Abstract
Acacia tortilis subsp.raddiana is one of the most uncommon species and is still present in North African regions. Indeed, the region of Bled Talah, in the south of Tunisia, constitutes today an ecosystem where the species continues to grow on a large surface of 16,488 ha. There is little information on the long-term dynamics of Acacia trees. The overall objectives of this study are to know the total individuals of Acacia trees which help to assess the density, to study the structure of Acacia population, to estimate the age of Acacia population and to determine the effects of A. tortilis canopy on soil properties. In this context, all individuals of A. tortilis covering 5,114 ha in the National Park Bov Hedma were systematically counted. Many dendrometric parameters were measured. In our study, age determination of A. tortilis is based on the counting of rings. Our results show that the population of A. tortilis subsp. raddiana in the region of Bou Hedma is characterised by an irregular structure, involved in a regressive dynamic. They show also that A. tortilis trees improve soil water availability and the nutrient status of soil.
Résumé
Acacia tortilis subsp. raddiana est une espèce clé de voute des zones arides et sahariennes de l’Afrique. En Tunisie, cette espèce occupe une superficie de 16,488 ha. Cependant, sa dynamique demeure encore peu ou pas connue. Ce travail a pour objectif l’analyse de la structure du peuplement, sur la base de certains paramètres dendrométriques et édaphiques (teneurs en azote, phosphore, matière organique et eau du sol sous et en dehors de la canopée), observés de façon exhaustive, pour tous les individus, occupant une superficie globale de 5,114 ha, au niveau du parc national du Bou Hedma. Parallèlement, un essai de détermination de l’âge des individus, par comptage de cernes fut réalisé sur un échantillon de 100 individus. Les résultats obtenus révèlent qu’en Tunisie, le peuplement d’A. tortilis subsp. raddiana se caractérise par une structure hétérogène, témoignant d’une dynamique plutôt régressive. En revanche, cette espèce exerce un effet positif sur les propriétés chimiques et hydriques du sol, sous canopée, ce qui facilite l’installation d’une riche strate herbacée.
Nomenclature: Le Floc’h et al. (Citation2010).
Introduction
Acacia tortilis (Forssk.) Hayne subsp. raddiana (Savi) Brenan (Fabaceae, Mimosoideae) has often been regarded as a “miracle” tree of circum-Saharian region. It has been suggested to function as a “Keystone species” in the arid and semi-arid regions of Africa and the Middle East (Aronson et al., 1993; Munzbergova & Ward, Citation2002; Abdallah et al., Citation2008; Noumi et al., Citation2010a,b). It is of importance for people and their domesticated animals, improves soil fertility and increases biodiversity (Abdallah et al., Citation2008; Noumi et al., Citation2011). It is well adapted to disturbances such as drought, fire and browsing.
In Tunisia, the geographical distribution of A. tortilis is limited to the region of Bou Hedma (16,488 ha, Bled Talah). In this area of very low mean annual rainfall (< 200 mm), this rare natural ecosystem has been subject to an increasing human pressure since the beginning of the 20th century. In 1986, an area of 5,114 ha was included in the Bou Hedma National Park and ranked by UNESCO (1986) as a biosphere reserve in order to preserve the Acacia population and impede desertification. Recently, there have been alarming reports of large-scale mortality, drying and lack regeneration of Acacia trees in this National Park (Noumi et al., Citation2010a). However, information is not complete and misinterpretations of natural regeneration are likely. As possible reasons for the lower regeneration of the Acacia trees, the following factors have been discussed: Acacia trees suffer from high infestation rates by Bruchidius raddianae and B. aurivillii (Derbel et al., Citation2007; Ben Jamâa et al., Citation2008). Moreover, Halevy & Orshan (Citation1972) reported high infestation rates of Acacia trees in African countries by seed beetles (Bruchidae, mostly Bruchidius raddianae and Caryedon palaestinicus).
There is the possibility of autoallelopathy in A. tortilis since several studies have shown that Acacia trees contain higher levels of bioactive chemicals (Reigosa et al., Citation1984; Rafiqul Hoque et al., Citation2003), suggesting a large inhibitory potential (Barnes et al., Citation1996; Noumi et al., Citation2010a).
Although, some studies have been conducted on this population of A. tortilis, in particular in the field of phenology and ecophysiology, the population dynamics of this tree species is not well known.
Although these plants play a key role in initiating the bush problem, little research has been directed at the autoecology of the species and the mechanisms which allow populations to coexist and replace each other over time. The knowledge of the dynamics of this population will certainly be helpful to different management scenarios (Swartzman & Kaluzny, Citation1987; Bart, Citation1995; Dunning et al., Citation1995). Martin & Moss (Citation1997) suggested that age determination on the basis of ring count holds great potential for the study of dry woodland ecosystem dynamics. They showed that age estimation enables the inference of population pattern and is essential to a fuller understanding of ecosystem dynamics.
In this study, we tried to further our knowledge about the long-term dynamics of A. tortilis trees under arid bioclimate. As such, it is important to estimate the population structure of these tree species as a prerequisite for developing sustainable use and management strategies. Focus here is on the hyper-arid ecosystem, where trees are key resources for local nomads. However, in arid and semiarid areas of the Mediterranean basin, restoration activities have been mainly based on plantations of trees (Maestre & Cortina, Citation2004).
The specific objectives studied here are 1) to determine the total individuals of Acacia trees which helps to assess the density, 2) to study the structure of Acacia populations, 3) to estimate the age of Acacia populations and 4) to determine the effects of A. tortilis tortilis on soil properties under the canopy, compared to open spaces among the trees.
Material and methods
Study area
The study was undertaken in the Bou-Hedma National Park which is located in the Governorate of Sidi Bouzid, central – southern of Tunisia (34.39° N, 9.38° E). Following the Emberger’s classification (Emberger, Citation1955), its climate is Mediterranean arid with temperate winters. The area prospected covers 5,114 hectares.
The average annual rainfall varies from 150 mm at 100 m elevation in the plain to 300 mm at 800 m elevation on the highest peak of the mountain range. However, like in other arid Mediterranean climates, inter-annual variability of precipitation is very high. In the plain area, mean minima temperature of the coldest month (January) is 3.9 °C and mean maxima temperature of the warmest month (August) is 36.2 °C.
According to the site geomorphology, soils are skeletal in the mountainous area, superficial and stony in the piedmont and become sandy to sandy-loamy in the flat area. On the mountainous massif of the park, natural vegetation is mainly dominated by vestigial forest species such as Juniperus phoenicea, Periploca angustifolia, Rhus tripartitum, Olea europea, Rosmarinus officinalis and Stipa tenacissima. Artemisia herba-alba, Anarrhinum brevifolium, Gymnocarpos decander and Helianthemum kahiricum colonize the piedmont. However, the flat area is covered by pseudo-savanna vegetation where only Acacia raddiana constitutes the tree stratum (Le Houérou, Citation1969; Noumi et al., Citation2011). The herbaceous stratum is dominated by chamaephytes such as Rhanterium suaveolens, Cenchrus ciliaris, Digitaria nodosa [= Digitaria commutata], Hammada schmittiana, Haloxylon scoparium Pomel [= Hammada scoparia] and Salvia aegyptiaca. With this original physiognomy of the herbaceous vegetation and in presence of A. raddiana, the only spontaneous tree species in southern Tunisia, it seems important to study the dynamics of A. tortilis under arid bioclimate.
Design and measurements
All individuals of A. tortilis covering the 5,114 ha were systematically counted and tree density was determined. For each tree, we measured trunk circumference at ground level because of heavy branching of the species at very low height.
In our study, age determination of A. tortilis is based on the counting of rings. In this context, a total of 100 discs from stems, with different circumferences, of Acacia tree from the National Park were collected for dendrochronological analysis. Regression analysis was assessed to estimate the age of the Acacia population covering the 5,114 ha.
Sample discs were examined under a binocular microscope (x10) and the rings identified and marked on three radii 120° to each other. When possible, rings were traced around the disc to check for continuity. Otherwise the final ring count was determined by taking an average of the counts on three radii. In order to predict the age of Acacia population, we used regression analysis.
Soil sampling
To investigate the effect of A. tortilis on soil fertility, we collected soil samples from two subhabitats (canopied subhabitats and uncanopied subhabitat or open areas). Soil samples were collected from the upper 10 cm soil, excluding litter and stones. Once in the laboratory, the samples were air-dried and passed through a 2 mm sieve for chemical analyses. Oxidizable soil organic matter was determined by the Walkly black procedure (Nelson & Sommers, Citation1982). Olsen’s bicarbonate extraction (Olsen & Sommers, Citation1982) and Kjeldahl’s method were used to analyse extractable phosphate and total nitrogen, respectively.
In parallel to soil fertility, soil moisture (volumetric soil water content) was measured in different subhabitats at a 10 cm depth with a FDR probe (ThetaProbe ML2x, Delta T, Cambridge, UK). The first measurements were made during the dry season, whereas the other ones were sampled after a 30 mm rain.
Statistical analysis
We used one-way analysis of variance (ANOVA) followed by Tukey’s HSD-tests to analyse differences in soil water and soil nutrient among the two subhabitats (canopied and uncanopied subhabitat). ANOVAs were conducted with SPSS version 12. Values of probability lower than 0.05 were considered as statistically significant. No transformations were required to meet parametric assumptions for ANOVA.
Results
Dynamics of A. tortilis
Density and size-class distribution of A. tortilis
A. tortilis specimens were systematically counted. A total of 23,191 individuals, covering an area of 5,114 ha were registered. The number of individuals per hectare, which represents the density, was about 5 individuals per hectare or less.
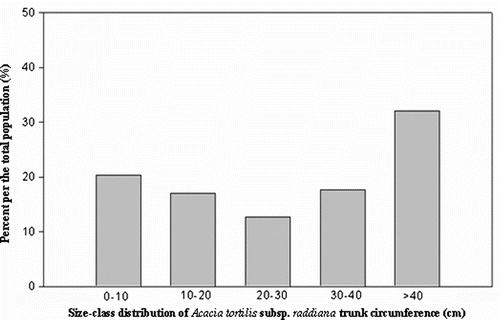
The frequency of trees in each circumference class for shown A. tortilis population sampled is shown in Fig. . Population structure shows the dominance of adult trees with high circumference. However, in the total of the 23,191 trees of Acacia, 32% a circumference superior to 40 cm. The juvenile classes (circumference < 10 cm), does not exceed 21% with 4,717 individuals. The other size classes (10-20, 20-30 and 30-40) are characterized by lower frequencies (< 20%). The population structure of A. tortilis could show an abormal trend in future, as there seems to be a decrease in natural regeneration.
Fig. 1 Size-class distribution of Acacia tortilis subsp. raddiana trunk circumferences (cm). Fig. 1. Structure du peuplement d’Acacia tortilis subsp. raddiana en fonction de la circonférence du tronc.
Age scenarios of A. tortilis
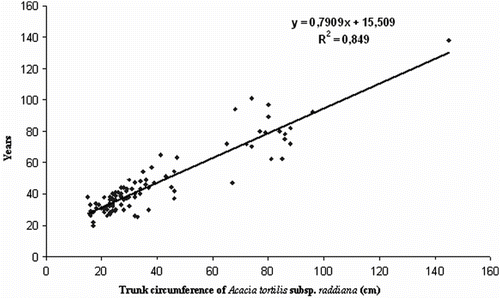
A plot of age on ring count for the one hundred stems with different circumferences is given in Fig. . The relationship between ring count and circumference shows a high correlation (R2 = 0.791). Using this regression, age of all individuals of Acacia tree is estimated with in a range from 16 to 316 years. This confirmes the dominance of aged trees in the total population, reflecting the lower natural regeneration.
Fig. 2 Relationship between age and trunk circumference of Acacia tortilis subsp. raddiana. Fig. 2. Corrélation entre la circonférence du tronc et l’âge des individus d’Acacia tortilis subsp. raddiana.
Soil properties
Soil fertility
The overall ANOVA of soil parameters shows that significantly higher amounts of nutrients in soils under canopy of the Acacia trees are observed compared to the open areas among the trees. Results of one-way ANOVA show a highly significant effect on the understorey compared to the open areas (Table .). However, the organic matter content of soil in open areas and under the canopy of A. tortilis ranges between 8.5 in open areas and 30.5 g.kg-1 in the canopied subhabitats. Concentrations of N and P range respectively between 0.49 and 0.02 in open areas, 1.32 and 0.07 g.kg-1 in the canopy of A. tortilis .
Table 1 Chemical properties of soils occurring under Acacia tortilis subsp. raddiana canopies and in open areas. Different letters denote significant differences between treatments (Tukey’s HSD-test at p < 0.05). Tableau 1. Effets d’Acacia tortilis subsp. raddiana sur les propriétés chimiques du sol.
Soil moisture
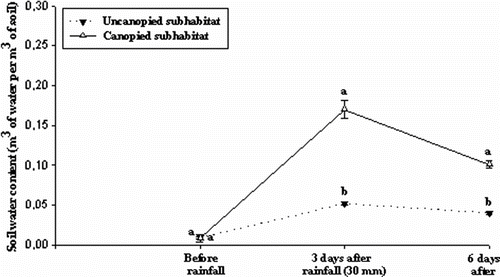
Figure shows the temporal evolution of soil water content of two subhabitats. During the dry period, there is no significant differences between subhabitats (p > 0.05). Three days after the rain (rainfall of 30 mm), results of one-way ANOVA show significant differences between the different subhabitats. However, the canopy subhabitat exhibited a quantity of water in the soil that was significantly higher than thaat sampled in the open subhabitats (p < 0. 05). Six days after rainfall, a significant effect was obvious between the understorey canopy and the open areas (p < 0.05).
Discussion
Acacia dynamics
This study improves general understanding of the population dynamics of Acacia trees in Southern of Tunisia. It emerges from this analysis that the population of Bou Hedma follows a certain regressive dynamics. In this context, our research shows the dominance of adult trees with big circumference of trunk. Our results agree with those reported in the Negev desert for the same species by several authors (Ashkenazi, Citation1995; Ward & Rhoner, Citation1997; Wiegand et al., Citation1998), and by Anthelme & Michalet (Citation2009) in the Air-Tenere Nature Reserve (Sahara, Niger) who showed the lack of natural regeneration of this species.
The dominance of large and aged individuals in the study area could be explained by the low rate regeneration of Acacia trees in this region. In this context, many factors could be discussed.
Taking into account that A. tortilis regenerates from seeds, it is possible that the seed properties could influence its regeneration levels. In fact, Acacia species have a hard seed coat which makes it difficult to absorb water unless they are firstly scarified (Demel, Citation1996). Moreover, it has been reported by Mekuria et al. (Citation1999) that seeds of tree legume species need pre-sowing treatments such as acid and mechanical scarification to make the hard seed coat permeable to water and improve germination. In the same way, Masamba (Citation1994) attributed the unsuccessful planting programs of A. tortilis to seed dormancy caused by the seed coat.
Furthermore, Acacia trees suffer extraordinarily high infestation rates by seed beetles (Bruchidae-mostly, Bruchidius raddianae and Caryedon palaestinicus; Halevy & Orshan, Citation1972). For example, Rohner & Ward (Citation1997) reported a seed infestation rate of 96% in A. tortilis and 98% in A. raddiana. Because of the substantially lower germination rates of infested seeds, recruitment might be seriously limited by Bruchidius (Halevy, Citation1974; Lamprey et al., Citation1974; Coe & Coe, Citation1987). In Tunisia, the presence of B. raddianae and B. aurivillii seems to be very dangerous, since our observations revealed the pod infestation from the beginning of its formation, where a majority of produced seeds were infested by this species (Derbel et al., Citation2007; Noumi et al., Citation2010a).
On the other hand, it is well known that natural regeneration of the woody species is threatened by the variation of water availability (Larwanou & Saadou, Citation2004). Van Bremen et al. (Citation1992) reported that protection encouraged the developments of surface pellicle in the soil which inhibits germination of seeds and reduces water infiltration. Working in a protected area (Bou Hedma National Park), the establishment of strong ferruginous crust does not encourage the regeneration of A. tortilis. These findings indicate that variation in water availability regime may have considerable effects on the dynamics and structure of Acacia populations.
An other possibility that can explain the lower regeneration of Acacia trees under arid bioclimate is the dominance of negative interaction or competition between the target species (Acacia) and others species in the same area. In this context, Noumi et al. (in press) showed that competition occurs in all experimental conditions for the Acacia target species by both the grass and the chenopod shrubs used as nurse species (Cenchrus ciliaris and Hammada scoparia) .
Fig. 3 Temporal evolution of soil water content for two subhabitats (canopied and uncanopied). Mean values which are not followed by the same letter are statistically significant (Tukey’s HSD-test at p < 0.05). Fig. 3. Evolution de la teneur en eau du sol dans les deux sous-habitats (sous la canopée et hors la canopée).
In conclusion, the population of A. tortilis subsp. raddiana in the region of Bou Hedma is characterised by an irregular structure, involved in a regressive dynamic of the species that could, by extrapolation, engender the turnover of the forest. Several activities and researches are still needed to protect this tree which is considered as a keystone species. Management intervention could be undertaken in the Bou Hedma National Park where the population density of A. tortilis is about 5 individuals or less per hectare.
Impact of Acacia tree on soil properties
In our field, A. tortilis has a significant effect on its understorey compared to open areas. The increased amount of soil nitrogen under trees in our study lends support to the influence of legumes on this important nutrient, as recorded in Barnes & Archer (Citation1996). The ability to fix nitrogen in A. tortilis was recorded by Assefa & Kleiner (Citation1998) and for A. raddiana by Ndoye et al. (Citation1995). Nitrogen concentrations under Acacia trees in our study correspond with those found by Belsky et al. (Citation1993) in low rainfall savannas in Africa under the canopies of A. tortilis and Adansonia digitata. As reported by Bernhard-Reversat (Citation1982) and Franco-Pizana et al. (Citation1995), we also showed the existence of a gradient of soil nutrients from the base of trunk to the edge of the canopy. The occurrence of N fixation due to microbial activity in association with the roots of leguminous trees is a possible source of N enrichment (Felker & Clark, Citation1982; Palm, Citation1995). Being a leguminous species, A. tortilis is classified among the trees with a high N2-fixing potential (Gueye & Ndoye, Citation2003).
In our study, total soil nitrogen tends to vary directly with soil organic matter. This is consistent with earlier findings showing high correlation between organic matter and nutrient content (Whitford et al., Citation1987). Our results are in agreement with those reported by Alstad & Vetaas (Citation1994), who found higher amounts of nitrogen and organic carbon and lower pH under large A. tortilis trees in one of their two study sites, and with observations on other shrubs in anarid zone of Spain by Pugnaire et al. (Citation2004). The increased level of phosphorus corroborates with those shown by Munzbergova & Ward (Citation2002) on A. raddiana. In the same way, but with others species, Turner et al. (Citation1966) and Franco-Pizana et al. (Citation1995) obtained the same results on Carnegia gigantea and Prosopis glandulosa respectively.
Although we do not know the exact source of the nutrient enrichment of the soils under tree canopies, many possible reasons can be proposed. It might be due, in part, to nutrient input by tree litter because with their roots, trees induce large volumes of soils for nutrients from surrounding surface and subsurface soils from their canopy and drop the nutrients in leaf and stem litter (Belsky et al., Citation1989; Amiotti et al., Citation2000). Another possible mechanism, the droppings from birds, is thought to be a major factor of increased soil nutrients built up under tree canopies. The long-term accumulation of nutrients at the trunk base through stem flow and bark sloughing is another explanation for nutrient enrichment (Kellman, Citation1979; Belsky et al., Citation1993).
In conclusion, the results of this study confirm the positive influence of A. tortilis in terms of soil enrichment and soil water availability. This is especially true with regards to OM, total N, P and soil water availability.
Acknowledgements
We are grateful to Lazhar Hemdi (Manager of the Bou Hedma National Park) for help. We would like to thank Guy-Georges Guittonneau for his valuable comments on an earlier version of this manuscript.
Related Research Data
References
- Abdallah , F. , Noumi , Z. , Touzard , B. , Ouled Belgacem , A. , Neffati , M. and Chaieb , M. 2008 . The influence of Acacia tortilis (Forssk.) subsp. raddiana (Savi) and livestock grazing on grass species composition, yield and soil nutrients in arid environments of South Tunisia . Flora , 203 : 116 – 125 .
- Alstad , G. and Vetaas , O.R. 1994 . The influence of Acacia tortilis stands on soil properties in arid north-eastern Sudan . Acta Oecol. , 15 : 449 – 460 .
- Amiotti , N.M. , Zolba , P. , Sanchez , L.F. and Peinemann , N. 2000 . The impact of single trees on properties of loess-derived grassland soils in Argentina . Ecology , 81 : 3283 – 3290 .
- Anthelme , F. and Michalet , R. 2009 . Grass-to-tree facilitation in an arid grazed environment (Aïr Mountains, Sahara) . Basic Appl. Ecol. , 10 : 437 – 446 .
- Aronson , J. , Floret , C. , Le Floc’h , E. , Ovalle , C. and Pontanier , R. 1993a . Restoration and rehabilitation of degraded ecosystems in arid and semi-arid lands. I. A view the South . Restor. Ecol , 1 : 8 – 17 .
- Aronson , J. , Floret , C. , Le Floc’h , E. , Ovalle , C. and Pontanier , R. 1993b . Restoration and rehabilitation of degraded ecosystems in arid and semi-arid lands. II. Case studies in southern Tunisia, central Chile, and northern Cameroon . Restor. Ecol. , 1 : 168 – 187 .
- Ashkenazi , S. 1995 . Acacia trees in the Negev and the Arava, Israel: A review following reported large-scale mortality (in Hebrew, with English summary) , Jerusalem : Hakeren HaKayemet LeIsrael, XVII . 121 p
- Assefa , F. and Kleiner , D. 1998 . Nodulation pattern and acetylene reduction (nitrogen fixation) activity of some highland and lowland Acacia species in Ethiopia . Biol. Fertil. Soils , 27 : 60 – 64 .
- Barnes , P.W. and Archer , S. 1996 . Influence of an overstory tree (Prosopis glandulosa) on associated shrubs in a savanna parkland: implications for patch dynamics . Oecologia , 105 : 493 – 500 .
- Barnes , R.D. , Filer , D.L. and Milton , S.J. 1996 . Acacia karroo Tropical Forestry Papers No 32 , Oxford Forestry Institute, Oxford University .
- Bart , J. 1995 . Acceptance criteria for using individual-based models to make management decisions . Ecol. Appl. , 5 : 411 – 420 .
- Belsky , A.J. , Amundson , R.G. , Duxburg , J.M. , Riha , S.J. , Ali , A.R. and Mwonga , S.M. 1989 . The effects of trees on their physical, chemical, and biological environments in a semi-arid savanna . J. Appl. Ecol. , 26 : 1005 – 1024 .
- Belsky , A.J. , Mwonga , S.M. , Amundson , R.G. , Duxbury , J.M. and Ali , A.R. 1993 . Comparative effects of isolated trees on their undercanopy environments in high and low rainfall savannas . J. Appl. Ecol. , 30 : 143 – 155 .
- Ben Jamâa , M.L. , Sliti , O. and Barbouche , N. 2008 . Les insectes bruchidés de l’Acacia tortilis subsp. raddiana en Tunisie . Ann. INGREF , 12 : 128 – 143 .
- Bernhard-Reversat , F. 1982 . Biogeochemical cycles of nitrogen in a semi-arid savanna . Oikos , 38 : 321 – 332 .
- Coe , M. and Coe , C. 1987 . Large herbivores, acacias trees and bruchid beetles . S. Afr. J. Sci. , 83 : 624 – 635 .
- Demel , T. 1996 . Germination ecology of twelve indigenous and eight exotic multipurpose leguminous species from Ethiopia . Forest Ecol. Manag. , 80 : 209 – 223 .
- Derbel , S. , Noumi , Z. , Anton , K.W. and Chaieb , M. 2007 . Life cycle of the coleopter Bruchidius raddianae and the seed predation of the Acacia tortilis subsp. raddiana in Tunisia . C. R. Biol. , 330 : 49 – 54 .
- Dunning , J.B. , Steward , D.J. , Danielson , B.J. , Noon , B.R. , Root , T.L. , Lamberson , R.H. and Stevens , E.E. 1995 . Spatially-explicit population models: current forms and future uses . Ecol. Appl. , 5 : 3 – 11 .
- Emberger , L. 1955 . Une classification biogéographique des climats. Faculté des Sciences, Service botanique . Revue des travaux de Laboratoire de Botanique et de Zoologie , 7 : 3 – 43 .
- Felker , P. and Clark , P.R. 1982 . Position of mesquite (Prosopis subsp.) nodulation and nitrogen fixation (acetylene reduction) in 3-m long phraetophytically simulated soil columns . Plant Soil , 64 : 297 – 305 .
- Franco-Pizana , J. , Fulbright , T.E. and Gardiner , D.T. 1995 . Spatial relations between shrubs and Prosopis glandulosa canopies . J. Veg. Sci. , 6 : 73 – 78 .
- Gueye , M. and Ndoye , I. 2003 . “ Le potential fixateur d’azote d’Acacia raddiana comparé à celui d’Acacia senegal, Acacia seyal et Faidherbia albia ” . In Un arbre au désert, Acacia raddiana , Edited by: Grouzis , M. and Le Floc’h , E. 201 – 227 . IRD Editions .
- Halevy , G. 1974 . Effets of gazelles and seed beetles (Bruchidae) on germination and establishment of Acacia species . Isr. J. Bot. , 23 : 120 – 126 .
- Halevy , G. and Orshan , C. 1972 . Ecological studies of Acacia raddiana, Acacia tortilis and Acacia gerardii ssp. negavensis . Isr. J. Bot. , 21 : 197 – 208 .
- Kellman , M. 1979 . Soil enrichment by neotropical savanna trees . J. Ecol. , 67 : 565 – 577 .
- Lamprey , H.F. , Halevy , G. and Makacha , S. 1974 . Interactions between Acacia, bruchid seed beetles and large herbivores . East Afr. Wildlife J. , 12 : 81 – 85 .
- Larwanou , M. and Saadou , M. 2004 . Biodiversity of ligneous species in semi-arid to arid zones of southwestern Niger according to anthropogenic and natural factors . Agr. Ecosyst. Environ. , 105 : 267 – 271 .
- Le Floc’h, E., Boulos, L. and E. Vela, 2010. Catalogue synonymique commenté de la flore de Tunisie. République Tunisienne, Ministère de l’Environnement et du Développement durable, 500 p. + errata (version numérisée).
- Le Houérou H.N., 1969. La végétation de la Tunisie steppique (avec références au Maroc, à l’Algérie et à la Libye). Analyses de l’INRAT-tunisie, 42, 617 p.
- Maestre , F.T. and Cortina , J. 2004 . Insights into ecosystem composition and function in a sequence of degraded semiarid steppes . Restor. Ecol. , 12 : 494 – 502 .
- Martin , D.M. and Moss , J.M.S. 1997 . Age determination of Acacia tortilis (Forsk.) Hayne from northern Kenya . Afr. J. Ecol. , 35 : 266 – 277 .
- Masamba , C. 1994 . Presowing seed treatments on four African Acacia species: appropriate technology for use in forestry for rural development . Forest Ecol. Manag. , 64 : 105 – 109 .
- Mekuria , A. , Demel , T. and Mats , O. 1999 . Soil seed flora, germination and regeneration pattern of woody species in Acacia woodland of the Rift Valley in Ethiopia . J. Arid Environ. , 43 : 411 – 435 .
- Munzbergova , Z. and Ward , D. 2002 . Acacia trees as keystone species in Negev desert ecosystems . J. Veg. Sci , 13 : 227 – 236 .
- Ndoye , I. , Gueye , M. , Danso , S.K.A. and Dreyfus , B. 1995 . Nitrogen fixation in Faidherbia albida, Acacia raddiana, Acacia senegal and Acacia seyal estimated using the 15N isotope dilution technique . Plant Soil , 172 : 175 – 180 .
- Nelson , D.W. and Sommers , L.E. 1982 . “ Total carbon, organic carbon, and organic matter ” . In Methods of Soil Analysis , Edited by: Page , A.L. Madison , Wis : American Society of Agronomy, Soil Science Society of America .
- Noumi , Z. , Abdallah , F. , Torre , F. , Michalet , R. , Touzard , B. and Chaieb , M. 2011 . Impact of Acacia tortilis ssp. raddiana tree on wheat and barley yield in the south of Tunisia . Acta Oecol. , 37 : 117 – 123 .
- Noumi , Z. , Ouled Dhaou , S. , Abdallah , F. , Touzard , B. and Chaieb , M. 2010a . Acacia tortilis subsp. raddiana in the North african arid zone: the obstacles to natural regeneration . Acta Bot. Gallica , 157 : 231 – 240 .
- Noumi , Z. , Touzard , B. , Michalet , R. and Chaieb , M. 2010b . The effects of browsing on the structure of Acacia tortilis (Forssk) Hayne subsp. raddiana (Savi) Brenan along a gradient of water availability in arid zones of Tunisia . J. Arid Environ. , 74 : 625 – 631 .
- Olsen , S.R. and Sommers , L.E. 1982 . “ Phosphorus ” . In Methods of Soil Analysis , Edited by: Page , A.L. 403 – 430 . Madison , Wis : American Society of Agronomy, Soil Science Society of America .
- Palm , C.A. 1995 . Contribution of agroforestry trees to nutrient requirements of intercropped plants . Agroforest. Syst. , 30 : 105 – 124 .
- Pugnaire , F. , Armas , C. and Valladares , F. 2004 . Soil as a mediator in plant-plant interactions in a semi-arid community . J. Veg. Sci. , 15 : 85 – 92 .
- Rafiqul Hoque , A.T.M. , Ahmed , R. , Uddin , M.B. and Hossain , M.K. 2003 . Allelopathic effect of different concentration of water extracts of Acacia auriculiformis leaf on some initial growth parameters of five common agricultural crops . Pak. J. Agr. , 2 : 92 – 100 .
- Reigosa , M.J. , Casal , J.F. and Carballeira , A. 1984 . Efectos alelopaticos de Acacia dealbata Link durante su floracion . Stud. Oecol. , 5 : 135 – 150 .
- Rohner , C. and Ward , D. 1997 . Chemical and mechanical defense against herbivory in two sympatric species of desert Acacia . J. Veg. Sci. , 8 : 717 – 726 .
- Swartzman , G.L. and Kaluzny , S.P. 1987 . Simulation model evaluation , New York , , USA : Macmillan .
- Turner , R.M. , Alcorn , S.M. , Olin , G. and Booth , J.A. 1966 . The influence of shade, soil and water on saguaro seedling establishment . Bot. Gaz. , 127 : 95 – 102 .
- Van Bremen H. and H.C.K. Kinyanjui, 1992. Soils of the Lodwar area (quarter degree sheets, 24, 25 and parts of 26, 38, 39 & 40): an inventory of soils, an evaluation of present land use and recommendation for future land use. Report (n°R17) of Kenya Soil Survey, Nairobi, Kenya.
- Ward , D. and Rhoner , C. 1997 . Anthropogenic causes of high mortality and low recruitment in three Acacia tree taxa in the Negev desert Israel . Bio. Conserv. , 6 : 877 – 893 .
- Whitford , W.G. , Reynolds , J.F. and Cunningham , G.L. 1987 . “ How desertification affects nitrogen limitation of primary production on Chihuahuan desert watersheds ” . In Proceedings of the Symposium on Strategies for Classification and Management of Natural Vegetation for Food Production in Arid Zones , Edited by: Moir , W. and Aldon , E.A. Denever , Colorado : U.S. Forest Service .
- Wiegand , K. , Jeltsch , F. and Ward , D. 1999 . Analysis of the population dynamics of Acacia trees in the Negev desert, Israel with a spatially-explicit computer simulation model . Ecol. Model. , 117 : 203 – 2 .