Résumé
L’étude de l’effet du NaCl a été suivie sur trois clones de peuplier blanc (MA-104, MA-195 et O.Gherib). Certains paramètres micromorphologiques foliaires ont été mesurés. Les résultats suggèrent que les différences inter et intraspécifiques relevées sont principalement liées à la diversité des conditions écogéographiques des lieux d’origine. En effet, le clone MA-104 a développé la plus forte densité de trichomes et de stomates. Il a montré une plasticité des stomates mise en évidence par une réduction de la longueur et de l’ouverture des ostioles. Le clone MA-195 a montré une moindre augmentation du nombre de stomates par comparaison à MA-104, mais l’ouverture de ses ostioles a été réduite. Le clone O.Gherib a révélé la plus faible densité stomatique et l’absence de réduction de l’ouverture des ostioles correspondant à une faible tolérance à la salinité.
Abstract
We study the salt effect on three clones of white poplar (MA-104, MA-195 and O.Gherib). Leaf micromorphological parameters were measured. This study has shown a high degree of variation of these characters mainly related to geographical origin. It was observed that the clone MA-104 has developed the highest density of trichomes and stomata. It showed a plasticity of it’s stomata as evidenced by a reduction in their length and the opening of the mudboils. The clone MA-195 showed a smaller increase in the number of stomata compared to MA-104, but the opening of its mudboils is greatly reduced. The clone O.Gherib developed the lower stomatal density and the absence of reduction of the opening of mudboils related to low tolerance to salinity.
Introduction
La salinité de l’eau et du sol compte parmi les facteurs majeurs limitant la croissance chez les non-halophytes. La salinité peut entraîner une inhibition de la croissance et une modification de l’anatomie et de la morphologie de la plante (Pallardy & Kozlowski, Citation1980). De fortes concentrations en sel peuvent provoquer, chez des plantes, une sécheresse physiologique suite à une diminution du potentiel osmotique du sol (Rengel, Citation1992). Ces plantes développent souvent divers mécanismes d’ordre morphologique et physiologique. Les adaptations morphologiques sont souvent associées à une réduction de la partie aérienne au travers d’une surface foliaire réduite et d’une modification des caractères micromorphologiques des feuilles. En effet, les paramètres stomatiques figurent parmi les meilleurs indicateurs sur l’adaptation d’une espèce à son milieu (Ait-Said et al., Citation2005). Comme exemple, sous l’effet d’un stress dû à la pollution, des changements se sont opérés dans la longueur, la largeur et la densité des stomates des feuilles d’arganier (Alilou et al., Citation2006). Par ailleurs, l’abondance et la petitesse des stomates, en condition limitante en eau, sont considérées comme un critère d’adaptation à la sécheresse (Albouchi et al., Citation1997). La présence de poils ou trichomes assure un rôle important dans la réduction de la transpiration. En effet, un tel caractère a pour effet de réduire la perte d’eau qui n’est pas liée à l’augmentation de la couche limite crée par les poils, mais à une réduction de l’interception de la lumière induisant une baisse de la température de la feuille (Ehleringer, Citation1980).
La forêt tunisienne est un écosystème fragilisé par les conditions climatiques défavorables et la pression permanente des usagers de la forêt et de leur cheptel. Elle est confrontée aux changements climatiques révélés principalement par l’augmentation des températures, un bouleversement dans le régime des précipitations et la fréquence d’évènements extrêmes tels la sécheresse et les vagues de chaleur. Dans le but de faire face à ces contraintes, des efforts considérables de reboisement ont été déployés dans l’objectif d’atteindre un taux de couverture forestière de 15%. Le recours au boisement des zones salines à l’aide d’essences ripicoles à croissance rapide telles que le peuplier blanc (Populus alba L.), pourrait contribuer à la mise en valeur de ces zones et à réduire le déficit en bois. Cette espèce présente des réponses intéressantes vis-à-vis de la salinité comparée à d’autres peupliers (Sixto et al., Citation2005). Cependant, diverses études ont montré que ce peuplier présente une variabilité intraspécifique dans sa sensibilité à la salinité (Beritognolo et al., Citation2007). Les travaux de Abassi et al. (Citation2004) ont conforté cette idée sur des clones de peuplier existants en Tunisie. Ils ont été suivis par une étude plus fine afin de déterminer les mécanismes impliqués dans la tolérance à la salinité de trois clones présélectionnés de peuplier blanc. Parmi les mécanismes développés figure une différence dans l’expression des modifications anatomiques foliaires exprimée par ces clones sous l’effet du stress salin (Abassi et al., Citation2009). La recherche de modifications morphologiques et micromorphologiques foliaires développées par le peuplier blanc face à ce stress constitue l’objectif du présent travail.
Matériel et méthodes
La présente étude s’est déroulée à la pépinière de l’institut national de recherches en génie rural, eaux et forêts de Tunis. Elle a porté sur des cépées âgées d’un an de trois clones de peuplier blanc, Populus alba L. var. hichkeliana (Dode) à tolérance croissante à NaCl: O.Gherib (sensible), MA-195 (moyennement tolérant) et MA-104 (tolérant) (Abassi et al., Citation2004). Les clones MA-104 et MA-195 importés du Maroc montrent une domestication dans les conditions du nord-ouest du pays. Le clone O.Gherib pousse sur les berges de l’oued Gherib (ancienne plantation située dans la même région). Ces cépées sont plantées dans des pots en plastique de 50 cm de diamètre et 60 cm de profondeur, contenant un substrat inerte, le sable grossier. Ces pots portent des drains évacuant l’eau en excès. Pour chaque clone, deux traitements sont appliqués: un témoin (T) irrigué à l’eau de robinet sans ajout de NaCl et 6 g/l NaCl. Cette dose de NaCl a été ainsi fixée car en moyenne la charge en NaCl des eaux de surface et d’irrigation en Tunisie surtout en période estivale avoisine les 6g/l (Hachicha et al., Citation1995). Le substrat est enrichi par une solution nutritive de Hoagland (Hoagland & Arnon, Citation1950) à moitié concentrée, une fois par semaine. Le maintien de la dose 6 g/l de NaCl a été assuré au niveau du substrat grâce à un suivi régulier de la conductivité électrique de l’eau de drainage à l’aide d’un conductimètre de type Cellox 325 WTW. Le dispositif expérimental adopté est du type Split-Plot, avec quatre blocs, où le clone constitue l’unité principale alors que la dose de NaCl représente l’unité secondaire. Chaque bloc comporte trois parcelles élémentaires contenant 18 plants du même clone répartis aléatoirement en deux lots de 9 plants soumis respectivement aux traitements (0 g/l, et 6 g/l NaCl), soit au total 216 plants. La densité, la longueur et la largeur des stomates ainsi que la densité des trichomes et leur diamètre ont été déterminés à l’aide de la méthode de l’empreinte épidermique (Jones, Citation1983). Les empreintes ont été obtenues par l’application d’une couche de vernis à ongle transparent à la face inférieure d’une feuille mature de peuplier, dans sa partie médiane. Une fois séchée, cette couche de vernis est enlevée à l’aide d’un ruban adhésif et placée sur une lame porte-objet. Les observations ont été réalisées grâce à une caméra vidéo type Hitachi KP-D 40 Color Digtal, montée à un microscope de type Leitz Dialux 22E (agrandissement 250 fois) et reliée à un ordinateur. L’image affichée sur l’écran de l’ordinateur est analysée par un logiciel Win Dias. Le calibrage de ce système se fait au préalable par une lame graduée au 1/10. Les mesures ont été réalisées sur des feuilles matures situées au même rang, sur un rameau bien éclairé. Chaque donnée est la moyenne de 48 observations (4 observations par feuille et 3 feuilles par plant x 4 répétitions / clone et par traitement).
La densité des stomates (Ds) a été déterminée par comptage de leur nombre par mm² de surface foliaire. La dimension des stomates est déterminée par la mesure en μm de la longueur (a) et de la largeur (b) de l’ouverture de l’ostiole. Ces paramètres ont servi à la détermination de la surface relative des pores stomatiques SR= (a x b)/4 x Ds x π x 10-4 en% (Meinder & Mansfield, Citation1968).
La densité des trichomes (DT) a été déterminée à partir du comptage du nombre de leurs empreintes par mm2 d’épiderme alors que leur diamètre est déterminé par la mesure en μm de chaque empreinte. La surface ou couverture des trichomes (ST) a été calculée par la formule suivante: ST (mm2) = (D/2)² x π x DT en mm², où D représente le diamètre d’un trichome.
La surface foliaire a été mesurée sur quatre plants par traitement à l’aide d’un planimètre (Patton electronic Pty Ltd, Adelaide, SA).
Les mesures ont fait l’objet d’une analyse de la variance à deux facteurs par le test F de Fisher pour vérifier l’hypothèse d’égalité des moyennes au seuil de risque de 5%, au logiciel STATISTICA (version-V) (Stat Soft France 1998).
Résultats
Structure des feuilles témoins
L’examen micromorphologique n’a révélé la présence de stomates et de poils que sur la face inférieure des feuilles (Fig. ). Le nombre des stomates a été significativement différent entre clones (Tableau ). Le clone MA-104 a présenté la densité en stomates la plus élevée (295 st/mm2), suivi par MA-195 (251 st/mm2) puis par O.Gherib (216 st/mm2) (Fig. ). Ces stomates ont été d’une longueur similaire de l’ordre de 19 μm, alors qu’une différence s’est exprimée pour la largeur (Tableau ). Les clones MA-104 et MA-195, ont montré des stomates à largeur statistiquement similaire de l’ordre de 9 μm, contre 7 μm pour O.Gherib. Une différence entre clones s’est exprimée pour le nombre de trichomes où MA-104 s’est distingué par la plus forte densité (259 tr/mm2) suivi par O.Gherib (190 tr/mm2) et MA-195 (157 tr/mm2). Cependant, ces clones ont montré des trichomes à diamètre statistiquement similaire (Tableau ).
Fig. 1 Epidermal imprint of a witness leaf of white poplar (G x 250). Fig. 1. Empreinte épidermique d’une feuille témoins de peuplier blanc (G x 250).
Tableau 1 Analyse de la variance relative à l’effet de NaCl sur la densité stomatique (Ds), la longueur des stomates (a), la largeur des stomates (b) et la surface relative des pores stomatiques (SR) des clones de peuplier blanc (MA-104, MA-195 et O.Gherib). Table 1. Summary of analysis of variance on the effect of NaCl on stomatal density (Ds), the stomatal length (a), the stomatal width (b) and the relative area of stomatal pores (SR) of white poplar clones (MA-104, MA-195 and O.Gherib).
Fig. 2 Effect of NaCl on the stomatal density of white poplar clones (MA-104, MA-195 and O. Gherib). The data are means values of forty-eight measurements. Fig. 2. Effet de NaCl sur la densité des stomates des clones de peuplier blanc (MA-104, MA-195 et O.Gherib). Les valeurs représentent les moyennes de quarante-huit mesures individuelles.
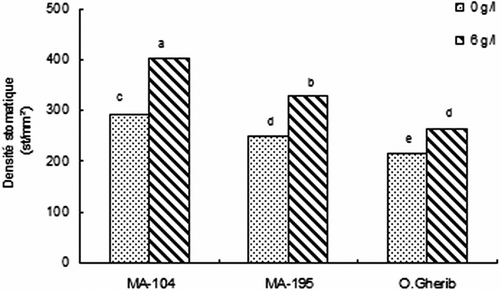
Tableau 2 Analyse de la variance relative à l’effet de NaCl sur la densité des trichomes (DT), le diamètre des trichomes (D) et la couverture foliaire en trichomes (ST) des clones de peuplier blanc (MA-104, MA-195 et O.Gherib). Table 2. Summary of analysis of variance on the effect of NaCl on the trichomes density (DT), the trichomes diameter (D) and the trichomes foliar cover (ST) of white poplar clones (MA-104, MA-195 and O.Gherib).
Influence de NaCl sur la densité et les dimensions des stomates
La salinité a induit des modifications significatives dans la densité et les dimensions des stomates du peuplier blanc.
La comparaison par rapport aux témoins montre que Ds a augmenté chez MA-104, MA-195 et O.Gherib de 37, 31 et 22%, respectivement (Fig. ). La comparaison entre clones indique que MA-104 a présenté la densité stomatique la plus élevée, dépassant respectivement celle de MA-195 et O.Gherib de 22 et de 53%. Le clone MA-195 a développé une densité stomatique dépassant celle de O.Gherib de 25%.
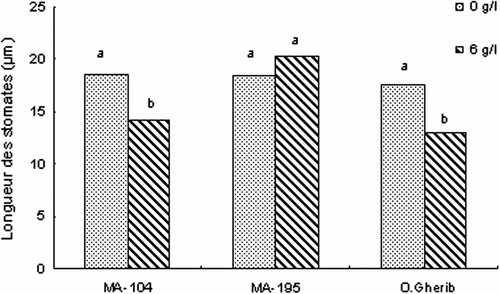
D’autre part, la salinité a induit une réduction significative de la longueur des stomates des clones MA-104 et O.Gherib de l’ordre de 25% du témoin, alors que le clone MA-195 a maintenu des longueurs stomatiques statistiquement similaires (Fig. ).
Fig. 3 -Effect of NaCl on the stomatal length of white poplar clones (MA-104, MA-195 and O. Gherib). Values represent the mean of forty-eight individual measurements. Fig. 3. Effet de NaCl sur la longueur des stomates des clones de peuplier blanc (MA-104, MA-195 et O.Gherib). Les valeurs représentent les moyennes de quarante-huit mesures individuelles.
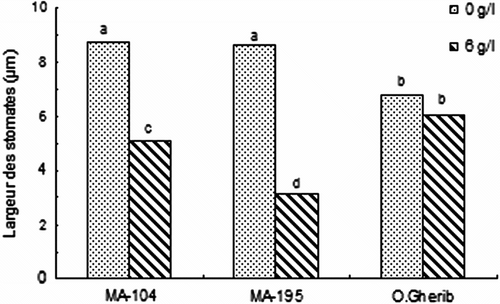
Une différence significative entre clones a concerné la largeur de l’ouverture stomatique (Tableau ). Les clones MA-104 et MA-195 ont subi une réduction significative de ce paramètre, alors que O.Gherib a maintenu des largeurs statistiquement similaires à celles des témoins (Fig. ). Par ailleurs, le clone MA-195 s’est distingué par la plus faible largeur des stomates (- 67% du témoin), suivi en deuxième position par MA-104 (- 44% du témoin).
Fig. 4 -Effect of NaCl on the stomatal width of white poplar clones (MA-104, MA-195 and O. Gherib). Values represent the mean of forty-eight individual measurements. Fig. 4. Effet de NaCl sur la largeur des stomates des clones de peuplier blanc (MA-104, MA-195 et O.Gherib). Les valeurs représentent les moyennes de quarante-huit mesures individuelles.
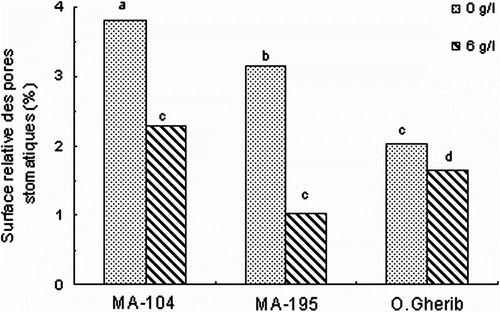
La salinité a réduit significativement la surface génératrice des pertes transpiratoires (évaluée à partir du calcul de la surface relative des pores stomatiques, ou ouverture des ostioles) (Tableau ), mais de manière variable selon le clone. Ainsi, MA-195 s’est distingué par la plus faible ouverture en ostioles avec une réduction par rapport aux témoins de l’ordre de 67%, suivi par MA-104 (40%) (Fig. ). Le clone O.Gherib a subi la moindre réduction en ouverture des ostioles (-16% du témoin). Par ailleurs, la comparaison entre clones a montré que MA-195 a développé la plus faible surface relative des pores stomatiques (1,03%), suivi par O.Gherib (1,8%) puis par MA-104 (2,27%).
Fig. 5 -Effect of NaCl on the relative area of stomatal pores of white poplar clones (MA-104, MA-195 and O. Gherib). Values represent the mean of forty-eight individual measurements. Fig. 5. Effet de NaCl sur la surface relative des pores stomatiques des clones de peuplier blanc (MA-104, MA-195 et O.Gherib). Les valeurs représentent les moyennes de quarante-huit mesures individuelles.
Influence de NaCl sur la densité et le diamètre des trichomes
Le sel a induit une augmentation significative de la densité des trichomes des feuilles de peuplier blanc (Tableau ).
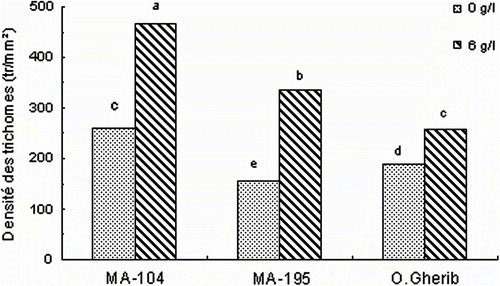
Par comparaison aux témoins, le clone MA-195 a montré la plus forte augmentation en densité des trichomes de l’ordre de 144%, le clone MA-104 (80%) alors que O.Gherib a montré la plus faible augmentation (36%) (Fig. ). La comparaison entre clones indique que MA-104 développe la plus importante densité des trichomes, dépassant respectivement celle de MA-195 et de O.Gherib de 39 et 81%. Il a été suivi en deuxième position par le clone MA-195 qui a montré une augmentation de l’ordre de 31% par comparaison au clone O.Gherib.
Fig. 6 -Effect of NaCl on the trichomes density of white poplar clones (MA-104, MA-195 and O. Gherib). Values represent the mean of forty-eight individual measurements. Fig. 6. Effet de NaCl sur la densité des trichomes des clones de peuplier blanc (MA-104, MA-195 et O.Gherib). Les valeurs représentent les moyennes de quarante-huit mesures individuelles.
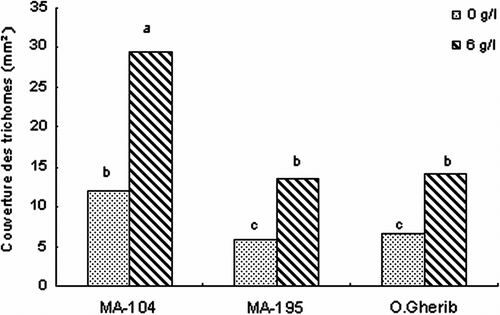
D’autre part, la salinité n’a pas eu d’effet significatif sur le diamètre des trichomes. Par contre, la surface en trichomes (ST) a augmenté sous ce stress (Tableau ). La comparaison entre clones suggère que MA-104 a développé la plus forte couverture en trichomes dépassant celle de MA-195 et O.Gherib de 110% (Fig. ).
Fig. 7 -Effect of NaCl on the trichomes foliar cover of white poplar clones (MA-104, MA-195 and O. Gherib). Values represent the mean of forty-eight individual measurements. Fig. 7. Effet de NaCl sur la couverture foliaire en trichomes des clones de peuplier blanc (MA-104, MA-195 et O.Gherib). Les valeurs représentent les moyennes de quarante-huit mesures individuelles.
Surface foliaire
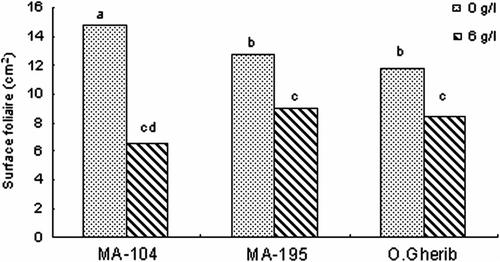
La réponse des plants de peuplier blanc à la salinité est exprimée par une réduction significative (p < 0,001) de la surface foliaire (Fig. ). La comparaison entre traitements indique une réduction significative (p < 0,001) et variable selon le clone de la surface foliaire des plants stressés par rapport aux plants témoins. Le clone MA-104 a enregistré la plus forte réduction (-55% du témoin) suivi des deux clones MA-195 et O.Gherib (-29%).
Fig. 8 -Effect of NaCl on the leaf area of white poplar clones (MA-104, MA-195 and O.Gherib). Values represent the mean of forty-eight individual measurements. Fig. 8. Effet de NaCl sur la surface foliaire des clones de peuplier blanc (MA-104, MA-195 et O.Gherib). Les valeurs représentent les moyennes de quarante-huit mesures individuelles.
Discussion
Les feuilles du peuplier blanc sont pourvues de stomates localisés pour leur totalité sur la face inférieure, contrairement au peuplier noir à titre d’exemple dont les stomates sont localisées sur la face inférieure et supérieure de la feuille. Ce sont des feuilles de type hypostomatique, qui est une forme d’adaptation contre une transpiration excessive liée au fort ensoleillement (Ait Said et al., Citation2005). Leur densité stomatique est élevée par comparaison à celles d’autres espèces ligneuses vivants sous un environnement humide (Ceulemans et al., Citation1995). Elle est comparable à celle du pistachier de l’Atlas, (283 stomates mm-2) (Ait Said et al., Citation2005) et à celle de divers hybrides de peuplier noir acclimatés aux conditions du semi-aride (170 – 288 stomates mm-2) (Pearce et al., Citation2005). Dans sa synthèse sur l’adaptation à la sécheresse des plantes, Ludlow (Citation1980) considère qu’un nombre élevé de stomates est perçu comme un caractère de xéromorphisme. Les clones de peuplier blanc ont montré des stomates à longueur relativement courte par comparaison à celle d’autres populations de peuplier noir sous environnements plus modérés (Pallardy & Kozlowsky Citation1979). Par contre, elle a été comparable à celle de Pinus sylvestris, (20 μm), (Meinder & Mainsfield Citation1968) et à celles de populations de peuplier adaptées à la sécheresse (19 – 22 μm) (Pearce et al., Citation2005). Ces feuilles renferment également des poils ou trichomes sur leur face inférieure leur donnant un aspect blanchâtre. Des différences entre les trois clones ont été observées. Les clones MA-195 et O.Gherib ont montré un nombre en trichomes comparable alors le clone MA-104 a développé un nombre plus élevé. Il a été démontré que la pubescence affecte directement la transpiration par la création d’une couche d’air permanente proche de la feuille réduisant le brassage de l’air et de ce fait réduisant la transpiration (Martin & Glover, Citation2007).Ces auteurs ont mentionné que les poils présents chez P. alba sont capables de refléter la lumière. Ce caractère permettrait de réduire la perte en eau, non pas par l’augmentation de la couche limite créée par ces poils, mais par une réduction de l’interception de la lumière induisant une baisse de la température de la feuille (Ehleringer, Citation1980).
L’application du traitement 6 g/l NaCl a induit chez le peuplier blanc des adaptations micromorphologiques mises en évidence par une augmentation de la densité des stomates (Fig. ) et par une réduction de leur dimension (Fig. et 4). La relation inverse entre dimension des stomates et densité stomatique reflète une adaptation à la salinité. En effet, l’augmentation de la densité des stomates confèrerait au peuplier un système de régulation stomatique plus efficace qui limiterait l’ouverture des ostioles et par conséquent réduirait la perte d’eau et le dessèchement. Un tel comportement a été noté chez des populations de peuplier noir confrontées à un déficit hydrique (Pearce et al., Citation2005), chez divers écotypes de mil tolérants le stress salin (Ben Jaballah, Citation2006) et chez l’arganier sous une pollution minière (Alilou et al., Citation2006). Cependant, des différences entre clones ont été révélées. Le clone MA-104 a montré la plus forte densité stomatique, comparé au clone O.Gherib qui en a montré la plus faible alors que le clone MA-195 a développé une réponse intermédiaire. L’accroissement de la densité stomatique peut augmenter l’assimilation nette du CO2 (Terashima et al., Citation2005) et diminuer la perte en eau: en effet un nombre élevé de stomates peut engendrer des stomates de petite taille et à fermeture rapide. Dans ce contexte, nos résultats ont montré que l’augmentation du nombre de stomates par unité de surface foliaire a été accompagnée par une réduction en longueur et en largeur de l’ouverture stomatique. Cette modification constitue un mécanisme d’adaptation au stress puisque les stomates deviennent globuleux et moins aplatis (Frank & Farquahr, Citation2001) avec fermeture rapide de l’ostiole. Cependant les clones de peuplier ont développé des stratégies d’adaptation différentes en réponse au stress. Le clone MA-104 a montré une réduction dans l’ouverture en longueur et en largeur de ses stomates alors que chez MA-195, l’ouverture en longueur des stomates n’a pas été modifiée. Par contre ce clone a développé la plus faible ouverture en largeur des stomates leur conférant une fermeture rapide. Chez le clone O.Gherib une réduction dans l’ouverture en longueur des stomates a été notée alors que leur ouverture en largeur n’a pas été affectée (Fig. et 4). Des accommodations de ce type ont été développées par divers écotypes de mil sous stress hydrique et salin (Ben Jaballah, Citation2006) et par l’arganier (Argania spinosa L.) sous pollution minière (Alilou et al., Citation2006). D’autre part, la réduction des dimensions des stomates s’est traduite par une réduction de la surface génératrice des pertes transpiratoires, évaluée par le calcul de la surface relative des pores stomatiques. Les valeurs moyennes de la SR sont relativement élevées par rapport à celles relevées chez la tomate (0,62 à 1,35%) Mougou (Citation1984), mais se rapprochent des surfaces relatives d’espèces désertiques (1,8 à 3,4%) (Al Farraji, Citation1983) et de l’olivier (1,96 à 2,89%) (Guerfel et al., Citation2007). Par ailleurs, Kramer (Citation1983) rapporte que ce paramètre ne dépasse pas 3% de la surface totale de la feuille quand les stomates sont complètement ouverts.
La salinité a augmenté le nombre de trichomes. La présence de nombreux trichomes joue un rôle très important dans la réduction des pertes d’eau à travers les stomates. En effet, l’abondance des trichomes ralentit la circulation de l’air en dessous de la feuille, ce qui entraîne une couche limite plus épaisse, diminuant ainsi la transpiration (Schwabe & Lionakis, Citation1996). Le changement de l’épaisseur de la couche limite affecte directement la diffusion du gaz carbonique et de la vapeur d’eau au niveau de la feuille et par conséquent la vitesse de la conductance stomatique (Bongi & Pallioti, Citation1994). D’autre part, les trichomes ont la capacité de réfléchir les radiations qui provoquent le réchauffement de la feuille, ce qui influence aussi bien la photosynthèse que la transpiration (Boujneh, Citation1997). Le même auteur souligne que des densités élevées en trichomes sont favorables pour la résistance à la sécheresse. Ces structures peuvent retenir l’eau sous forme de gouttelettes ou rosée (Benzing, Citation1980). Par ailleurs, le diamètre des trichomes du peuplier blanc n’a pas été affecté par la contrainte saline traduisant une caractéristique propre à l’espèce, non dépendante des facteurs du milieu. Par contre chez l’olivier, ce diamètre a augmenté chez la variété Chemlali, tolérante au stress hydrique (Guerfel, Citation2008). L’importance de ces structures foliaires face au stress salin a été évaluée par la couverture foliaire assurée par les trichomes. Les trois clones ont montré une augmentation de cette couverture mais de façon variable (Fig. ). D’après Flowers (Citation1985), les espèces possédant des poils supportent et tolèrent de plus fortes concentrations de sel dans leur xylème comparativement à celles qui en sont dépourvues. D’autre part, l’effet inhibiteur du sel sur la croissance des parties aériennes s’est répercuté négativement sur la surface foliaire qui a accusé une réduction plus prononcée chez le clone MA-104 (Fig. ). Cet effet pourrait être attribué à une inhibition de l’initiation foliaire (Zekri & Parsons, Citation1991) et/ou à un ralentissement de la division ou de l’élongation cellulaire (Cramer & Bowman, Citation1991a). Ce phénomène est considéré comme une tolérance à la salinité dans la mesure où des surfaces foliaires plus réduites, assurent une meilleure efficience de l’utilisation de l’eau (Omami & Hammes, Citation2006).
Une différence dans l’expression des mécanismes d’adaptation à la salinité, traduite par des modifications micromorphologiques des stomates et des trichomes et par une réduction de la surface foliaire, a été relevée entre les clones de peuplier blanc. Le clone MA-104 (tolérant) a montré une meilleure adaptation au seuil de salinité appliqué à travers les effets combinés d’une surface foliaire plus réduite, d’une densité stomatique élevée et d’une régulation plus sensible de l’ouverture des ostioles. Il a montré également la densité en trichomes la plus élevée et la plus importante couverture par unité de surface foliaire assurée par ces structures. De telles modifications constituent des critères de choix pour l’adaptation au stress osmotique. Chez le clone MA-195 (moyennement tolérant), une augmentation de la densité en stomates et en trichomes a été notée mais de manière moins intense comparativement à MA-104. Par contre, son accommodation à la salinité s’est reflétée à travers une faible ouverture en largeur des stomates qui est un mécanisme d’adaptation contre la déperdition de l’eau. Les modifications moins intenses observées chez le clone O.Gherib (sensible), notamment la non élasticité des ostioles expliquent sa moindre adaptation à la dose de sel appliquée. Ainsi, les clones de peuplier blanc ont développé des modifications anatomiques foliaires (Abassi et al., Citation2009) et micromorphologiques spécifiques à chacun pour s’adapter à la salinité. Les clones MA-195 et notamment MA-104 sont les plus préconisés à s’implanter dans des sites à salinité avoisinant 6 g/l NaCl.
Related Research Data
Bibliographie
- Abassi , M. , Albouchi , A. , Ben Mansoura , A. , Béjaoui , Z. , Rejeb , M.N. and Mougou , A. 2004 . Tolérance de divers clones de peuplier à la salinité . Ann. INRGREF , 6 : 17 – 34 .
- Abassi , M. , Albouchi , A. , Béjaoui , Z. , Rejeb , M.N. and Mougou , A. 2009 . Adaptations anatomiques foliaires développées par le peuplier blanc (Populus alba L.) face au stress salin . Ann. INRGREF , 13 : 153 – 163 .
- Ait-Said , S. , Smail-Saadoun , N and Kadi-Bennane , S . 2005 . Etude comparative de trois populations de Pistacia atlantica Desf. (Ain Oussera – Messad – Tamanrasset) par le biais des dimensions stomatiques. Actes des Journées Scientifiques de l’INRGREF . Ann. INRGREF , 7 : 19 – 26 .
- Al Farraji F. A. H., 1983.- Drought resistance evaluation of six desert species commonly used for sand dune fixation in Iraq. Ph.D. University of Gent, Belgium, 182 pp.
- Albouchi , A. , Ghrir , R. and El Aouni , M.H. 1977 . Endurcissement à la sécheresse et accumulation de glucides solubles et d’acides aminés libres dans les phyllodes d’Acacia cyanophylla Lindl . Ann.Sci.For. , 54 : 155 – 168 .
- Alilou , H. , Rouhi , R. and Idrissi Hassani , L.M. 2006 . Effet de la pollution minière sur les paramètres stomatique chez Argania spinosa (L.) . Biol. Biotec. , 5 : 41 – 45 .
- Ben Jaballah L., 2006. Diversité marpho-phenologique, caractérisation agro-physiologique et mecanismes d’adaptation aux contraintes hydriques et salines de populations autochtones de Mil (Pennisetum glaucum (L.) R. BR.). Thèse, INAT, Tunis, 291p.
- Benzing , D.H. 1980 . The biology of the bromeliads. Eureka , California : Mad River Press . 305 p.
- Beritognolo , I. , Piazzai , M. , Benucci , S. , Kuzminsky , E. , Sabatti , M. , Scarascia , G. and Muleo , R. 2007 . Functional characterization of three Italian Populus alba L. genotypes under salinity stress . Trees , 21 ( 4 ) : 465 – 477 .
- Bongi , G. and Palliotti , A. 1994 . “ Olive ” . In Hand book of environmental physiology of fruit crops , Edited by: Schaffer , B. and Anderson , P.C. 165 – 187 . Boca Raton Florida , , USA : CRC Press .
- Boujneh D., 1997. Variations morphologiques anatomiques et écophysiologiques en rapport avec la résistance à la sécheresse chez l’olivier (Olea europeaa L.). Thèse, Fac. Sci. Agr. Univ. Gend, Belgique.
- Ceulemans , R. , van Pract , L. and Jiang , X.N. 1995 . Effects of CO2 enrichment leaf position and clone on stomatal index and epidermal cell density in poplar (Populus) . New Phytol. , 131 : 99 – 107 .
- Cramer , G.R. and Bowmann , D.C. 1991 . Kinetics of maize leaf elongation. I. Increase yield threshold limits short-term, steady state elongation rates after exposure to salinity . J.Exp.Bot. , 42 : 1417 – 1426 .
- Ehleringer , J. 1980 . “ Leaf morphology and reflectance in relation to water and temperature ” . In Adaptation of plants to water and high temperature stress , Edited by: Turner , N.C. and Kramer , P.J. 123 – 134 . New York : A Wiley Inter Science Pub .
- Flowers , T.J. 1985 . Physiology of Halophytes . Plant Soil , 89 : 41 – 56 .
- Franks , P. and Farquah , G.D. 2001 . The effect of exogenus abcisic acid on stomatal development, stomatal mechanics and leaf gaz exchange in Trade scantia virginia . Plant Physiol. , 125 : 935 – 942 .
- Guerfel , M. , Boujneh , D. , Baccouri , B. and Zarrouk , M. 2007 . Evaluation of morphological and physiological traits for drought tolerance in 12 Tunisian Olive varieties (Olea europea L.) . J. Agro. , 6 ( 2 ) : 356 – 361 .
- Guerfel, M., 2008. Caractérisation morphologique structurale et biochimique de l’adaptation à la sécheresse chez l’olivier (Olea europea L). Thèse, Fac. Sci. Tunis, 161 p.
- Hachicha , M. , Job , J.O. and Mtimet , M. 1995 . Sols irrigués et sols sodiques. Les sols salés et la salinisation en Tunisie . Sols de Tunisie, Bulletin de la Direction des sols. , 15 : 341
- Hoagland D.R and D.I. Arnon, 1950. The water culture method for growing plants without soil. Circ. 347 Univ. Of Calif. Agric. Exp. Station, Berkley.
- Jones H.G., 1983. A quantitative approach to environmental plant physiology. In: Plants and microclimat: a quantitative approach to environmental plant physiology. Jones H.G, (2nd ed.), Cambridge University Press, Cambridge. 289 p.
- Kramer , P.J. 1983 . Water relations of plants , New York , ND : Academic Press . 489
- Ludlow , M.M. 1980 . “ Adaptative significiance of stomatal response to water stress ” . In Adaptation of Plants and High temperature stress , Edited by: Turner , N.C. and Kramer , P.J.A. 123 – 128 . New York : Wiley Interscience Publi .
- Martin , C. and Glover , B.J. 2007 . Functional aspect of patterning in aerial epidermis . Plant Biol. , 10 : 70 – 82 .
- Meinder , H. and Mansfield , T.A. 1968 . Physiology of stomata , New York : Mc Grow-Hill . 179
- Mougou A., 1984. Evaluation de la résistance à la sécheresse par des paramètres morphologiques, écophysiologiques et biochimiques chez plusieurs espèces de tomate. Thèse, Fac. Sci. Agr. Univ. Gend. Belgique.
- Omami , E.N. and Hammes , P.S. 2006 . Interactive effects of salinity and water stress on growth, leaf water relations, and gas exchange in amaranth (Amaranthus spp.) . N. Z. J. Crop Hortic Sci , 34 ( 1 ) : 33 – 44 .
- Pallardy , S.G. and Kozlowski , T.T. 1979 . Frequency and length of stomata of 21 Populus clones . Can. J. Bot. , 57 : 2519 – 2523 .
- Pallardy , S.G. and Kozlowski , T.T. 1980 . Cuticle development in the stomatal region of Populus clones . New. Phytol. , 85 : 363 – 368 .
- Pearce , D.W. , Millard , S. , Bray , D.F. and Rood , S.B. 2005 . Stomatal characteristics of riparian poplar species in a semi-arid environment . Tree Physiol. , 26 : 211 – 218 .
- Rengel , Z. 1992 . The role of calcium in salt toxicity . Plant. Cell. Env. , 15 : 625 – 632 .
- Schwabe , W.W. and Lionakis , S.M. 1996 . Leaf attitude in olive in relation to drought resistance . J. Hortic. Sci. , 71 : 157 – 166 .
- Sixto , H. , Grau , J.M. , Alba , N. and Alia , R. 2005 . Response to sodium chloride in different species and clones of genus Populus L . Forestry , 78 ( 1 ) : 93 – 104 .
- Terashima , I. , Araya , T. , Miyazawa , S.I. , Sone , K. and Yano , S. 2005 . Construction and maintenance of the optimal photosynthetic systems of the leaf, herbaceous plant and tree: an eco-development treatise . Ann. Bot , 95 : 507 – 519 .
- Zekri , M. and Parsons , L.R. 1991 . Response of split–root sour orange seedlings to NaCl and polyethylene glycol stresses . J. Exp. Bot , 41 : 35 – 40 .