Résumé
La présente étude montre la variation saisonnière des teneurs en substances qualifiées de stress des principaux organes végétatifs (feuilles, tiges et rhizome) de l’oyat (Ammophila arenaria (L.) Link) poussant en conditions naturelles sur la côte ouest de l’Algérie. Ces substances sont la proline et les sucres solubles qui sont généralement libérés et stockés dans les cellules des organes végétatifs des plantes supérieures en réponse aux différentes contraintes environnementales. Les résultats montrent d’une part que les teneurs respectives en proline et en sucres solubles sont relativement élevées comparativement à celles d’espèces glycophiles et halophiles étudiées, d’autre part ces substances ont tendance à s’accumuler dans la partie aérienne plutôt que dans la partie souterraine (rhizome), dans les organes les plus âgés et pendant les saisons les plus sèches et les plus chaudes, les sites de stockage seraient les feuilles les plus âgées, les rhizomes sont les organes les moins riches en ces substances.
Abstract
The present study brings to the seasonal variation of the contents in qualified substances of the stress of the principal vegetative organs (leaves, stems and rhizomes) of oyat (Ammophila arenaria (L.) Link) growing in natural conditions in the Algerian western coast. These substances are the proline and soluble sugars which are generally freed and stocked in the cells of the vegetative organs of superior plants in response to various environmental constraints. The results show on one side that the respective contents in proline and soluble sugars are relatively high compared to the ones of glycophiles and halophiles species already studied and, on the other side, that substances are quantitatively better represented in the aerial part than in the underground part (rhizome), in older organs and during the driest and the hottest seasons, the stocking and resistance sites would be the older leaves, the rhizomes are the organs the least rich in these substances.
Introduction
Le substrat sableux de la partie continentale du littoral marin n’est pas un milieu idéal pour un bon développement des végétaux qui doivent développer des stratégies spécifiques de survie pour palier les nombreuses contraintes qu’elles affrontent. Les plus importantes de ces contraintes sont la pauvreté en apports nutritifs, la mobilité du sable qui les menace en permanence de déchaussement ou d’ensevelissement, le manque d’eau en surface et en profondeur car le sable est extrêmement poreux, perméable et filtrant, ainsi que les vents fréquents très souvent violents et chargés d’embruns salés. Les milieux littoraux subissent donc de plein fouet l’action des vents chargés d’humidité et d’embruns salés, le mitraillage par le sable, la déflation, le dessèchement du substrat, la perte des nutriments par lessivage et la chaleur solaire. Ces différents facteurs sont ainsi des paramètres de stress qui s’imposent en permanence aux composantes biocénotiques de ce type de milieu.
Bien que les effets des stress dépendent du stade de développement des plantes, des conditions climatiques et édaphiques, plusieurs caractères physiologiques et biochimiques se manifestent chez les plantes et sont directement corrélés à la tolérance à ces stress (Rathert, Citation1984; Zid and Grignon, Citation1991; Belkhodja, Citation2000; Hernandez et al., Citation2000). Pour leur survie, les espèces qui poussent dans ces conditions extrêmes et précaires présentent un arsenal particulier d’adaptations qu’il est fondamental de comprendre dans le but de leur valorisation. L’oyat (Ammophila arenaria (L.) Link), variété arundinacea (Host) (Quezel & Santa, Citation1963), objet de notre travail, compte parmi les rares espèces vivaces les plus caractéristiques de ce type de milieu et les mieux adaptées à ces différents facteurs contraignants, elle développe dans ces conditions extrêmes une stratégie offensive rare chez la plupart des autres plantes et évoquée par un certain nombre d’auteurs dont Kühnholtz Lordat (1928, in Leclerc, Citation1985).
Graminée vivace strictement psammophile poussant sous forme de touffes, l’oyat peut atteindre une hauteur de 1,50 m et se reproduit particulièrement par voie végétative à travers des bourgeons se formant le long d’un rhizome traçant de plusieurs mètres de longueur qui suit la déclinaison de la dune.
La plupart des travaux de recherche effectués de par le monde portent essentiellement sur les aspects phytoécologiques et écophysiologiques qui convergent tous vers la voie de la résolution de ce que les chercheurs qualifient de « problème Ammophila » (Maun & Baye, Citation1989) et pratiquement aucun travail n’est consacré à sa valorisation biochimique. Largement représentée en Algérie le long de la frange littorale méditerranéenne continentale, cette espèce montre une très grande adaptation à son milieu, elle se développe avec une vigueur sans égal dans les dunes en cours de formation (Aimé & Penven, Citation1982; Bendimered, Citation1997), c’est une espèce pionnière des dunes vives proliférant là où aucune autre espèce ne peut survivre dans les mêmes conditions.
Pour valoriser un des aspects de forte adaptation de cette espèce aux pressions environnementales, nous nous sommes intéressés à l’étude de deux substances largement reconnues comme substances de stress, à savoir la proline et les sucres solubles. Le présent travail consiste en la détermination quantitative et la variation saisonnière de ces deux substances accumulées dans la partie aérienne et rhizomateuse de l’oyat en conditions naturelles.
De nombreux auteurs admettent depuis environ deux décennies que l’accumulation des sucres et de la proline est une réponse métabolique commune aux plantes exposées à des contraintes de l’environnement (Weimberg et al., Citation1986; Wang & Stuttle, Citation1992; Taylor, Citation1996). En effet dans les milieux salés, les plantes s’adaptent métaboliquement en accumulant dans le cytoplasme des composés azotés quaternaires tels que la proline (Smirnoff & Stewart, Citation1985; Goldhirs et al., Citation1990; Belkhodja, Citation1996, Citation2000) pour exercer un ajustement osmotique dans les cellules (Goldhirs et al., Citation1990; Ottow et al., Citation2005.).
Matériel biologique et méthodes d’étude
Matériel biologique
Les prélèvements sont effectués sur des touffes d’oyat poussant dans les conditions naturelles durant les quatre saisons de l’année 2008. Le site de prélèvement des échantillons choisi est une dune du cordon de Cap Falcon du littoral de la wilaya d’Oran, à l’ouest de l’Algérie, (Fig. ). Dans ce site, les dunes sont en voie de remobilisation marquée par une extension importante de l’oyat avec de nombreuses jeunes pousses qui proviennent de quelques touffes plus âgées. Ce site fait partie d’une région caractérisée par un bioclimat semi-aride à hiver doux subissant une longue saison sèche et chaude de cinq mois en moyenne: de mai à septembre, il est exposé à des vents souvent violents de secteur ouest et chargés d’embruns salés (Bendimered, Citation1997; Bendimered et al., Citation2007). Les taux de sels dans l’atmosphère et à la surface des végétaux et des sols sont très élevés et l’effet est accentué en période sèche.
Fig. 1 Situation géographique de la zone de prélèvement des échantillons d’organes de Ammophila arenaria. Fig. 1. The geographical situation of the area of Ammophila arenaria organs samples collections.
Méthodes d’étude
Préparation des échantillons de la plante
La plante entière est soigneusement prélevée. La partie aérienne (tiges et feuilles) est séparée de la partie souterraine (rhizomes), les racines n’étant pas considérées dans cette étude (elles feront l’objet d’une autre). Les feuilles qualifiées de «âgées» sont celles du troisième rang ayant achevé leur croissance et atteint le maximum de leur longueur dont la valeur se situe entre 50 et 60 cm, tandis que les feuilles jeunes sont celles du même rang que les feuilles âgées mais en pleine croissance et présentent une longueur égale à la moitié de celle de ces dernières (de 20 à 30 cm) (Fig. ). Chaque organe est lavé délicatement pour supprimer les poussières et les impuretés, égoutté puis pesé.
Fig. 2 Organes de Ammophila arenaria utilisés pour les dosages de la proline et des sucres solubles (le trait d’échelle représente 3 cm). Fa: feuilles âgées, Fj: feuilles jeunes, T: Tiges, Rh: rhizomes. Fig. 2. Ammophila arenaria organs used for the assay of the proline and soluble sugars (the scale feature represents 3 cm). F: leaves, Fa: old leaves, Fj: young leaves, T: stems, Rh: rhizome.

Extraction et dosage de la proline
L’extraction de la proline s’est faite à l’éthanol selon la méthode de l’AOAC (Association of Official Analytical Chemists, Citation1955) modifiée par Nguyen & Paquin (Citation1971) utilisant successivement de l’éthanol 95%, de l’éthanol 70% et du chloroforme à froid.
La proline est dosée selon la méthode colorimétrique de Bergman & Loxley (Citation1970) utilisant les solutions respectives de NaCl 5M, de tampon phosphate et de ninhydrine. La densité optique est lue, sur un spectrophotomètre à absorption moléculaire, à une longueur d’onde de 505 nm.
Les résultats sont exprimés en μg.100 mg-1 de matière végétale en référence à une courbe-étalon obtenue à partir de solutions pures de proline à concentrations croissantes préparées dans les mêmes conditions que les solutions à doser.
Extraction et dosage des sucres solubles
La méthode au phénol de Dubois et al. (Citation1956) est utilisée pour l’extraction et le dosage des sucres après un traitement à l’éthanol à 80% et une solubilisation à l’eau distillée. Le dosage colorimétrique se fait par addition respective de solutions de phénol 5% et d’acide sulfurique concentré. Les lectures de l’absorbance sont effectuées à une longueur d’onde de 485 nm.
Les résultats des concentrations en sucres solubles sont déduits à partir d’une courbe-étalon de solutions de glucose à différentes concentrations préparées dans les mêmes conditions que les échantillons et exprimés en μg.100 mg-1 de matière végétale.
Traitement des données
Les résultats sont traités statistiquement par l’analyse des séries appariées du test de Student à une probabilité de 5% pour tous les échantillons, pour les quatre saisons, avec cinq répétitions pour chaque échantillon en utilisant le logiciel StatView pour la comparaison des résultats et du logiciel Stat.12 pour déterminer la corrélation entre les deux substances de stress étudiées en fonction des saisons et de l’âge de la plante.
D’autre part un dendrogramme interprétatif de la matrice de Pearson est établi dans un but de récapitulation de tous les résultats obtenus et l’analyse est effectuée à partir de l’indice de Pearson qui permet d’établir le degré de ressemblance entre les organes par rapport aux concentrations des deux substances de stress étudiées pendant les quatre saisons.
Résultats et discussion
Résultats
Variation des teneurs en proline et en sucres solubles dans différents organes pour la même saison
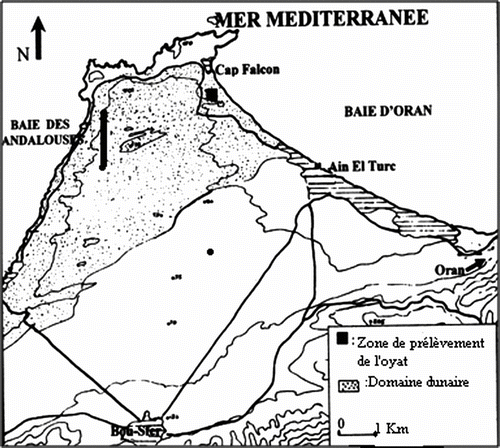
En été, la quantité de proline est la plus élevée au niveau des feuilles âgées (62,3 ± 0,277 μg.100 mg-1 de matière végétale MV); ces valeurs diminuent depuis les feuilles jeunes jusqu’aux rhizomes en passant par les chaumes avec des valeurs respectives de 48,6 ± 0,443, 40,5 ± 0,353 et 28,4 ± 0,412 μg.100 mg-1 de MV (Fig. ). L’accumulation des sucres solubles avec de fortes teneurs est constatée au niveau des feuilles les plus âgées (179,1 ± 0,420 μg. 100 mg-1 de MV) et ces valeurs diminuent progressivement des feuilles jeunes vers le rhizome en passant aussi par les chaumes (131,3 ± 0,299, 98,1 ± 0,611 et 46,2 ± 0,422 μg. 100 mg-1 MV (Fig. ).
Fig. 3 Variation des teneurs moyennes en proline dans les quatre organes de l’oyat pour chaque saison. Fa: feuilles âgées, Fj: feuilles jeunes, T: Tiges, Rh: rhizomes. Fig. 3. Variation of the average contents in proline of the oyat four organs for each season. Fa: old leaves, Fj: young leaves, T: stems, Rh: rhizomes.
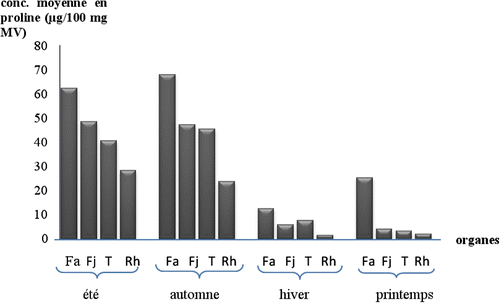
Fig. 4 Variation des teneurs moyennes en sucres solubles dans les quatre organes de l’oyat pour chaque saison. Fa: feuilles âgées, Fj: feuilles jeunes, T: tiges, Rh: rhizomes. Fig. 4. Variation of the average contents in soluble sugars of the oyat four organs for each season. Fa: old leaves, Fj: young leaves, T: stems, Rh: rhizomes.
En automne, le taux de proline le plus élevé est enregistré dans les feuilles âgées avec une valeur moyenne de 68,1 ± 0,499 μg. 100 mg-1 de MV suivies par celui des tiges dont la valeur moyenne est 45,5 ± 0,480 μg. 100 mg-1 MV. Dans les feuilles jeunes la valeur moyenne est de 47,4 ± 0,459 μg.100 mg-1 de MV et celle des rhizomes est la plus faible avec 23,6 ± 0,947 μg. 100 mg-1 MV (Fig. ). Concernant les sucres solubles, les plus fortes valeurs sont enregistrées également dans les feuilles âgées suivies de celles des feuilles jeunes puis des tiges et les plus faibles dans les rhizomes avec des valeurs moyennes respectives de 234,7 ± 2,943 μg.100 mg1, 101,7 ± 1,445 μg. 100 mg-1, 94,7 ± 1,160 μg. 100 mg-1 et 84,3 ± 0,511 μg. 100 mg-1 de MV (Fig. ).
La concentration de la proline au printemps est moins importante que celle enregistrée en automne et en été mais elle est toujours plus importante au niveau des feuilles âgées (25,2 ± 0,280 μg.100 mg-1) et la diminution de la quantité de cet acide aminé est très marquée à partir des feuilles jeunes jusqu’au rhizome, les valeurs trouvées au niveau des feuilles jeunes, de la tige et du rhizome sont presque six fois moins importantes que celle des feuilles âgées et sont respectivement: 4,1 ± 0,072, 3,3 ± 0,233 et 2,1 ± 0,013 μg. 100 mg-1 MV (Fig. ). Aussi, les valeurs des concentrations en sucres solubles les plus basses ont été obtenues durant la période printanière avec une valeur toujours plus élevée dans la feuille âgée (80,7 ± 0,277 μg. 100 mg-1 MV). La diminution des concentrations est remarquable au niveau de tous les autres organes étudiés. Celles obtenues au niveau des rhizomes (25,1 ± 0,630 μg. 100 mg-1 MV) sont trois fois moins importantes que celles trouvées au niveau des feuilles âgées, et moins de la moitié de la valeur moyenne trouvée dans le chaume (56 ± 0,374 μg. 100 mg-1 MV) (Fig. ).
L’hiver est la saison des plus basses concentrations en proline au niveau de tous les organes, de la feuille âgée (12,5 ± 0,354 μg. 100 mg-1) aux rhizomes (1,4 ± 0,226 μg. 100 mg-1) (Fig. ). Les valeurs sont toujours plus importantes au niveau des feuilles âgées, deux fois plus élevées que celles des jeunes feuilles (5,8 ± 0,461 μg.100 mg-1) et des tiges (7,5 ± 0,288 μg. 100 mg-1 MV). La plus grande accumulation en sucres solubles est également observée au niveau des feuilles âgées (133,6 ± 0,927μg. 100 mg-1 MV), la concentration diminue dans les feuilles jeunes (80,9 ± 0,460 μg. 100 mg-1) et dans les tiges (72,3 ± 0,661 μg. 100 mg-1) jusqu’à ce qu’elle atteigne moins du tiers de la valeur au niveau du rhizome (59,3 ± 0,477 μg. 100 mg-1 MV) (Fig. ).
L’analyse statistique montre des différences significatives entre les valeurs moyennes relatives aux concentrations respectives en proline et en sucres solubles des organes en fonction de chaque saison considérée.
Variation des teneurs en proline et en sucres solubles du même organe pendant les quatre saisons
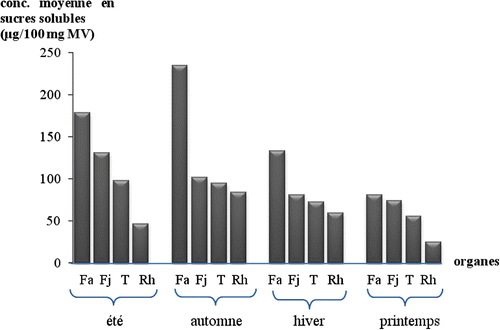
Les feuilles âgées concentrent le maximum de proline en automne (68,1 ± 0,499 μg. 100 mg-1 de MV) contre un minimum enregistré en hiver avec une valeur moyenne de 12,4 ± 0,354 μg. 100 mg-1, elle augmente progressivement au printemps et en été, les valeurs passant du simple au double (Fig. ). Parallèlement, ces mêmes organes accumulent le maximum de sucres solubles en automne (234,7 ± 2,943 μg. 100 mg-1), le minimum au printemps (80,7 ± 0,277 μg. 100 mg-1), les valeurs des moyennes en sucres solubles en été et en hiver avoisinent le double de celles du printemps, tout en restant significativement différentes entre elles: 179,1 ± 0,421 μg. 100 mg-1, contre 133,6 ± 0,927 μg.100 mg-1 (Fig. ).
Fig. 5 Variation saisonnière des teneurs moyennes en proline pour chaque organe de l’oyat. E: été, A: automne, H: Hiver, P: printemps, Fa: feuilles âgées, Fj: feuilles jeunes, T: tiges, Rh: rhizomes. Fig. 5. Seasonal variation of the average contents in proline for each oyat organ. Fa: old leaves, Fj: young leaves, T: stems, Rh: rhizome, A: autumn, H: winter, P: spring, S: summer.
Fig. 6 Variation saisonnière des teneurs moyennes en sucres solubles dans chaque organe de l’oyat. E: été, A: automne, H: hiver, P: printemps, Fa: feuilles âgées, Fj: feuilles jeunes, T: tiges, Rh: rhizomes. Fig. 6. Seasonal variation of the average contents in soluble sugars for each oyat organ. Fa:old leaves, Fj: young leaves, T: stems, Rh: rhizome, A: autumn, H: winter, P: spring, S: summergr.
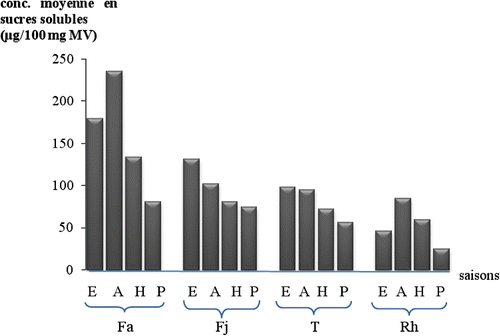
Les feuilles jeunes accumulent significativement moins de proline et de sucres solubles que les feuilles âgées pour les quatre saisons, ces valeurs étant les plus importantes en été (48,6 ± 0,443 μg. 100 mg-1) puis diminuent significativement en automne et atteignent le minimum au printemps (4,1 ± 0,072 μg.100 mg-1), (Fig. ). De même pour les sucres, les mêmes variations sont enregistrées pour les quatre saisons considérées. Le maximum en été (131,3 ± 0,299 μg. 100 mg-1) et le minimum au printemps (80,7 ± 0,277 μg. 100 mg-1) (Fig. ).
Dans les tiges, les concentrations en proline varient dans le sens décroissant depuis l’automne (45,5 ± 0,480 μg. 100 mg-1 de MV) jusqu’au printemps (3,3 ± 0,233 μg. 100 mg-1), en passant par l’été (40,5 ± 0,354 μg. 100 mg-1) et l’hiver (7,5 ± 0,288 μg. 100 mg-1), avec des valeurs significativement plus faibles que pour les feuilles âgées et les feuilles jeunes (Fig. ) de même que pour les sucres solubles (Fig. ) tandis que leurs concentrations diminuent significativement depuis l’été (98,1 ± 0,612 μg. 100 mg-1) jusqu’au printemps (56 ± 0,374 μg. 100 mg-1) en passant par l’automne (94,7 ± 1,160 μg. 100 mg-1) puis l’hiver (72,3 ± 0,661 μg. 100 mg-1).
Les rhizomes montrent les valeurs les plus faibles en proline et en sucres solubles pour les quatre saisons par rapport aux autres organes étudiés; pour la proline, elles varient entre 28,4 μg. 100 mg-1 de MV en été, valeur maximale, et 1,4 μg. 100 mg-1 en hiver, valeur minimale; en automne elle est de 23,6 μg. 100 mg-1 et au printemps 2,0 μg. 100 mg-1 (Fig. ) et pour les sucres entre 84,3 μg. 100 mg-1, valeur maximale, et 25,0 μg.100 mg-1 de MV, valeur minimale (Fig. ).
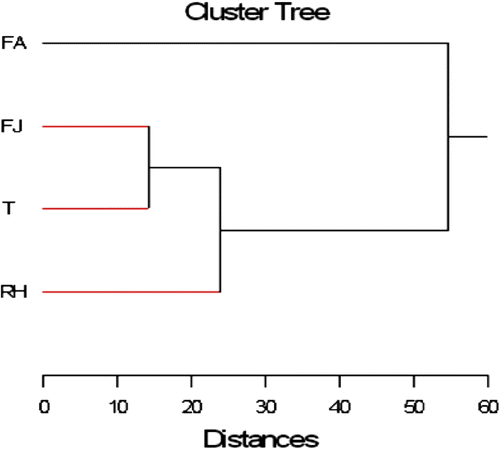
L’étude statistique a démontré que le maximum d’accumulation en proline et sucres solubles a été enregistré au niveau de la partie aérienne et de manière notable dans les feuilles âgées et ceci pendant les périodes les plus sèches et chaudes de l’année d’étude, à savoir l’automne et l’été. Ainsi, le dendrogramme récapitulatif des différents résultats obtenus dans cette étude montre l’individualisation de quatre groupes: le paramètre FA (feuilles âgées) se trouve toujours en «out group» par rapport aux autres paramètres (FJ: feuilles jeunes, T: tiges et RH: rhizome) (Fig. ), c’est-à-dire que les taux en sucres et en proline y sont les plus importants et se démarquent largement par rapport aux autres organes. D’autre part, FJ et T forment un groupe très proche où la distance ne dépasse pas 13. Ce groupe formé recompose avec RH une autre classe très proche par la distance montrant, entre ces derniers groupes, une forte corrélation.
Fig. 7 Dendrogramme interprétatif des matrices des corrélations à partir de l’indice de Pearson des résultats obtenus sur les teneurs en proline et en sucres solubles dosés sur les différents organes de l’oyat aux quatre saisons d’étude. FA: feuilles âgées, Fj: feuilles jeunes, T: tiges, RH: rhizomes. Fig. 7. Interpretative dendrogram of correlation matrix from Pearson index of the results obtained on the contents in proline and soluble sugars assayed in different oyat organs for the four study seasons. FA: old leaves, Fj: young leaves, T: stems, RH: rhizomeFeuilles Feuilles 2.
Discussion
Les métabolites de stress, la proline et les sucres solubles, sont accumulés naturellement dans les feuilles d’un certain nombre d’espèces des sables dunaires (Smirnoff & Stewart, Citation1985). À l’échelle cellulaire, leur accumulation dans le cytosol est accompagnée d’une baisse de la concentration de solutés moins compatibles (les sels) et d’une augmentation du volume d’eau du cytosol assurant le maintien de l’équilibre osmotique (Patel & Vora, Citation1985). La proline fonctionnerait comme stabilisateur de membranes par interaction avec les phospholipides car, si la concentration en sels excède le niveau de tolérance de la plante, des perturbations fonctionnelles apparaissent au niveau de la photosynthèse, par effet du sel dans le stroma des chloroplastes qui perturbe le transport des électrons; la glycolyse et le cycle de Krebs sont eux aussi affectés (Guillaume, Citation2006).
Les sables ont une faible capacité de rétention d’eau, l’accumulation de solutés est donc une réponse adaptative à la sécheresse du substrat. Elle est accentuée durant les deux saisons chaudes et sèches de l’année car les températures sont élevées et les précipitations sont rares, deux variables du climat qui jouent un rôle prépondérant dans la définition des réponses éco-physiologiques des espèces telles Ammophila arenaria. En conditions de stress, la plante contrôle son architecture et réduit la croissance foliaire qui est généralement plus importante que la réduction de la photosynthèse, il en résulte un excédent de sucres produits et donc une augmentation de leur teneur dans les feuilles (INRA, Citation2006). En même temps, exposé à une atmosphère humide et salée, l’oyat subit la salinité plus dans sa partie aérienne que dans sa partie souterraine. La production de ces composés osmoprotecteurs et leur accumulation assure l’ajustement osmotique entre le cytosol et la vacuole des cellules des feuilles plus que celles des tiges et d’une manière moindre de celles des rhizomes, assure une réserve de carbone et d’azote pour la croissance et la stabilisation des protéines et des membranes (Solomon, Citation1985) et assure une protection contre les lésions osmotiques (Schaller, Citation2005).
Par ses teneurs élevées, la proline paraît être l’osmolyte le plus largement distribué sous les conditions de stress dans lesquelles vit notre plante: climatiques (périodes sèches), pédologiques (substrat sableux) et atmosphériques (embruns). Son action serait plus efficace au niveau des feuilles; quand à sa teneur saisonnière, les périodes les plus sèches et les plus chaudes de l’année semblent être plus favorables à l’accumulation de cet acide aminé (été). Au stade âgé de la feuille, la quantité de proline augmente par rapport au stade jeune; cela peut être dû aussi à une plus forte lignification des tissus (Bendimered, Citation1997). Nos résultats rejoignent ceux d’un certain nombre de travaux tels que ceux effectués sur cinq variétés d’orge qui montrent qu’il y a une relation entre la survie des feuilles et leur teneur en proline (Hassani et al., Citation2008; Zid & Grignon, Citation1991). L’accumulation foliaire de la proline est observée chez les plantules de tomate cultivées sous stress hydrique ou salin (Hernandez et al., Citation2000) et chez Nicotiana sylvestis (Dix & Pearce, Citation1981). Selon Belkhodja & Bidai (Citation2007), l’accumulation de proline augmente significativement avec l’augmentation de la concentration de la salinité chez Atriplex halimus, espèce halophile par excellence, et varie en fonction de l’organe, de la nature et de l’intensité du stress (Djerroudi Zidane et al., Citation2010). Aussi, il a été montré chez Arabidopsis thaliana que l’accumulation de la proline observée à la suite d’un traitement salin est la conséquence d’une régulation fine de son métabolisme (Pare et al., Citation2007).
L’accumulation de la proline serait provoquée par la synthèse accrue à partir du glutamate et de l’ornithine (Hare & Cress, Citation1997). En comparant avec la majorité d’autres acides aminés, la synthèse de la proline a un avantage métabolique en tant que produit terminal d’une voie relativement courte et hautement régulée. Sous conditions de stress, le métabolisme des acides aminés est largement altéré et la synthèse des protéines augmente comme une conséquence de ces conditions métaboliques, la synthèse de la proline peut être promue par une augmentation des concentrations des métabolites accompagnée d’une production de précurseurs de proline et peut être la principale cause de l’accumulation de la proline dans les tissus des plantes exposées à des conditions de stress (Silveira et al., Citation2002). En plus, la biosynthèse de la proline peut être associée à la régulation du pH cytosolique ou la production de NADPH pour la stimulation de la voie des pentoses-phosphates (Lutts et al., Citation1996).
Aussi, nos résultats montrent que la quantité de proline accumulée chez l’oyat est assez élevée, ce qui laisse penser que cet osmolyte compte parmi les paramètres biochimiques les plus efficaces permettant la résistance de cette plante aux conditions environnementales extrêmes et s’exprimant de manière plus marquée dans les feuilles les plus âgées. Il migrerait depuis les différents organes vers ces feuilles dans un but de protection et d’augmentation de l’efficacité photosynthétique car certains travaux rapportent que la proline serait synthétisée dans les feuilles et transportée vers les sites de résistance aux agressions (Le Saint, Citation1966; Vezina & Paquin, Citation1982; La Liberte & Paquin, Citation1984) et d’autres signalent que la proline migre vers les feuilles et s’y localise sous contrainte saline; tel est le cas pour le sorgho (Weimberg et al., Citation1986), l’aubergine (Joshi, Citation1984), le coton (Boutelier, Citation1996), la vigne (Imamul Huq & Larher, Citation1984), les fèves (Ait Sadi, Citation1990; Belkhodja, Citation1996; Belkhodja & Benkablia, Citation2000). Bellinger et al. (Citation1989) rapportent que cette accumulation ne représente pas un indicateur de la sensibilité de la plante aux stress mais un indicateur de l’acquisition de la tolérance aux agressions.
Les résultats des dosages des sucres solubles nous ont permis également de noter des valeurs élevées au niveau des différents organes étudiés comparativement à ceux d’autres espèces étudiées en situation de stress hydrique ou salin, avec une variation de la teneur en fonction de l’âge de la plante pendant les quatre saisons de l’année. Les saisons les plus sèches et chaudes et l’âge de la plante conditionneraient l’accumulation de ces carbohydrates. Ammophila arenaria est caractérisée par de fortes capacités d’ajustement osmotique conditionnées par l’accumulation de quantités élevées en sucres solubles qui est sans doute responsable de sa stratégie. Les teneurs en sucres solubles des feuilles, notamment les plus âgées, sont en effet indicatrices du degré de résistance de l’espèce à la salinité: plus la plante accumule de carbo-hydrates, plus elle est résistante (Rathert, Citation1984). L’accumulation accrue des sucres participerait au maintien de l’hydratation des feuilles et l’amplitude accrue serait alors un caractère important du renforcement apparent de la tolérance (Alarcon et al., Citation1994). La teneur élevée des hexoses, essentiellement au niveau des feuilles âgées, pourrait témoigner soit d’une limitation de leur utilisation suite à l’arrêt de croissance (Lawlor, Citation2002), soit d’une force d’appel importante et leur hydrolyse par des enzymes présentes dans ces organes.
La diminution des teneurs en sucres dans les feuilles pendant la période de récupération (hiver) semble rejoindre l’hypothèse émise sur leur réutilisation par la plante, contribuant à la reprise de la croissance. Les sucres solubles participent à l’établissement des relations entre organes sources et organes puits, dépendant de l’âge et du degré d’organisation des tissus. Les différences observées entre les quantités des sucres soulèvent la question de l’importance des processus métaboliques impliqués dans la synthèse, la dégradation, le transport et le stockage des sucres et leur dérégulation en cas de stress environnemental; l’importance quantitative des sucres suggère aussi leur implication dans le processus d’endurcissement des plantes (Aminata Ould El Hadj, Citation2001). Des études menées sur de très nombreuses plantes naturelles et cultivées soumises à différents stress hydriques et salins telles que celles de Adda et al. (Citation2005) effectuées sur le blé et Ardnt et al. (Citation2007) sur Zizyphus rotundifolia ont révélé des accumulations plus ou moins importantes de carbohydrates en réponse à différents degrés de stress et que l’ajustement osmotique s’explique principalement par une importante accumulation des sucres au niveau des différents organes. Ces teneurs pourraient être attribuées aux besoins accentués en carbohydrates et en composés azotés dont la proline, qui représente l’un des composés les plus facilement mobilisables (Zerrad et al., Citation2008).
Les résultats des corrélations établies dans la présente étude pourraient contribuer à faire ressortir la relation existant entre le métabolisme azoté et le métabolisme carboné déjà évoqué par un certain nombre de travaux. Ainsi, la faible synthèse de la proline pourrait être attribuée à une faible activité de la nitrate réductase foliaire due à la présence de NaCl dans le milieu, Aslam et al. (Citation1984) ont montré que le chlorure de sodium affecte l’activité nitrate réductase in vitro et in vivo et sa diminution pourrait être aussi expliquée par une forte synthèse glucidique, la proline étant un acide aminé gluconéoformateur selon Zerrad et al. (Citation2008) et, parallèlement, le catabolisme des glucides dans les feuilles fournit à la plante les composés cétoniques essentiels à la synthèse des composés azotés (Laouar, Citation1977; Schaffer et al., Citation1999).
Conclusion
L’oyat (Ammophila arenaria (L.) Link) est une espèce du littoral halophile, xérophile et psammophile montrant de grandes performances d’adaptabilité aux conditions extrêmes de son milieu tant sur le plan morphologique qu’anatomique et biochimique et offrirait de larges possibilités de valorisation et d’exploitation dans le domaine de la biologie appliquée, et plus précisément en biologie moléculaire, qui devraient permettre d’établir des relations entre les variabilités génotypiques et phénotypiques d’adaptation et de résistance aux différents facteurs de stress.
Il s’agit d’une étude préliminaire de mise en évidence de la variation saisonnière des teneurs en sucres solubles et en proline qui doit être complétée par des études plus élargies sur les sites de synthèse de ces substances, les modalités et la dynamique de leur transport, les sites exacts de leur accumulation et les niveaux les plus optimums permettant à cette espèce la meilleure adaptation et enfin le déterminisme génétique régulant les différentes voies de synthèses et les facteurs conditionnant leur transport, leur utilisation et leur accumulation.
Related Research Data
References
- Adda A., M. Sahnoune, M. Kaid Harche and O. Merah, 2005. Impact of water deficit intensity on durum whent seminal roots. Plant biology and pathology. Science direct. C.R. Biologies, 328, 918-927.
- Aimé , S. and Penven , M.J. 1982 . Le complexe dunaire de Cap Falcon (Oran). Etude morpho-dynamique appliquée et perspectives d’aménagement . Méditerranée , 2 : 2 – 13 .
- Ait Sadi M., 1990. Comportement biochimique de quelques lignées de fève (Vicia faba L.) soumises à la salinité: étude particulière de la proline. Mémoire DES, Univ. Sénia, 65p.
- Alarcon , J.J. , Sanchez-Blanco , M.J. , Bolarin , M.C. , Higazy-Shehata , M. and Allam , A. 1994 . Free proline relation to salinity tolerance of three sugar beet varieties . Egypt. J. Agric. Res. , 73 ( 1 ) : 175 – 189 .
- Aminata Ould Al Hadj K., 2001. Contribution à l’étude des réponses métaboliques de la tomates à la salinité. Thèse Doc. Univ. Rennes (France). 118p.
- AOAC, 1955. Association of official analytical chemists, official methods of analysis. Published by the AOAC., Po.box 540, Washington.
- Ardnt , S.K. , Clifford , S.C. , Wanek , W. , Jones , H.G. and Popp , M. 2007 . Physiological and morphological adaptations of the fruit free Ziziphus rotundifolia in response to progressive drought stress . Tree Physiol , 21 : 705 – 715 .
- Aslam , M. , Huffaker , R.C. and Rain , D.W. 1984 . Early effects of salinity on nitrate assimilation in barbey seedlings . Plant Physiol , 76 : 321 – 325 .
- Belkhodja , M. 1996 . Action de la salinité sur les teneurs en proline des organes adultes de trois lignées de fève (Vicia faba (L.)) au cours de leur développement . Acta Botanica Gallica, Soc. Bot. de France , 143 ( 1 ) : 21 – 28 .
- Belkhodja M., 2000. The Biochimical markers of the satlt plants (Faba Bean) tolerance in arid countries: a Particular Study of the proline Biosynthesis. 8e Conf. Agricul. Dev. Res. Fac. Agrc. Ain Shams Univ. Cairo. Nov. 20-22. Ann. Agrc. Sci, Sp 3, 1041-1051.
- Belkhodja , M. and Benkablia , M. 2000 . Proline of Faba bean (Vicia faba) under salt stress . Egypt. J. Agric. Res. , 78 ( 1 ) : 185 – 195 .
- Belkhodja M. and Y. Bidai, 2007. Analyse de la proline pour l’étude de la résistance d’une halophyte (Atriplex halimus(L.)) à la salinité. http://www.telabotanica.org/page:atriplex_halimus_salinite8p.
- Bellinger , Y. , Bensaouda , A. and Larher , F. 1989 . Physiology breeding of winter cereals for stress environments , Colloque, N°3 France : Monpellier .
- Bendimered F.Z., 1997. Contribution à l’étude de l’Oyat (Ammophila arenaria (L.) Link). Approche écologique. Germination des caryopses, croissance, anatomie et histochimie des feuilles. Th. Magister. Univ. Tlemcen. Algérie. 110p + annexes.
- Bendimered , F.Z. , Mehdadi , Z. and Benhassaini , H. 2007 . Etude de la germination et de croissance foliaire de l’oyat (Ammophila arenaria (L) Link. en conditions contrôlées . Acta Bot. Gallica , 154 ( 1 ) : 129 – 140 .
- Bergman , I. and Loxley , R. 1970 . New spectrophotometric method for the determination of prolinein tissue hydrolysates . Analytical Chemistry , 42 ( 7 ) : 702 – 706 .
- Boutelier E., 1996. Effet du NaCl sur la physiologie du cotonnier (Gossypium hirsutum L.). Son rôle dans l’acquisition de la résistance à la sécheresse. Thèse de Doc. Univ. Paris 6, 142p.
- Dix , P.J. and Pearce , P.S. 1981 . Proline accumulation in NaCl-resistant and sensitive cell pines of Nicotiana sylvestis . Z Pflanzen physiol Bd , 102 : 243 – 248 .
- Djerroudi Zidane O., M. Belkhodja, S. Bissati and S. Hadjadj, 2010. Effet du Stress Salin sur l’accumulation de proline chez deux espèces d’Atriplex: Atriplex halimus L. et Atriplex canescens (Pursh) Nutt. European Journal of Scientific Research, ISSN 1450-216X, 41, (2), 249-260.
- Dubois , M. , Gilles , K.A. , Hamilton , P.A. , Robeg , A. and Smith , F. 1956 . Colometric method for determination of suggars and related substances . Analytical chemistry , 28 ( 3 ) : 350 – 356 .
- Goldhirs , A.G. , Hankamer , B. and Lirs , SH. 1990 . Hydroxy-proline and proline content and cell wall of sunflower, Peanut and cotton under salt stress . Plant Sci , 69 : 27 – 32 .
- Guillaume C., 2006. Arabidopsis thaliana et Thellungiella halophila, plantes modèles dans l’étude du stress salin. Spectrosciences. http://www.spectrosciences.com/spip.php?article23.
- Hare , P.D. and Cress , W.A. 1997 . Metabolic implications of stress induced proline accumulation in plants . Plant Growth Regulation , 21 : 79 – 102 .
- Hassani , A. , DellaL , A. , Belkhodja , M. and KaidHarche , M. 2008 . Effet de la salinité sur l’eau et certains osmolytes chez l’orge (Hordeum vulgare) . European Journal of Scientific Research , 23 ( 1 ) : , 61 – 69 .
- Hernandez S., C. Deleu and R.F. Larhe., 2000. Accumulation de proline dans les tissus foliaires de la tomate en réponse à la salinité. Comptes Rendus Académie des Sciences. Paris, Sciences de la vie/ Life Sciences, 323, 551–557.
- Imamul Huq , S.M. and Larher , F. 1984 . Effect of maintaining a constant Na: Ca ratio on the growth, ion balance and organic solute status of NaCl treated cowpea . Vigna sinensis L.Z. Pflanze. Physiol , 113 : 163 – 176 .
- INRA, 2006. Communiqué Press Info. Juin/Juillet. Service de presse et de relations publiques.
- Joshi , S. 1984 . Effet of salinity stress on organic and mineral constituents in the leaves of Pigeonpea cajanus L . Plant and soil , 82 : 69 – 76 .
- Laouar S., 1977. Caractéristiques écophysiologiques et aspects de l’économie de l’eau de l’olivier (Olea europea (L.)) et de l’oranger (Citrus sinensis (L .) obsbeck). Th. doct. d’état ès. Sciences. p. 247. Univ. Paris VII.
- La Liberte , G. and Paquin , R. 1984 . Effets des basses températures avec ou sans apport d’acide glutamique et de saccharose sur la teneur en proline et la tolérance au gel du blé d’hiver . Physiol. vég , 22 : 305 – 315 .
- Lawlor , D.W. 2002 . Limitations to Photosynthesis in water-stressed leaves: stomatal vs metabolism and the role of ATP . Annals of Botany , 89 : 871 – 885 .
- Leclerc M.C., 1985. Alimentation azotée des plantes des dunes littorales de la méditerranée. Relation avec la disponibilité en eau et en azote du sol .Th. Doc. d’état en Sciences. Acad. Montpellier. Univ. Sc. et tech. Du Languedoc, 240p.
- Le Saint A.M., 1966. Observations physiologiques sur le gel et l’endurcissement chez le chou de Milan. Thèse Doct. Es Sciences, Paris, série A, N° 4669.
- Lutts , S. , Kinet , J.M. and Bouharmont , J. 1996 . Effects of various salts and mannitol on ion and proline accumulation in relation to osmotic adjustment in Rice callus cultures . J. Plant physiol. , 149 : 186 – 195 .
- Maun , M.A. and Baye , P.R. 1989 . The ecology of Ammophila breviligulata Fernon coastal duns systems . Aquatic. Sciences , 4 : 661 – 681 .
- Nguyen , S. and Paquin , R. 1971 . Méthodes d’extraction et de purification des acides aminées libres et des proteines des tissus végétaux J . Chromatography , 61 : 349 – 351 .
- Ottow , E. , Brinker , M. , Fritz , E. , Teichman , T. , Kaiser , W. , Brosche , M. , Kangasjarvi , J. , Jiang , X. and Polle , A. 2005 . Populus euphratia Displays Apoplastic sodium Accumulation, Osmotic Adjustment by decreases in calcium and soluble Carbohydrates and delops leaf succulence under salt stress 1 . Plant Physioology , 139 : 1762 – 1772 .
- Parre , E. , Chars , M.A. , Leprince , A.S. and Thiery , L. 2007 . Calcium signaling via phospholipase cis essential for proline accumulation upon ionic but not non-ionic hyperosmotic stresses in . Arabidopsis thaliana. Plant physiol , 16 : 588 – 599 .
- Patel , J.A. and Vora , A.B. 1985 . Free proline accumulation in drought stressed plants . Plant and soil , 84 : 427 – 429 .
- Quezel P. and S. Santa, 1963. Nouvelle flore de l’Algérie et des régions désertiques méridionales, vol.1. C.N.R.S., Paris VII, 565p.
- Rathert , G. 1984 . Sucrose and Starch content of plant parts as a possible indicators for salt tolerance . Aust. J. Plant Physiol , 11 : 491 – 495 .
- Schaffer , B. , Whiley , A.W. and Searle , C. 1999 . Atmospheric CO2 enrichment, root restriction, photosynthesis, and dry matter partionning in subtropical and tropical fruits crops . Hort. Science , 34 : 1033 – 1037 .
- Schaller , K. 2005 . Proline accumulation in grapevine berries during growth and development . Is it a quality indicator? Bull. OIV , 78 ( 891–92 ) : 321 – 333 .
- Silveira , P. , Meloar , B. and Viegras , R.A. 2002 . Effets salinité induits sur l’assimilation d’azote liée à la croissance aux usines de doliques de Chine . Exp. Bot , 46 : 171 – 179 .
- Smirnoff N. and G.R. Stewart, 1985. Stress metabolites and their role in coastal plants. Plant Ecology. Spinger Neth. (eds.), 62 (1–3), 273–278.
- Solomon , K.H. 1985 . Water-salinity-production functions . Trans. ASA , 28 : 1975 – 1980 .
- Taylor , C.R. 1996 . Proline and water deficit: Ups, Downs and Outs . The Plant Cell , 8 : 1221 – 1224 .
- Vezina , L. and Paquin , R. 1982 . Effects des basses températures sur la distribution de la proline dans les plants de Luzerne . Medicago mediapers. Physiol. Vég , 20 : 101 – 109 .
- Wang , Z. and Stuttle , G.W. 1992 . The role of carbohydrates in active osmotic adjustment in apple under water stress . J. Am. Soc. Hort. Sci. , 117 : 816 – 823 .
- Weimberg , R. , Lerner , H.R. and Poljakooff–Mayber , A. 1986 . Changes in growth and water-soluble solute concentrations in Sorghum bicolor stressed with sodium and potassium salts . Physiol. plant , 62 : 472 – 480 .
- Zerrad , W. , Maataoui , B.S. , Hilali , S. , El Antri , S. and Hamyene , A. 2008 . Etude comparative des mécanismes biochimiques de résistance au stress hydrique de deux variété de blé dur . Lebanese science Journal , 9 ( 2 ) : 1 – 10 .
- Zid E. and C. Grignon, 1991. Les tests de sélection précoce pour la résistance des plantes aux stress: cas des stress salin et hydrique. IIè journées scientifiques du réseau de biotechnologie végétale. L’amélioration des plantes pour l’adaptation aux milieux arides. John Libbey Eurotext (eds.), Paris, 91–108.