Abstract
Festuca paniculata communities in Valle d’Aosta (N.W. Italy) can contribute to the definition of the geographic boundaries between the western Alps phytosociological associations, due to their intermediate position between Insubrian and inner-alpine communities. In order to describe the phytosociology and syntaxonomy of F. paniculata communities in the study area, 17 original relevés data were analyzed. The communities were referred to Junipero-Festucetum spadiceae juniperetosum nanae, previously described in the Meje-Ecrins-Pelvoux Massif (France). Our results exclude the presence in Valle d’Aosta of Centaureo uniflorae-Festucetum spadiceae, supposed by previous works, and define in Colma di Mombarone (Biella and Torino districts) the western boundary of the Insubrian association Polygalo chamaebuxi-Festucetum paniculatae. The border between Junipero-Festucetum spadiceae and Polygalo chamaebuxi-Festucetum paniculatae is in accordance to differences in continentality conditions, that divide the outer areas of the Insubrian region from the subcontinental and continental zones of Valle d’Aosta.
Résumé
Les communautés à Festuca paniculata en val d’Aoste (nord ouest de l’Italie) jouent un rôle intéressant dans la définition des limites géographiques entre les associations des Alpes occidentales en raison de leur position intermédiaire entre les communautés insubriennes et intra-alpines. Afin de décrire la phytosociologie et la syntaxonomie des communautés à F. paniculata dans la zone d’étude, les données de 17 relevés ont été analysées. Les communautés ont été attribuées au Junipero-Festucetum spadiceae juniperetosum nanae, précédemment décrit dans le massif des Écrins-Mèje-Pelvoux (France). Les résultats excluent la présence en val d’Aoste du Centaureo uniflorae-Festucetum spadiceae, supposée par des travaux précédents, et définit la limite ouest des associations insubriennes du Polygalo chamaebuxi-Festucetum paniculatae à Colma di Mombarone (provinces de Biella et Turin). La limite entre Junipero-Festucetum spadiceae et Polygalo chamaebuxi-Festucetum paniculatae est conforme aux différences des conditions continentales, qui divisent les zones périphériques de la région insubrienne avec les zones sous-continentales et continentales du val d’Aoste.
Introduction
Festuca paniculata (L.) Schinz. & Thell. communities are extensively used grasslands occurring in a large part of mountain regions of southern Europe (Sburlino et al., Citation1996). In the Alps, four associations are actually distinguished within Festucion variae (Vittoz et al., Citation2005; Lonati & Siniscalco, Citation2006) (Fig ): (i) Centaureo uniflorae-Festucetum spadiceae, widespread in the Ligurian, Marittime, Cottian and Graian Alps (W.-Alps, France and Italy), (ii) Junipero nanae-Festucetum spadiceae, limited in the Meje-Ecrins-Pelvoux Massif (W.-Alps, France), (iii) Polygalo chamaebuxi-Festucetum paniculatae, described in the Insubrian Pennine and Lepontine Alps (W.-Alps, Switzerland and Italy), and (iv) Hypochaerido uniflorae-Festucetum paniculatae, confined in the eastern Alps (Austria and Italy).
Fig. 1 Distribution range of Festuca paniculata associations in the Alps. Fig. 1. Distribution des associations à Festuca paniculata dans les Alpes.
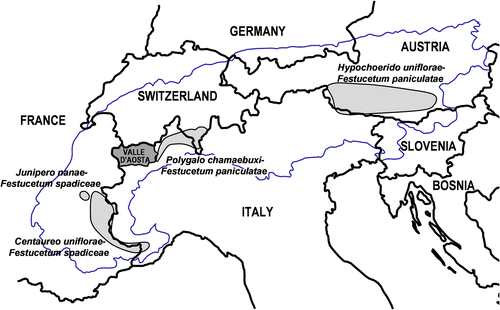
Polygalo chamaebuxi-Festucetum paniculatae (including Hypochaerido uniflorae-Festucetum paniculatae polygaletosum chamaebuxi) was recently described in the subatlantic Insubrian area, characterized by high summer precipitations and mild winter temperatures (Vittoz et al., Citation2005; Lonati & Siniscalco Citation2006). The western limit of the association and the border between Polygalo chamaebuxi-Festucetum paniculatae and Centaureo uniflorae-Festucetum spadiceae are actually unknown. The western range of Polygalo chamaebuxi-Festucetum paniculatae was described at Colma di Mombarone, between Biella and Torino districts (Lonati & Siniscalco, Citation2006) (Fig. ). This association could probably spread eastwards to the Dora Baltea River, that divides the Valle d’Aosta region into its right (Lepontine Alps) and left sides (Graian Alps). This statement agrees with the single phytosociological survey reported by Kaplan (Citation1983) from the Gran Paradiso massif (left side of the Valle d’Aosta), referred by the same author to Centaureo uniflorae-Festucetum spadiceae.
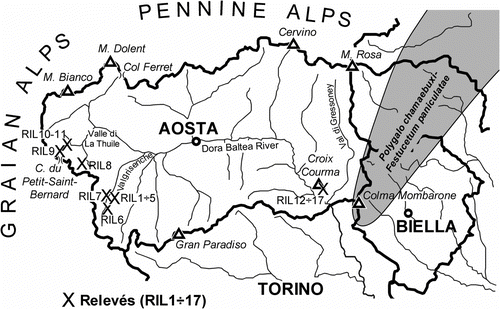
Fig. 2 Study area and location of relevés. The grey area defines the border of the Polygalo chamaebuxi-Festucetum paniculatae. Fig. 2. Zone d’étude et emplacement des relevés. La zone grise définit les limites du Polygalo chamaebuxi-Festucetum paniculatae.
Festuca paniculata communities are not very frequent in Valle d’Aosta. The species is reported in bib-liography in the inner valley (Kaplan, Citation1983; Buffa & Dal Vesco, Citation1988; Revel et al., Citation2001). Many unknown populations were recently observed by M. Bovio (personal communication) both in the inner (Valle di La Thuile, Valgrisenche) and in the outer Valle d’Aosta (Val di Gressoney). Except for the previously mentioned single relevés of Kaplan (Citation1983), no other syntaxonomical data are available for Valle d’Aosta. The attribution to Centaureo uniflorae-Festucetum spadiceae is uncertain: in a recent comparison of alpine Festuca paniculata associations, Vittoz et al. (Citation2005) highlighted a great floristic similarity between Kaplan’s survey and Junipero-Festucetum spadiceae.
On these bases, a phytosociological study was conducted in Valle d’Aosta in order to (i) describe the floristic composition and the syntaxonomy of F. paniculata communities and (ii) clarify the geographical and ecological borders between Polygalo chamaebuxi-Festucetum paniculatae and the other associations growing in the study area.
Material and methods
Relevés were carried out according to the phytosociological method, using the abundance-dominance values proposed by Braun-Blanquet (Citation1932). Site characteristics such as geographical position, morphological and topographical features (altitude, aspect, slope), evidence of current and past management, and soil and water conditions were also logged.
Phytosociological data were transformed into numerical values according to van der Maarel (Citation1979) and a total of 17 relevés were classified by cluster analysis (average link, similarity ratio). Cluster analysis was performed using Syn-Tax V statistical package (Podani, Citation1995).
To compare the differences in climatic conditions between the studied communities and the Insubrian Polygalo chamaebuxi-Festucetum paniculatae, the Gams’ continentality index (Gams, Citation1931, Citation1932a, b) was calculated for each survey. Annual precipitations for the study area were assigned according to data reported in Mercalli et al. (Citation2003). Annual precipitations for the Polygalo chamaebuxi-Festucetum paniculatae surveys (Vittoz et al., Citation2005; Lonati & Siniscalco Citation2006) were derived from the Climatologic Atlas of Piedmont (Biancotti et al., Citation1998) and from the Hydrological Atlas of Switzerland (Schwarb et al., Citation2001).
Forage quality of swards was assessed by pastoral values (PV), which were calculated for each site according to Daget & Poissonet (Citation1971) weighted using van der Maarel (Citation1979) numerical values. Carrying capacity was computed using quality species indices reported by Cavallero et al. (Citation2007).
Floristic nomenclature follows Pignatti (Citation1982) and Aeschimann et al. (Citation2004). The syntaxonomical nomenclature follows Grabherr & Mucina (Citation1993), integrated with Theurillat et al. (Citation1994). Authors’ standard form of syntaxon names follows Izco (Citation2002).
Study area
Valle d’Aosta is surrounded by the highest summits of the Alps: Monte Bianco (4810 m a.s.l.), Monte Rosa (4634 m a.s.l.), Gran Paradiso (4061 m a.s.l.), and Cervino (3480 m a.s.l.). The orography is strongly influenced by glacial dynamics of the last glaciation (Burga, Citation1988). The orogaphic seclusion, the West-East direction and the southern-foehn influence, cause an intrinsic climate with lower precipitations, particularly in the centre of the valleys, and relative lower precipitation typical of inner-alpine regions (Schwabe & Kratochwil, Citation2004).
The geology of the study area is characterized mainly by silicate-dominant rocks and partly calcareous schists. In silicate rock dominant areas, often small base-rich or calcium carbonate-rich patches are observed, while serpentine rocks are very rare (Guyot, Citation1935).
Many of the investigated populations are actually grazed with very low stocking density, mainly with cattle and goats. Festuca paniculata stands occupy marginal areas around mountain farms, far-away from the cattleshed or located on very steep areas.
Results and discussions
Distribution range of Festuca paniculata communies
Festuca paniculata communities grow in Valle d’Aosta in small patches located in the subalpine(-alpine) belt from approximately 1800 to 2350 m. The studied populations are situated in two different valley districts (Fig. ). A first group of surveys is localized in the inner valley (Graian Alps), from Val di La Thuile (RIL08÷11) to Valgrisenche (RIL01÷07). In particular, on the right side of Valgrisenche, Festuca paniculata communities are widely spread. A second population (RIL12÷17) is localized in the outer valley (Pennine Alps), in Val di Gressoney, restricted to the top of Croix Courma (Fig. ).
Syntaxonomy and synecology
Vegetation can be referred to Junipero nanae-Festucetum spadiceae due to the constant presence of many character species: Juniperus nana, Pulsatilla alpina subsp. apiifolia, Silene nutans, Senecio doronicum, Helianthemum nummularium and Centaurea nervosa (Table ). This association was first described by Nègre (Citation1950) in the siliceous Meje-Ecrins-Pelvoux Massif.
Table I. Phytosociological table and synop tical table for the comparision between the Junipero-Festucetum spadiceae (Jun-Fest) and the Polygalo chamaebuxi-Festucetum paniculatae (Pol-Fest). Tableau 1. Tableau phytosociologique.
The attribution of the studied communities to Centaureo uniflorae-Festucetum spadiceae is unlikely, because many character species of this association were totally absent in our relevés (Centaurea uniflora, Hypericum richeri, Anemone narcissiflora, Hypochaeris maculata). Among the character species of Centaureo uniflorae-Festucetum spadiceae reported by Barbero (Citation1970) we found Trifolium montanum and Asphodelus albus with very low frequency (18% and 6% respectively). Paradisea liliastrum, here considered among the Festucetalia spadiceae character species (Grabherr, Citation1993), accounts for a frequency of 41%. Silene nutans, reported as character species both in Junipero-Festucetum spadiceae (Nègre, Citation1950) and in Centaureo uniflorae-Festucetum spadiceae (Barbero, Citation1970), was here considered as a companion species.
The comparison with Polygalo chamaebuxi-Festucetum paniculatae reveals in Valle d’Aosta communities the almost total absence of the character species of this association (Table ). Also, the communities localized in the outer part of Valle d’Aosta (Croix Courma) are not referable to the Insubrian Polygalo chamaebuxi-Festucetum paniculatae described in Colma di Mombarone (Biella and Torino districts), despite the whichdistance between Croix Courma and Colma di Mombarone which is only about 10 km.
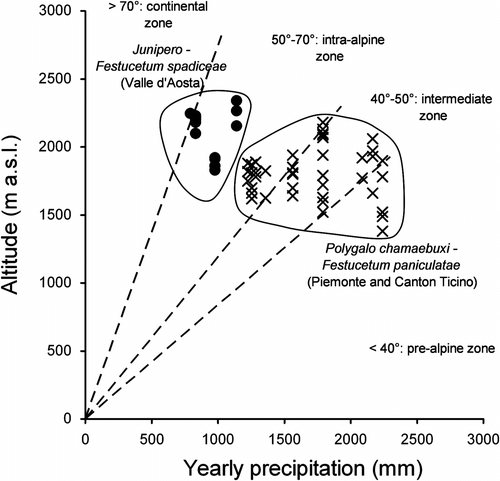
The explanation for this change in species composition is most likely linked to climatic conditions (Fig. ). In particular, the Insubrian Polygalo chamaebuxi-Festucetum paniculatae is localized between the outer-alpine and the inner-alpine zone, with optimum in the intermediate zone, with Gam’s continentality index ranging between 31.6 and 56.9° (Ozenda, Citation1985). Junipero-Festucetum spadiceae is localized between the intra-alpine and the continental zone, with a Gam’s continentality index ranging between 61.9 and 70.6°.
Fig. 3 Relation between altitude (m) and yearly precipitation (mm) for two Festuca paniculata associations: (•) Junipero-Festucetum spadiceae, (×) Polygalo chamaebuxi-Festucetum paniculatae. Lines show Gams’ continentality index thresholds (Gams’ angle) and define the ecological districts according to Ozenda (1985). Fig. 3. Relations entre l’altitude (m) et les precipitations annuelles (mm) pour les deux associations à Festuca paniculata: (•) Junipero-Festucetum spadiceae, (×) Polygalo chamaebuxi-Festucetum paniculatae. Les lignes indiquent les limites de l’indice de continentalité de Gams (angle de Gams) et définissent les domaines écologiques selon Ozenda (1985).
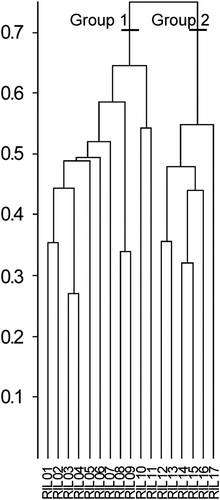
The cluster analysis identifies two groups of relevés (Fig. ): (i) Group 1, corresponding to the inner district of the valley (Graian Alps), and (ii) Group 2, corresponding to the outer district of the valley (Pennine Alps).
Fig. 4 Dendrogram of phytosociological surveys (average link, similarity ratio). Fig. 4. Dendrogramme des relevés phytosociologiques (lien moyen, similitude ratio).
The differences between the two groups were mainly explained by different ingressive species (Table ). In F. paniculata communities located in the outer district three subatlantic chamaephytic species ingressive from Calluno-Ulicetea: Chamaecytisus hirsutus, Genista tinctoria and Genista germanica. are very frequent The two latter species are localized in the outer Valle d’Aosta and in the lateral valley mouth region (Bovio et al., Citation2008). In the intra-alpine communities, these subatlantic species were totally replaced by continental species like Vaccinium myrtillus, Vaccinium gaultherioides and Vaccinium vitis-idaea, ingressive from Vaccinio-Piceetea.
The two groups differ also by the presence of many thermophilous species, ingressive from Festuco-Brometea (i.e. Brachypodium caespitosum, Centaurea triumfetti, Allium lusitanicum, Dianthus carthusianorum subsp. carthusianorum, Minuartia laricifolia, etc), exclusives of Group 2 (Table ) located at low altitudes (average altitude of 1868 m in Group 2 vs. 2214 m in Group 1, F-test p < 0.001).
The differences in floristic composition between the two groups do not justify a separation of the studied communities in two different subassociations. All the relevés were placed in the same subassociation Juniperetosum nanae, described by Nègre (Citation1950) in the more extensive areas, mainly grazed by wild ruminants. This attribution is confirmed by the frequent presence of differential species like Arctostaphylos uva-ursi, Thesium alpinum, Cotoneaster integerrimus, Bupleurum stellatum and Lilium bulbiferum. Compared with the Juniperetosum nanae described by Nègre (Citation1950) in the Meje-Ecrins-Pelvoux Massif (France), the Valle d’Aosta communities show many frequent ingressive species from Molinio-Arrhenetheretea, related to the grazing by domestic ruminants (goats or cattle), especially at the border of easily accessible pastures.
Syndynamics
The association can mainly be found on secondary stands which have been conserved by grazing pressure. The outer Valle d’Aosta populations, placed from the intermediate to subcontintal zone and located at low altitudes, are linked to the western acidophilous series of Pinus sylvestris. The inner-alpine Valle d’Aosta populations are connected to the western and central acidophilous inner alpine series of Pinus cembra-Larix decidua (Blasi, Citation2010).
Festuca paniculata stands, as also observed for many other gregarious species (Schwabe & Kratochwil, Citation2004), are always rich in an evident litter layer and may remain fallow lands for a long time and hinder the rejuvenation of woody species (Ozenda, Citation1985). Nevertheless, with the absence of grazing activities, a slow evolution towards the mature woody communities occurs, although the successional processes can be very slow at high altitude. Grazing extensification could probably induce in the future a surface increase of Festuca paniculata stands, with particular reference to marginal areas (Jouglet & Dorée, Citation1987). However, land abandonment, inducing an excessive litter accumulation, could have negative effect with a strong loss of biodiversity (Garde, Citation1996).
The PV calculated for our surveys highlights very low values, between 6 and 20 (0-100 scale), due to the occasional presence and to the low percentage cover of the palatable species of Molinio-Arrhenatheretea. The corresponding stocking-rate recommended for communities conservation ranged also between very low values (average 39.3 livestock units ha-1 d-1).
Conclusion
The Festuca paniculata communities in Valle d’Aosta are referable to Junipero-Festucetum spadiceae Juniperetosum nanae, previously described by Nègre (Citation1950) in the Meje-Ecrins-Pelvoux Massif (France).
Our results exclude the presence in the study area of Centaureo uniflorae-Festucetum spadiceae, supposed by Kaplan (Citation1983). This statement is in accordance with the previous observations of Vittoz et al. (Citation2005) that demonstrated a floristic similarity between the Kaplan survey and Junipero-Festucetum spadiceae.
Our phytosociological investigation defines the border between Polygalo chamaebuxi-Festucetum paniculatae and Junipero-Festucetum spadiceae. The analyses led us to the conclusion that the Insubrian Polygalo chamaebuxi-Festucetum paniculatae finds its western limit in Colma di Mombarone (Biella and Torino districts; Lonati & Siniscalco, Citation2006). Its western border is now better defined in correspondence of the administrative border of Aosta district. Data show clear and corresponding results with respect to an existing continental gradient that divides the suboceanic Colma di Mombarone with the Valle d’Aosta region. The potential forest communities in the different sites confirm also the continental gradient: Fagus sylvatica series under subatlantic influence for Polygalo chamaebuxi-Festucetum paniculatae in Colma di Mombarone, Pinus sylvestris series with intermediate to subcontinental climate for the communities observed in the Croix Courma and Pinus cembra-Larix decidua series under continental conditions for the inner Valle d’Aosta communities (Blasi, Citation2010).
The Festuca paniculata communities localized in Valle d’Aosta mouth region at Croix Courma (Valle di Gressoney, Group 2) occupy a floristic and ecological intermediate position between Polygalo chamaebuxi-Festucetum paniculatae and the inner Valle d’Aosta communities. This is confirmed by the presence in Group 1 of some subatlantic chamaephytic species of Calluno-Ulicetea. We further observe many ingressive species from Festuco-Brometea, due to the low altitude. Despite these differences between outer and inner-alpine communities, character species of the Polygalo chamaebuxi-Festucetum paniculatae association are totally absent in our relevés, supporting the hypothesis of a defined boundary between the two associations.
Acknowledgements
We thank Maurizio Bovio from the Museo Regionale di Scienze Naturali of the Valle d’Aosta who kindly provided indications of the Festuca paniculata populations in the study area.
References
- Aeschimann D., K. Lauber, M.D. Moser and J.P. Theurillat, 2004. Flora alpina. 3 vol. Zanichelli, Bologna.
- Barbero , M. 1970 . Les pelouses orophiles acidophiles des Alpes maritimes et ligures, leur classification phytosociologique . Nardetalia strictae, Festucetalia spadiceae et Caricetalia curvulae. Ann. Fac. Sci. Marseille , 43 : 173 – 195 .
- Biancotti , A. , Bellardone , G. , Bovo , S. , Cagnazzi , B. , Giacomelli , L. and Marchisio , C. 1998 . Distribuzione regionale di piogge e temperature , Torino : Cima Icam . 80 p
- Blasi , C. , ed. 2010 . La vegetazione d’Italia , Roma : Palombi and Partner S.r.l . 539 pp
- Bovio , M. , Broglio , M. and Poggio , L. 2008 . Guida alla flora della Valle d’Aosta , Torino : Blu edizioni . 335 p
- Braun-Blanquet , J. 1932 . Plant sociology , New York and London : McGraw-Hill Book Company . 439 p
- Buffa , G. and Dal Vesco , G. 1988 . Contributo alla conoscenza della flora del Piccolo San Bernardo: il Vallone di Chavannes (La Thuile, Valle d’Aosta) . Rev. Valdôtaine Hist. Nat , 42 : 5 – 30 .
- Burga , C.A. 1988 . Swiss vegetation history during the last 18.000 years . New Phytol , 110 : 581 – 602 .
- Cavallero , A. , Aceto , P. , Gorlier , A. , Lombardi , G. , Lonati , M. , Martinasso , B. and Tagliatori , C. 2007 . I tipi pastorali delle Alpi piemontesi , Bologna : Alberto Perdisa Editore . 464 p
- Daget , P. and Poissonet , J. 1971 . Une méthode d’analyse phytologique des prairies . Ann. Agron , 22 ( 1 ) : 5 – 41 .
- Gams , H. 1931 . Die klimatische Begrenzung von Pflanzenarealen und die Verteilung der hygrischen Kontinentalität in den Alpen I . Z. Ges. Erdkunde , 9 ( 10 ) : 321 – 346 .
- Gams , H. 1932a . Die klimatische Begrenzung von Pflanzenarealen und die Verteilung der hygrischen Kontinentalität in den Alpen II . Z. Ges. Erdkunde , 1 ( 2 ) : 52 – 68 .
- Gams , H. 1932b . Die klimatische Begrenzung von Pflanzenarealen und die Verteilung der hygrischen Kontinentalität in den Alpen III . Z. Ges. Erdkunde , 5 ( 6 ) : 178 – 198 .
- Garde , L. 1996 . Guide pastoral des espaces naturels du sud-est de la France , Louis-Jean , Gap : CERPAM/Méthodes et Communication . 250 p
- Grabherr G., 1993. Caricetea curvulae. In: Die Pflanzengesellschaften Österreichs 2, Natürliche waldfreie Vegetation. Grabherr G. and L. Mucina (eds.), Jena, 2, 343-372.
- Grabherr , G. and Mucina , L. 1993 . Die Pflanzengesellschaften Österreichs 2 , Fischer , Jena : Natürliche waldfreie Vegetation. G . 523 p
- Guyot , H. 1935 . Phytogéographie comparée du Valais et de la vallée d’Aoste . Bull. Maurithienne , 52 : 16 – 35 .
- Izco J., 2002. Authors of syntaxon names. In: Vascular plant communities of Spain and Portugal. S. Rivas-Martínez et al. (eds.), Itin. Geobot., 15, 413-432.
- Jouglet, J.P., Dorée, A., 1987. Paturage mixte: ovins-équins. Incidence sur la dynamique d’une pelouse subalpine à fétuque paniculée. Rapport 215, CEMAGREF/INERM, Grenoble, 31 p.
- Kaplan , K. 1983 . Über Gesellschaften des Festucion variae-Verbandes in den östlichen Grajischen Alpen (Aosta, Italien) . Ber. Geobot. Inst. ETH , 50 : 97 – 118 .
- Lonati M. and C. Siniscalco, 2006. Le praterie a Festuca paniculata (L.) Sch. et Th. (Polygalo chamaebuxi-Festucetum paniculatae ass. nova) della Alpi Pennine (Piemonte, Italia). Fitosociologia, 43 (1), 55-66.
- Mercalli , L. , Cat Berro , D. , Montuschi , S. , Castellano , C. , Ratti , M. , Di Napoli , G. , Mortasa , G. and Guineani , N. 2003 . Atlante climatico della valle d’Aosta , Torino : Graficat . 403 pp
- Nègre , R. 1950 . Contribution à l’étude phytosociologique de l’Oisans: la haute vallée du Vénéon (massif Mèije-Écrins-Pelvoux) . Phyton , 2 : 23 – 50 .
- Ozenda , P. 1985 . La végétation de la chaîne alpine dans l’espace montagnard européen , Paris : Masson . 330 p
- Pignatti, S., 1982. Flora d’Italia. 3 vol., Edagricole, Bologna.
- Podani , J. 1995 . Syn-Tax V. Computer program for data analysis in ecology and systematics , Trieste : Unido . 104 p
- Revel , V. , Siniscalco , C. and Montacchini , F. 2001 . Indagini sulla flora e sulla vegetazione della Val Sapin (Comune di Cormayeur, Valle d’Aosta) . Rev. Valdôtaine Hist. Nat , 55 : 25 – 43 .
- Sburlino , G. , Buffa , G. and Ghirelli , L. 1996 . L’analisi corologica nell’interpretazione sintassonomica: l’esempio delle praterie a Festuca paniculata (L.) . Sch. et Th. Giorn. Bot. It. , 130 ( 1 ) : 236 – 247 .
- Schwabe , A. and Kratochwil , A. 2004 . Festucetalia valesiacae communities and xerothermic vegetation complex in the Central Alps related to environmental factor . Phytocoenologia , 34 ( 3 ) : 329 – 446 .
- Schwarb M., C. Daly, C. Frei and C. Schär. 2001. Mean annual precipitation throughout the European Alps 1971–1990. In: Hydrological Atlas of Switzerland. Federal Office for Water and Geology, Berne, Plate 2.6.
- Theurillat , J.-P. , Aeschimann , D. , Küpfer , P. and Spichiger , R. 1994 . The higher vegetation units of the Alps . Colloques phytosociologiques , 23 : 190 – 239 .
- van der Maarel , E. 1979 . Transformation of cover-abundance values in phytosociology and its effects on community similarity . Vegetatio , 39 : 97 – 144 .
- Vittoz , P. , Selldorf , P. , Eggenberg , S. and Maire , S. 2005 . Les pelouse à Festuca paniculata du Tessin (Suisse) dans un contexte alpin . Bot. Helv , 115 : 33 – 48 .
Appendix 1
Syntaxonomic list.
Caricetea curvulae Br.-Bl. 1948 nom. cons. propos.
Festucetalia spadiceae Barbero Citation1970 em. Grabherr Citation1993
Festucion variae Guinochet 1938
Junipero nanae-Festucetum spadiceae Nègre Citation1950
Appendix 2
Other syntaxa quoted in the text.
Calluno vulgaris-Ulicetea minoris Br. Bl. & Tüxen ex Klika & Hadač 1944
Centaureo uniflorae-Festucetum spadiceae Guinochet 1938
Festuco valesiacae-Brometea erecti Br.-Bl. & Tüxen ex Klika & Hadač 1944
Hypochaerido uniflorae-Festucetum paniculatae Hartl ex Theurillat 1989
Hypochaerido uniflorae-Festucetum paniculatae Hartl ex Theurillat 1989 polygaletosum chamaebuxi Vittoz, Selldorf, Eggenberg & Maire Citation2005
Molinio caerulae-Arrhenatheretea elatioris Tüxen 1937 em. 1970
Polygalo chamaebuxi-Festucetum paniculatae (Vittoz, Selldorf, Eggenberg & Maire) Lonati & Siniscalco Citation2006
Vaccinio myrtilli-Piceetea abietis Br.-Bl. in Br.-Bl., Sissing & Vlieger 1939
Appendix 3
Date and UTM coordinates (WGS84) of the relevés (Table ).
RIL01: 08/07/2006, N 51758.42, E 349779.06; RIL02: 08/07/2006, N 52063.38, E 349392.49; RIL03: 08/07/2006, N 51888.36, E 349757.92; RIL04: 08/07/2006, N 51946.12, E 349727.24; RIL05: 08/07/2006, N 52460.74, E 349292.73; RIL06: 08/07/2006, N 49640.82, E 347569.04; RIL07: 08/07/2006, N 52691.67, E 347289.25; RIL08: 22/07/2010, N 59879.21, E 340105.09; RIL09: 23/07/2010, N 62996.27, E 333875.17; RIL10: 23/07/2010, N 64797.25, E 336017.21; RIL11: 23/07/2010, N 65016.25, E 335921.21; RIL12: 15/07/2006, N 53133.96, E 405241.68; RIL13: 15/07/2006, N 53165.96, E 405325.68; RIL14: 15/07/2006, N 53125.96, E 405428.67; RIL15: 15/07/2006, N 53141.96, E 405515.67; RIL16: 15/07/2006, N 53073.96, E 405503.67; RIL17: 15/07/2006, N 53058.96, E 405614.67.
Appendix 4
‘RIL01: Achillea moschata +, Pedicularis verticillata +, Potentilla crantzii +, Primula farinosa +, Veratrum album +; RIL02: Achillea moschata +, Arenaria serpyllifolia +, Chaerophyllum hirsutum +, Erigeron alpinus +, Oxytropis campestris +, Saxifraga aspera +, Sempervivum arachnoideum +, Silene vulgaris +, Veratrum album +; RIL03: Agrostis tenuis +, Carduus carlinaefolius +, Potentilla crantzii +, Sempervivum arachnoideum +, Sempervivum tectorum +; RIL04: Achillea moschata +,
Agrostis tenuis +, Carduus carlinaefolius +, Erigeron alpinus +, Polygala alpestris +, Sempervivum arachnoideum +; RIL05: Larix decidua (A) 1, Larix decidua (as) 1, Hieracium x auriculiforme +; RIL06: Gentiana nivalis +, Hieracium x auriculiforme +, Pedicularis verticillata +, Peucedanum ostruthium +, Rhinanthus aristatus +, Silene vulgaris +; RIL07: Agrostis tenuis +, Chaerophyllum hirsutum +, Convallaria majalis +, Epilobium angustifolium +, Polygonatum verticillatum +, Rhinanthus aristatus +, Sempervivum tectorum +, Silene vulgaris +, Trausteinera globosa +, Valeriana tripteris +; RIL08: Euphrasia hirtella +, Hieracium intybaceum +; RIL09: Hieracium intybaceum 1, Euphrasia hirtella +, Hieracium x auriculiforme +; RIL10: Carduus carlinaefolius +,
Carex ornithopoda +, Valeriana tripteris +; RIL11: Euphrasia hirtella +, Rhinanthus aristatus +; RIL12: Pulmonaria australis 1; RIL13: Pulmonaria australis +; RIL14: Asphodelus albus +, Astragalus penduliflorus +, Sempervivum tectorum +; RIL16: Amelanchier ovalis +, Betula pendula +, Calamagrostis arundinacea +, Carex ornithopoda +; RIL17: Larix decidua (A) 2, Pinus sylvestris (A) 2, Amelanchier ovalis (ai) +, Betula pendula (ai) +, Calamagrostis arundinacea +, Carex ornithopoda +, Convallaria majalis +, Laserpitium latifolium +.