Résumé
La diversité floristique de deux lots de vergers traditionnels de ± 20 et ± 40 ans est étudiée en fonction du degré de l’activité anthropique. Mis en place en savane, les vergers de ± 20 ans sont entretenus, alors que ceux de ± 40 ans sont abandonnés depuis plus d’une vingtaine d’année. L’inventaire recense 24 espèces dont 21% sont communes aux deux lots de vergers. Les densités moyennes respectives sont de 94 ± 3,46 pieds.ha-1 et 102 ± 2 pieds.ha-1. Les coefficients de similarité de Jaccard et de Sørensen discriminent les deux lots, alors que les indices de diversité de Shannon (0,79 ± 0,38 et 0,93 ± 0,02) et de Simpson (5,63 ± 1,05 et 13,74 ± 1,79) soulignent leur faible diversification. Quant à l’indice de Pielou (0,42 ± 0.17 et 0,46 ± 0.02), il marque une dominance dans les deux cas. Les données phytogéographiques caractérisent l’élément de base Guinéo-Congolais (29,16%), quoique que l’on note une dominance des espèces allochtones (50%) de distribution pantropicale.
Abstract
The floristic diversity of two batches of traditional orchards of ± 20 and ± 40 years were studied according to the degree of anthropogenic activity. Installed in savannah, the orchards of ± 20 years are maintained, whereas those of ± 40 years are abandoned since more than one score of year. The inventory counts 24 species of which 21% are common to both batches of orchards. The respective average densities are of 94 ± 3.46 trees.ha-1 and 102 ± 2 trees.ha-1. The coefficients of similarity of Jaccard and Sørensen discriminate the two batches, whereas the diversity indexes of Shannon (0.79 ± 0.38 and 0.93 ± 0.02) and of Simpson (5.63 ± 1.05 and 13.74 ± 1.79), underline their weak diversification. As for the index of Pielou (0.42 ± 0.17 and 0.46 ± 0.02), it marks a predominance in the two batches of orchards. The phytogeographic data characterise the Guineo-Congolian basic element (29.16%), although the flora is being dominated by the settler species (50%) of pantropical distribution
Introduction
Le Congo, traversé par l’équateur, s’étire entre 4° N - 5° S et 11°-19° E. D’une superficie de 342.000Km2, il est recouvert à environ 65% de forêts et à 35% de savanes. Cette couverture forestière représente environ 12% des forêts du Bassin du Congo, soit environ 10% des forêts tropicales (UICN, Citation1996). La végétation forestière se présente sous la forme de trois grands massifs discontinus se répartissant comme suit: le Mayombe (3%) et le Chaillu (11%) dans le Sud-Ouest, et le massif du Nord Congo (51%) dans la partie septentrionale. Ce dernier, plus diversifié que les deux autres, se subdivise en deux entités écologiques et phytogéographiques basées sur la nature du substrat (Descoings, Citation1975; Kimpouni et al., Citation1992). Il s’agit des forêts de la Haute Sangha (31%) sur la terre ferme et de la Basse Sangha (20%) dominée par la forêt inondée (Descoings, Citation1975; Adjanohoun et al., Citation1988).
Les savanes recouvrent majoritairement le Sud Ouest (la Vallée du Niari et le littoral) et le centre (plateaux Téké) du territoire (Adjanohoun et al., Citation1988; Hecketsweiler, Citation1990). Ces formations, très diversifiées dans leur physionomie et composition floristique se repartissent comme suit: savane du littoral (1%); savane de la Vallée du Niari (8%); savane des plateaux de Cataractes (2%) et savane des plateaux Tékés (24%).
La valeur agro-industrielle et pastorale de la savane de la Vallée du Niari, associée à l’inégale répartition de la population congolaise, fait que cette région est densément peuplée (Vennetier, Citation1977). Si la densité nationale est d’environ 3,8 hab./km2, celle de la Vallée du Niari oscille entre 10 et 18 hab./km2 tandis que la moyenne nationale en zone rurale est d’environ 1,7 hab./km2. Fortement colonisée, la physionomie floristique de la végétation de cette zone est très marquée par l’action anthropique, suite aux implantations humaines et à leurs corollaires qui sont les agroforêts et les jardins de cases (Koechlin, Citation1961; Vennetier, Citation1977; Adjanohoun et al., Citation1988).
Les arbres fruitiers cultivés forment un système agroforestier de type traditionnel qui concour et à la satisfaction des besoins économiques, alimentaires, thérapeutiques et sert occasionnellement de source de bois de chauffage pour les populations. Les études de la flore de ces systèmes ligneux artificiels (jardins de cases et agroforêts) sont très peu nombreuses. Celles qui existent se focalisent sur l’inventaire floristique et les usages ethnobotaniques des plantes (Koussibila & Makaya, Citation1997; Biyoudi, Citation1998). Nonobstant les valeurs ethnobotaniques, reconnues à la flore des systèmes agroforestiers et des jardins de cases, le rôle écologique de ces formations reste presque occulté. En effet, les seuls travaux relatifs à la dynamique floristique des vergers (jardins de cases et agroforêts) en cours d’exploitation et abandonnés, en milieu savanicole sont ceux de Kouvoukana (Citation2008) menés dans le district de Kayes.
Les zones de savanes de la Vallée du Niari étant régulièrement soumises aux feux de brousse tardifs, leurs suites se résument en un appauvrissement de la biodiversité. Ce phénomène est marqué par la sélection d’un petit nombre d’espèces végétales (pyrophytes) adaptées à ces conditions (Trochain, Citation1980; Bergonzini & Lanly, 2000). La mise en place des systèmes agroforestiers en savane est intéressante à plus d’un titre, dans la mesure où elle lutte contre l’effet de serre par l’augmentation du potentiel ligneux et concomitamment la conversion des surfaces moins productives, contribuant ainsi à l’enrichissement et la conservation de la biodiversité spécifique, génétique et écosystémique (Bergonzini & Lanly, 2000; Bergonzini, 2004).
Dans les zones de savane, surtout la Vallée du Niari, les fruitiers constituent un système agroforestier traditionnel autour des villages. Ces écosystèmes anthropiques contrastent fortement avec la physionomie naturelle de la savane dans le sud ouest du Congo (Koechlin, Citation1961; Adjanohoun et al., Citation1988; Hecketsweiler, Citation1990). Suite aux mouvements migratoires et socioéconomiques ayant marqué les populations du district de Kayes au cours des années 1930 (Anonyme, Citation2003; Kouvoukana, Citation2008), plusieurs vergers, naguère entretenus et enrichis en espèces héliophiles généralement, ont été progressivement abandonnés au profit des nouveaux qui jouxtent les villages récents. Une fois le système livré au gré de la nature, la densité des fruitiers baisse et le cortège floristique spontané, à base d’espèces forestières notamment, prend la succession.
La présente étude qui est une contribution à l’amélioration des connaissances de ces écosystèmes anthropiques, se base sur deux lots de vergers de ± 20 et 40 ans et répond à trois hypothèses:
| • | le verger abandonné favorise le développement forestier en zone de savane; | ||||
| • | Les espèces cultivées sont mal adaptées à la compétition interspécifique; | ||||
| • | L’action anthropique ralentit la colonisation des agroforêts par les espèces forestières. | ||||
Présentation du site d’étude
Localisation du site
Les différents vergers étudiés se localisent dans le district de Kayes (région de la Bouenza). Cette localité qui encercle la ville de Nkayi (13° 16’ E, 04° 13’ S) s’étire entre 13° 03’ et 13° 33’ de longitude est et 4° et 4° 30’ de latitude sud (Fig. ).
Fig. 1 Localisation de la zone d’étude. Fig. . Localization of study zone.
Climat
La zone d’étude est sous l’influence du climat bas congolais (Aubréville, Citation1949; Vennetier, Citation1977). Celui-ci est marqué par une température moyenne annuelle d’environ 25 °C et une amplitude thermique comprise entre 4–6 °C et une alternance de deux saisons de durée inégale. La saison de pluies, chaude et humide, s’étend de juin à septembre, avec un ralentissement pluviométrique de janvier à février; la saison sèche, caractérisée par un arrêt de précipitations, est fraîche et s’installe de juillet à septembre. La pluviométrie moyenne est comprise entre 1200 et 1400 mm/an (Adjanohoun et al., Citation1988; Hecketsweiler, Citation1990; U.C.N., Citation1996; Samba et al., Citation2008; Samba & Nganga, Citation2011). Les pics de pluies sont observés au mois de novembre, pour le premier, et entre les mois de mars et d’avril, pour le second.
L’humidité relative est particulièrement élevée et se maintient au dessus de 70%. Le minimum absolu est enregistré entre les mois d’août et de septembre, alors que le minimum relatif est relevé aux mois de février et de mars. Les minima oscillent entre 49 et 59%, pendant que les maxima restent au dessus de 80% et varient de 88 à 90% (Koechlin, Citation1961; Makany, Citation1976; Adjanohoun et al., Citation1988).
L’insolation moyenne mensuelle oscille de 120 à 190 heures, soit une moyenne annuelle comprise entre 1700 et 2000 heures. Les pics d’insolation sont observés au mois d’avril, d’une part, et d’autre part, entre les mois de novembre et de décembre, alors que le minimum intervient en juillet (Koechlin, Citation1961; Samba et al., Citation2008)
Les vents sont généralement faibles et leur vitesse ne dépasse pas les 5 m/s. Exceptionnellement, la zone est balayée par des vents violents de courte durée, dont la vitesse est supérieure à 10 m/s (Koechlin, Citation1961; Vennetier, Citation1977; Adjanohoun et al., Citation1988).
Sols
Les sols appartiennent généralement à la classe des sols ferrallitiques fortement désaturés. Leur teneur en argile varie entre 64 et 70% (Koechlin Citation1961; Trochain, Citation1980; Nzila, Citation1986; Loumeto, Citation2002).
Végétation
La végétation du district de Kayes est essentiellement formée de savane à haute herbe (Hyparrhenia diplandra (Hack.) Stapf) et à strate ligneuse moins fournie. Les principaux ligneux composant la strate arbustive sont Annona senegalensis Pers., Bridelia ferruginea Benth., Psorospermum febrifugum Spach., Vitex madiensis Oliv., Sarcocephalus latifolius (Smith) Bruce, Millettia versicolor Welw. ex Bak.. Les formations boisées se limitent à des étroites forêts galeries dégradées et à des formations anthropiques (Koechlin, Citation1961; Kimpouni, Citation2009).
Hydrographie
Le district de Kayes est arrosé par plusieurs affluents du fleuve Niari qui le traverse d’Ouest en Est. Les principaux affluents sont la Loudima et son bras, la Louamba au Sud-Ouest, la Loango au Nord, la Divouba et la Louadi au Sud. Toutes ces rivières prennent leur source sur le plateau des cataractes, dans le secteur dit, plateau Doondo à l’exception de la Milombo, affluent de la Divouba dont l’écoulement est intermittent, tous les cours d’eau sont permanents. En dehors des rivières et fleuves, on dénombre plusieurs lacs dont les plus grands sont Mbouma, Bodissa et Kimbonga-Louamba (Koechlin, Citation1961; Moukolo, Citation1992).
Matériel et Méthodes
Matériel
Le matériel est constitué de deux lots de vergers âgés de ± 20 et 40 ans. Chaque lot représente une aire de 3 ha, soit trois vergers d’un hectare. La différence entre les deux lots réside dans le fait que les vergers de ± 20 ans sont encore entretenus, alors que ceux de ± 40 ans sont abandonnés depuis plus d’une vingtaine d’années au moins. Le matériel végétal d’inventaire se compose d’arbres, d’arbustes et de lianes. L’identification taxonomique de ce matériel s’est faite in situ pour les espèces couramment rencontréeset, le cas échéant, au Laboratoire de botanique de l’École Normale Supérieure, voire à l’herbier national (IEC) à Brazzaville. Les différents taxons sont ordonnés suivant APG III (2009) et répartis en classe de diamètre suivant la section du tronc à 1,3 m.
Méthodologie
La méthodologie se base sur trois parties complémentaires (la revue de la littérature, l’inventaire floristique et l’entretien oral). La revue de la littérature a permis de faire l’état des lieux des connaissances acquises dans l’étude des vergers traditionnels et des jardins de cases. L’entretien oral est semi directif et à questions ouvertes; il est soit individuel, soit mené en “focus group”. Son intérêt est d’éclairer les questions ethnolinguistiques de la dénomination des plantes et de confirmer le statut des ligneux dans les vergers (plantes cultivées ou spontanées). L’inventaire floristique est mené dans les parcelles de 100 x 50 m, soit 0,5 ha. Le principe est de mesurer tous les ligneux dont le dbh (diameter at breast height) pris à 1,30 m du sol, est supérieur ou égal à 10 cm (Dallmeier, Citation1992; Lejoly, Citation1993). Au cours de cet inventaire, les caractères suivis sont:
| - | le type biologique, notamment phanérophytes au sens large (arbre, arbuste, liane), qui correspond au stade ultime de développement de l’individu; | ||||
| - | le statut de l’espèce (plante cultivée ou spontanée); | ||||
| - | le type phytogéographique pour déterminer l’aire de répartition des spécimens. Il est défini suivant les traits généraux de la phytochorologie africaine (White, Citation1979, Citation1986); les types retenus sont: Afrotropical (Aftr), Centroguinéen (Cg), Guinéo-Congolais (GC), Paléotropical (Paléo), Pantropical (Pantr). | ||||
Les types de diaspores retenus, afin de suivre la dissémination des différents taxons sont en accord avec les catégories définies par Dansereau et Lems (1957) et Évrard (Citation1968). Celui-ci s’agit des ballochores (Ballo): diaspores expulsées par la plante elle-même; des pogonochores (Pogo): diaspores pourvues d’appendices plumeux ou soyeux; des ptérochores (Ptéro): diaspores munies d’appendices aliformes; des sarcochores (Sarco): diaspores totalement ou partiellement charnues.
Dans le traitement des données, les arbres sont regroupés en sept classes de diamètre de 10 cm. La septième classe regroupe les arbres de dbh supérieur ou égal à 70 cm.
Mode d’expression des résultats
| - | la diversité floristique est exprimée par le calcul des spectres bruts et pondérés | ||||
| - | le coefficient de similarité, en utilisant le mode de calcul de Jaccard et de Sørensen | ||||
na = nombre de spécimens du 1er relevé; nb = nombre de spécimens du 2e relevé;
nc = nombre de spécimens communs aux deux relevés.
Le double calcul de coefficients de similarité de Jaccard et de Sørensen se fonde sur la valeur que l’on attribue à la présence et à l’absence. Si le calcul de Jaccard donne la même valeur à la présence et à l’absence, il n’accorde pas assez de poids aux espèces communes (en mettant seulement une fois nc au numérateur et 3 fois nc au dénominateur car, na et nb contiennent 2 fois nc); celui de Sørensen, par contre, donne un avantage deux fois plus important à la présence. Dans ce dernier cas la présence est plus informative que l’absence (Legendre & Legendre, Citation1984a, Citation1984b).
| - | la surface terrière: il s’agit de la surface de la section transversale des troncs d’arbres au niveau de référence de 1,3 m. | ||||
Avec: = constante
; R = rayon du tronc au niveau de référence
| - | Le rapport de la surface terrière (ST) sur la densité (D): indice écologique traduisant la dynamique de l’évolution diamétrique et de la structure de la population, et le changement de la composition floristique. | ||||
| - | les indices de diversité de Shannon (ou Shannon-Weiner) et Simpson. | ||||
Avec: pi = ni/n = nombre d’individus (ni) de l’espèce i divisé par le nombre total (n) d’individus;
S = nombre d’espèces; log = logarithme de base 10 (décimal).
La différence entre les deux méthodes de calcul de l’indice de diversité se fonde sur la valeur accordée aux espèces. L’indice de Simpson penche en faveur des espèces communes, alors que celui de Shannon favorise les espèces rares (Odum, Citation1976). En effet, l’indice de Shannon qui est plus utilisé dans l’analyse de la végétation ne privilégie aucune espèce quel que soit son statut (espèce rare ou abondante), tandis que l’indice de Simpson accorde plus de poids aux espèces abondantes à travers (pi)².
La diversité floristique maximale est calculée par la formule suivante: H’max = log(s). Elle renseigne sur le degré de diversité que peut atteindre une phytocénose dans le cas d’une distribution équitable des individus constitutifs de sa flore.
l’indice d’équitabilité de Pielou (Eq) ou indice de régularité: cet indice est égal au rapport de l’indice de Shannon sur la diversité maximale (H’/H’max). Variant de 0 à 1, il indique le degré de diversité atteint par rapport au maximum possible et exprime mieux la variation intra-communauté. Lorsque la régularité est faible (tend vers 0), elle dénote un phénomène de dominance; cependant, quand elle est élevée (tend vers 1), on note une répartition régulière des individus entre les espèces, d’où l’absence de dominance.
Résultats
Inventaire floristique
L’inventaire des deux lots de vergers a permis de recenser 588 individus de dbh ⩾ 10 cm (Tableau ) dont 282 individus dans les vergers de ± 20 ans et 306 dans les ± 40 ans. Suivant l’APG III (Citation2009), ces individus représentent 24 espèces distribuées dans 16 familles et 23 genres (Tableau ). La diversité des familles varie de 4,17 à 12,5%. En fonction de l’âge des vergers, on relève 7 espèces dans les vergers de ± 20 ans, contre 24 dans ± 40 ans, dont 17 (soit 62,5%) qui y sont strictement associées. Notons que des 7 espèces des vergers de ± 20 ans, 5 sont présentes dans les ± 40 ans. Elles sont les seules espèces communes, inventoriées dans les deux types de formation.
Table 1. Distribution of taxa listed in the orchards and their phytogeographical type Legend: V = orchard; (2/8,33 %) = number of species by family / relative diversity of the family in %. Tableau 1.- Distribution des taxons recensés dans les vergers et leur type phytogéographique Légende : V = verger ; (2/8,33%) = nombre d’espèces par famille/diversité relative de la famille en %.
Table 2. Synoptic of the floristic diversity listed in the orchards. Tableau 2. Synoptique de la diversité floristique recensée dans les vergers.
Analyse de la composition floristique
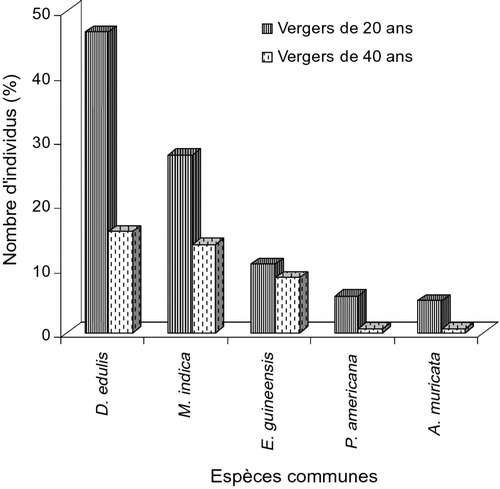
L’inventaire de la flore des deux lots de vergers présente 20,83% d’espèces communes. Ainsi, la diversité floristique caractéristique est de 70,83% pour les vergers de ± 40 ans, contre 8,33% aux ± 20 ans. Une comparaison des densités des espèces communes montre qu’elle est plus élevée dans les vergers de ± 20 ans que dans les ± 40 ans (Fig. ). Quant à la diversité floristique, elle est plus élevée dans les vergers de ± 40 ans que dans les ± 20 ans.
Fig. 2 Comparaison des effectifs des espèces communes dans les deux types de vergers. Fig. 2. Comparison of the common species in the two orchard types.
Variation des caractéristiques écologiques et de la diversité floristique
Les densités (Tableau ) présentent une variation de 92 à 98 pieds.ha-1, soit une moyenne de 94 ± 3,46 pieds.ha-1 dans les vergers de ± 20 ans, contre 100 à 104 pieds.ha-1, soit une moyenne de 102 ± 2 pieds.ha-1.
Table 3. Ecological indices of the various orchards. Tableau 3. Récapitulatif des indices écologiques dans les différents vergers.
La diversité floristique relevée dans les vergers de ± 20 ans est comprise entre 5 et 7 espèces, soit une moyenne de 6 ± 1 espèces.ha-1. Les Anacardiaceae avec 28,55% des espèces inventoriées constituent la famille dominante, alors que les moins représentées ont 14,29% (Planche , Planche 1A), Quant au nombre d’individus, les Burseraceae sont les plus abondantes avec 46,81% de pieds (Planche 2A). Ce faible degré de diversification floristique est corrélé à une forte échelle de sélection, favorisant l’émergence de quelques espèces aux densités modestement élevées. Dans ce cas, il s’agit de Dacryodes edulis (G.Don) H.J.Lam., de Mangifera indica L. et de Elaeis guineensis Jacq.
Planche 1 Évolution comparée des taxons dans les deux lots de vergers. Board 1. Species evolution compared in the two batches of orchards.
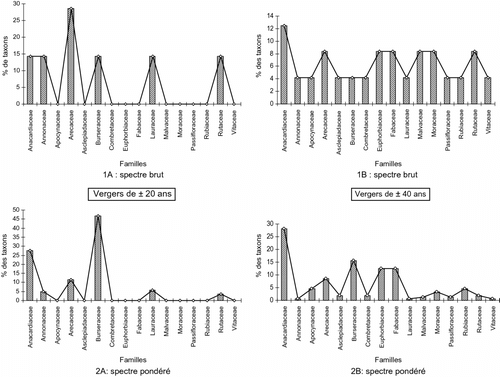
Dans les vergers de ± 40 ans, la diversité floristique par hectare est comprise entre 12 et 15 espèces, soit une moyenne de 14 ± 1,73 espèces.ha-1. La famille la plus diversifiée est celle des Anacardiaceae avec 12,50% d’espèces, et la moins pourvue a 4,17% d’espèces (Planche 1B). Les Anacardiaceae avec 28,10% de pieds sont les plus présentes à ce stade de développement (Planche 2B). Il est noté dans ces vergers un accroissement de la diversité floristique, sans réelle dominance de taxons et une baisse des densités des espèces autrefois dominantes. Ce système agroforestier a une composition floristique composite favorisée par deux raisons, la cessation de la sélection d’une part, et d’autre part les modifications écologiques du milieu dont le corollaire est l’envahissement du système par une flore spontanée, mieux adaptée et plus compétitive.
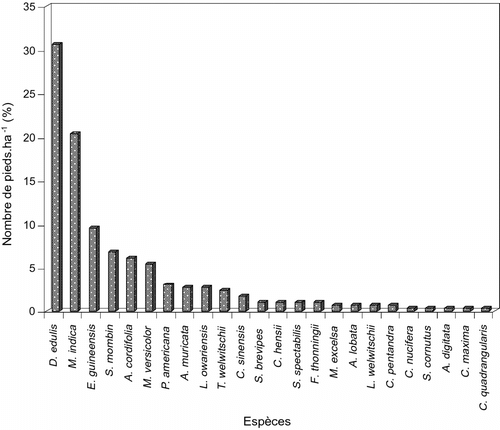
L’inventaire dans son ensemble montre que Dacryodes edulis est dominante avec 30,61% de spécimens (Fig. ). Les espèces les moins représentées ont une contribution de 0,34%. Au niveau spécifique, le nombre de pieds des espèces dominantes, pour chacun de lot, se présente comme suit:
| • | Vergers de ± 20 ans: Dacryodes edulis avec 46,81% d’individus est la plus représentée, alors que Cocos nucifera est la moins représentée avec 0,70% de pieds. | ||||
| • | Vergers de ± 40ans: Dacryodes edulis avec 15,62% de pieds est dominante, contre 0,65% de pieds pour les moins représentées. | ||||
Fig. 3 Spectre de la densité des ligneux des deux lots de vergers. Fig. 3. Spectrum of the density of the ligneous family of the two batches of orchards.
Structure diamétrique et occupation spatiale
Distribution des taxons en classes de diamètre
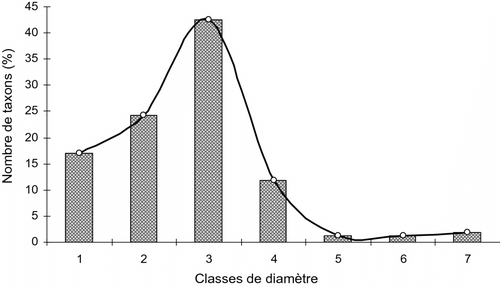
L’étude de la structure diamétrique globale de l’inventaire indique qu’environ 2/3 des 588 individus se retrouvent dans les classes de diamètre de 20 et 30 cm. Elles représentent ensemble une contribution de ± 65,31%. Toutes les autres classes réunies ont une contribution de 34,69%. Suivant l’âge des vergers, cette étude ordonne les ligneux des vergers de ± 20 ans en 5 classes de diamètre, avec une nette prépondérance des classes de 20 et 30 cm. Ces deux classes regroupent environ 64% de l’effectif (Fig. ). Quant aux vergers de ± 40 ans, cette analyse identifie 7 classes de diamètre avec une meilleure représentation de la classe 3 qui compte 42,5% de l’inventaire (Fig. ). Cette courbe traduit l’évolution de la flore et montre le changement en cours dans la composition floristique du système.
Fig. 4 Distribution des individus inventoriés dans les vergers de ± 20 ans en classe de diamètre. Fig. 4. Distribution in diameter class of the individuals inventoried in the orchards of ± 20 years.
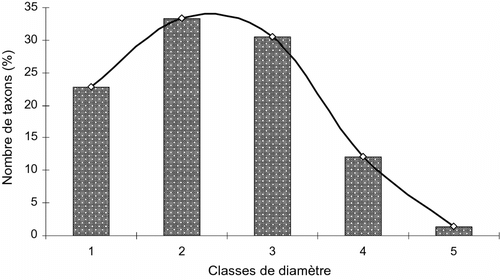
Fig. 5 Distribution des individus inventoriés dans les vergers de ± 40 ans en classe de diamètre. Fig. 5. Distribution in diameter class of the individuals inventoried in the orchards of ± 4 0 years.
Variation de la surface terrière
La surface terrière des vergers de ± 20 ans varie de 6 à 7 m².ha-1, soit une moyenne de 6,66 ± 0,67 m².ha-1. Quant aux vergers de ± 40 ans, elle varie de 8 à 11,4 m².ha-1, soit 9,8 ± 1,70 m².ha-1 en moyenne (Tableau ).
Données écologiques et floristiques
Dissémination des taxons
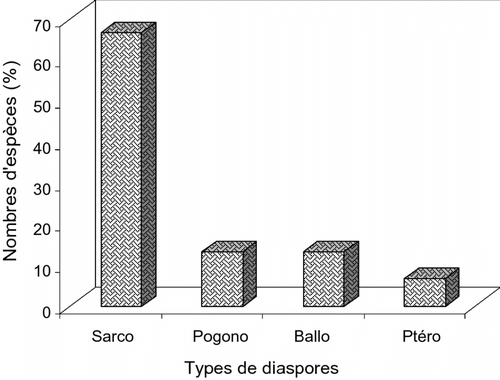
Sur l’ensemble des d’espèces ligneuses spécifiques recensées dans les vergers de ± 40 ans, on dénombre 4 types de diaspores. Parmi lesquels, les sarcochores sont prépondérantes (Fig. ) d’où la zoochorie se trouve être le principal mode de dissémination avec 66,67% de l’effectif total. Les autres formes (ballochorie, pogonochorie et ptérochorie) réunies contribuent à hauteur de 33,33%.
Fig. 6 Distribution des types de diaspores des espèces spécifiques aux vergers de ± 40 ans. Fig. 6. Distribution of diaspores types of specific species to the orchards of ± 40 years.
Indices de diversité floristique
Les indices moyens de diversité floristique de Shannon et de Simpson pour les vergers de ± 20 ans sont respectivement de 0,79 ± 0,38 et de 5,63 ± 1,05. Quant aux vergers de ± 40 ans, ils sont de 0,93 ± 0,02 et de 13,74 ± 1,79. Dans les deux cas, la diversité maximale moyenne atteinte par le système est de 1,97 ± 0,01 et de 2 ± 0,01. Enfin, le coefficient moyen d’équitabilité (régularité) des deux lots de vergers est respectivement de 0,42 ± 0,170 et de 0,46 ± 0,02 (Tableau ). Dans les deux types de vergers, la composition floristique présente une nette dominance.
Table 4. - Variations of floristic diversity in the two batches of orchards. Tableau 4. Variations des indices de diversité floristique dans les deux lots de vergers.
Coefficients de similarité entre vergers
Le coefficient de similarité entre les vergers (Tableau ) distingue deux lots basés sur la composition floristique en fonction de l’âge. Les vergers de même âge sont semblables entre eux, ceci quel que soit le coefficient de similarité (Jaccard ou Sørensen).
Table 5. Correlation of similarity between orchards Tableau 5. Corrélation de similarité entre vergers
Analyse morphologique et phytogéographique
Données morphologiques
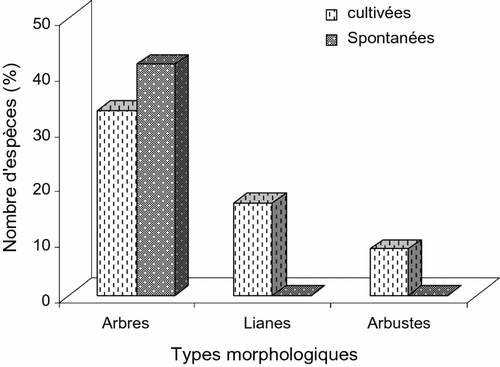
Le type morphologique dans les deux lots caractérise les taxons recensés en trois groupes: les arbres, les arbustes et les lianes (Fig. ). Les proportions des effectifs montrent que les arbres (75%) constituent le type morphologique dominant devant les arbustes (8,33%). Sur les 24 taxons recensés, les plantes cultivées représentent 33,33%, soit 8 espèces (Dacryodes edulis, Mangifera indica, Citrus maxima (Burm.) Merrill, Citrus sinensis Osbeck, Persea americana Mill., Annona muricata L., Elaeis guineensis et Cocos nucifera L.).
Fig. 7 Distribution des taxons inventoriés suivant les types morphologiques. Fig. 7. Distribution of taxa inventoried according to the morphological types.
Données phytogéographiques
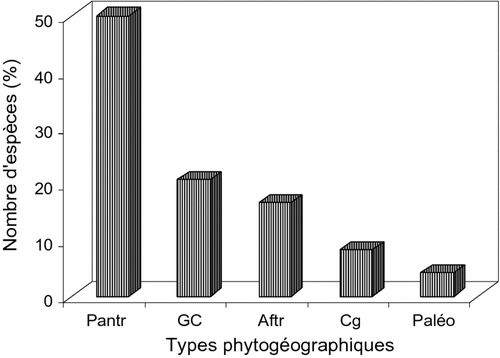
L’analyse phytogéographique de la flore des vergers (Fig. ) identifie 5 éléments phytogéographiques d’importance inégale. L’élément pantropical (50%) est le mieux représenté alors que le paléotropical est marqué par une très faible contribution (4,17% de l’effectif).
Fig. 8 Répartition phytogéographique des différentes espèces recensées. Fig. 8. Phytogeographical distribution of the listed species.
Discussions
Évolution floristique
L’analyse de la composition floristique des vergers marque un net enrichissement des vergers de ± 40 ans, en espèces spontanées d’origine forestière. Ces systèmes qui se développent dans la savane où la strate ligneuse est formée de Sarcocephalus latifolius, Bridelia ferruginea, Psorospermum febrifugum, Ficus sp., Vitex madiensis et Annona senegalensis, jouent un rôle écologique majeur dans la contrée. En effet, la savane étant soumise systématiquement aux feux de brousse pendant la saison sèche, les vergers abandonnés, voire actuels servent de refuge aux animaux vertébrés. Cette migration des animaux serait le fondement de leur enrichissement, car elle s’accompagne d’une dissémination importante des diaspores. Les taxons enregistrés comme exclusifs aux vergers de ± 40 ans sont dominés par les espèces sarcochores dont la dissémination est principalement endozoochore. Cette caractéristique montre une dynamique évolutive les rattachant aux forêts d’Afrique Centrale où généralement la proportion des plantes sarcochores est supérieure à 50% (Sonke, Citation1988). Ces faits symbolisent les liens de co-évolution et de mutualisme existant entre les plantes et les animaux vertébrés (Puig, Citation2001). Concomitamment à cet enrichissement, on observe une diminution des densités des espèces cultivées. Cette baisse du nombre d’individus s’explique par l’inadaptation des plantes cultivées, à tendance héliophile, à la compétition interspécifique engendrée par les nouvelles conditions du milieu. On note, au cours du temps, un remplacement de la flore cultivée par une flore spontanée mieux adaptée aux conditions écologiques qu’offrent les vergers (De Namur, 1978; Kimpouni et al., Citation2008a, Citation2008b).
Analyse des données écologiques
L’analyse des caractéristiques écologiques de la flore montre que les vergers de ± 40 ans ont une hétérogénéité structurale et une complexité fonctionnelle supérieures aux vergers de ± 20. De facto, ils présentent une meilleure résilience en cas de perturbation majeure du milieu [Bond & Chase, Citation2002; Giller & O’Donovan, Citation2002; Caldeira et al., Citation2005; Wallington et al., Citation2005; Mikkelson (sous presse)]. Comparativement aux données écologiques connues en forêt tropicale, notamment sur la densité et la surface terrière (23 à 50 m2.ha-1), cette donnée écologique montre que les écosystèmes étudiés sont foncièrement distincts et encore jeunes (Huetz de lemps, Citation1970; Schnell, Citation1971, Citation1977; Trochain, Citation1980; Ozenda, Citation1982; Mosango, Citation1991; Puig, Citation2001; Kimpouni et al., Citation2008a, Citation2008b). Cependant, la distribution des différents taxons en classes de diamètre relève un recrutement insuffisant au sein de celles-ci, surtout pour les plantes cultivées (Peters, Citation1997; Kpérkouma et al., Citation2005). Elle traduit aussi une régénération insuffisante de ces taxons dans ces écosystèmes artificiels. Ces résultats expliquent le comportement héliophiles des espèces cultivées, et leur incapacité à se développer sous couvert végétal. La courbe de la Fig. marque non seulement l’évolution de la flore, mais aussi le changement en cours dans la composition floristique du système. En tenant compte des données sur la densité, de la croissance en épaisseur des arbres tropicaux qui est d’environ 1 cm/an (Normand, Citation1971; Bedel et al., 1998; Fargeot et al., Citation2004; Fuhr et al., Citation2001) et de la dominance des classes de diamètre 2 et 3 dans les vergers de ± 20 ans et de la classe 3 dans les vergers de ± 40 ans, on relève que ceux-ci sont en harmonie avec l’âge des deux lots de vergers. La faible représentativité des individus de plus petit diamètre s’explique par l’effet conjugué des phénomènes d’entretiens et du ramassage systématique des fruits. Dans ce système, la régénération se fait très mal à cause du contrôle strict des densités des arbres utiles d’une part; et d’autre part, le ramassage systématique des produits induit une réduction de la réserve séminale du sol.
Le rapport de la surface terrière (ST) sur la densité (D) est un indice écologique mettant en évidence les individus les moins représentés dans l’inventaire. Ce rapport qui ne caractérise pas seulement les taxons de gros diamètre, permet aussi de confirmer la dynamique de l’évolution diamétrique, d’une part, et, la dynamique de la structure de la population et le changement de la composition floristique, d’autre part (Doumenge, Citation1992). C’est ainsi qu’il constitue un indicateur de la succession et de l’évolution des ligneux. Les valeurs de cet indice étant inférieur à 1, surtout dans les vergers abandonnés, montrent une nette prépondérance des individus de faible diamètre issus des apports spontanés (Tableau ), cette phase correspondant à l’installation d’un important cortège d’espèces forestières dans les vergers abandonnés. Elle représente le signe majeur du remplacement des ligneux cultivés par une population spontanée, mieux adaptée aux nouvelles conditions du milieu (De Namur, 1978; Doumenge, Citation1992).
La dominance de la sarcochorie dénote une dissémination des diaspores sous la dépendance des animaux, notamment des vertébrés, au sein du système de ± 40 ans. Cette observation sous-tend un haut degré de diversité floristique du système (Puig, Citation2001). Les autres modes de dissémination ne jouant qu’un rôle mineur, le système de ± 40 ans présente une dynamique évolutive assez avancée. En effet, aux espèces pogonochores et ptérochores est associée une dissémination anémochore, caractéristique des essences pionnières des systèmes jeunes (Mosango, Citation1991; Puig, Citation2001).
Au plan phytogéographique, si l’on soustrait l’élément pantropical centré presque exclusivement sur les espèces allochtones dont certaines devenues subspontanées, l’élément de base Guinéo-Congolais est dominant dans les forêts galeries de la contrée (Kimpouni, Citation2008, Citation2009). En effet, l’analyse des nouveaux taxons montre que la conversion est en faveur de l’élément de base Guinéo-Congolais (29,16%), Cette observation montre les tendances évolutives futures de la flore et de ces écosystèmes.
En matière de gestion et d’aménagement des vergers et des types de végétation du milieu d’étude, on retiendra la grande variété des services environnementaux assurés par les forêts par rapport à la savane, pour une exploitation durable et rationnelle. Ces services se scindent en deux ordres qualifiés de directs et d’indirects (Convery, Citation1990). Les fonctions régulatrices se résument au climat, à la protection des sols contre les érosions, au maintien de la biodiversité, à la séquestration du carbone et au recyclage de la matière organique. Elles sont le support des activités économiques et du bien-être humain (Bergonzini & Lanly, Citation2000). La fonction productrice cependant, met en valeur les ressources de base, notamment les matériaux de construction, le bois énergie, les ressources alimentaires et phytopharmaceutiques, tant végétales qu’animales (produits forestiers non ligneux), et génétiques. Enfin, la fonction récréative, cultuelle, éducationnelle et scientifique est plus enrichissante et valorisée (Cunningham, Citation1997; Bergonzini, Citation2004).
Les enjeux planétaires de la question environnementale, notamment du réchauffement climatique et ses conséquences, la conversion partielle des terres savanicoles entretenues régulièrement par les feux de brousse en « écosystème forestier », par le biais des vergers et jardins de cases présente des atouts indéniables dans l’approche des solutions. On passe d’un écosystème naturel moins productif (la savane) à un type forestier plus rentable (Bergonzini & Lanly, 2000; Riedacker, Citation2004). Ainsi, un nouveau regard devrait être porté sur la valeur de ces écosystèmes (agroforêts), en matière de conservation de la biodiversité, d’une part, et, d’autre part, sur la problématique de la gestion des feux de brousse dans la savane (Hecketsweiler, Citation1990).
Conclusion
L’analyse de l’évolution floristique des vergers de ± 20 et 40 ans montre que les espèces spontanées prennent le dessus sur les cultivées. L’absence detaxons spontanés dans les vergers de ± 20 ans est liée à l’action anthropique. Ces espèces cultivées (Dacryodes edulis, Persea americana, Elaeis guineensis,…) à tempérament héliophile subissent la concurrence des espèces spontanées, car non adaptées à compétition interspécifiques et aux nouvelles conditions écologiques. Ainsi, les fourrés anthropiques (agroforêts) préparent les conditions d’installation des espèces forestières en milieu savanicole. Le développement et l’évolution des espèces mieux adaptées aux nouvelles conditions du milieu et aussi plus compétitives induit une diminution des densités des espèces cultivées au profit des spontanées.
Au stade actuel des connaissances sur la dynamique floristique des vergers abandonnés conduisant aux bosquets forestiers dans la savane, un nouveau regard devra être porté sur la gestion de ces écosystèmes. En effet, les bénéfices directs et indirects que l’on tire de leur du développement ont des implications d’ordre privé et communautaire.
Bibliographie
- Adjanohoun E.J., A.M.R. Ahyi, L. Ake Asi, J. Baniakina, P. Chibon, G. Cusset, V. Doulou, A. Enzanza, J. Eyme, E. Goudote, A. Keita, C. Mbemba, J. Mollet, J.-M. Moutsambote, J. Mpati & P. Sita, 1988. Contribution aux études ethnobotaniques et floristiques en République Populaire du Congo: Médecine traditionnelle et pharmacopée. ACCT, Paris, 605 p.
- Anonyme, 2003. Historique du district de Kayes. District de Kayes, 3 p.
- Apg , I.I.I. 2009 . An update of the Angiosperm phylogeny group classification for the orders and families of flowering plants: APG III . Bot. J. Linn. Soc. , 161 : 105 – 121 .
- Aubréville A., 1949. Climats, forêts et désertification de l’Afrique tropicale. Société d’Éditions Géographiques, Maritimes et Coloniales, paris, 351 p.
- Bedel F., L. Durrieu De Madron, B. Dupuy, V. Favrichon, H.-F. Maitre, A. Bar Hen & P. Narboni, 1998. Dynamique de croissance dans des peuplements exploités et éclaircis de forêt dense africaine: le dispositif de M’Baiki en République centrafricaine (1982–1995). Série Forafri n° 1, Cirad-forêt, Cifor, Montpellier, France, 72 p.
- Bergonzini J.-C., 2004. Changements climatiques, désertification, diversité biologique et forêts. Silva, RIAT, 146 p.
- Bergonzini J.-C. & J.-P. Lanly, 2000. Les forêts tropicales. Karthala, CIRAD, 164 p.
- Biyoudi A., 1998. Étude sur les jardins de case de Kinkala. Mémoire CAPES, ENS/UMNG, Brazzaville, 49 p.
- Bond , E.M. and Chase , J.M. 2002 . Biodiversity and ecosystem functioning at local and regional spatial scales . Ecology Letters , 5 : 467 – 470 .
- Caldeira , M.C. , Hector , A. , Loreau , M. and Pereira , J.S. 2005 . Species richness, temporal variability and resistance of biomass production in a Mediterranean grassland . Oikos , 110 : 115 – 123 .
- Convery F.J., 1990. Intégration d’un processus de plan national d’action pour l’environnement dans un cadre macro-économique. In: Plans nationaux d’action pour l’environnement en Afrique, Compte rendu atelier du 13–14 décembre 1990, Gouvernement Irlandais, Institut pour l’environnement (University College, Dublin), Banque Mondiale (EDIAR et AFTEN), pp. 33–47.
- Cunningham , A.B. 1997 . Revue de la documentation ethnobotanique relative à l’Afrique orientale et australe . Bulletin Réseau Africain d’Ethnobotanique , 1 : 23 – 87 .
- Dallmeier F., 1992. Long-term Monotoring of Biological Diversity in Tropical Forest Areas, Methods for Establishment and Inventory of Permanent Plots. MAB Digest 11, UNESCO, Paris, 112 p.
- Dansereau P. & K. Lems, 1957. The grading of dispersal types in plant communities and their ecological significance. Contr. Inst. Bot. Univ. Montreal n° 71, 52 p.
- Descoings , B. 1975 . Les grandes régions naturelles du Congo . Candollea , 30 : 92 – 120 .
- Doumenge C., 1992. La réserve de Conkouati: Congo, le secteur Sud-ouest. UICN, Chevron, Gland-Suisse, 213 p.
- Évrard C., 1968. Recherches écologiques sur le peuplement forestier des sols hydromorphes de la Cuvette centrale congolaise. Publ. INEAC n° 110, 295 p.
- Fargeot , C. , Forni , É. and Nasi , R. 2004 . Réflexions sur l’aménagement des forêts de production dans le bassin du Congo . Bois et Forêts Des Tropiques , 281 ( 3 ) : 19 – 34 .
- Fuhr , M. , Nasi , R. and Délègue , A.-M. 2001 . Vegetation structure, floristic composition and growth characteristics of Aucoumea klaineana Pierre stands as influenced by stand age and thinning . Forest Ecology and Management , 14 : 117 – 132 .
- Giller , P.S. and O’Donovan , G. 2002 . Biodiversity and ecosystem function: do species matter? . Biology and Environment, Proceeding of the Royal Irish Academy , 1028 ( 3 ) : 129 – 139 .
- Hecketsweiler P., 1990. La conservation des écosystèmes forestiers du Congo. UICN, Gland-Suisse, 187 p.
- Huetz de Lemps , A. 1970 . La végétation de la Terre , Paris : Masson & Cie . 132
- Kimpouni , V. 2008 . Premières données sur la diversité floristique de la forêt d’Aubeville (Congo – Brazzaville) . Syst. Geogr. Plants , 78 : 47 – 62 .
- Kimpouni , V. 2009 . Contribution à l’inventaire et à l’analyse de la flore ligneuse du plateau des Cataractes (Congo – Brazzaville) . Acta Bot. Gallica , 156 ( 2 ) : 233 – 244 .
- Kimpouni , V. , Lejoly , J. and Lisowski , S. 1992 . Les Eriocaulaceae du Congo . Fragm. Flor. Geobot. , 37 : 127 – 145 .
- Kimpouni , V. , Loumeto , J. and Mizingou , J. 2008 . Diversité du peuplement ligneux de la forêt monodominante à Aucoumea klaineana (okoumé) du littoral congolais . Annales Université Marien Ngouabi , 9 ( N° spécial ) : 9 – 21 .
- Kimpouni , V. , Loumeto , J. and Mizingou , J. 2008 . Diversité floristique du faciès forestier à Aucoumea klaineana (Okoumé) du littoral congolais . Acta Bot. Gallica , 155 ( 3 ) : 323 – 334 .
- Koechlin J., 1961. La végétation des savanes dans le sud de la République du Congo. IRSC- ORSTOM, 337 p.
- Koussibila Dibansa G.M. & D. Makaya, 1997. Contribution à l’étude de la flore des jardins de case de Brazzaville. Mémoire CAPES, ENS/UMNG, Brazzaville, 47 p.
- Kouvoukana V., 2008. Chronoséquence et dynamique floristique des vergers traditionnels dans le district de Kayes. Mém. CAPES-SN, ENS/UMNG, 47 p.
- Kpérkouma Wala , B. , Sinsin , G. , Kudzo Atsu , Kouami Kokou and Koffi Akpagana . 2005 . Typologie et structure des parcs agroforestiers dans la préfecture de Doulelgou (Togo) . Science et changements planétaires / sécheresse , 16 ( 3 ) : 209 – 216 .
- Legendre L. & P. Legendre, 1984a. Écologie numérique. 1. Le traitement multiple des données écologiques. Masson, Paris & Les Presses de l’Université du Québec, Montréal, 260 p.
- Legendre L. & P. Legendre, 1984b. Écologie numérique. 2. La structure des données écologiques. Masson, Paris & Les Presses de l’Université du Québec, Montréal, 335 p.
- Lejoly J., 1993. Méthodologie pour les inventaires forestiers (partie flore et végétation). AGRECO-CTFT, Bruxelles, 57 p.
- Loumeto J.J., 2002. Le système litière des forêts à peuplements d’okoumé (Aucoumea klaineana). Exemples de deux sites congolais: la forêt du Chaillu et la forêt du littoral. Thèse de Doctorat, Université de Paris VI, 159 p.
- Makany L., 1976. Végétation des plateaux Teke (Congo). Coll. Travaux Université de Brazzaville, 301 p.
- Mikkelson G.M. (sous presse). Diversity–stability hypothesis. (Accessible au http://www.arts.mcgill.ca/philo/mikkelson/dsh.pdf).
- Moukolo N., 1992. Hydrogéologie du Congo. Document BRGM n° 210, 112p.
- Mosango , M. 1991 . Contribution à l’étude botanique et géochimique de l’écosystème forêt en région équatoriale (île Kongolo – Zaïre) . Belg. Journ. Bot. , 124 ( 2 ) : 167 – 194 .
- Namur (De) , C. 1978 . Quelques caractéristiques du développement d’un peuplement ligneux au cours d’une succession secondaire . Cah. ORSTOM, sér. Biol. , XIII ( 3 ) : 211 – 221 .
- Normand D., 1971. Forêts et bois tropicaux. Que sais-je ? Presse Universitaire, France, 127 p.
- Nzila J.D.D., 1986. Étude expérimentale de l’effet des amendements calcaires sur un sol ferrallitique acide de la vallée du Niari. (Congo). Mémoire de DEA fédéral de Pédologie (Sciences des sols). INA-PG, Université Paris VI, 86 p.
- Odum E.P., 1976. Écologie. Doin, Paris, 257 p.
- Ozenda P., 1982. Les végétaux dans la biosphère. Doin, Paris, 432 p.
- Peters C.M., 1997. Exploitation soutenue de produits forestiers autres que le bois en forêt tropicale humide: Manuel d’initiation écologique. Programme d’appui à la biodiversité n° 2, USAID, Biodiversity Support Program, Washington, 49 p.
- Puig H., 2001. La forêt tropicale et humide. Belin, Paris, 334 p.
- Riedacker A., 2004. Changements climatiques et forêts. Silva, RIAT, 231 p.
- Samba G. & Nganga D., 2011. Rainfall variability in Congo-Brazzaville: 1932–2007. Int. J. Climatolog, DOI; 10.1002/joc.2311.
- Samba , G. , Nganga , D. and Mpounza , M. 2008 . Rainfall and temperature variations over Congo-Brazzaville betwen 1950 and 1998 . Theor. Appl. Climatol. , 91 : 85 – 97 .
- Samba Kimbata M.-J., 1978. Le climat du bas-Congo, Thèse Doctorat, Université de Dijon, 280 p.
- Schnell R., 1971. Introduction à la phytogéographie des pays tropicaux. Problèmes généraux (vol. 2: les milieux, les groupements végétaux). Gauthier-Villars, Paris, 425 p.
- Schnell R., 1977. La flore et la végétation d’Afrique tropicale. Gauthier-Villars, Paris, 837 p.
- Sonke B., 1998. Étude floristique et structurale des forêts de la réserve de faune du Dja (Cameroun). Thèse de doctorat, Université Libre de Bruxelles, Bruxelles, 256 p.
- Trochain J.L., 1980. Écologie végétale de la zone intertropicale non désertique. Université Paul Sabatier, Toulouse, 439 p.
- U.I.C.N., 1996. L’atlas pour la conservation des forêts tropicales d’Afrique. UICN, Gland, Suisse, pp. 143–150.
- Vennetier P., 1977. Atlas de la République Populaire du Congo. Éditions Jeune Afrique, 64 p.
- Wallington T.J., R.J. Hobbs & S.A. Moore, 2005. Implications of current ecological thinking for biodiversity conservation: a review of the salient issues. Ecology and Society, 10 (1), 1-15 (online) URL. http://www.ecologyandsociety.org/vol10/iss1/art15/
- White , F. 1979 . The Guineo-congolian region and its relationships to other phytochoria . Bull. Jard. Bot. Nat. Belg. , 49 ( 1/2 ) : 11 – 55 .
- White F., 1986. La végétation de l’Afrique. Mémoire accompagnant la carte de végétation de l’Afrique. UNESCO/AETFAT/UNSO, ORSTOM, Paris, 384 p.