
Abstract
Ecological restoration can accelerate the resilience of degraded dry grasslands. Among the ecological restoration techniques used, soil transfer has already given promising results for restoring species-rich plant communities by providing the ability to recreate rapidly a habitat that corresponds to the reference ecosystem (the non disturbed dry grasslands). However, soil transfer is a destructive method, since it involves the use of non-renewable resources such as grassland soils of the donor sites that are the produce of centuries of interactions between climate, plants and animals. In south-eastern France, the plain of La Crau is an example of Mediterranean dry grasslands where the resilience of the steppe vegetation is extremely slow after land use changes which have impacted soil and vegetation. On 7th August 2009, a major oil leak occurred in the steppe center, destroying more than 13 acres of steppe vegetation. As a consequence, in 2010, the polluted soil was excavated and evacuated in a specialised dump. This operation was, then, combined with various in situ experiments of soil transfer, with exclusion of traditional sheep grazing management, to test (i) the importance of respecting the vertical organization of the main soil horizons and (ii) some opportunities to save this non-renewable resource. In May 2011, just after the soil transfer (72,000 tons) was achieved in April 2011 from a nearby quarry which extension had been authorised prior to the oil leak, different quadrats were materialized in the reference steppe vegetation around and in the restored site, with at the soil surface: the organic layer (top-soil, treatment ABC) with or without compaction (treatment ABCnc), the mineral layer (sub-soil, treatment BC) or the altered bedrock only (treatment C). After three years of vegetation monitoring, all the different treatments of soil transfer resulted in a rapid resilience of steppe vegetation in terms of floristic composition and plant species richness. Indeed, in 2013, the treatment with the transfer of mineral layer only at the soil surface, showed no significant difference in terms of plant species richness with the reference steppe plant community. Furthermore, there was a significant higher species richness for the treatments with the transfer of organic layer, compacted or not, in comparison with the reference steppe plant community. In addition, between 2011 and 2013, the natural colonisation by some ruderal opportunistic species was very low. Nevertheless, the reference steppe plant community organization was not restored for none of the different soil transfer treatments, as revealed by the calculation of the Bray-Curtis index. In addition, species abundances in the uncompacted organic layer, mean vegetation height and plant cover were significantly higher, than in the reference steppe plant community. Our results show that after only 3 years, the results of the different soil transfer treatments seem very promising. It appears from this three year monitoring that soil transfer of a mixture of the organic (top soil) and mineral (sub soil) layers, without final compaction, could be sufficient to boost the natural resilience of the typical plants of the reference steppe plant community. It might also be possible to increase the surface where the soil will be spread according to ratios of 1 to 2 or 1 to 3, instead of 1 to 1. This method will allow the protection of the steppe soil that is still a non-renewable resource. Because none of the different soil transfer treatments has allowed for the restoration of the integral steppe plant community, it seems now necessary to reinstall the traditional sheep grazing system with the aim to increase the restoration of the reference steppe plant community spatial organization.
Résumé
La restauration écologique peut accélérer la résilience des pelouses sèches dégradées. Parmi les techniques utilisées, le transfert de sol a, ainsi, déjà permis d’obtenir des résultats encourageants pour rétablir des communautés végétales riches en espèces. En revanche, il s’agit d’une méthode non durable, car destructrice du site donneur. Le 07 août 2009, une importante fuite d’un oléoduc est survenue au milieu de la steppe de La Crau. En 2010, plus de 5,5 ha de végétation ont été détruits par excavation et évacuation en décharge du sol pollué par les hydrocarbures. La restauration de ce site a, alors, été associée à différentes expérimentations in situ de remise en place du sol, en l’absence du pâturage ovin traditionnel (exclos), pour tester l’importance du respect de l’organisation verticale des principaux horizons pédologiques. Après trois années de suivi, nos premiers résultats montrent une forte résilience de la végétation en termes de composition et de richesse spécifique. De plus, peu d’espèces rudérales ont réussi à coloniser le site après transfert du sol. Cependant, aucune des modalités de remise en place du sol n’a pas permis de restaurer l’intégrité de la communauté végétale steppique de référence. Il semble, alors, maintenant nécessaire de remettre en place le système de pâturage ovin ancestral pour permettre la restauration de la structuration spatiale et de la diversité de la végétation steppique.
Introduction
En Europe occidentale, les pelouses sèches possèdent généralement une importante biodiversité (Gibson Citation2009). Lorsqu’un écosystème subit des régimes de stress et de perturbations sur le long terme, il présente souvent une forte richesse spécifique ainsi que des communautés fortement structurées (Weiher and Keddy Citation1999; Alard and Poudevigne Citation2002; Gibson Citation2009). De ce fait, en Europe, les pelouses sèches sont les habitats les plus riches en espèces à une échelle locale et régionale (Willems Citation2001; Poschold and WallisdeVries Citation2002).
Dans le sud-est de la France, la plaine de La Crau est un bon exemple de pelouse sèche méditerranéenne (Molinier and Tallon Citation1950; Devaux et al. Citation1983; Römermann, Tackenberg, and Poschold Citation2005; Tatin et al. Citation2013). Sa végétation steppique résulte d’un régime de stress et de perturbations particulier: l’interaction entre un sol rouge peu profond, un climat méditerranéen et un pastoralisme ancestral a été à l’origine de la création et du maintien d’une communauté végétale unique de par sa richesse spécifique, sa composition et sa structure (Devaux et al. Citation1983; Cherel Citation1988; Badan, Congés, and Brun Citation1995; Henry Citation2009).
Malgré cela, depuis la deuxième moitié du 20e siècle, une diminution drastique des surfaces, de la qualité des habitats et de la biodiversité des pelouses sèches a été mesurée (Dzwonko and Loster Citation1998; Lee et al. Citation2001; Dutoit et al. Citation2011; Adriaens, Honnay, and Hermy Citation2006). Les changements d’usages des sols, la fragmentation du paysage et l’invasion par des espèces non-indigènes (Saunders, Hobbs, and Margules Citation1991) en sont les principales causes. Dès le 16e siècle, la plaine de La Crau a, ainsi, subi d’importants changements d’usages (Masson and Estrangin Citation1928), dus au développement de systèmes d’irrigation pour l’agriculture et à la création de prairies de fauche. A la fin des années 1980, une accélération de sa destruction est constatée par le développement de cultures industrielles (maraîchage, arboriculture fruitière) et par l’installation de structures militaires et industrielles (Deverre Citation1996; Etienne, Aronson, and Le Floc’h Citation1998). Aujourd’hui, cette plaine a perdu quasiment 80 % de sa superficie originelle.
De nombreuses pelouses sèches ont bénéficié de mesures conservatoires (Wells Citation1969). A titre d’exemple, la plaine de La Crau est protégée en partie grâce à la création, en 2001, d’une Réserve Naturelle Nationale sur 7400 ha (Buisson et al. Citation2006). Cependant, ce type d’écosystèmes a souvent une faible capacité de résilience et franchit des seuils de non retour sur des pas de temps historiques (Clewell and Aronson Citation2007). La « résilience naturelle » est de fait limitée par la rémanence des effets des altérations anciennes (élimination des espèces clé, destruction du sol, etc.) et de la modification de la matrice paysagère (fragmentation du paysage). La « régénération naturelle », aux échelles de temps historiques, est alors compromise, même avec la remise en place du système de gestion conservatoire ayant assuré auparavant sa pérennité. A partir de la levée de la contrainte, la résilience correspond à la vitesse de retour d’un écosystème à un état d’équilibre antérieur à la perturbation (Holling Citation1973; Holling and Gunderson Citation2002; Hirst et al. Citation2003, Citation2005). La restauration de ces pelouses peut alors nécessiter des interventions humaines (SER Citation2004), comme dans la plaine de La Crau (Buisson et al. Citation2006; Coiffait-Gombault Citation2011; Jaunatre, Buisson, and Dutoit Citation2012), où la végétation steppique a montré une résilience extrêmement lente après des changements d’usages (Römermann, Tackenberg, and Poschold Citation2005; Buisson et al. Citation2006; Dutoit et al. Citation2013; Dutoit, Jaunatre, and Buisson Citation2013).
La restauration écologique se distingue de la réhabilitation en se fixant pour objectif de rétablir non seulement les processus de l’écosystème, sa productivité et ses services, mais également son intégrité biotique préexistante en termes de composition spécifique et de structure de la communauté (SER Citation2004; Clewell and Aronson Citation2007). Elle permet aussi de comprendre l’organisation et les mécanismes de rétablissement des communautés après une perturbation, par la manipulation des différents filtres qui gouvernent leurs processus de coexistence (Pimm Citation1991; Luh and Pimm Citation1993; Weiher and Keddy Citation1999; Young, Petersen, and Clary Citation2005).
Parmi les nombreuses techniques de restauration écologique testées, le transfert de sol, appliqué en vrac ou en mottes (entières ou fragmentées), a déjà permis d’obtenir des résultats encourageants pour rétablir des communautés végétales riches en espèces (Pywell, Webb, and Putwain Citation1995; Bullock Citation1998; Vécrin and Muller Citation2003; Khiel et al. Citation2010; Jaunatre, Buisson, and Dutoit Citation2014). En effet, cette méthode permet d’agir à la fois sur les filtres biotiques et abiotiques qui contraignent l’établissement et le développement des communautés végétales. Elle consiste à déplacer un sol préalablement prélevé dans un site donneur vers une zone à restaurer. Sous certaines conditions de transport, cette technique permet alors de transférer des éléments physico-chimiques (nutriments, particules fines et grossières, etc.), de la matière organique (litière, humus, etc.), une partie de la pédofaune et de la flore (graines, algues, parties de plantes, etc.), mais aussi de la biomasse microbienne (bactérienne, algale et fongique) du sol (Clewell and Aronson Citation2007). Les plantes pérennes à reproduction végétative ayant des rôles structurants dans la communauté, peuvent aussi être transplantées. Cette technique offre, alors, la possibilité de recréer rapidement un habitat correspondant à l’écosystème de référence, à condition que le site donneur soit proche de celui originellement dégradé et/ou détruit. Cela a déjà été démontré pour certaines pelouses calcaires en Angleterre pour lesquelles un bilan de restauration positif a été mesuré (Bullock Citation1998; Box Citation2003). En revanche, le transfert de sol n’est absolument pas une technique durable puisque la surface donneuse est détruite pour restaurer la surface receveuse et que le sol ne constitue pas une ressource rapidement renouvelable. Cette technique doit donc être limitée à des cas bien particuliers (Bullock Citation1998; Bruelheide and Flintrop Citation2000), lorsque la destruction du sol destinée au prélèvement est programmée indépendamment de la mise en place de l’opération de transfert (McLean Citation2003; Vécrin and Muller Citation2003).
Dans la plaine de La Crau, le 7 août 2009, une superficie de 5,5 ha a totalement été détruite par la rupture d’un oléoduc en plein cœur d’une Réserve Naturelle Nationale (Wolff Citation2014). La surface, le sous-sol et la nappe phréatique ont été pollués par le déversement de plus de 5 400 m3 de pétrole brut. La réhabilitation écologique a alors consisté à excaver et à évacuer 72 000 hydrocar-bures tonnes de sol pollué vers une décharge spécialisée. La restauration écologique a, ensuite, à l’aide d’engins de travaux publics, permis de combler la cavité créée par le transfert d’un même sol, avec un rapport 1/1, destiné à être détruit suite à l’extension d’une carrière autorisée avant l’accident. Ce transfert de sol est très original de par son exécution effectuée pour favoriser au maximum le développement d’une végétation steppique identique à celle détruite par la pollution aux hydrocarbures, l’excavation et la mise en décharge du sol (Dutoit Citation2011).
Dans cette étude, nous avons mesuré la résilience de la végétation steppique en fonction de différents traitements de remise en place du sol. Pour cela, notre premier objectif a été (1) de mesurer l’effet de ce transfert de sol sur la composition, la richesse spécifique et la structure de la végétation en comparant la zone de restauration avec la communauté steppique de référence située autour du site restauré. Notre second objectif a été (2) de tester l’importance du respect de l’organisation verticale des principaux horizons pédologiques (A1, (B), C) pour vérifier s’il est nécessaire de tous les reconstituer ou s’il est plus pertinent de ne pas remettre l’horizon organique A1, afin de rendre le sol plus oligotrophe. L’hypothèse est que le transfert de l’horizon organique A1 pourrait favoriser l’installation des espèces non caractéristiques de l’écosystème de référence (espèces rudérales, cosmopolites, ubiquistes, exotiques envahissantes, etc.) qui se développent à partir de la banque et de la pluie de graines, ce qui permettrait d’économiser le sol en le transférant à des rapports plus élevés, par exemples à 1/2 ou 1/3.
Enfin, notre troisième objectif a été (3) de tester l’effet du compactage de la surface du sol pour favoriser l’installation des espèces cibles de la restauration. Notre hypothèse était alors que cela permettrait de réduire un foisonnement du sol et une oxydation concomitante de la matière organique qu’il contient. Ces processus peuvent en effet être responsables in fine d’une accélération de la minéralisation de l’azote et, donc, favoriser l’installation d’espèces nitratophiles au détriment des espèces stress-tolérantes, caractéristiques de la végétation steppique de référence.
Matériels et Méthodes
Site atelier – La plaine de La Crau
Notre étude a été menée dans la plaine de La Crau, localisée dans le sud-est de la France (Bouches-du-Rhône) (Figure ). Cet ancien delta fossile a été formé par les dépôts de la Durance pendant la période du Pléistocène lorsque cette rivière était encore un fleuve qui se déversait directement dans la mer Méditerranée (Rainaud Citation1893; Colomb and Roux Citation1978; Devaux et al. Citation1983). Sa topographie est extrêmement plate, avec une surface de sol recouverte à plus de 50 % par des galets siliceux charriés depuis le massif alpin. Entre 40 cm et 60 cm de profondeur dans le sol, un poudingue, composé de galets enrobés dans une matrice de carbonates de calcium, empêche les racines des plantes d’atteindre la nappe phréatique (Rainaud Citation1893; Colomb and Roux Citation1978; Römermann, Tackenberg, and Poschold Citation2005). Le climat méditerranéen est sec et venteux avec un fort ensoleillement durant l’année: la pluviométrie est d’environ 540 mm/an, principalement au printemps et à l’automne; le mistral souffle à plus de 50 km/h pendant 110 jours/an et le soleil est présent pendant plus de 3000 h/an (Devaux et al. Citation1983). En combinaison avec le sol et le climat, un pâturage ovin, attesté depuis au moins 2000 ans, parcourt et façonne une communauté végétale exceptionnelle par sa richesse spécifique, sa composition et sa diversité (Cherel Citation1988; Badan, Congés, and Brun Citation1995; Henry Citation2009).
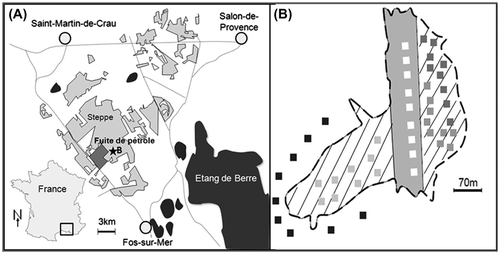
Figure 1. A, Situation géographique de la plaine de La Crau en France et du site où a eu lieu la fuite de pétrole; B, Schématisation du site restauré (hachures), de la zone de passage des canalisations (bande grise), de la steppe de référence (fond blanc) et positionnement des quadrats (100 m²) correspondants aux différents traitements de transfert sol effectués par camions (traitement C: blanc; traitement BC: gris clair; traitement ABCnc : gris foncé; horizon ABC: GRIS; steppe de référence: gris très foncé).
Figure 1. A, Location of the plain of La Crau in France and location of the site where there was the oil spill, B, Schema of the restored site (hatched), zone crossing by oil pipes (gray line), reference steppe (white) and positioning of quadrats (100 m²) corresponding to different treatments of soil transfer realized by trucks (treatment C: white; treatment BC: light gray; treatment ABCnc: dark gray; treatment ABC: gray; reference steppe: very dark gray).
La végétation qui recouvre entre 50 % et 80 % de la surface du sol, comprend en moyenne 30 à 40 espèces végétales par mètre carré (Römermann, Tackenberg, and Poschold Citation2005) et est composée à 50 % par des espèces annuelles. Des espèces pérennes, telles que Brachypodium retusum (Pers.) P. Beauv. (Poaceae), représentant 50 % de la biomasse herbacée estimée à 2,11 tonnes de matière sèche/ha/an, et Thymus vulgaris L. (Lamiaceae) dominent et structurent la communauté (Buisson et al. Citation2006). Les autres espèces végétales les plus caractéristiques sont Asphodelus ayardii Jahand. & Maire (Xanthorrhoeaceae), Linum trigynum L. (Linaceae) ou encore Stipa capillata L. (Poaceae) (Molinier and Tallon Citation1950; Devaux et al. Citation1983).
Le sol de la steppe est un sol fersialitique rouge méditerrannéen tronqué (Duchaufour Citation1984), encore appelé aujourd’hui Fersiasol leptique tronqué (AFES Citation2008), constitué par trois horizons pédologiques principaux: un horizon organique A1 correspondant à un horizon mixte composé de matière organique (moins de 30 %) mélangée à de la matière minérale, et à une subdivision de l’horizon A de surface; un horizon (B) constituant l’horizon « structural » ou d’altération, différant de la roche-mère par son degré d’altération plus fort (présence de Fe2O3 libre) et de l’horizon A1 de surface par sa structure différente, et un horizon C, correspondant au matériau originel aux dépens duquel se sont formés A et (B) (Duchaufour Citation1984).
Site d’étude et techniques de restauration
Notre site d’étude correspond aux 5,5 ha détruits par une fuite de pétrole brut le 7 août 2009 (43° 31’ 36.77’’ N, 4° 53’ 04.50’’ E) (Figure ). Après excavation et évacuation de 72 000 tonnes de terres polluées, de janvier à avril 2011, un transfert de sol direct sans stockage a été effectué, avec un rapport 1/1, à l’aide de camions, en reconstituant les trois principaux horizons pédologiques. Ce sol a été prélevé dans la steppe intacte au niveau d’une carrière située à quelques kilomètres de distance.
Pour les quatre traitements testés, neuf parcelles de 100 m² ont été matérialisées avec la remise en place : (1) des horizons C + (B) + A1 (traitement ABC), (2) des horizons C + (B) + A1 sans compactage en surface (traitement ABCnc), (3) des horizons C + (B) (traitement BC), et (4) seulement de l’horizon C (traitement C).
Après le transfert du sol au printemps 2011, l’ensemble de la surface du site a été compacté à l’aide d’un rouleau compresseur à l’exception des neuf parcelles du traitement sans compactage (Figure ). À cause d’un problème logistique et de pluviométrie, la mise en place des quadrats du traitement C + (B) n’a pu être achevée que deux mois après celles des autres traitements à la fin du mois d’avril 2011.
Figure 2. A, Photographie du site où a eu lieu la fuite de pétrole après l’excavation du sol pollué correspondant à la phase de réhabilitation écologique (premier plan) et de la steppe de référence (arrière plan); B, Photographie représentant la collecte du sol dans le site donneur (à droite) et de la steppe intacte (à gauche); C, Photographie représentant le transfert de sol sur le site receveur correspondant à la phase de restauration écologique (arrière plan) et de deux quadrats pour le suivi de la végétation déjà mis en place (premier plan); D, Photographie représentant les principaux horizons pédologiques en cours de transfert (horizon organique A1; horizon minéral (B); cailloutis C) et de la roche mère R.
Figure 2. A, View of the site in 2010 where the leak occured and after the excavation of the polluted soil corresponding at the depollution step (forefront); and view of the reference steppe (background); B, collecting of soil in 2011 in the donor site (at right) and the undisturbed steppe (at left); C soil transfer in march 2011 on the receive site (background) and disposition of two quadrats for the vegetation survey (foreground); D, main soil layers being transferred (top soil A1, subsoil (B), altered bedrock C) and the unaltered bedrock.

Analyses de la végétation
Pour analyser la dynamique de régénération de la végétation, neuf quadrats de 4 m² ont été délimités au centre des parcelles de 100 m² afin d’éviter tout effet de bordure dans tous les traitements. De même les quadrats dans la steppe de référence ont été positionnés à l’ouest du site et à au moins 20 m de la lisière (Buisson et al. Citation2006). La surface des parcelles de 100 m² a été choisie pour être nettement supérieure à l’aire minimale des formations végétales herbacées de la plaine de La Crau, soit environ 20 m2 (Devaux et al. Citation1983). Seule la steppe de référence a été pâturée au cours des années de suivi pour assurer au maximum la régénération des espèces végétales sur le site restauré en évitant des effets de surpâturage les premières années.
Dans chaque quadrat, la présence et l’abondance des espèces végétales ont été enregistrées aux mois de mai 2011, 2012 et 2013, correspondant à la période de floraison maximale de la végétation de la plaine de La Crau (Bourrely et al. Citation1983). Un coefficient d’abondance-dominance de Braun-Blanquet a été défini pour toutes les espèces végétales relevées (Braun-Blanquet et al. Citation1952). Celles-ci ont ensuite été classées comme « espèces cibles » ou comme « espèces non cibles ». Les espèces cibles sont les espèces les plus caractéristiques de la steppe de référence et qui répondent à nos objectifs de restauration tandis que les espèces non cibles sont celles de type rudéral, cosmopolites, ubiquistes, exotiques envahissantes, etc. (Devaux et al. Citation1983). Le pourcentage de recouvrement total de la végétation a été estimé visuellement et la hauteur moyenne a été mesurée dans chaque quadrat, à l’aide d’un mètre pliable, afin de mesurer la densité de la végétation entre les zones de traitements et la steppe de référence. Si le taux de recouvrement et la hauteur moyenne de la végétation sont significativement supérieurs dans les quadrats, correspondant aux traitements de restauration, cela indique que la physionomie de la steppe de référence n’est pas atteinte et que la réintroduction du pâturage sur le site en cours de restauration est nécessaire.
Analyses des teneurs en carbone et en azote totaux
Des échantillons de sol ont été prélevés et analysés juste après le transfert de sol en avril 2011 dans les traitements ABCnc, BC et C. Pour cela, cinq échantillons de sols de 200 g ont été extraits à 0–10 cm de profondeur dans cinq parcelles de 100 m² sélectionnées au hasard parmi les neuf parcelles de chaque traitement et dans la steppe de référence. Chaque échantillon a été constitué en mélangeant trois autres sous-échantillons prélevés aléatoirement dans chaque parcelle de 100 m² pour avoir une meilleure représentativité. Après séchage et tamisage (2 mm), les teneurs en carbone organique total (Corg) et en azote (N) total ont été mesurées en suivant des méthodes standards (Baize Citation2000) par le laboratoire d’analyses des sols de l’INRA (Institut National de la Recherche Agronomique) d’Arras.
Analyses statistiques des données
Pour tester l’effet du transfert de sol sur la régénération de la végétation du site restauré, nous avons comparé la composition, la richesse spécifique (totale, en espèces cibles et en espèces non cibles) et la structure de la communauté végétale entre le traitement ABC et la steppe de référence. La structure de la communauté végétale est mesurée à l’aide des coefficients d’abondance/dominance des espèces, de la diversité spécifique, de la hauteur moyenne et de l’estimation du recouvrement de la végétation.
Pour vérifier ensuite l’importance du respect de l’organisation verticale des principaux horizons pédologiques, ces mêmes paramètres ont été comparés entre les traitements ABC, BC, C et la steppe de référence et entre les traitements ABC et ABCnc pour mettre en évidence un éventuel effet du compactage de la surface du sol sur le rétablissement de la végétation steppique. Pour vérifier les niveaux trophiques des traitements ABC, BC, C et ceux de la steppe de référence, nous avons comparé leurs teneurs en Corg et en N totaux.
L’interaction entre les trois années de suivi et les différences traitements n’a pas été calculée, puisque les conditions paramétriques n’ont pas pu être validées. Les analyses statistiques ont, donc, toutes été utilisées pour comparer les différents traitements entre eux au cours de la troisième année de suivi (2013), ou pour comparer les trois années de suivi (2011, 2012, 2013) dans un même traitement.
Les caractéristiques de la composition et des abondances des espèces végétales dans tous les traitements et dans la steppe de référence ont été ordonnées à l’aide d’une NMDS (Non-metric Multidimensional Scaling) basée sur la distance de Bray-Curtis pour l’année 2013 (Borcard, Gillet, and Legendre Citation2011). La distance de Bray-Curtis (Bray and Curtis Citation1957) est un index de dissimilarité utilisé pour mesurer la distance entre les abondances des espèces végétales présentes dans les traitements et dans la steppe de référence. Elle varie entre 0 et 1, 0 signifiant que les abondances des espèces végétales sont strictement identiques et 1 signifiant qu’elles sont strictement différentes entre les traitements et la steppe de référence.
La diversité spécifique a été calculée grâce aux indices de Shannon et d’Équitabilité qui renseignent sur la distribution des espèces dans chaque traitement. L’indice de Shannon est égal à zéro lorsque la population est représentée par une seule espèce et devient maximal lorsque la population est répartie de manière homogène entre toutes les espèces. L’indice d’Équitabilité, basé sur l’indice de Shannon, varie entre 0 et 1. Sa valeur est maximale lorsque la répartition de la population est homogène entre toutes les espèces et elle est minimale lorsqu’une espèce domine (Pielou Citation1969).
Pour comparer le recouvrement et la hauteur moyenne de la végétation, les richesses spécifiques, les indices de Shannon et d’Équitabilité, les distances de Bray-Curtis, ainsi que les teneurs en Corg et N totaux, nous avons utilisé un test ANOVA lorsque les données suivaient une loi normale (Test de Lilliefors), suivi par des tests post-hoc de Tukey quand une différence significative était détectée et un test non-paramétrique de Kruskal-Wallis lorsque les données ne suivaient pas une loi normale, suivi par un test non-paramétrique de Mann-Whitney-Wilcoxon, dans lequel la valeur de p était ajustée selon la méthode de Benjamini-Hochberg (Benjamini and Hochberg Citation1995) quand une différence significative était détectée.
Toutes les analyses statistiques ont été effectuées en utilisant le logiciel R version 2.15.2. (The R Foundation for Statistical Computing 2012) avec ses packages pour les analyses univariées, et les packages additionnels ade4 (Chessel, Dufour, and Thioulouse Citation2004; Dray and Dufour Citation2007) et vegan (Oksanen et al. Citation2012) pour les analyses multivariées.
Résultats
Effet du transfert de l’intégralité du sol
Sur l’ordination NMDS, le traitement ABC se distingue nettement des relevés de la steppe de référence. Celle-ci est caractérisée par la dominance de l’espèce pérenne, Brachypodium retusum (Pers.) P. Beauv. (Poaceae) et par la présence d’espèces annuelles telles que Asterolinon linum-stellatum (L.) Duby (Primulaceae), Aira cupanania Guss. (Poaceae) ou encore Asphodelus ayardii Jahand & Maire (Xanthorrhoeaceae). Le traitement ABC est, quant à lui, caractérisé par de nombreuses espèces cibles telles que Neatostema apulum (L.) I.M. Johnst. (Boraginaceae), Evax pygmaea (L.) Brot. (Asteraceae) ou encore Plantago bellardii All. (Plantaginaceae), mais également par quelques espèces non-cibles telle que Cynodon dactylon (L.) Pers. (Poaceae). De plus, le brachypode le plus corrélé à ce traitement est l’espèce annuelle Brachypodium distachyon (L.) P. Beauv. (Poaceae) et non l’espèce pérenne Brachypodium retusum (Pers.) P. Beauv. (Poaceae) (Figure -A).
Figure 3. Analyse de la composition et des abondances des espèces végétales de chaque traitement et de la steppe de référence par une ordination NMDS basée sur 45 échantillons de végétation au printemps 2013. Le graphique A montre les échantillons de 2012 regroupés par traitement à l’aide de polygones: traitement C (Trait C, blanc), traitement BC (Trait BC, gris clair), traitement ABCnc (ABCnc, gris foncé), traitement ABC (Trait ABC, gris) et la steppe de référence (Steppe de référence, gris très foncé). Pour plus de clarté, seules les espèces végétales les mieux corrélées sont inscrites littéralement Le graphique B montre la dynamique de la végétation pour chaque modalité, selon la position de leur barycentre. Les lignes pointillées représentent la succession de la végétation de 2011 à 2012 et les flèches représentent la succession de la végétation de 2012 à 2013.
Figure 3. Composition and abundances analyses of plant species in each treatment and in the reference steppe with a NMDS ordination based on 45 vegetation samples. The A plot shows only the 2013 samples grouped by treatment with polygons: treatment C (Trait C, blanc), treatment BC (Trait BC, gris clair), treatment ABCnc (ABCnc, gris foncé), treatment ABC (Trait ABC, gris) and the reference steppe (Steppe de référence, gris très foncé). To clarify, only the most correlated plant species, at two first axes, are literally inscribed. The B plot shows the vegetation dynamic for each treatment, according the position of their barycenter during the tree years of the vegetation survey.
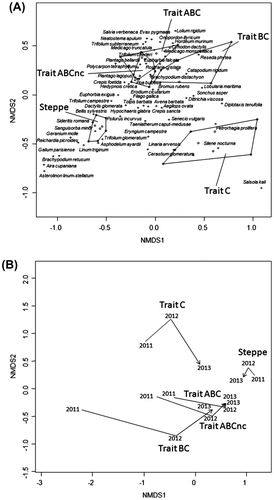
Les trajectoires des barycentres montrent que la steppe a eu une dynamique assez lente de 2011 à 2013 avec très peu de changements dans sa composition et les abondances des espèces végétales, tandis que le traitement ABC a eu une forte dynamique de 2011 à 2012 et une dynamique plus lente de 2012 à 2013 (Figure -B).
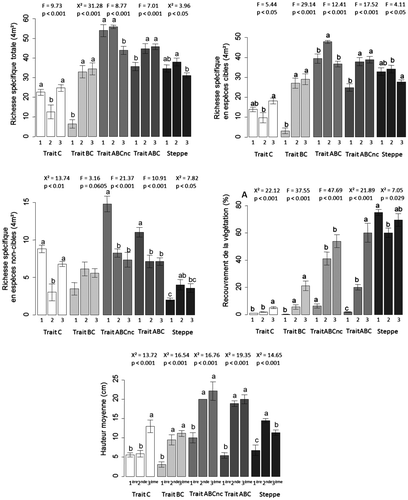
Sur le traitement ABC, la richesse spécifique totale de la végétation a significativement augmenté entre 2011 et 2012 (Figure -A), du fait de l’augmentation du nombre d’espèces cibles (Figure -B) et de la diminution du nombre d’espèces non cibles (Figure -C). En revanche, dans la steppe de référence, la richesse spécifique totale ne présentait pas de différence significative entre 2011 et 2013, même si un nombre d’espèces significativement plus élevé a été mesuré en 2012 (Figure -A). Sur le traitement ABC, le pourcentage de recouvrement de la végétation a également augmenté de façon significative de 2011 à 2013, alors que celui de la steppe de référence ne présentait pas de différence entre les années 2011 et 2013, mais était plus faible en 2012 (Figure -D). Enfin, entre 2011 et 2012, la hauteur moyenne de la végétation a significativement augmenté sur le traitement ABC ainsi que celle de la steppe de référence, mais celle-ci a ensuite diminué entre 2012 et 2013 (Figure -E).
Figure 4. Moyennes et erreurs standards de la richesse spécifique totale (4 m²) (A), de la richesse spécifique en espèces cibles (cm) (B), de la richesse spécifique en espèces non cibles (4 m²) (C), du recouvrement (%) (4 m²) (D) et de la hauteur moyenne de la végétation (E) au cours des trois années de suivi (2011, 2012 et 2013) pour chaque modalité de traitement de transfert de sol et pour la steppe de référence: traitement ABCnc (Trait ABCnc, gris foncé), traitement ABC (Trait ABC, gris), traitement BC (Trait BC, gris clair), traitement C (Trait C, blanc) et la steppe de référence (Steppe, gris très foncé). Les valeurs de F, de X² et de p des ANOVAs ou des tests de Kruskal-Wallis appliquées pour comparer chaque modalité entre années sont reportées au dessus des barres. A l’intérieur de chaque modalité, les barres partageant une lettre commune ne montrent aucune différence significative selon les tests post-hoc de Tukey ou de Mann-Withney-Wilcoxon.
Figure 4. Means and error standards of total species richness (4 m²) (A), target specifies richness (4 m²) (B), non-target species richness (4 m²) (C), vegetation cover (%) (D) and mean height (cm) (E) during three years of vegetation survey (2011, 2012 et 2013) for each treatment in spring 2013: treatment C (Trait C, white), treatment BC (Trait BC, light gray), treatment ABCnc (Trait ABCnc, gray), treatment ABC (Trait ABC, dark gray) and the reference steppe (Steppe, very dark gray). F and p values of ANOVA tests or X² and p values of Kruskal-Wallis tests, performed within each treatment, to compare years, are reported above each bar. Within a treatment, bars sharing common letters don’t have significant differences according to Tukey post-hoc test or Mann-Withney-Wilcoxon test.
Au printemps 2013, sur le traitement ABC, la richesse spécifique en espèces cibles et non cibles, les indices de Shannon et d’Équitabilité, ainsi que la hauteur moyenne de la végétation étaient significativement plus élevés que dans la steppe de référence. Cependant, il n’y avait aucune différence significative concernant le pourcentage de recouvrement de la végétation (Tableau ).
Tableau 1. Caractéristiques de la végétation dans chaque traitement de remise en place du sol et dans l’horizon A1 de la steppe de référence au printemps 2013, après 3 années de suivi: traitement C (Trait C), traitement BC (Trait BC), traitement ABCnc (ABCnc), traitement ABC (Trait ABC) et la steppe de référence (Steppe). Les valeurs mesurées correspondent à la moyenne ± l’erreur standard; df, F, X², p correspondent au degré de liberté, à la valeur de F, à la valeur du chi-carré et à la valeur de p résultant du test ANOVA ou du test de Kruskal-Wallis testant l’effet de chaque modalité sur chaque variable. Dans une même rangée, les cases partageant une lettre commune ne montrent aucune différence significative de leurs valeurs selon le test post-hoc de Tukey ou de Mann-Withney-Wilcoxon.
Table 1. Plant community characteristics for each treatment after 3 years, namely only for spring 2012: treatment C (Trait C), treatment BC (Trait BC), treatment ABCnc (Trait ABCnc), treatment ABC (Trait ABC) and the reference steppe (Steppe). The measured values are means ± standard errors; df, F, X², p correspond to the degree of freedom, the F value, the X² value and the p value resulting from ANOVA tests or Kruskal-Wallis tests to test the effect of each treatment on each variable. Within a row, two cases with a common letter have significantly different values according to Tukey post-hoc tests or Mann-Whitney-Wilcoxon tests.
Importance du respect de l’organisation verticale des horizons pédologiques
En 2011, les taux de C total et de N total mesurés ne sont pas significativement différents entre le traitement ABCnc et la steppe de référence, mais sont significativement plus faibles dans le traitement BC et encore plus faible dans le traitement C (Tableau ).
Tableau 2. Analyses chimiques des teneurs en carbone organique total et en azote total dans trois traitements et dans la steppe de référence juste après le transfert de sol en avril 2011 : traitement C (Trait C), traitement BC (Trait BC), traitement ABCnc (ABCnc), traitement ABC (Trait ABC) et la steppe de référence (Steppe de référence). Les valeurs mesurées correspondent à la moyenne ± l’erreur standard; df, F, X², p correspondent au degré de liberté, à la valeur de F, à la valeur du chi-carré et à la valeur de p résultant du test ANOVA ou du test de Kruskal-Wallis testant l’effet de chaque modalité sur chaque variable. Dans une même rangée, les cases partageant une lettre commune ne montrent aucune différence significative de leurs valeurs selon le test post-hoc de Tukey ou de Mann-Withney-Wilcoxon.
Table 2. Soil analyses of total organic carbon and total nitrogen contents for each treatment just after the soil transfer in april 2011: treatment C (Trait C), treatment BC (Trait BC), treatment ABCnc (Trait ABCnc), treatment ABC (Trait ABC) and the reference steppe (Steppe). The measured values are means ± standard errors; df, F, X², p correspond to the degree of freedom, the F value, the X² value and the p value resulting from ANOVA tests or Kruskal-Wallis tests to test the effect of each treatment on each variable. Within a row, two cases with a common letter have significantly different values according to Tukey post-hoc tests or Mann-Whitney-Wilcoxon tests.
Sur l’ordination NMDS, les traitements ABC et BC sont extrêmement proches; ils sont corrélés aux mêmes espèces en termes de composition et d’abondance. Cependant, le traitement C se distingue nettement des autres traitements et de la steppe de référence. Il est caractérisé par des espèces cibles telles que Petrorhagia prolifera (L.) P.W. Ball & Heywood (Caryophyllaceae), Silene nocturna L. (Caryophyllaceae) ou encore Cerastium glomeratum Thuill. (Caryophyllaceae), mais également par quelques espèces non cibles telles que Salsola kali L. (Caryophyllaceae) et Diplotaxis tenuifolia (L.) DC. (Brassicaceae) (Figure -A).
Les trajectoires des barycentres des traitements BC et C montrent une forte dynamique de la végétation au cours des trois années de suivi, mais plus élevée entre 2011 et 2012 pour le traitement BC et entre 2012 et 2013 pour le traitement C (Figure -B).
Sur le traitement BC, la richesse spécifique totale a significativement augmenté entre 2011 et 2012 du fait d’un nombre plus élevé d’espèces cibles. Pour le traitement C, elle n’était pas significativement différente entre 2011 et 2013, mais elle était plus faible en 2012 (Figure -A et -B). Le pourcentage de recouvrement et la hauteur moyenne de la végétation ont eu la même évolution sur les traitements BC et ABC, avec une augmentation significative entre 2011 et 2012, mais sur le traitement C, ils ont augmenté seulement entre 2012 et 2013 (Figure -D et -E).
Au printemps 2013, par rapport à la steppe de référence, la richesse spécifique totale sur le traitement BC n’était pas significativement différente, même si le nombre d’espèces non cibles était supérieur. Sur le traitement BC, la richesse spécifique en espèces cibles étaient significativement plus faible que celle sur le traitement ABC mais similaire à celle de la steppe de référence. Les indices de Shannon et d’Équitabilité ainsi que la hauteur moyenne de la végétation ne montraient pas non plus de différence significative avec la steppe de référence. En revanche, le pourcentage de recouvrement de la végétation était significativement plus faible.
Ces mêmes paramètres étaient significativement plus faibles sur le traitement C comparés à la steppe de référence, mais également aux traitements BC et ABC. Seule la hauteur moyenne de la végétation n’était pas significativement différente de celle mesurée dans la steppe de référence, et la richesse spécifique en espèces non cibles était similaire à celle mesurée sur les autres traitements. Le traitement C avait une dissimilarité plus élevée avec la steppe de référence que le traitement BC (Tableau ).
Effet du compactage de la surface du sol
Sur l’ordination NMDS, le traitement ABCnc se superpose avec le traitement ABC. Ces deux traitements sont donc également extrêmement proches en matière de composition et d’abondances des espèces végétales (Figure -A). Les dynamiques de la végétation révélées par la position de leurs barycentres étaient également similaires entre 2011 et 2013, mais la végétation avait une dynamique plus forte entre 2012 et 2013 sur le traitement ABCnc que sur le traitement ABC (Figure -B).
La richesse spécifique totale a significativement augmenté entre 2011 et 2012 sur le traitement ABC dû à un nombre plus élevé d’espèces cibles, puis elle s’est stabilisée entre 2012 et 2013 alors que celle sur le traitement ABCnc diminuait de manière significative à cause d’une perte d’espèces cibles (Figure -A et -B). Le nombre d’espèces non cibles n’a pas changé significativement entre 2011 et 2013 sur les traitements ABCnc et ABC, alors qu’il a diminué entre 2011 et 2012 (Figure -C). La hauteur moyenne de la végétation a eu la même évolution sur ces deux traitements, c’est-à-dire une augmentation significative de 2011 à 2012, puis une stabilisation entre 2012 et 2013 (Figure -E). En revanche, le pourcentage de recouvrement de la végétation n’a cessé d’augmenter entre 2011 et 2013 sur le traitement ABCnc, alors qu’il s’est maintenu entre 2012 et 2013 sur le traitement ABC.
Au printemps 2013, aucune différence significative n’a été détectée entre le traitement ABCnc et le traitement ABC pour tous les paramètres de la végétation mesurés (Tableau ).
Discussion
Dans ce travail, nous avons testé l’effet d’un transfert de sol printanier, sans stockage, sur la régénération de la végétation steppique dans la plaine de La Crau. L’originalité de ce transfert réside entre autres dans la vérification de l’importance du respect de l’organisation verticale des principaux horizons pédologiques (A1, (B), C) et de la nécessité de compacter le sol afin de choisir le meilleur compromis entre l’efficacité de cette technique de restauration écologique et sa durabilité écologique, en économisant le sol et en limitant le nombre de passages des engins de travaux publics.
Effet du transfert de sol
Le transfert de sol a permis d’obtenir sur le court terme des résultats très encourageants sur le rétablissement de la communauté végétale steppique. Nous obtenons un taux de transfert de 100 % des espèces végétales de la steppe deux ans seulement après la fin des travaux. En Grande-Bretagne, après un transfert de mottes dans des pelouses sèches, sur 18 expérimentations Bullock et al. (1998), ont obtenu un taux de transfert de 54 % à 91 % après trois à sept ans, et Trueman, Mitchell, and Besenyei (Citation2007) ont atteint 100 % mais seulement après cinq années.
Dans notre cas, après trois années, la communauté végétale restaurée est constituée à plus de 50 % par des espèces annuelles, comme dans la steppe de référence (Molinier and Tallon Citation1950; Devaux et al. Citation1983; Römermann, Tackenberg, and Poschold Citation2005). La grande richesse spécifique de la communauté restaurée, composée à 84 % par des espèces cibles, peut être expliquée par la germination de graines viables contenues dans la banque de graines du sol transféré. Contrairement à un premier transfert de sol réalisé dans la plaine de La Crau, le succès de cette opération réside également dans le fait que peu d’espèces non désirées, par rapport à nos objectifs de restauration, se sont installées. Jaunatre, Buisson, and Dutoit (Citation2014) ont mesuré que le transfert de sol n’avait pas affecté les teneurs du sol en éléments nutritifs, contrairement à ce qui peut être obtenu après un transfert de mottes (Trueman, Mitchell, and Besenyei Citation2007). On peut, donc, supposer dans notre cas que le transfert de sol n’a pas augmenté suffisamment sa fertilité (notamment par oxydation de la matière organique) pour permettre l’installation d’espèces nitratophiles dans ce traitement.
La diversité spécifique, bien que supérieure à celle de la steppe de référence, reste relativement faible. En effet, la communauté végétale du site restauré semble moins hétérogène que celle de la steppe, et légèrement mieux répartie entre les espèces. Cela peut s’expliquer par le fait que l’espèce structurante de la steppe, Brachypodium retusum (Pers.) P. Beauv. (Poaceae), ne domine toujours pas à l’issue des observations. Cependant, les abondances des espèces végétales ne sont pas dissimilaires à celles de la steppe de référence, ce qui représente un résultat innovant comparé à un transfert de sol par épandage selon un rapport 1/3 sur une friche dans la plaine de La Crau où les abondances des espèces végétales se sont révélées très différentes de celles de l’écosystème de référence même trois ans après la fin des travaux (Jaunatre, Buisson, and Dutoit Citation2014).
Le pourcentage de recouvrement de la végétation, bien qu’équivalent à celui de la steppe de référence en 2013, n’a cessé d’augmenter significativement au cours des trois années de suivi. De plus, la végétation possède également une hauteur moyenne bien supérieure à celle de la steppe de référence. La végétation semble donc suffisamment régénérée pour permettre le retour d’un système de gestion qui maîtrisera son développement. Le retour du pâturage ovin est donc nécessaire (Gibson and Brown Citation1992; Bullock Citation1998; Stroh et al. Citation2002; Henry Citation2009). Nous faisons, en effet, l’hypothèse que les différences mesurées entre 2011 et 2012/2013 dans la steppe de référence sont dues aux variations interannuelles de pression de pâturage.
Importance du respect de l’organisation verticale des horizons pédologiques
Au printemps 2013, avec la reconstitution des horizons pédologiques C et (B) (transfert des horizons C et (B)) jusqu’à la surface du sol, la communauté végétale présente un taux d’espèces cibles inférieur de 20 % à celui de la steppe de référence. Néanmoins, la proportion des espèces cibles par rapport au nombre d’espèces total atteint quand même 84 %. Lorsque les horizons pédologiques (B) et A1 ne sont pas remis en place, la richesse spécifique de la végétation reste plus faible que celles des traitements BC (transfert des horizons C et (B)) et ABC (transfert des horizons C + (B) + A1), avec respectivement 28 % et 56 % d’espèces en moins. Cependant, 73 % des espèces identifiées sur le traitement C sont des espèces cibles.
En ce qui concerne la composition spécifique, le traitement C (transfert seul de l’horizon C) se distingue nettement des traitements BC (transfert des horizons C et (B)) et ABC (transfert des horizons C + (B) + A1) par la présence d’espèces pionnières telles que Petrorhagia prolifera (L.) P.W. Ball & Heywood (Caryophyllaceae) ou Silene nocturna L. (Caryophyllaceae).
Les analyses des teneurs en carbone organique total et en azote total indiquent que les traitements C et BC sont moins fertiles que le traitement ABCnc (transfert des horizons C + (B) + A1 sans compactage en surface) et que le sol de la steppe de référence. Bien que n’ayant pas permis le développement d’espèces nitratophiles, le caractère oligotrophe du traitement BC (transfert des horizons C et (B)) et le caractère encore plus oligotrophe du traitement C (transfert seul de l’horizon C) ont limité le développement d’autres espèces cibles. En effet, dans les sols fersialitiques, c’est surtout l’horizon A1 qui est riche en matières organiques, carbone, azote et en autres éléments minéraux comme le potassium, par exemple (Duchaufour Citation1984; AFES Citation2008).
L’augmentation très rapide du nombre d’espèces mesurée entre la première et la deuxième année sur le traitement BC (transfert des horizons C et (B)) correspondrait plutôt à un artéfact, puisque la végétation ne s’y était pas encore développée entre la fin de la mise en place des quadrats du traitement BC (transfert des horizons C et (B)) (avril 2011) et les relevés de la végétation (mi-mai 2011).
Sur les traitements C et BC (transfert des horizons C et (B)), la diversité spécifique est similaire à celle de la steppe de référence, mais l’indice de Bray-Curtis indique une dissimilarité significative avec celle-ci en ce qui concerne l’abondance des espèces végétales. Cependant, le traitement ABC (transfert des horizons C + (B) + A1) ne présente pas cette dissimilarité. Cela peut être expliqué par la dominance des graminées telles que Avena barbata Pott ex Link (Poaceae), Bromus sp. (Poaceae), Lolium rigidum Gaudin (Poaceae) ou encore Hordeum murinum L. (Poaceae) sur ces traitements, au même titre que la steppe de référence est dominée par Brachypodium retusum (Pers.) P. Beauv. (Poaceae).
Entre la deuxième et la troisième année de suivi, le pourcentage de recouvrement et la hauteur moyenne de la végétation augmentent significativement, mais le pourcentage de recouvrement reste encore très faible sur le traitement C (transfert seul de l’horizon C), certainement à cause du caractère très stressant de ce milieu. Sur le traitement BC (transfert des horizons C et (B)), au contraire, la végétation semble se stabiliser entre la deuxième et la troisième année, même en l’absence de pâturage. En effet, le pourcentage de recouvrement et la hauteur moyenne de la végétation cessent d’augmenter et deviennent semblables à ceux de la steppe de référence.
Le retour du pâturage ovin ancestral peut être envisagé sur le traitement BC (transfert des horizons C et (B)) pour conserver, à long terme, des espaces de sols nus favorables au développement des espèces cibles tolérantes aux stress (Gibson and Brown Citation1992; Bullock, Citation1998) et sur le traitement C (transfert seul de l’horizon C), pour maîtriser la hauteur de la végétation.
Effet du compactage de la surface du sol
Même si la végétation a eu une dynamique différente entre les trois années, au printemps 2013, aucune différence significative n’a été détectée entre les traitements ABC (transfert des horizons C + (B) + A1 avec compactage en surface) et ABCnc (transfert des horizons C + (B) + A1 sans compactage en surface).
Le compactage du sol ne semble donc pas augmenter ou accélérer le succès du transfert dans cet écosystème, bien qu’il paraisse ralentir la fermeture et la colonisation du milieu. En effet, un an après le transfert de sol, le pourcentage de recouvrement est de 20 % contre 40 % sans compactage du sol. II faut noter, cependant, que le retour du pâturage ovin pourrait avoir le même rôle (Gibson and Brown Citation1992; Bullock Citation1998; Stroh et al. Citation2002; Henry Citation2009).
Conclusion
A la suite du transfert de sol, nous avons pu mesurer une très forte résilience de la végétation steppique en matière de composition et de richesse spécifique. Ces premiers résultats sont en accord avec ceux d’études précédentes menées dans des prairies tempérées (Bullock Citation1998; Good et al. Citation1999; Box Citation2003; Trueman, Mitchell, and Besenyei, Citation2007; Eichberg et al. Citation2010) ou dans la plaine de La Crau (Jaunatre, Buisson, and Dutoit Citation2014). Les meilleurs résultats ont été obtenus avec le transfert de sol dans lequel les trois principaux horizons pédologiques ont été reconstitués, même en matière d’abondance des espèces végétales. Cependant, éventuellement même lorsque l’horizon organique A1 n’est pas reconstitué, les résultats sont également très prometteurs. Un transfert de sol par l’épandage d’un mélange organo-minéral, sans compactage final, pourrait être alors suffisant pour obtenir des résultats aussi rapidement. Cela pourrait permettre d’augmenter la surface sur lequel peut être épandue le sol transféré avec un étalement selon des rapports 1/2 ou 1/3. De plus, le transfert d’un volume de sol selon un rapport 1/1 n’entraîne pas un gain du point de vue de la restauration. Pour cela, le transfert de plaques, qui permet une recolonisation dans le site receveur et une cicatrisation dans le site donneur, constitue aussi une piste technique à explorer dans un cadre de recherche de durabilité des pratiques.
Le compactage du sol ne semble, donc, pas nécessaire. De plus, cela induit des émissions de CO2 supplémentaires et un coût de l’opération plus élevé.
Au final, aucun traitement n’a cependant permis de restaurer l’intégrité de la communauté végétale steppique de référence, notamment en matière d’abondance/dominance des espèces végétales mais également en matière de densité de la végétation. Aujourd’hui, il semble nécessaire de remettre en place le système de pâturage ovin ancestral (Gibson and Brown Citation1992; Bullock Citation1998; Stroh et al. Citation2002) dont les actions de prélèvement, piétinement et dispersion (endozoochorie, exozoochorie) devraient permettre d’accélérer la restauration de la structuration spatiale de la végétation steppique.
Bien qu’obtenant des résultats très prometteurs pour restaurer des communautés végétales à forte richesse spécifique (Pywell, Webb, and Putwain Citation1995; Bullock Citation1998; Good et al. Citation1999; Box Citation2003; Vécrin and Muller Citation2003; Trueman, Mitchell, and Besenyei Citation2007; Eichberg et al. Citation2010; Jaunatre, Buisson, and Dutoit Citation2014), le transfert de sol est une méthode de restauration écologique qualifiée de non-durable car destructrice d’une ressource non renouvelable, le sol. De plus, le transfert de sol véhiculé est une méthode polluante (émissions de CO2, NOx, huiles, etc.) et utilisant des énergies non-renouvelables. Elle ne doit donc en aucun cas représenter un substitut à une conservation in situ (McLean Citation2003).
Remerciements
Nous remercions la Société du Pipeline Sud-Est Européen, le Conseil Général des Bouches-du-Rhône, la région Provence-Alpes-Côtes-d’Azur (PACA), la FR ECCOREV et la SFR Tersys pour le financement de nos travaux. Nous remercions également le Conservatoire des Espaces Naturels PACA, la Chambre d’Agriculture PACA et la Réserve Naturelle Nationale des Coussouls de Crau pour nous avoir donné la permission de réaliser nos travaux et d’accéder au site d’étude.
Note biographique courte des auteurs:
Adeline Bulot est doctorante; elle a mis en place les protocoles expérimentaux, réalisé les suivis, interprété les données et rédigé l’article.
Erick Provost est chargé de recherches au CNRS; il a contribué à la mise en place de cette problématique de recherche.
Thierry Dutoit est directeur de recherches au CNRS; il est à l’origine de cette expérimentation dont il a mis en place les différents protocoles. II a réalisé les suivis et validé les analyses, leurs résultats et leurs interprétations.
References
- Adriaens, D., O. Honnay, and M. Hermy. 2006. “No evidence of a plant extinction debt in highly fragmented calcareous grasslands in Belgium.” Biological Conservation 133 (2): 212–224.
- AFES. Association Française pour l’Étude des Sols. 2008. Référentiel pédologique. Versailles: Éditions Quae.
- Alard, D., and I. Poudevigne. 2002. “Biodiversity in changing landscapes: from species to patch assemblages to system organization.” In: Application of G.I.S and remote sensing in river studies, edited by R.S.W. Leuwen, I. Poudevigne, and R. Teeuw eds, 9–27. Leiden: Backhuys.
- Baize, D. 2000. Guide des analyses pédologiques. Paris : 2ème édition INRA.
- Badan, O., G. Congés, and J.P. Brun. 1995. “Les bergeries romaines de La Crau d’Arles. Les origines de la transhumance en Provence.” Galia 52: 263–310.
- Benjamini, Y., and Y. Hochberg. 1995. “Controlling the false discovery rate: a practical and powerful approach to multiple testing.” Journal of the Royal Statistical Society, Serie B 57 (1): 289–300.
- Borcard, D., F. Gillet, and P. Legendre. 2011. Numerical ecology with R. New York: Springer.
- Bourrely, M., L. Borel, J.P. Devaux, J. Louis-Palluel, and A. Archiloque. 1983. “Dynamique annuelle et production primaire nette de l’écosystème steppique de Crau (Bouches du Rhône).” Biologie-Ecologie méditerranéenne 10: 55–82.
- Box, J. 2003. “Critical factors and evaluation criteria for habitat translocation.” Journal of Environmental Planning and Management 46(6): 839–856.
- Bray, J.R., and J.T. Curtis. 1957. “An ordination of upland forest communities of southern Wisconsin.” Ecological Monographs 27: 325–349.
- Braun-Blanquet J., N. Roussine, R. Nègre, and L. Emberger. 1952. Les groupements végétaux de la France Méditerranéenne. Paris: édition CNRS.
- Bruelheide, H., and T. Flintrop. 2000. “Evaluating the transplantation of a meadow in the Harz Mountains, Germany.” Biological conservation 92(1): 109–120.
- Buisson, E., T. Dutoit, F. Torre, and P. Poschold. 2006. “The implications of seed rain and seed bank patterns for plant succession at the edges of abandoned fields in Mediterranean landscapes.” Agriculture, Ecosystems and Environment 11: 6–14.
- Bullock, J.M. 1998. “Community translocation in Britain: Setting objectives and measuring consequences.” Biological conservation 84(3): 199–214.
- Cherel, O. 1988. “Contribution à l’étude des relations végétation-mouton sur les parcours de Crau (S.E France): adaptation et développement de méthodes d’étude du régime alimentaire.” PhD diss.: Aix-Marseille Université, Marseille.
- Chessel, D., A.B. Dufour, and J. Thioulouse. 2004. “The ade4 package-I- one-table methods.” R News 4: 5–10.
- Clewell, A.F., and J. Aronson. 2007. Ecological restoration: principles, values and structure of an emerging profession. 1st ed. Washington, Covelo, London: Island Press.
- Coiffait-Gombault, C. 2011. “Règles d’assemblage et restauration écologique des communautés végétales herbacées méditerranéennes. Le cas de la Plaine de Crau (Bouches-du-Rhône, France).” PhD diss., Université d’Avignon et des Pays du Vaucluse, Avignon.
- Colomb, E., and R.M. Roux. 1978. “La Crau, données nouvelles et interprétations.” Géologie méditerranéenne 3: 303–324.
- Devaux, J.P., A. Archiloque, L. Borel, M. Bourrely, and J. Louis-Palluel. 1983. “Notice de la carte phyto-écologique de la Crau (Bouches-du-Rhône).” Biologie et écologie méditerranéenne 10(1–2): 5–54.
- Deverre, C. 1996. “La nature mise au propre dans la steppe de Crau et la forêt du Var.” Etudes rurales 141: 45–61.
- Dray, S., and A.B. Dufour. 2007. “The ade4 package: Implementing the duality diagram for ecologists.” Journal of statistical software 22: 1–20.
- Duchaufour, P. 1984. Pédologie. Paris: Masson.
- Dutoit, T., E. Buisson, F. Fadda, F. Henry, C. Coiffait-Gombault, and R. Jaunatre. 2011. “Dix années de recherche dans une pseudo-steppe méditerranéenne: impact des changements d’usage et restauration écologique.” Sécheresse, 22: 75–85.
- Dutoit, T. 2011. “Fuite d’hydrocarbures en Crau: Mesures compensatoires, dernier acte d’une catastrophe patrimoniale?” Le courrier de la Nature 263: 7–9.
- Dutoit, T., E. Buisson, S. Fadda, F. Henry, C. Coiffait-Gombault, R. Jaunatre, J.F. Alignan, S. Masson, and A. Bulot. 2013. “The pseudo-steppe of La Crau (South-Eastern France): origin, management and restoration of a Mediterranean rangeland.” In Steppe Ecosystems: Biological Diversity, Management and Restoration, edited by J. Traba, J. Morales and M. Morales, 287–301. USA: Nova Publishers.
- Dutoit, T., R. Jaunatre, and E. Buisson. 2013. “Mediterranean steppe restoration in France.” In: Ecological Restoration Principles, Values, and Structure of an Emerging Profession, edited by A.F. Clewell, and J. Aronson, second edition, 60–64. Washington: Island Press.
- Dzwonko, Z., and S. Loster. 1998. “Dynamics of species richness and composition in limestone grassland restored after tree cutting.” Journal of Vegetation Sciences 9(3): 387–394.
- Eichberg, C., C. Storm, M. Stroh, and A. Schwabe. 2010. “Is the combination of topsoil replacement and inoculation with plant material an effective tool for the restoration of threarened sandy grassland?” Applied Vegetation Science 13: 425–438.
- Etienne, M., J. Aronson, and E. Le Floc’h. 1998. “Abandoned lands and land use conflicts in southern France. Piloting ecosystem trajectories and redesigning outmoded landscapes in the 21st century.” In: Landscape degradation and biodiversity in Mediterranean-type ecosystems, edited by P.W. Rundel, G. Montenegro, and F. Jaksic. 127–140. Ecological studies n° 136, Berlin: Springer.
- Gibson, D.J. 2009. Grasses and grassland ecology. Oxford: Oxford University Press.
- Gibson, C.W.D., and V.K. Brown. 1992. “Grazing and vegetation change: deflected or modified succession?” Journal of Applied Ecology 29: 120–131.
- Good, J.E.G., H.L. Wallace, P.A. Stevens, and G.L. Radford. 1999. “Translocation of herb-rich grassland from a site in wales prior to opencast coal extraction.” Restoration Ecology 7: 336–347.
- Henry, F. 2009. “Origine et dynamique à long terme d’un écosystème herbacé pseudo steppique, le cas de la plaine de la Crau (Bouches-du-Rhônes, France).” PhD diss.: Université Paul Cézanne, Marseille.
- Hirst, A.H., R.F. Pywell, R.H. Marrs, and P.D. Putwain. 2003. “The resistance of a chalk grassland to disturbance.” Journal of Applied Ecology 40: 368–379.
- Hirst, A.H., R.F. Pywell, R.H. Marrs, and P.D. Putwain. 2005. “The resilience of calcareous and mesotrophic grasslands following disturbance.” Journal of Applied Ecology 42 (3): 498–506.
- Holling, C.S. 1973. “Resilience and stability of ecological systems.” Annual review of Ecology and Systematics 4: 1–23.
- Holling, C.S., and L.H. Gunderson. 2002. “Resilience and adaptative cycles.” In: Understanding transformations in human and natural systems, edited by L.H. Gunderson, and C.S. Holling, 25–62. Panarchy, Washington: Island Press.
- Jaunatre, R., E. Buisson, and T. Dutoit. 2012. “First year results of a multi-treatment steppe restoration experiment in La Crau (Provence, France).” Plant Ecology and Evolution 145: 13–23.
- Jaunatre, R., E. Buisson, and T. Dutoit. 2014. “Can ecological engineering restore Mediterranean rangeland after intensive cultivation? A large-scale experiment in southern France.” Ecological Engineering 64: 202–212.
- Kiehl, K., A. Kirmer, T.W. Donath, L. Rasran, and N. Hölzel. 2010. “Species introduction in restoration projects – Evaluation of different techniques for the establishment of semi-natural grasslands in Central and Northwestern Europe.” Basic and Applied Ecology 11: 285–299.
- Lee, T.D., M.G. Tjoelker, D.S. Ellsworth, and P.B. Reich. 2001. “Leaf gas exchange responses of 13 prairie grassland species to elevated CO2 and increased nitrogen supply.” New phytologist 150(2): 405–418.
- Luh, H.K., and S.L. Pimm. 1993. “The assembly of ecological communities: a minimalist approach.” Journal of Animal Ecology 62: 749–765.
- Masson, P., and E. Estrangin. 1928. “Le mouvement économique: l’agriculture. Le bilan du XIXe siècle.” Encyclopédie départementale des Bouches-du-Rhône 10: 353–379.
- McLean, I.F.G., 2003. A habitats translocation policy of Britain. Joint Nature Conservation Committee on behalf of The Countryside Council for Wales, English Nature and Scottish Natural Heritage.
- Molinier, R., and G. Tallon. 1950. “La végétation de la Crau (Basse-Provence).” Revue générale de botanique 56: 1–111.
- Oksanen, J., F.G. Blanchet, R. Kindt, P. Legendre, P.R. Minchin, R.B. O’Hara, G.L. Simpson, P. Solymos, M.H.H. Stevens, and H. Wagne. 2012. “Vegan: community ecology package.” R package version 2.0-5. Accessed March 12 2014. http://CRAN.R-project.org/package=vegan.
- Pielou, E.C. 1969. An introduction to mathematical ecology. New York: Wiley-Interscience.
- Pimm, S.L. 1991. The balance of nature? Ecological issues in the conservation of species and communities. Chicago: University of Chicago Press.
- Poschold, P., and M.F. WallisdeVries. 2002. “The historical and socioeconomic perspective of calcareous grasslands: lessons from the distant and recent past.” Biological Conservation 104: 361–376.
- Pywell, R.F., N.R. Webb, and P.D. Putwain. 1995. “A comparison of techniques for restoring heathland on abandoned farmland.” Journal of Applied Ecology 32: 400–411.
- R foundation for Statistical computing. 2014. “The R project for Statistical Computing.” Accessed March 12. http://www.r-project.org/index.html.
- Rainaud, A. 1893. “La Crau.” Annales de géographie 2: 189–211.
- Römermann, C., O. Tackenberg, and P. Poschold. 2005. “How to predict attachment potential of seeds to sheep and cattle coat from simple morphological seed traits.” Oikos 110: 219–230.
- Saunders, D.A., R.J. Hobbs, and C.R. Margules. 1991. “Biological consequences of ecosystem fragmentation: a review.” Conservation Biology 5: 18–32.
- SER International Primer on Ecological Restoration. 2004. “Society for Ecological Restoration International Sciences and Policy Working Group.” Accessed March 12 2014. https://www.ser.org/resources/resources-detail-view/ser-international-primer-on-ecological-restoration.
- Stroh, M., C. Storm, A. Zehm, and A. Schwabe. 2002. “Restorative grazing as a tool for directed succession with diaspore inoculation: the model of sand ecosystem.” Phytocoenologia 32: 595–625.
- Tatin, L., A. Wolff, J. Boutin, E. Colliot, and T. Dutoit. 2013. Ecologie et conservation d’une steppe méditerranéenne: la plaine de Crau. Versailles: Éditions Quae.
- Trueman, I., D. Mitchell, and L. Besenyei. 2007. “The effects of turf translocation and other environmental variables on the vegetation of a large species-rich mesotrophic grassland.” Ecological Engineering 31: 79–91.
- Vécrin, M.P., and S. Muller. 2003. “Top-soil translocation as a technique in the re-creation of species-rich meadows.” Applied Vegetation Science 6: 271–278.
- Weiher, E., and P.A. Keddy. 1999. Ecological assembly rules: perspectives, advances, retreats. Cambridge: Cambridge University Press.
- Wells, T.C.E. 1969. “Botanical aspects of conservation management of chalk grasslands.” Biological Conservation 2 (1): 36–44.
- Willems, J.H. 2001. “Problems, approaches and results in restoration of Dutch calcareous grassland during the last 30 years.” Restoration ecology 9(2): 147–154.
- Wolff, A. 2014. “Risques industriels dans les espaces naturels: retour d’expérience après la rupture d’un pipeline.” In: Écologie et conservation d’une steppe méditerranéenne, La plaine de Crau, edited by L. Tatin, A. Wolff, J. Boutin, E. Colliot and T. Dutoit, 309–323. Versailles: Éditions Quae.
- Young, T.P., D.A. Petersen, and J.J. Clary. 2005. “The ecology of restoration: historical links, emerging issues and unexplored realms.” Ecology Letters 8: 662–673.