of Class 2 fungi is also friendlier to the host plant; the fungi can penetrate both seeds and tissues, with the possibility of vertical and horizontal transmission [Citation28]. Therefore, rather than clarifying the ecological scale suggested in previous studies, the preference of host plants and endophytic fungi should be considered as the key study topic, and study methods that can distinguish those preferences should be applied.
7. Ecology of endophytic fungi
Endophytic fungi are found in most plant species, including mosses, Cephalanthera longibracteata Blume, and Thuja koraiensis Nakai [Citation29–31]. Those that coexist with plants have a positive effect on host plants, aiding them in adapting to their environment and ultimately affecting the distribution of plants. Endophytic fungi are directly affected, just as plants are affected, by the distribution and dispersal of seeds under the influence of endophytic fungi. The biodiversity of plants in tropical regions is higher than that in temperate regions since the distribution of plants is influenced by temperature and humidity [Citation32].
Korea experiences seasonal changes and, consequently, variability in plant growth. Plant growth is observed in spring and summer, but in autumn and winter, leaves are abscised, seeds are produced, and the plants enter a potentially dormant state. Endophytic fungi are inevitably affected by such changes in the state of the host plant, and it is vital to better understand this phenomenon. Despite this, only a few studies have been recorded in Korea addressing this. However, some studies conducted on a quarterly basis confirmed that in conifers, the number of fungal strains and species increased as the season progressed from spring to winter [Citation33]. Studies on how seasonal changes affect the biodiversity of endophytic fungi are required. From such studies, it will be possible to identify the occurrence of endophytic fungi according to the season. Finally, the dynamic state of endophytic fungal communities can be elucidated by tracking their seasonal appearance and extinction patterns.
It is necessary to know which endophytic fungi are present in plants and to what extent to study the interactions and ecological functions of endophytic fungi and host plants. To this end, the structure of non-cultured endophytic fungi should be observed to supplement the outcomes of the isolation–culture method. For this, NGS and digital PCR are required. These experimental methods can more closely reflect reality. Molecular ecological monitoring of non-cultured endophytic fungi is possible through NGS, and absolute quantification of endophytic fungi at the host plant or tissue level is possible through digital PCR. If the components of the endophytic fungal community and their abundance are determined, it may be possible to realistically interpret what environmental and ecological functions the endophytic fungi perform within the host plant.
Finally, studies of the mitochondrial genome of fungi have recently developed as an important topic. Considering the relatively weak ability of mitochondria to repair their damaged DNA, the frequency of mutations in mitochondrial DNA is approximately 10–20 times higher than that in nuclear DNA. Artificial mutations in the color, size, and wrinkle formation of a fungal colony were reported due to point mutations in the tRNA of Candida albicans (C.-P. Robin) Berkhout [Citation34]. Thus, the mitochondrial genome could be utilized as an indicator of heterogeneity. The distribution and geographic types of host plants are most often identified using chloroplast or mitochondrial genome sequencing. Based on the infection patterns of endophytic fungi, it is believed that there are instances where endophytic fungi and the host plant form a specific relationship according to the host plant’s preference for the endophytic fungi. Therefore, if the mitochondrial genome of the host plant and endophytic fungi are studied simultaneously in the context of the distribution area of host plants, it may be possible to gather insight into the biogeographical processes through which endophytic fungi and host plants have been dispersed.
8. Conclusion
A total of 5 phyla, 16 classes, 49 orders, 135 families, 305 genera, and 855 taxa of endophytic fungi were confirmed in 50 families, 108 genera, and 132 species of host plants through the review of studies on endophytic fungi conducted in Korea over the last 30 years. Among the fungi, Ascomycetes accounted for approximately 93% of the total number of species. Although 4596 species of vascular plants are distributed in the Korean Peninsula, studies of endophytic fungi have been conducted on only 3% of these potential host species. Further taxonomic and ecological studies are required to expand the understanding of endophytic fungi.
In particular, it is necessary to extend the biodiversity study of endophytes intensively investigated in crops and conifers to other host plants. In addition, the study area for endophytes was also targeted at study sites focusing on specific landscapes, but now it should be expanded to urban areas with perspective of host plants distribution and floristic region. Because studies on endophytic fungi in Korea mainly considers host plants rather than geographical, genetic, or environmental factors, study topics should also be diversified. To do this, it is necessary to increase the efficiency of species identification through a reverse taxonomy approach. Endophytic fungi are greatly influenced by the distribution of host plants, and the host plants are influenced by seasonal changes in Korea. We do not know how endophytic fungi existing in Korea are changing within host plants and how much they exist in it. Therefore, basic ecological monitoring of host plants and endophytic fungi and molecular biological techniques are required, and mitochondrial genomic study related to mutation and dispersal mechanisms of endophytic fungi are also required. Studies should also focus on their function of endophytic fungi in the ecosystem. In addition to the ecological aspect, several developments are expected to make human life more prosperous through the study of beneficial secondary metabolites or functional genes in endophytic fungi. This is the point where more energy needs to be invested in endophytic fungal research for our sustainable development.
Disclosure statement
No potential conflict of interest was reported by the author(s).
References
- Petrini O. Taxonomy of endophytic fungi of aerial plant tissues. In: Fokkema NJ, Huevel JVD, editors. Microbiology of the phyllosphere. Cambridge: Cambridge University Press; 1986. p. 175–187.
- Kumaran RS, Hur BK. Screening of species of the endophytic fungus Phomopsis for the production of the anticancer drug taxol. Biotechnol Appl Biochem. 2009;54(1):21–30.
- Rubini MR, Silva-Ribeiro RT, Pomella AWV, et al. Diversity of endophytic fungal community of cacao (Theobroma cacao L.) and biological control of Crinipellis perniciosa, causal agent of witches’ broom disease. Int J Biol Sci. 2005;1(1):24–33.
- Ganley RJ, Brunsfeld SJ, Newcombe G. A community of unknown, endophytic fungi in Western white pine. Proc Natl Acad Sci USA. 2004;101(27):10107–10112.
- Kong WS. Biogeography of native Korean Pinaceae. J Korean Geog Soc. 2006;41:73–93.
- Im HT. Plant geographical study for the plant of Cheju. Korean J Pl Taxon. 1992;22(3):219–234.
- Lee WC, Yim YJ. Plant geography. Chuncheon: Kangwon National University Press; 2002.
- Kong WS. Species composition and distribution of native Korean conifers. J Korean Geog Soc. 2004;39:528–543.
- National Institue of Biological Resources. National list of species of Korea [Internet]. [cited 2022 Jun 10]. Available from: https://species.nibr.go.kr.
- Paul NC, Deng JX, Sang H-K, et al. Distribution and antifungal activity of endophytic fungi in different growth stages of chili pepper (Capsicum annuum L.) in Korea. Plant Pathol J. 2012;28(1):10–19.
- Eo JK, Park E. One unrecorded endophytic fungi from sub-alpine conifer, Rhizosphaera pini. Kor J Mycol. 2019;47:121–124.
- National Institue of Biological Resources. Endangered wildlife in Korea [Internet]. [cited 2022 Jun 10]. Available from: https://species.nibr.go.kr
- National Institue of Biological Resources. 100 Climate-sensitive biological indicator species [Internet]. [cited 2022 Jun 10]. Available from: https://species.nibr.go.kr
- Nam GH, Kwak MH, Lim CK, et al. Endemic species of Korea. Revised ed. National Institute of Biological Resources (NIBR). Incheon: NIBR; 2018.
- Korea Meteorological Administration. Annual report 2014 [Internet]. [cited 2022 Jun 10]. Available from: https://www.kma.go.k
- Lee S, Rhyu S. Impacts of global temperature rise on the change of snowfall in Korea. J Korean Geograph Soc. 2003;38:463–477.
- Koo KA, Park WK, Kong WS. Dendrochronological analysis of Abies koreana E. H. Wilson at Mt. Halla, Korea: Effects of climate change on the growths. Korean J Ecol. 2001;24:281–288.
- Kim CK, Eo JK, Eom AH. Diversity of foliar endophytic fungi isolated from Lindera obtusiloba in Korea. Kor J Mycol. 2012;40(3):136–140.
- Carroll F, Muller E, Sutton B. Preliminary studies on the incidence of needle endophytes in some European conifers. Sydowia. 1977;29:103.
- Prakash PY, Irinyi L, Halliday C, et al. Online databases for taxonomy and identification of pathogenic fungi and proposal for a cloud-based dynamic data network platform. J Clin Microbiol. 2017;55(4):1011–1024.
- Stielow JB, Levesque CA, Seifert KA, et al. One fungus, which genes? Development and assessment of universal primers for potential secondary fungal DNA barcodes. Persoonia. 2015;35:242–263.
- Medicine NLo. National Center for Biotechnology Information [Internet] [cited 2022 Jul 8]. Available from: https://www.ncbi.nlm.nih.gov/.
- Hawksworth DL. The fungal dimension of biodiversity: magnitude, significance, and conservation. Mycol Res. 1991;95(6):641–655.
- Petrini O, Andrews J, Hirano S. Microbial ecology of leaves. New York: Spring-Verlag; 1991. p. 179–197.
- Sinclair J, Cerkauskas R. Endophytic fungi in grasses and woody plants systematics, ecology and evolution. Minnesota: APS Press; 1996.
- Tedersoo L, Sánchez-Ramírez S, Kõljalg U, et al. High-level classification of the fungi and a tool for evolutionary ecological analyses. Fungal Divers. 2018;90(1):135–159.
- Göker M, García-Blázquez G, Voglmayr H, et al. Molecular taxonomy of phytopathogenic fungi: a case study in Peronospora. PLOS One. 2009;4(7):e6319.
- Rodriguez R, White J, Jr Arnold A, et al. Fungal endophytes: diversity and functional roles. New Phytol. 2009;182(2):314–330.
- Choi HS, Park H, Eo JK, et al. Unrecorded endophytic fungi isolated from Mnium heterophyllum and Hypnum plumaeforme in Korea: Biscogniauxia petrensis and Cercophora thailandica. Kor J Mycol. 2020;48:55–61.
- Eo JK, Lee BH, Eom AH. Diversity of endophytes isolated from Thuja koraiensis Nakai in the Korean peninsula. Kor J Mycol. 2016;44:113–117.
- Lee BH, Eom AH. Diversity of endophytic fungi associated with roots of Calanthe discolor and Cephalanthera longibracteata in Korea. Kor J Mycol. 2018;46:427–435.
- Cannon P, Hawksworth D. The diversity of fungi associated with vascular plants: the known, the unknown and the need to bridge the knowledge gap. Adv Plant Pathol. 1995;11:277–302.
- Kim CK, Eo JK, Eom AH. Diversity and seasonal variation of endophytic fungi isolated from three conifers in Mt. Taehwa, Korea. Mycobiology. 2013;41(2):82–85.
- Elson SL, Noble SM, Solis NV, et al. An RNA transport system in Candida albicans regulates hyphal morphology and invasive growth. PLOS Genet. 2009;5(9):e1000664.