Résumé
Cette étude consiste à effectuer une évaluation préliminaire des propriétés édaphiques et floristiques des parcours steppiques d’armoise blanche dégradés de la région de Saïda (Algérie occidentale), réhabilités par la plantation d’Atriplex halimus subsp. schweinfurthii et d’Atriplex canescens. Les résultats obtenus montrent l’adaptation de ces deux espèces aux conditions édaphiques et climatiques de la région d’étude avec un taux de réussite supérieur à 80%. Comparativement au périmètre steppique dégradé non réhabilité (témoin), dans les périmètres où ont été introduites les deux espèces d’Atriplex, nous avons noté l’amélioration du taux de recouvrement de la végétation, de la phytomasse, de l’épaisseur et de l’humidité du sol. Cependant, une légère augmentation de la salinité du sol caractérise le périmètre d’Atriplex canescens. L’inventaire floristique dénote une diversité spécifique similaire dans les trois périmètres explorés.
Abstract
This study consist in performing a preliminary assessment of soil and floristic properties of degraded steppes of white wormwood in the region of Saïda (western Algeria), rehabilitated by the planting of Atriplex halimus subsp. schweinfurthii and Atriplex canescens. The results show the adaptation of these two species to edaphic and climatic conditions of the study area with a success rate above 80%. Compared to the non rehabilitated perimeter (control), in the perimeters where the two species of Atriplex were introduced, we note improved recovery rate of vegetation, phytomass, thickness and humidity of soil. However, a slight increase of soil salinity characterizes the perimeter of Atriplex canescens. The plant inventory indicates a similar specific diversity in the three explored perimeters.
Introduction
La désertification est et restera un défi de taille aux chercheurs qui s’occupent de ce péril pesant continuellement sur la qualité et la quantité des ressources naturelles dont dépend l’existence des êtres vivants à l’échelle de la planète. Dans l’ensemble des pays du Maghreb, depuis plusieurs années, nous assistons à un déséquilibre agro-sylvo-pastoral qui se manifeste par une dégradation des potentialités pastorales et forestières (Abdelguerfi & Laouar, Citation2000).
En Algérie, la désertification concerne essentiellement les steppes des régions arides et semi-arides qui ont toujours été l’espace privilégié de l’élevage ovin extensif. Depuis plus d’une trentaine d’années, ces écosystèmes connaissent une dégradation de plus en plus accentuée de toutes leurs composantes (flore, couvert végétal, sol et ses éléments, faune et son habitat; Le Houérou, Citation1985; Aidoud &Touffet, Citation1996; Nedjraoui & Bedrani, Citation2008; Wael et al., Citation2009).
Actuellement, le constat majeur est celui d’une diminution de la superficie de ces steppes et de leur dégradation parfois extrême. Ces transformations au niveau spatial se sont aussi accompagnées de modifications qualitatives importantes suite à une diminution ou une disparition totale des espèces ayant de bonnes valeurs pastorales (Floret et al., Citation1981) en laissant la place à des espèces non palatables (espèces épineuses ou toxiques: Atractylis humilis, Noaea mucronata, Peganum harmala, etc.; Kadi-Hanifi, Citation1998; Aidoud et al., Citation2006). Il en ressort que la production pastorale dans ces parcours a globalement été marquée par un déclin significatif. Des changements particulièrement rapides et intenses se sont opérés dans ces milieux sous la pression des besoins croissants des populations. Ces derniers sont à l’origine de l’extension des cultures, des changements de politique de gestion et donc des usages et pratiques d’élevage, le tout aggravé par des sécheresses périodiques plus ou moins sévères et prolongées (Bouchetata Tarik & Bouchetata Arslan, Citation2005; Aidoud et al., Citation2006).
Compte tenu de l’état de dégradation de ces écosystèmes naturels, la reconstitution du couvert végétal ne peut plus être assurée dans la plupart des cas par les mécanismes de la régénération naturelle et donc le recours à des techniques sophistiquées d’aménagement et de gestion des terres sont devenues nécessaires. Ces techniques se basent sur l’utilisation judicieuse des eaux de pluie et la plantation d’espèces ligneuses contribuant aussi bien à l’accroissement de la production qu’à la protection des sols contre l’érosion (Nefzaoui & Chermiti, Citation1991).
L’introduction d’espèces fourragères, souvent exotiques (des genres Opuntia, Atriplex et Acacia), en plantations monospécifiques, a en général été préférée à la réintroduction d’espèces autochtones (Aidoud et al., Citation2006). Les avantages de l’utilisation de ces espèces, en particulier les Atriplex, réside dans leurs stratégies d’adaptations écophysiologiques, leur grande résistance à l’aridité et à la salinité (Pujol et al., Citation2001; Belkhodja & Bidai, Citation2004), leur capacité à produire une biomasse utilisée dans des conditions marginales comme fourrage pour le bétail (Naggar, Citation2000; Mulas & Mulas, Citation2004; Ighilhariz et al., Citation2008). En plus, leurs systèmes racinaires très ramifiés jouent un rôle important dans la réhabilitation des sols dégradés, la lutte contre l’érosion des sols et la désertification (Abbad et al., Citation2004).
Dans ce contexte, nous avons établi un premier diagnostic des principales caractéristiques édaphiques et floristiques relatives à des périmètres steppiques d’armoise blanche (Artemisia herba alba Asso) dégradés, réhabilités par la plantation d’Atriplex halimus subsp. schweinfurthii et d’Atriplex canescens dans la commune de Moulay Larbi (wilaya de Saïda). Ce travail s’intègre dans le cadre d’un suivi de la dynamique de ces écosystèmes artificiels qui ont tendance à occuper des superficies croissantes suite à l’intervention de l’homme.
Zone et méthodes d’étude
Zone d’étude
Notre zone d’étude est située dans la commune de Moulay Larbi (wilaya de Saïda), entre 34° et 35° de latitude nord et entre 1° et 0° de longitude ouest. Sur la base de vingt années de données climatiques, de 1985 à 2005 (Office national de la météorologie, 2006), il ressort que cette zone est caractérisée par un climat aride à hiver frais, avec une pluviométrie moyenne annuelle de 327 mm et une période sèche qui s’étale sur sept mois, d’avril à octobre. La pluviométrie de la région est caractérisée par une grande variabilité intra et interannuelle.
Les témoignages des techniciens du Haut Commissariat au développement de la steppe (HCDS) de Saïda confirment que notre zone d’étude est un parcours steppique à base d’armoise blanche (Artemisia herba-alba Asso.) dégradée, réhabilité en 2003 par plantation d’Atriplex halimus subsp. schweinfurthii et d’Atriplex canescens.
Dans cette zone d’étude, trois périmètres steppiques contigus sont choisis: le premier périmètre de 45 ha est planté en Atriplex canescens, le deuxième de 25 ha est planté en Atriplex halimus subsp. schweinfurthii et le troisième de 40 ha, non planté, est considéré comme témoin.
Les plants d’Atriplex sont préalablement cultivés en pépinière durant trois mois avant d’être transplantés en zone steppique. L’agréage des plants avant leur transfert aux périmètres est basé essentiellement sur la hauteur de la partie aérienne (entre 15 et 20 cm), sur leur vitalité et leur état sanitaire.
Les plantations sont effectuées dans des potêts établis le long des sillons. La distance séparant un sillon de l’autre est de 5 m. L’écartement entre deux potêts est de 2.5 m. La densité des plantations est estimée à 800 plants / ha. Après plantation, trois arrosages sont effectués à raison de 20 litres / plant sur une période de deux mois.
Méthodes d’étude
Étude de la végétation et du sol
L’état du développement et de la croissance des arbustes d’Atriplex dans les périmètres de plantation est mis en évidence en tenant compte des trois paramètres suivants:
| - | le taux de réussite des Atriplex, exprimé en pourcentage, correspond au rapport du nombre de plants réussis au nombre total de plants plantés; | ||||
| - | la hauteur et le diamètre moyens des plants présents à l’intérieur des placettes, au niveau des relevés floristiques effectués. | ||||
Ensuite, pour la caractérisation de la composition floristique au niveau des trois périmètres choisis, six relevés phytoécologiques correspondant chacun à une aire minimale de 100 m2 y sont établis aléatoirement, durant la période de végétation (mars-avril 2008) (Braun-Blanquet, Citation1951; Djebaili, Citation1984). Les espèces inventoriées sont réparties en familles et en types biologiques correspondants. Pour chaque relevé, sont notées les principales données stationnelles (altitude, pente, exposition, texture du sol et recouvrement de la végétation). Chaque espèce présente dans le relevé est affectée des coefficients d’abondance-dominance et de sociabilité, estimés selon l’échelle de Braun-Blanquet (Citation1951); le recouvrement basal des espèces pérennes, correspondant à la surface (S) occupée par les partie aériennes de leurs individus au niveau du sol, est également noté. Cette surface est exprimé en m2 et est calculée par la formule S = π x (D / 2)2, D étant le diamètre de la partie aérienne de l’espèce (Gounot, Citation1969).
En vue de comparer la productivité des périmètres choisis, nous avons évalué la phytomasse du cortège floristique accompagnant les Atriplex (Aidoud, Citation1983). Ceci permettra de savoir dans quel périmètre la contribution du couvert végétal à la protection du sol et l’offre fourragère sont les plus importantes. Ces mesures concernent la partie aérienne de la végétation (phytomasse épigée). Pour cela, sur six placettes de 1 m2 établies dans chacun des trois périmètres, les parties aériennes des plantes herbacées (pérennes et annuelles) sont prélevées puis pesées pour avoir le taux de matière fraîche. Le taux de matière sèche est obtenu après séchage à l’étuve, à 105°C pendant 24 heures.
Pour l’étude du sol, trois profils pédologiques sont réalisés dans chaque périmètre. Au niveau de chaque profil, nous avons mesuré l’épaisseur du sol (de la surface jusqu’à la dalle calcaire). Les paramètres pédologiques évalués sont: la texture, la conductivité électrique, le pH, l’humidité, le taux de matière organique et le taux de calcaire actif et total (Aubert, Citation1978).
Traitement des données
Nous avons utilisé des indices de similarité (indice de Sorensen et indice de Jaccard) et de diversité spécifique (indice de Shannon et indice d’équitabilité) pour comparer respectivement la similarité de la composition floristique et la richesse spécifique des trois périmètres d’étude (Gounot, Citation1969).
Les paramètres édaphiques et floristiques des trois périmètres étudiés sont traités par l’analyse en composantes principales (ACP) en utilisant le logiciel STATISTICA 6.0. La matrice de cette analyse est composée de trois lignes correspondant aux trois périmètres et vingt-et-une colonnes présentant les différents paramètres édaphiques et floristiques. C’est une analyse qui résume la variabilité de ces paramètres pour un nombre réduit de variables synthétiques, appelées axes factoriels définissant le premier plan factoriel de l’ACP. Les valeurs des différents paramètres édaphiques et floristiques projetés ont des coordonnées comprises entre –1 et +1 à l’intérieur d’un cercle de corrélations. L’interprétation des axes factoriels se fait à partir de l’examen du cercle de corrélation et de la position des périmètres retenus sur les axes factoriels (Phillippeau, Citation1986).
Résultats et discussion
Résultats
Analyse de la végétation
Les données enregistrées indiquent l’adaptation du genre Atriplex aux zones arides de la région d’étude, justifiée par un taux de réussite élevé (> 80%): 89.4% pour Atriplex canescens et 94.2% pour Atriplex halimus. Comparativement à Atriplex canescens (48.5 cm), les touffes d’Atriplex halimus présentent une croissance en diamètre plus importante (60.7 cm). Leurs hauteurs moyennes sont comparables, avoisinant 53 cm.
Le recouvrement global de la végétation le plus important correspond au périmètre de plantation à base d’Atriplex halimus avec un taux de 20%, dont la contribution des touffes est estimée à 4% et le reste, soit 16%, correspond à la strate herbacée représentée par les espèces spontanées. Le périmètre de plantation à base d’Atriplex canescens est caractérisé par un recouvrement global de 15%, dont 2% pour Atriplex canescens et 13% pour la strate herbacée. Le taux de recouvrement du périmètre non planté est de 12% (Tableau ).
Tableau 1 Taux de recouvrement et phytomasse des périmètres d’étude. Table 1: Cover rate and phytomass of study perimeters.
L’analyse du Tableau montre que le recouvrement des espèces pérennes, en particulier celui des touffes d’Artemisia herba-alba est plus important dans le périmètre d’Atriplex canescens (4%) par rapport au périmètre d’Atriplex halimus (2.5%). En revanche, le taux de sable élevé dans le périmètre d’Atriplex halimus (Tableau ) a favorisé l’installation de Lygeum spartum, espèce psammophile, avec un recouvrement de l’ordre de 4.2%. Le recouvrement de l’espèce Noaea mucronata est plus important dans le périmètre non planté (6%).
Tableau 2. Relevés phytoécologiques. Table 1: Phytoecological statments.
La production de matière sèche des herbacées est meilleure dans les deux périmètres de plantation à base d’Atriplex. Elle est de l’ordre de 255 et 360 kg MS / ha, respectivement dans les périmètres d’Atriplex canescens et d’Atriplex halimus. Cependant, dans le périmètre non planté, nous avons noté un taux de 150 kg MS / ha (Tableau ).
Sur le Tableau est représentée la liste des espèces inventoriées dans les trois périmètres étudiés ainsi que leurs coefficients d’abondance-dominance et de sociabilité correspondants. Vingt-quatre espèces appartenant à la strate herbacée sont recensées, dont 58.3% sont communes aux trois périmètres. Les espèces exclusives à chaque périmètre sont très peu nombreuses (au plus trois espèces).
Tableau 3 Analyses pédologiques. Table 3: Soil analysis.
L’ensemble des espèces est réparti sur onze familles caractérisant les périmètres non plantés et plantés d’Atriplex canescens et dix familles pour le périmètre d’Atriplex halimus. La famille dominante est celle des Astéracées avec cinq espèces dans les périmètres non planté et d’Atriplex halimus et quatre espèces dans le périmètre d’Atriplex canescens. Les familles des Poacées et des Cistacées viennent en deuxième et troisième position, avec respectivement trois et deux espèces dans chacun des trois périmètres. Les autres familles (Caryophyllacées, Fabacées, Plantagacées, Amaranthacées, Malvacées, Lamiacées, Euphorbiacées, Lilliacées et Valérianacées) sont monospécifiques.
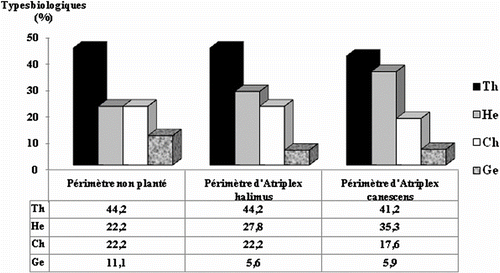
Les vingt quatre espèces recensées sont réparties en onze thérophytes, sept hémicryptophytes, quatre chaméphytes et deux géophytes. Sur les trois périmètres, les thérophytes présentent une forte contribution au tapis végétal avec un pourcentage oscillant entre 41.2% et 44.2% (Fig. ). Les hémicryptophytes et les chaméphytes sont moyennement représentés. Le pourcentage des hémicryptophytes varie entre 22.2% et 35.3% dans les trois périmètres, celui des chaméphytes entre 17.6% et 22.2%. Les géophytes sont faiblement représentés avec des valeurs se situant entre 5.6% et 11.1%.
Fig. 1 Spectres biologiques des trois périmètres Th: thérophytes; He: hémicryptophytes; Ch: chaméphytes; Ge: Géophytes. Fig. . Biological spectrum of the three perimeters Th: therophytes; He: hemicryptophytes; Ch: chamaephytes; Ge: Geophytes.
Analyse du sol
Les résultats des analyses pédologiques (Tableau ) indiquent, pour les trois périmètres étudiés, un pH légèrement alcalin, variant en moyenne entre 7.2 et 7.4. L’épaisseur maximum du sol dans le périmètre non planté est de 17 cm, alors qu’elle est de 20 à 21 cm dans les périmètres de plantation à base d’Atriplex. La texture du sol est argilo-limoneuse dans le périmètre non planté et le périmètre planté en Atriplex canescens; dans le périmètre planté en Atriplex halimus, elle est sablo-argilo-limoneuse. Le taux d’humidité moyen varie d’un périmètre à l’autre: faible (5.3%) dans le périmètre non planté, moyen dans les périmètres plantés en Atriplex halimus (6.8%) et Atriplex canescens (8.3%).
Le taux moyen de la matière organique dans les trois périmètres est faible avec un maximum de 4.6% caractérisant le périmètre d’Atriplex canescens.
La conductivité électrique des sols analysés varie entre 0.2 mmhos / cm et 0.7 mmoh / cm dans le périmètre non planté et entre 0.4 mmhos / cm et 0.8 mmhos / cm dans le périmètre d’Atriplex halimus. Cette conductivité est plus ou moins élevée dans le périmètre d’Atriplex canescens, oscillant entre 0.5 mmhos / cm à 1.3 mmhos / cm. La teneur en calcaire total est élevée dans les trois périmètres, en particulier pour le périmètre non planté où elle varie entre 17.7% et 28.2%.
Traitement des données
Les valeurs des indices de similarité de Sorensen et de Jaccard permettant de comparer les listes floristiques des trois périmètres, montrent que la quasi-totalité des indices sont supérieurs à 50% (Tableau ).
Tableau IV Similarité et diversité spécifique des périmètres d’étude. Table 4: Similarity and species diversity of study perimeters.
Les valeurs de l’indice de diversité de Shannon (H’) sont très proches dans les trois périmètres d’étude: 3.1 bits / individu pour le périmètre non planté, 3.2 bits / individu pour le périmètre planté en Atriplex canescens et 2.9 bits / individu pour le périmètre planté en Atriplex halimus. Les valeurs de l’indice d’équitabilité montrent une répartition équitable des espèces dans les trois périmètres, il est de l’ordre de 0.7 pour le périmètre non planté et le périmètre planté en Atriplex halimus, 0.8 pour le périmètre planté en Atriplex canecsens.
Dans l’analyse en composantes principales effectuée sur les paramètres édaphiques et floristiques des trois périmètres (Fig. ), le plan F1-F2 est retenu car il rend compte d’un maximum d’informations (Fact. 1: 61.2% et Fact. 2: 38.8%) sur les corrélations existantes entre les trois périmètres et leurs caractéristiques floristiques et édaphiques. L’axe F1 exprime un gradient de dégradation et l’ensemble des trois périmètres est classé sur cet axe en fonction de la composition floristique et l’épaisseur du sol qui présentent les fortes contributions: les thérophytes (0.077), les géophytes (0.077), les hémicryptophytes (0.076), l’épaisseur du sol (0.076). Sur l’axe 2, les trois périmètres sont séparés suivant leurs principales caractéristiques édaphiques où le sable, la salinité et le pH présentent les plus fortes contributions (0.121). La contribution de la matière organique est de 0.105.
Fig. 2 Cercle des corrélations. Ep: épaisseur du sol; Hum: humidité du sol; He: hémicryptophytes; Sali: salinité du sol; Armo: recouvrement d’Artemisia herba-alba; M.O: matière organique; Stpa: recouvrement de Stipa parviflora; Arg: argile; Limo: limon; Noae: recouvrement de Noaea mucronata; Th: thérophytes; Ge: géophytes; Spar: recouvrement de Lygeum spartum; CaCo: calcaire actif; MSha: taux de matière sèche; Reco. Annu: recouvrement des espèces annuelles; Nbsp: nombre d’espèces; Ch: chaméphytes; Sabl: sable. Fig. . Correlations circle Ep: thickness of soil; Hum: soil humidity; He: hemicryptophytes; Sali: soil salinity; Armo:Artemisia herba-alba cover; M.O: rate of organic matter; Stpa: Stipa parviflora cover; Arg: clay; Limo: loam; Noae: Noaeamucronata cover; Th: therophytes; Ge: geophytes; Spar: Lygeum spartum cover; CaCo: active calcium carbonate; MSha: dry matter rate; Reco. Annu: annual plants cover; Nbsp: number of species; Ch: chamaephytes; Sabl: Sand.
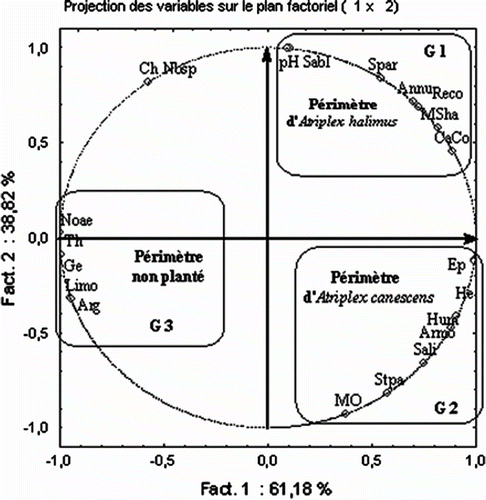
Le cercle des corrélations permet la distinction des trois groupes de variables suivants (Fig. ):
| - | le groupe G 1 corrélé au périmètre d’Atriplex halimus est composé de: sable (Sabl), recouvrement global (Reco), taux de matière sèche / ha (MSha), recouvrement de Lygeum spartum (Spar), pH du sol, taux de recouvrement des annuelles (Annu), calcaire actif (CaCo); | ||||
| - | le groupe G 2 corrélé au périmètre d’Atriplex canescens est composé de: épaisseur du sol (Ep), humidité relative (Hum), hémicryptophytes (He), salinité (Sali), recouvrement d’Artemisia herba-alba (Armo), matière organique (M.O), recouvrement de Stipa parviflora (Stpa); | ||||
| - | le groupe G 3 corrélé au périmètre non planté est composé de: argile (Arg), limon (Limo), recouvrement basal de Noaea mucronata (Noae), thérophytes (Th), géophytes (Ge). | ||||
Nous remarquons que sur le côté positif de l’axe F1 sont positionnés les deux périmètres de plantation à base d’Atriplex halimus et d’Atriplex canescens, présentant respectivement des taux de recouvrement (Reco) de l’ordre de 20% et 15%. Le taux de recouvrement est corrélé positivement à d’autres variables avec des coefficients importants: Reco-Annu (+ 0.999), Reco-Spar (+ 0.973), Reco-MSha (+ 0.989), Reco-He (+ 0.630).
Sur le côté négatif de cet même axe, le recouvrement est corrélé négativement aux variables suivantes: Reco-Th (- 0.785), Reco-Ge (- 0.785), Reco-Noae (- 0.709). Ces variables caractérisent l’état de dégradation du couvert végétal à l’intérieur du périmètre non planté où le recouvrement est de 12% (Fig. ).
Sur le côté positif de l’axe F2, la forte teneur du sol en sable dans le périmètre d’Atriplex halimus est corrélée positivement au recouvrement de Lygeum spartum (Sabl-Spar: + 0.890) et aux espèces annuelles (Sabl-Annu: + 0.785). Sur le côté négatif de l’axe F2, nous remarquons que l’abondance de Artemisia herba-alba est fortement corrélée à l’humidité, l’épaisseur et le taux de matière organique du sol: Armo-Hum (+ 0.997), Armo- Ep (+ 0.929), Armo-MO (+ 0.773).
Discussion
Les résultats obtenus indiquent l’adaptation de Atriplex halimus et de Atriplex canesens aux conditions climatiques et édaphiques de la commune de Moulay Larbi où elles ont été transplantées en 2003. Cette adaptation s’explique par le pourcentage de réussite encourageant, dépassant les 80%. Elle est assurée essentiellement par le système racinaire pivotant et par la morphologie des feuilles de ces espèces qui présentent une anatomie de type Kranz, caractéristique des plantes appartenant physiologiquement au groupe C4 ayant grande productivité (Mulas & Mulas, Citation2004; Smail-Saadoun, Citation2005; Ighilhariz et al., Citation2008).
Il ressort également qu’une amélioration du recouvrement végétal du sol est observée dans les périmètres plantés en Atriplex avec un taux qui varie entre 15% et 20% contre 12% dans le périmètre non planté. Cette amélioration est attribuée essentiellement au bon développement de la végétation en présence d’Atriplex, favorisée certainement par l’épaississement du sol (piégeage du sable), la diminution de l’évapotranspiration (effet self mulching) et la bonne protection du sol.
Le recouvrement des touffes d’Artemisia herba-alba est plus important dans la plantation d’Atriplex canescens (4%) où la texture fine à base de limons et d’argiles et l’épaisseur du sol favorisent sa croissance (Pouget, Citation1980; Djebaili, Citation1984; Aidoud et al., Citation2006). En revanche, l’installation de Lygeum spartum, espèce psammophile, avec un recouvrement de l’ordre de 4.2% en association avec Artemisia herba-halba est favorisée par la nature sablo-argilo-limoneuse du terrain dans le périmètre d’Atriplex halimus. En effet, Lygeum spartum apparaît dans les steppes à Artemisia herba-alba chaque fois que la surface est ensablée (Aidoud et al., Citation2006). Le recouvrement de Noaea mucronata est plus important dans le périmètre non planté (6%), ce qui atteste l’état de dégradation de ce périmètre.
La production des herbacées (phytomasse) en matière sèche est caractérisée par une nette augmentation dans les deux périmètres plantés en Atriplex: 360 kg MS / ha et 255 kg MS / ha respectivement dans les périmètres d’Atriplex halimus et d’Atriplex canescens. Cette augmentation trouve son origine essentiellement dans la présence de touffes d’Artemisia herba-alba et de Lygeum spartum plus volumineuses et bien développées à l’intérieur des plantations d’Atriplex. Cependant, la faible phytomasse dans le périmètre non planté (150 kg MS/ha) est assurée par des espèces naines, épineuses et à faible valeur fourragère, en l’occurrence Noaea mucronata.
L’inventaire de la végétation montre l’existence de vingt-quatre espèces appartenant à la strate herbacée, dont 58.3% sont communes aux trois périmètres, expliquant ainsi la forte similarité floristique (> 50%). Le nombre d’espèces recensées dans les trois périmètres au moment du relevé (avril 2008) est très faible. Ceci s’explique notamment par les insuffisances pluviométriques ayant marqué la période de notre étude où il a été enregistré, selon l’Office national de la météorologie de Saïda, un cumul pluviométrique de 57,1 mm entre janvier et avril 2008. Ces espèces sont réparties équitablement entre les trois périmètres: 18 espèces dans les périmètres non planté et d’Atriplex halimus, 17 espèces dans le périmètre d’Atriplex canescens. Cette équitabilité (environ 0,7) s’est traduite par une diversité spécifique comparable entre les trois périmètres avec des valeurs moyennes oscillant entre 2.9 et 3.2 bits / individu. Ces espèces appartiennent à onze familles dont les plus importantes sont les Astéracées et les Poacées et à trois types biologiques importants, à savoir les thérophytes, les hémicryptophytes et les chaméphytes. Les thérophytes dominent dans les trois périmètres à cause de l’aridité du climat, signalée par de nombreux auteurs (Gounot, Citation1969; Daget, Citation1980; Barbero et al., Citation1990; Dahmani-Megrourouche,Citation1997; Kadi-Hanifi, Citation1998) et de l’anthropisation (Grime, Citation1977).
Les géophytes présentent un faible taux dans les trois périmètres étudiés (11.1% dans le périmètre non planté, 5.9% dans le périmètre d’Atriplex canescens et 5.6% dans le périmètre d’Atriplex halimus), confirmant ainsi leur régression dans les pelouses et les zones steppiques (Barbero et al., Citation1989).
Les paramètres physico-chimiques du sol varient entre les trois périmètres. En effet, l’épaisseur maximum du sol dans le périmètre non planté est de 17 cm, atteint 20 à 21 cm dans les périmètres de plantation à base d’Atriplex. Cette amélioration reflète une bonne protection du sol qui serait due essentiellement au piégeage du sable par les touffes d’Atriplex, en particulier Atriplex halimus, au ralentissement de la vitesse du vent par ces touffes constituant un brise-vent, au recouvrement global assez important du sol dans les périmètres plantés en luttant ainsi contre l’érosion et le desséchement du sol, renforcé par un réseau dense de systèmes racinaires stabilisant le sol.
La variabilité de la texture du sol des trois périmètres s’observe par une augmentation du taux de sable, en particulier dans le périmètre de plantation d’Atriplex halimus. Cette texture légère présente un double effet: d’une part, elle améliore les conditions hydriques du sol par l’amélioration de la perméabilité; d’autre part, elle favorise l’installation d’espèces psammophiles (Lygeum spartum, Atractylis serratuloides, etc.) et certaines thérophytes (Pontanier et al., Citation1995).
L’humidité varie entre les trois périmètres, elle est fonction principalement de l’épaisseur, de la texture du sol et du recouvrement global de la végétation. Le périmètre non planté affiche le taux d’humidité le plus faible (5.3%). La texture argilo-limoneuse du périmètre planté en Atriplex canescens ainsi que son taux de recouvrement agissent en synergie pour retenir une quantité suffisante d’eau, se traduisant par l’amélioration du taux d’humidité (8.3%). Dans le périmètre planté en Atriplex halimus, l’amélioration du taux d’humidité (6.8%), mais à moindre degré par rapport au périmètre précédent, s’explique également par le taux de recouvrement suffisant et le taux élevé de sable. Ce dernier, ayant une forte porosité, favorise la perméabilité mais diminue la rétention de l’eau, en particulier dans les premiers centimètres de la surface du sol.
Le taux de matière organique dans les trois périmètres est faible. Le pourcentage le plus élevé est noté dans le périmètre d’Atriplex canescens avec une moyenne de 4.6%. Cela s’expliquerait par l’abondance du couvert végétal dominé par Artemisia herba-alba, espèce possédant un système racinaire développé représentant 50% à 80% de la biomasse totale (Pouget, Citation1980), par la texture plus ou moins fine (argilo-limoneuse) assurant un temps de résidence prolongé de la matière organique dans l’horizon superficiel (Bernoux, Citation2008) et par le pourcentage élevé de la matière organique d’Atriplex canescens en se référant à la composition chimique établie par Nefzaoui & Chermiti (Citation1991) sur les différentes espèces d’Atriplex.
Malgré l’amélioration du taux de recouvrement dans le périmètre d’Atriplex halimus, la teneur moyenne du sol en matière organique est la plus faible (3.9%). Cela s’expliquerait en partie par le taux élevé de sable dans ce périmètre, favorisant l’aération et le drainage du sol permettant ainsi une biodégradation importante de sa matière organique (Bernoux, Citation2008). Dans le périmètre non planté, le faible taux de la matière organique (4.1%) est dû au faible recouvrement (12%) et à la nature des espèces présentes dans ce périmètre dont la majorité sont naines et à surface foliaire réduite.
Les mesures de la conductivité du sol montrent des variations entre les trois périmètres avec des valeurs plus élevées dans les périmètres où l’Atriplex a été introduit. En effet, la conductivité varie entre 0.2 mmhos / cm et 0.7 mmhos / cm dans le périmètre non planté et entre 0.4 et 0.8 mmhos / cm dans le périmètre d’Atriplex halimus. Cette variation est plus marquée dans le périmètre d’Atriplex canescens, entre 0.5 à 1.3 mmhos / cm. Ces observations rejoignent celles de Pouget (Citation1980) en régions steppiques sud-algéroises. Le taux élevé de sels dans les périmètres plantés est dû certainement au caractère xérohalophytique du genre Atriplex. En effet, ce genre a la propriété de pomper le sel du sol et de l’accumuler dans ses feuilles. Une fois étalées sur le sol, ces dernières contribuent à l’élévation de sa conductivité électrique. Ces variations dans la teneur du sol en sels nécessitent un suivi pour mettre en exergue l’influence de l’utilisation des Atriplex sur l’évolution des sels dans les sols steppiques.
La forte teneur en calcaire dans les trois périmètres est justifiée par la présence d’une dalle calcaire très proche de la surface enrichissant le sol en carbonate de calcium.
La synthèse de toutes ces données par l’ACP fait ressortir sur les trois groupes de variables dégagés (Fig. ) que l’amélioration des caractéristiques édaphiques dans les périmètres plantés en Atriplex, en particulier l’épaisseur du sol et sa texture, s’est répercutée d’une part sur l’état du couvert végétal par une augmentation du recouvrement et de la production de matière sèche et, d’autre part, sur la composition floristique. En effet, nous avons remarqué, en particulier, l’installation de Lygeum spartum en affinité avec la texture sablo-limoneuse dans le périmètre d’Atriplex halimus. Ajoutons à cela que la présence et le développement d’Artemisia herba-alba dans le périmètre d’Atriplex canescens où la texture est argilo-limoneuse ont contribué essentiellement à l’accroissement de l’épaisseur du sol et son taux de matière organique. Dans le périmètre non planté, le sol, à faible épaisseur, reste pauvre en éléments nutritifs. Il ne permet que l’installation d’espèces annuelles peu exigeantes en éléments nutritifs et d’espèces épineuses peu palatables, très résistantes aux différents stress climatiques, édaphiques et anthropiques comme Noaea mucronata. Cette dégradation est à l’origine d’une thérophytisation qui caractérise essentiellement les zones arides (Grime, Citation1977; Daget, Citation1980; Amghar, Citation2002).
Conclusion
Les résultats obtenus mettent en lumière l’adaptation du genre Atriplex aux zones steppiques arides de la wilaya de Saïda (Algérie occidentale) et donc le rôle que pourraient jouer ces arbustes fourragers dans l’amélioration des disponibilités fourragères, la valorisation des terres marginales et la conservation des ressources naturelles dans les zones à faible pluviométrie (Chriyaa & El-Mzouri, Citation2004). En effet, cette première évaluation révèle une tendance à l’amélioration des conditions édaphiques dans les périmètres plantés en Atriplex. Le piégeage du sable en particulier par les arbustes d’Atriplex halimus permet l’amélioration de la perméabilité du sol, la diminution de l’évaporation (effet self mulching) et l’augmentation de l’épaisseur du sol. D’autre part, la présence d’Atriplex permet de créer certainement un microclimat en réduisant la vitesse du vent et en procurant de l’ombre permettant aux espèces herbacées qui l’accompagnent d’augmenter leur productivité. Ceci diminue l’évapotranspiration, conserve l’humidité, améliore la fertilité du sol et protège ce dernier contre l’érosion éolienne. Comparativement au périmètre témoin non planté, ces conditions favorables permettent, dans les périmètres plantés en Atriplex, l’installation d’une végétation herbacée avec de meilleurs taux de recouvrement et de production en matière sèche.
La richesse spécifique des trois périmètres est similaire et les espèces inventoriées appartiennent principalement aux Astéracacées et aux Poacées.
En outre, il est à noter l’installation de Lygeum spartum (espèce psammophile) dans le périmètre d’Atriplex halimus, en raison du pourcentage élevé de sable piégé. Alors que dans le périmètre d’Atriplex canescens la texture argilo-limoneuse a favorisé la régénération d’Artemisia herba-alba, dans le périmètre non planté, nous notons la dominance de Noaea mucronata indiquant l’état de dégradation. Par ailleurs, l’évolution des sels dans les sols sous Atriplex doit retenir notre attention.
Enfin, cette évaluation représente des résultats préliminaires contribuant à la caractérisation des premiers stades d’évolution des périmètres analysés, d’où il est important et nécessaire de poursuivre nos investigations afin de retracer la dynamique de ces plantations et d’affirmer ou d’infirmer leurs intérêts dans la remise en valeur de zones pastorales dégradées. Les efforts doivent aussi se multiplier par l’utilisation d’autres espèces et techniques permettant la conservation de la biodiversité et l’amélioration du rendement de ces parcours steppiques et leur exploitation durable.
Related Research Data
References
- Abbad , A. , El-Hadrami , A. and Benchaabane , A. 2004 . Atriplex halimus (Chenopodiaceae): a halophytic species for restoration and rehabilitation of saline degraded lands . Pakistan journal of biological sciences , 7 ( 6 ) : 1085 – 1093 .
- Abdelguerfi , A. and Laouar , M. 2000 . Conséquences des changements sur les ressources génétiques du Maghreb . Rev. Opt. Médit. , 39 : 77 – 87 .
- Aidoud, A. 1983. Contribution à l’étude des écosystèmes steppiques du Sud-Oranais: phytomasse, productivité primaire et applications pastorales. Thèse Doctorat 3éme Cycle, université des sciences et de la technologie Houari Boumédiène d’Alger, 250 p.
- Aidoud , A. , Le Floc’h , E. and Le Houérou , H.N. 2006 . Les steppes arides du nord de l’Afrique . Sécheresse , 17 ( 1 ) : 19 – 30 .
- Aidoud , A. and Touffet , J. 1996 . La régression de l’alfa (Stipa tenacissima L.), graminée pérenne, un indicateur de désertification des steppes algériennes . Sécheresse , 7 ( 3 ) : 187 – 193 .
- Amghar, F. 2002. Contribution à l’étude de la biodiversité de quelques formations de dégradations en Algérie. Thèse de magister, université des sciences et de la technologie Houari Boumédiène d’Alger, 188 p.
- Aubert, G. 1978. Méthodes d’analyses du sol. C.R.D.P., Marseille, 189 p.
- Barbero , M. , Quézel , P. and Loisel , R. 1989 . Perturbations et incendies en région méditerranéenne . Int. Estud. Pyrenaicos Jaca , 12 : 194 – 215 .
- Barbero , M. , Quézel , P. and Loisel , R. 1990 . Les apports de la phytoécologie dans l’interprétation des changements et perturbations induits par l’homme sur les écosystèmes forestiers méditerranéens . Forêt méditerranéenne , 12 : 194 – 215 .
- Belkhodja , M. and Bidai , Y. 2004 . Réponse des graines d’Atriplex halimus L. à la salinité au stade de la germination . Sécheresse , 4 ( 15 ) : 331 – 335 .
- Bernoux, M. 2008. Les matières organiques du sol. Colloque de restitution du programme GESSOL 2 Paris. Les matières organiques du sol: rôles, risques et enjeux.
- Bouchetata Tarik B., and A. Bouchetata Arslan. 2005. Dégradation des écosystèmes steppiques et stratégie de développement durable. Mise au point méthodologique appliquée à la wilaya de Nâama (Algérie). Développement durable et territoires [En ligne]. URL: http://developpementdurable.revues.org/1339.
- Braun-Blanquet, J. 1951-. Pflanzensoziologie (2ten Aufl.). Springer, Vienne, 631 p.
- Chriyaa Abdelouahid , A. and El-Mzouri , H. 2004 . Introduction des arbustes fourragers dans les systèmes de production en zones à faible pluviométrie . Options Méditerranéennes , 62 : 203 – 206 .
- Daget Ph., 1980. Sur les types biologiques en tant que stratégie adaptative. Cas des thérophytes. In: Recherches d’écologie théorique, les stratégies adaptatives, Paris, 89–114.
- Dahmani-Megrourouche M., 1997. Le chêne vert en Algérie, syntaxonomie, phytoécologie et dynamique des peuplements. Thèse de doctorat d’état, université des sciences et de la technologie Houari Boumédiène d’Alger, 330 p.
- Djebaili , S. 1984 . Steppe algérienne, phytosociologie et écologie. Recherche phytosociologique et écologique sur la végétation des hautes plaines steppiques de l’Atlas saharien , Alger : Office des publications universitaires (OPU) .
- Floret , C. , Le Floc’h , E. , Pontanier , R. and Romane , F. 1981 . Dynamique des systèmes écologiques de la zone aride. Application à l’aménagement sur les bases écologiques d’une zone de la Tunisie présaharienne . Acta Oecologica, Oecol. Applic. , 2 ( 3 ) : 195 – 214 .
- Gounot, M. 1969. Méthodes d’études quantitatives de la végétation. 1 vol. Masson et Cie, Paris, 314 p.
- Grime , J.P. 1977 . Evidence for existence of three primary strategies in plants and its relevance to ecological and evolutionary theory . The American Naturalist , 111 : 1169 – 1194 .
- Ighilhariz , Z. , Bouabdallah , L. and Belkhodja , M. 2008 . Influence hormonale sur l’induction de la callogénèse chez Atriplex halimus L. et Atriplex canescens . European Journal of Scientific Research , 24 ( 2 ) : 211 – 218 .
- Kadi-Hanifi, H. 1998. L’alfa en Algérie: syntaxonomie, relation milieu-végétation, dynamique et perspectives d’avenir. Thèse de doctorat d’état, université des sciences et de la technologie Houari Boumédiène d’Alger, 228 p.
- Le Houérou, HN. 1985. La régénération des steppes algériennes. Rapport de mission de consultation et d’évaluation. 42 p.
- Mulas, M., and G. Mulas 2004. Potentialités d’utilisation stratégique des plantes des genres Atriplex et Opuntia dans la lutte contre la désertification. Université des études de SASSARI-Groupe de recherche sur la désertification, 112 p.
- Naggar , M. 2000 . Eléments de base d’une stratégie de sylvopastoralisme en Afrique du Nord . Options Méditerranéennes, Série A , 39 : 191 – 202 .
- Nedjraoui ,D., and S. Bedrani 2008. La désertification dans les steppes algériennes: causes, impacts et actions de lutte. [Vertigo] La revue électronique en sciences de l’environnement, 8 (1).
- Nefzaoui , A. and Chermiti , A. 1991 . Place et rôles des arbustes fourragers dans les parcours des zones arides et semi-arides de la Tunisie . Options Méditerranéennes, Série Séminaires , 16 : 119 – 125 .
- Office national de la météorologie (Saïda-Algérie). Données 2003–2009.
- Phillippeau, G. 1986. Comment interpréter les résultats d’analyse en ACP? Institut technique des céréales et fourrages, Paris, 63 p.
- Pontanier R., A. M’Hiri, J. Aronson, N. Akrimi, and E. Le Floc’h, 1995. L’homme peut-il refaire ce quil a défait? Edition John Libbey Eurotext, Paris, 494 p.
- Pouget, M. 1980. Les relations sol-végétation dans les steppes sud-algéroises. ORSTOM, Paris, 555 p.
- Pujol , J.A. , Calvo , J.F. and Ramirez Diaz , L. 2001 . Seed germination, growth and osmotic adjustment in response to NaCl in a rare succulent halophyte from south eastern Spain . Journal Wetlands , 21 ( 2 ) : 256 – 264 .
- Smail-Saadoun , N. 2005 . Anatomical adaptation of Algerian Sahara Chenopodiaceae to severe drought conditions . Sécheresse , 16 ( 2 ) : 121 – 124 .
- Wael E., S.E. BachirBouiadjra, M. Benslimane, and K. Mederbal, 2009. L’écosystème steppique face à la désertification: cas de la région d’El Bayadh, Algérie. [Vertigo] La revue électronique en sciences de l’environnement, 9 (2).