
Abstract
Throughout the world, dunes are being altered by human activities. Dunes can be restored to naturally functioning systems to protect human settlements and coastal strips and preserve biotopes. This study took place on the Jaï dunes, on the shore of a coastal lagoon in southeastern France, which were mainly degraded by vehicles. The objectives of this work were (1) to define the reference ecosystem by gathering historical information and exploring other regional sand dunes; (2) to find out about the restoration potential of the site by studying the degraded seed bank; (3) to test the efficacy of a few basic restoration actions. Historical information was of little help in defining the reference ecosystem. The description of EU Habitat Directive habitat types and the selection of regionally less degraded sites allowed us to compile the following list of plant species that we could expect to find in the restored area: Anthemis maritima, Elytrigia juncea, Glaucium flavum and Sporobolus pungens. The seed bank of the road was mainly composed of Juncus acutus, which is not a typical dune species and therefore did not have much potential for restoration. Ploughing can therefore be used to decompact the sand road without hampering colonization from the seed bank. Ploughing was only efficient when the following other conditions were met: environmental conditions adequate for the establishment of dune vegetation, and a sufficient connection between the beach, the foredune and the restored area.
Introduction
Throughout the world, shorelines are being altered by human activities. Dunes are eliminated in support of construction projects such as buildings, roads, or other infrastructure; beaches and dunes are often subject to grading; and recreational or agricultural activities can involve mechanical cleaning and sand extraction (Nordstrom Citation2008; Lithgow et al. Citation2013). However, healthy dune and beach systems can provide useful goods and services: they can mitigate the impact on human-made structures arising from floods and severe storms, and they can provide nesting sites and refuge areas for wildlife (MEA Citation2005; Nordstrom Citation2008). Ecological restoration, which is the process of assisting the recovery of degraded ecosystems (SER Citation2004), can be used to restore naturally functioning dunes and beaches, even in developed areas with aims as varied as: protecting human settlements, protecting the coastal strip, preserving natural coastal dynamics, providing sustainable and environmentally friendly tourism and development, and protecting biotopes, landscapes and seascapes (Nordstrom Citation2008).
Various actions already take place to restore coastal dunes, and they span contrasting activities that range from re-vegetating and stabilizing the mobile substrate to removing plant cover and increasing substrate mobility (Lithgow et al. Citation2013; Martínez et al. Citation2013). Restoration actions are based on (1) the specific properties of the designated reference ecosystem, (2) a good knowledge of healthy ecosystem functioning, and (3) the state of degradation, or potential for restoration, of the ecosystem to be restored.
For each restoration project, defining a reference ecosystem is essential to the goals of the project and can provide a basis for monitoring progress and measuring success (Holl and Cairns Citation2002; Bainbridge Citation2007). The pre-disturbance state can be a helpful guide to restoration if enough is known of the historical condition of the site in question and/or if large areas of the pre-disturbance state are still found in the landscape. However, when using the pre-disturbance state, one has to keep in mind that the restored ecosystems have to be dynamic and resilient to disturbances, because of continuously evolving constraints (e.g. technical, socio-economic) and conditions (e.g. climatic, physical, biological) (Choi Citation2007). The reference ecosystem should be based on the range of what is possible, as illustrated by spatial and temporal variation in natural ecosystems, and local, ecological, cultural, and historical information (Parker and Pickett Citation1997; Clewell and Aronson Citation2007).
To plan restoration appropriately, a good knowledge of the reference ecosystem functioning is needed. The ecological restoration of dunes and beaches must take into account their inherent characteristics driven by particular environmental factors such as wind, sand movement and burial, salt spray, which act on a gradient (Maun Citation2009). This leads to a division of these landscapes into a number of ecosystems: ephemeral beach communities on upper beaches, perennial herbaceous communities on embryo dunes, perennial herbaceous communities on mobile dunes, perennial or annual interdune communities on transition dunes, and fixed dune communities (Carboni, Santoro, and Acosta Citation2010).
Not only does the reference ecosystem need to be described and understood, but the disturbance(s) that were responsible for degrading the site, together with the restoration potential of the project site, have to be assessed before designing a restoration programme. Within the framework of plant community restoration, the community assembly filter model (Lortie et al. Citation2004) is widely used in summarizing the four main barriers to restoration: the global species pool available for assembling the community, species dispersal, environmental conditions, and biotic interactions. Some authors propose dividing the global species pool into two categories: the external species pool, which are species found in the landscape, and the internal species pool, which are species already present on site (i.e. in the seed bank or as living specimens that collectively survived the disturbance) (Fattorini and Halle Citation2004). The internal species pool has to be studied in detail: because it has not passed through the dispersal filter, it can provide information on potential seed sources at the site to be restored (Willems and Bik Citation1998).
In southeastern France, coastal sand dunes are mainly found in the Camargue area (Bensettiti et al. Citation2004) and are therefore quasi-restricted to the Rhône delta. Other dunes of the type found on lagoon and pond shores, such as Jaï dunes, are extremely rare. The Jaï dunes and the surrounding wetlands, located on the shores of the 15,500-ha brackish to marine Berre Lagoon and the brackish to freshwater Bolmon pond, were placed under the protection of the Conservatoire du Littoral between 1992 and 2008, and have been managed by the Syndicat Intercommunal du Bolmon et du Jaï (SIBOJAI) since 1992 (43°24’52.992” N, 5°10’0.0114” E). The shores of the Berre Lagoon have been strategic sites for major economic developments, especially since the 1920s: refineries, a hydroelectric plant, the steel industry, urban development, and an airport, to cite only a few examples. The lagoon became dramatically polluted by the end of the 1950s; this was followed by an input of 3.3 billion m3/year of fresh water (about 3.7 times the volume of the lagoon/year), and of 520,000 t/year of sediments between 1966 and 1993 (GIPREB Citation2007). This drastically reduced the productivity of sea-grass beds and molluscs in the lagoon, the latter being the main source of material for the Jaï dunes, 80% of which are composed of shellfish debris. Since 2002, the flora and fauna have begun to resettle, partly as a result of a programme of freshwater input curtailment intended to protect the lagoon (EU habitat-type 1150). Within this improved context, the restoration of the Jaï dunes is planned.
The objectives of this work were (1) to define the reference ecosystem for the restoration, a narrow portion of these dunes, by gathering historical information and exploring other regional sand dunes; (2) to find out about the restoration potential of the site by studying the seed bank of the degraded system; (3) to test the efficacy of a few basic restoration actions to restore the dune plant community designated in step 1.
Material and methods
The site to be restored
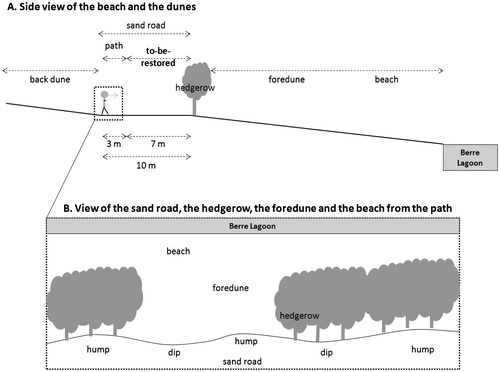
Some of the Jaï dunes were completely destroyed by urban development and these areas are not part of the restoration project. The remaining dunes, which are part of the project, stretch over 6.5 km, are 150–250 m wide, and have a maximum height of 2 m. They were mainly degraded by vehicles driving on the foredune and the beach. A hedgerow of Tamarix gallica was planted in the 1990s to prevent vehicles from parking on the beach and to separate the beach and foredune from the road and back dune. From the Berre Lagoon, located north of the Jaï, are found various ecosystems: first, the beach, then, the foredune, followed by the Tamarix gallica hedgerow and the 10-m wide sand road (7 m of which are to be restored and 3 m of which are to remain a path for visitors) which were both part of the foredune, and the back dune (Figure A). Because the sand road had been used for years, the road does not have much remaining vegetation, and the soil is highly compacted and has many small humps and dips. The hedgerow is not continuous (Figure B).
Figure 1. View of the Jaï dunes and of the site to be restored. (A) Side view of the beach and dunes. (B) View from the path looking toward Berre Lagoon.
Reference ecosystem
In order to describe the vegetation of regional foredunes, historical information on the Jaï were gathered from an updated catalogue (Crouzet, Pavon and Michaud Citation2009) in which all botanical data for the Bouches-du-Rhône département are compiled (Molinier Citation1981), and reference ecosystems in the region were searched for and characterized by their vegetation. All the sites with a remaining dune ecosystem around the Berre Lagoon were sampled (Berre l’étang and Figuerolles beach). To increase the number of sites, two more sites, also found on a lagoon shore located about 100 km away, were selected (Aresquiers beach and Airolles beach), resulting in four reference sites with similar characteristics (Table ).
Table 1. List of the sites that were evaluated as candidate reference ecosystems.
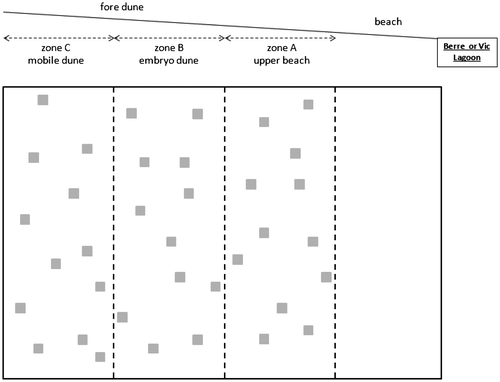
At each site (the four reference sites and the Jaï site), three zones parallel to the shore were defined to encompass the vegetation gradient of the foredune and were designated as follows: upper beach, embryo dune and mobile dune (Figure ). The width of each zone, visually determined based on the percentage of vegetation cover, differed from one site to the other, as sites varied considerably in size (Figure ). In May 2013, the percentage of cover of all species was visually estimated in 12 randomly set 50 × 50-cm quadrats at each site. The nomenclature used for plants in the study is that of TAXREF (Muséum national d’Histoire naturelle 2003–13).
Figure 2. Sampling design on the four candidate reference ecosystem sites (see Table ) and on the Jaï site.
Restoration potential: on-site seed bank
The persistent seed bank, consisting of seeds that persist in the seed bank for > 1 year (Thompson, Bakker, and Bekker Citation1997), was sampled on the Jaï foredunes in February 2013 before the input of fresh seeds and after the germination of most seeds in the autumn and winter. At each sampling point, three depths were sampled: between 0 and 5 cm, between 5 and 15 cm, and deeper than 15 cm. At each depth, three sub-samples were collected and mixed together to obtain a 1-litre sample. Four replicates were taken in each area sampled: the actual foredune, within the hedgerow, the sand road to be restored: on humps and in dips in the sand road, which could either be adjacent to unbroken sections of the hedgerow or to gaps in the hedgerow (Figure ). The 72 samples were set to germinate using the standard procedure recommended by Ter Heerdt et al. (Citation1996). Soil samples were concentrated by rinsing them with water and straining them through 2-mm and 200-μm sieves in order to reduce bulk and eliminate clay. The concentrated samples were then thinly spread in plastic vermiculite-filled trays topped with medical compresses (mesh size 100 μm) to prevent seed from migrating through the substrate. All trays were watered frequently from below. Emerged seedlings were identified (Mamarot, Psarski, and Rouquier Citation1997), counted, and removed weekly. Unidentified seedlings were allowed to grow so that their identities could be established later. Three trays with no sample were also set in the greenhouse to make sure that no seed could come with the seed rain.
Restoration techniques and monitoring
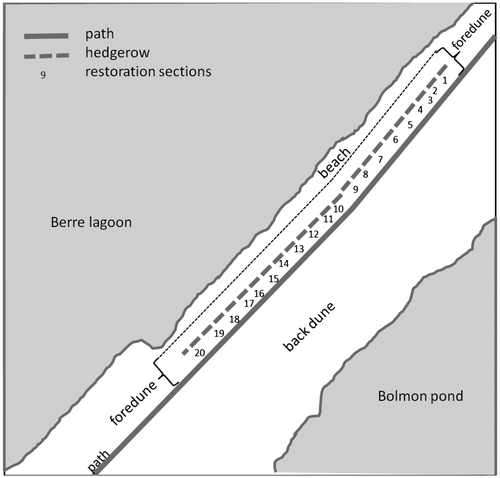
Restoration was applied on part of the 10-m wide sand road: 3 m of this road remain a path for visitors and 7 m were to be restored (Figure A). The 1000-m long sand road to be restored was divided into 20 sections (Figure ), of approximately 50 m average length, and allocated to the following treatments: control, ploughed, and ploughed with soil transfer (Table ). In order to decompact the soil, the sand road was ploughed to a depth of 15 or 40 cm using a donkey (15 cm) or small tractor (40 cm) and chisel plough. This method was chosen because the site is small and to minimize the environmental and actual costs. Soil from the beach that was likely to contain seeds was then hand-transferred by local volunteers. Some sections of the sand road were left without any treatment to serve as controls.
Figure 3. Aerial view of the Jaï site: the 1000-m long sand road was divided into 20 restoration sections of approximately 50-m average length where restoration treatments were applied (see Table
Table 2. Restoration treatments applied to the sand road.
In May 2013, the per cent cover of all species was visually estimated in four randomly set 50 × 50-cm quadrats in each restoration section (Figure ).
Statistical analyses
As data did not meet the conditions for parametric analyses (data were not normally distributed or variances were not homogeneous), four Kruskal–Wallis tests were performed, followed by Mann–Whitney post-hoc tests with Bonferroni p-adjustment to compare (1) vegetation species richness between sites and zones (in the reference ecosystem; Figure ), (2) seed-bank species richness and number of seeds in the seed bank between areas (Figure ) and depths, and (3) vegetation species richness between treatments (Figure and Table ).
Three correspondence analyses were performed in order: (1) to obtain an overview of the vegetation composition on the foredunes between the site to be restored (Jaï) and the four potential reference sites, (five sites, three zones, Table , Figure ) (175 quadrats × 44 species), (2) to describe the seed bank composition at the site to be restored (Jaï) at three depths, on the foredune, the hedgerow and the sand road to be restored (sand road humps and dips, either adjacent to unbroken hedgerow or to hedgerow gaps) (72 quadrats × 24 species), and (3) to describe the vegetation composition that results from the different restoration treatments (Table , sand road humps and dips, either adjacent to unbroken hedgerow or to hedgerow gaps) (136 quadrats × 39 species).
All analyses were performed with R software (R Development Core Team, Citation2008) using the following R packages: “ade-4” and “stat”.
Results
Reference ecosystem
To help in defining the reference ecosystem for the restoration, the historical information on the site was found in Molinier’s catalogue (Molinier Citation1981) of Bouches-du-Rhône Vascular Plants, which documents all of the species found in the département, as observed by different authors at different times. In total, 4133 species were identified throughout the département, of which 190 were recorded on the periphery the Berre Lagoon. Among these, 27 were reported on the Jaï dunes, without making a distinction between foredune and back dune species: Allium chamaemoly, Anthemis maritima, Asparagus officinalis, Avellinia michelii, Blackstonia imperfoliata, Centaurea diffusa, Cerastium siculum, Cochlearia glastifolia, Dichanthium ischaemum, Elytrigia juncea, Ephedra distachya, Euphorbia esula, Euphorbia paralias, Hypochaeris radicata, Lotus corniculatus subsp. preslii, Medicago marina, Melilotus indicus, Myosotis discolor, Plantago crassifolia, Polygonum maritimum, Polygonum robertii, Rubia tinctorium, Senecio crassifolius, Sporobolus pungens, Tribulus terrestris, Trifolium spumosum and Vicia lutea. Three more species were found in 2009: Cynaria cardunculus, Hypochaeris achyrophorus and Ophrys ciliata (Crouzet, Pavon, and Michaud Citation2009).
To further define the reference ecosystem for the restoration, other regional sand dunes were explored and 44 species were identified in total over the five sites with an average of 3.1 species per 50 × 50-cm quadrat. Mean species richness was found to be lower on Aresquiers beach at Vic Lagoon, and on the upper beach (zone A, Figure ) of Figuerolle and Airolles (χ² = 88.0, p < 0.001).
The correspondence analyses performed on these regional data aimed to characterize the vegetation of the foredunes of the site to be restored (Jaï) and the four potential reference sites in order to find out: (1) if the Jaï foredunes are similar to other regional healthier foredunes and can therefore be their own reference, (2) if not, which other site can serve as a reference for the restoration of the narrow portion which has been compacted, and (3) if there is a zonation of the foredune vegetation at all sites (upper beach, embryo dune and mobile dune). Together with the two Vic Lagoon sites, Figuerolle beach at Berre lagoon was considered and found to be separated from Jaï and Berre l’étang by Axis 1 (6.6%) of the correspondence analysis (175 quadrats × 44 species). The first group of sites was found to be characterized by Salsola kali (contribution to Axis 1: 14.9), Glaucium flavum (10.5) and Beta vulgaris (5.75), with Jaï and Berre l’étang being better characterized by Anthemis maritima (18.9), Elytrigia juncea (16.5), Sporobolus pungens (3.0) and Valentia muralis (1.9) (Figure ). Axis 2 (6.3%) of the correspondence analysis separated the zones on the two Vic Lagoon sites. Zone A, which is closer to the lagoon (Figure ), was found to be characterized by Salsola kali (contribution to Axis 2: 44.9) and Beta vulgaris (9.2), and Zone C by Glaucium flavum (9.0), Geranium molle (8.4), Cerastium pumilum (8.1), Vulpia fasiculata (4.6), Centranthus calcitrapa (2.2) and Arenaria leptoclados (1.3) (Figure ). The portion of the sand road to be restored, being away from the lagoon and closer to the back dune, would most probably correspond to a plant community where the latter set of species would be found.
Figure 4. Correspondence analysis performed on species per cent cover for the five sites [the four (see Table ) candidate reference ecosystems + the Jaï site] on the three zones (A, B and C; Figure ) (175 quadrats × 44 species). Only the most correlated species are shown. Samples are grouped according to site (name of the lagoon on which the site is found – Vic or Berre, name of the site). Zones are indicated only for the two sites on the Vic Lagoon (Airolles and Aresquiers) because zones overlapped on the other sites.
![Figure 4. Correspondence analysis performed on species per cent cover for the five sites [the four (see Table 1) candidate reference ecosystems + the Jaï site] on the three zones (A, B and C; Figure 2) (175 quadrats × 44 species). Only the most correlated species are shown. Samples are grouped according to site (name of the lagoon on which the site is found – Vic or Berre, name of the site). Zones are indicated only for the two sites on the Vic Lagoon (Airolles and Aresquiers) because zones overlapped on the other sites.](/cms/asset/2b736d0d-b687-436d-a949-58d0d7c9010d/tabg_a_919875_f0004_b.gif)
Restoration potential: on-site seed bank
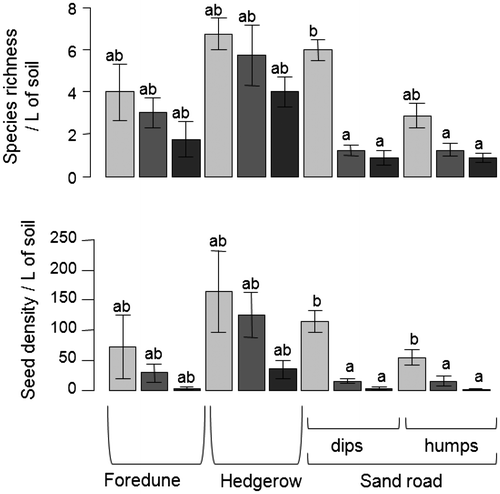
To find out about the restoration potential of the site to be restored, the seed bank of the degraded system was studied, along with the seed bank of the foredune, which played the role of donor site for some of the restoration treatments (soil transfers, Table ). Twenty-four species were identified in total in the seed bank with an average of 2.9 species and 46.6 seeds per sample. Species richness was found to be slightly higher in the hedgerow, on the foredune, and in the sand road surface soil, than in the sand road’s deeper layers (χ² = 47.7, p < 0.001; Figure ). Seed density was also observed to be higher in surface soil than deeper down (χ² = 47.7, p < 0.001; Figure ).
Figure 5. Mean number of species and of seeds in the seed bank on the Jaï site on the foredune, on the hedgerow and on the sand road: depth between 0 and 5 cm, light grey bars; between 5 and 15 cm, dark grey bars; deeper than 15 cm, black bars. Kruskal–Wallis test for species richness: χ² = 47.7, p < 0.001; Kruskal–Wallis test for seed density: χ² = 47.7, p < 0.001; the boldface letters indicate significant differences between areas, results of the Mann–Whitney post-hoc tests with Bonferroni p-adjustment.
The three main areas that were sampled, namely, the sand road, the hedgerow and the foredune, appeared to be well separated by Axis 1 (39.9%) of the correspondence analysis on the number of seeds in the seed bank (72 quadrats × 24 species). These three areas were respectively characterized by Juncus acutus (21.0), Anthemis maritima (contribution to axis 1: 11.7) and by Plantago coronopus (64.5). Axis 2 (16.2%) appeared to separate various areas of the foredune, which are characterized either by Anthemis maritima (contribution to axis 2: 76.2) or by Plantago coronopus (19.7). There was no clear difference in seed bank composition between depths or between humps and dips on the sand road.
Restoration techniques and monitoring
At the Jaï site, an average of 4.1 species per 50 × 50 cm quadrat were identified on the foredune, in the hedgerow, and on the sand road, with 39 distinct species found overall. On the sand road, a lower species richness was found on the humps than in the dips (χ² = 47.0, p < 0.001).
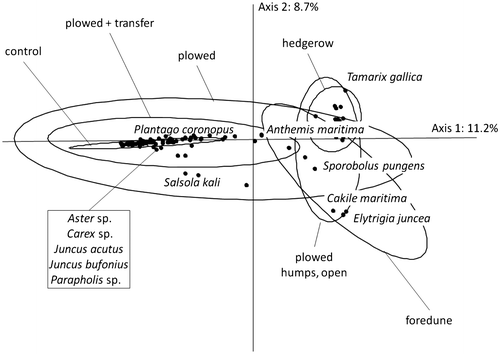
In the correspondence analysis on the species per cent cover (136 quadrats × 39 species) performed to identify the best treatment for the area to be restored (i.e. the treatment leading to a plant composition closest to that of the Jaï foredune, which was the donor site for the soil transfers), it was found that the control group, together with most of the restoration-treated areas, are separated from the foredune by Axis 1 (11.2%; Figure ). The latter treated areas are characterized by Juncus acutus (contribution to Axis 1: 13.1), Juncus bufonius (10.4) and Parapholis sp. (25.5). The hedgerow and the “ploughed” portions of the sand road humps adjacent to hedgerow gaps appear to be characterized by Elytrigia juncea (11.5), Anthemis maritima (10.5) and Sporobolus pungens (4.4). Axis 2 (8.7%) was found to separate the hedgerow characterized by Tamarix gallica (contribution to axis 2: 21.6) from the foredune, which is characterized by Elytrigia juncea (55.9) and Cakile maritima (5.6).
Figure 6. Correspondence analysis performed on species per cent cover for restoration-treated areas (see Table , sand road humps and dips, either adjacent to unbroken hedgerow or to hedgerow gaps) (136 quadrats × 39 species). Only the most correlated species are shown. Samples are grouped according to restoration treatment.
Discussion
Reference ecosystem
Because the Jaï dune species list in Molinier’s Catalogue (Molinier Citation1981) of Bouches-du-Rhône Vascular Plants does not make any distinction between foredune and back dune species, it is of very limited use in elaborating the future species list at the restored site, which does not include any back dunes. Among the 12 species found in the Jaï site in our 2013 plant survey, three were mentioned by Molinier’s catalogue (Citation1981): Anthemis maritima (present in 29 quadrats out of 35), Sporobolus pungens (28 quadrats) and Elytrigia juncea (26 quadrats). The other nine species may have had too much of a wide distribution range in the département to be mentioned in the catalogue or were not present at the time: Atriplex tornabenei, Cakile maritima, Cerastium semidecandrum, Geranium molle, Inula crithmoides, Medicago littoralis, Plantago coronopus, Salsola kali and Tamarix gallica.
The four sites selected as potential reference sites, so selected because they were located on lagoon shores, shared some species with the catalogue and the Jaï site, in particular, Anthemis maritima (present in 41 quadrats out of 171). The Berre l’étang site, on Berre Lagoon, had a vegetation composition relatively close to that of the Jaï site (e.g. Anthemis maritima present in 35 and 29 quadrats out of 36, respectively). This is most probably because these two sites are highly frequented by human visitors; Berre l’étang site was also probably degraded by the nearby installation of salt ponds and of a refinery. On the three other sites (Figuerolle on Berre Lagoon, Airolles and Aresquiers on Vic Lagoon), species richness was low (2.8 species per quadrat, with a total of 29 species in Figuerolle, 16 species in Airolles and 12 species in Aresquiers on Vic Lagoon), but comparable to other coastal foredunes of the world (e.g. van der Valk Citation1975; Costa Citation1997; Isermann Citation2005). Usually, the Mediterranean foredunes are composed of an ephemeral beach community on the upper beach (Habitat Directive 92/43/EEC: 1210 annual vegetation of drift lines) with Cakile maritima and Salsola kali, a perennial herbaceous community on embryo dunes (Habitat Directive 92/43/EEC: 2110 embryonic shifting dunes) with Elymus sp. and Sporobolus sp., and a perennial herbaceous community on mobile sand (Habitat Directive 92/43/EEC: 2120 shifting dunes along the shoreline) with Ammophila arenaria, Anthemis maritima and Pancratium maritimum (Acosta et al. Citation2007; Carboni, Santoro, and Acosta Citation2010). Both Vic Lagoon sites exhibited this structure: an ephemeral beach community with Salsola kali and Beta vulgaris, a perennial herbaceous community on embryo dunes with Sporobolus pungens and Elytrigia juncea and a perennial herbaceous community on mobile sand with Anthemis maritima and Glaucium flavum. They can therefore be considered as good local references for foredune restoration in the region. While the Jaï and Figuerolle sites did not have such a clear structure, they still had many of the common species expected on foredunes and can also be considered for the restoration of this narrow stretch of dune on the Jaï. The portion of the sand road to be restored, being away from the lagoon and closer to the back dune, would most probably correspond to embryo and mobile dune plant communities rather than that of the upper beach and would be characterized by Anthemis maritima, Elytrigia juncea, Glaucium flavum and Sporobolus pungens.
Restoration potential
The seed bank on the road to be restored has a lower seed density than the foredune and is species-poor, being mainly composed of Juncus acutus, which is not a typical dune species (target species). Restoration potential from the topsoil seed bank is therefore low, as observed by Mason, French and, Russell (Citation2007) on coastal dunes. Many dune annuals have no buried seed bank (García Novo et al. Citation2004). Moreover, seed incorporation into the seed bank is lower on compacted than on non-compacted soils (Chambers and MacMahon Citation1994). Species richness and seed density in the deeper road soil layers is even lower (Qiaoling et al. Citation2005). Whenever ploughing is used as a restoration treatment, few seeds of target species are buried, and few seeds of target species are likely to be brought back to the surface. The topsoil of the foredune and the hedgerow was observed to contain slightly more species and more seeds, mainly consisting of Anthemis maritima and Plantago coronopus, two of the target species intended for the road restoration. We can therefore hypothesize that transferring soil from the foredune to the road will promote the dispersal of these target species and that, maybe, with enough time, the target species’ seeds can be wind dispersed from the foredune to the road, especially if the hedgerow is removed. As shown in other studies, vegetation restoration of sand dunes depends on the dispersal of seeds from nearby plant communities on the site to be restored (Qiaoling et al. Citation2005; Vestergaard Citation2013).
Restoration treatment
Although the restoration treatments did not allow the establishment of a striking species-rich dune vegetation in the short-term, they did promote the establishment of a few target species. Control sections, in which no restoration treatment was applied, were mainly composed of Juncus sp. and Parapholis sp., whereas the restoration treatments had much less Juncus sp. and were mainly composed of Parapholis sp., Salsola kali and Anthemis maritima. Although reintroducing plant species is a widespread technique to restore dune ecosystems (e.g. Doody Citation2013), ploughing is not. Dune restoration spans from re-vegetating and stabilizing the mobile substrate to removing plant cover and increasing substrate mobility (Lithgow et al. Citation2013; Martínez et al. Citation2013), so the available techniques are numerous and have to be adapted to the particular degradation encountered. Ploughing can be used to bury the upper nutrient-rich soil layer (topsoil inversion: Jones, Norman, and Rhind Citation2010), to rejuvenate sand movement (Pye, Saye, and Blott Citation2007; Rhind, Jones, and Jones Citation2013) or, as in this study, to loosen compacted soil. Deep ploughing or topsoil inversion can lead to promising results, but, as in the present study, vegetation colonization was rather slow (Jones, Norman, and Rhind Citation2010). Introducing species may speed up the restoration, but it may still take several years for them to actually establish (Hölzel and Otte Citation2003; Rozé and Lemauviel Citation2004). Here, we do not see any significant positive effect of soil transfer on target species establishment, but we can hope for more individuals of target species to germinate in the years to come. It might also be that soil transfers were not performed at the ideal season, which would be just after most plant species set their seeds. This aspect would have to be investigated and monitoring should be carried out to see if the foredune plant community will establish in the future.
The best treatment included ploughing on the road humps where there was no hedgerow. Humps might have environmental characteristics in general, and soil conditions in particular, that are more adapted to dune vegetation. This is in contrast to the dips, which are subject to flooding for a portion of the winter and more resemble the dune slacks (Doody Citation2013). Target species seeds may have been wind dispersed from the foredune to this treatment (road humps where there was no hedgerow), as the hedgerow gaps were not retentive of seeds. Removing the Tamarix gallica hedgerow could be considered and tested.
Conclusion
In our study, historical information was of little help in defining the reference ecosystem. The description of EU Habitat Directive habitat types and the selection of regionally less degraded sites allowed us to compile the following list of plant species that we could expect to find in the Jaï restored area: Sporobolus pungens, Elytrigia juncea and Anthemis maritima, which are halophilous species and Medicago littoralis, Plantago coronopus, Glaucium flavum, which are haloresistant species. The seed bank of the road does not have much potential for restoration, so ploughing was possible. At the same time ploughing is only efficient when the following other conditions are met: environmental conditions adequate for the establishment of dune vegetation, and a sufficient connection between the beach, the foredune and the restored area.
Acknowledgements
The implementation of this study has been made possible thanks to the support of the SIBOJAÏ – Espace Naturel Protégé du Bolmon et du Jaï and the Institut Méditerranéen de Biodiversité et d’Ecologie (IMBE). We thank the staff at SIBOJAÏ for their help and logistical support. We thank the various individuals who helped with our field work, in particular IMBE botanist Daniel Pavon, who provided invaluable assistance in identifying plants and seedlings. We also thank Jean-François Alignan, Prof. Catherine Fernandez, Dr Raphaël Gros, Dr Isabelle Laffont-Schwob and two reviewers for their suggestions on early versions of the manuscript.
References
- Acosta, A., S. Ercole, A. Stanisci, V. De Patta Pillar, and C. Blasi. 2007. “Coastal Vegetation Zonation and Dune Morphology in Some Mediterranean Ecosystems.” Journal of Coastal Research 23: 1518–1524.
- Bainbridge, D.A. 2007. A guide for desert and dryland restoration: new hope for arid lands. Washington, D.C.: Island Press.
- Bensettiti, F., F. Bioret, J. Roland, and J.-P. Lacoste. 2004. “Dunes fixées du littoral méditerranéen du Crucianellion maritimae.” In Cahiers d’habitats Natura 2000. Connaissance et gestion des habitats et des espèces d’intérêt communautaire. Tome 2 - Habitats côtiers, edited by MEDD/MAAPAR/MNHN, 319–320. Paris: La Documentation française.
- Carboni, M., R. Santoro, and A.T.R. Acosta. 2010. “Are some communities of the coastal dune zonation more susceptible to alien plant invasion?” Journal of Plant Ecology 3: 139–147.
- Chambers, J.C., and J.A. MacMahon. 1994. “A day in the life of a seed: movements and fates of seeds and their implications for natural and managed systems.” Annual Review of Ecology and Systematics 25: 263–292.
- Choi, Y.D. 2007. “Restoration ecology to the future: a call for new paradigm.” Restoration Ecology 15: 351–353.
- Clewell, A.F., and J. Aronson. 2007. Ecological Restoration: Principles, Values, and Structure of an Emerging Profession. Washington, D.C.: Island Press.
- Costa, J.C. 1997. “Dune vegetation of the Ria Formosa Natural Park in The Ecology and Conservation of European Dunes.” In The Ecology and conservation of European Dunes, edited by F. García Novo, R.M.M. Crawford and M.C. Díaz Barradas, 83–88. Sevilla: Universidad de Sevilla.
- Crouzet, N., D. Pavon, and H. Michaud. 2009. “Mise à jour de la liste des plantes vasculaires du département des Bouches-du-Rhône.” Bulletin de la Société Linnéenne de Provence 60: 55–73.
- Doody, J.P. 2013. Sand dune conservation, management and restoration. Berlin, Heidelberg: Springer.
- Fattorini, M., and S. Halle. 2004. “The dynamic environmental filter model: how do filtering effects change in assembling communities after disturbance?” In Assembly rules and restoration ecology—bridging the gap between theory and practice, edited by V.M. Temperton, R.J. Hobbs, T. Nuttle and S. Halle, 96–114. Washington, D.C.: Island Press.
- García Novo, F., M.C. Díaz Barradas, M. Zunzunegui, R. García Mora, and J.B. Gallego Fernández. 2004. “Plant functional types in coastal dune habitats.” In Ecological Studies, Vol. 171 – Coastal Dunes, Ecology and Conservation, edited by M.L. Martínez and N.P. Psuty, 155–169. Berlin Heidelberg: Springer-Verlag.
- GIPREB, 2007. “Histoire d’Etang. Vision d’Etang.” Le journal de GIPREB, 2: 8.
- Holl, K.D., and J. Jr. Cairns. 2002. “Monitoring and appraisal.” In Handbook of Ecological Restoration Volume 1: Principles of Restoration, edited by M.R. Perrow and A.J. Davy, 411–432. Cambridge: Cambridge University Press.
- Hölzel, N., and A. Otte. 2003. “Restoration of a species-rich flood meadow by topsoil removal and diaspore transfer with plant material.” Applied Vegetation Science 6: 131–140.
- Isermann, M. 2005. “Soil pH and species diversity in coastal dunes.” Plant Ecology 178: 111–120.
- Jones, M.L.M., K. Norman, and P.M. Rhind. 2010. “Topsoil inversion as a restoration measure in sand dunes, early results from a UK field-trial.” Journal of Coastal Conservation 14: 139–151.
- Lithgow, D., M.L. Martínez, J.B. Gallego-Fernández, P.A. Hesp, P. Flores, S. Gachuz, N. Rodríguez-Revelo, O. Jiménez-Orocio, G. Mendoza-González, and L.L. Álvarez-Molina. 2013. “Linking restoration ecology with coastal dune restoration.” Geomorphology 199: 214–224.
- Lortie, C.J., R.W. Brooker, P. Choler, Z. Kikvidze, R. Michalet, F.I. Pugnaire, and R.M. Callaway. 2004. “Rethinking plant community theory.” Oikos 107: 433–438.
- Mamarot, J., P. Psarski, and R. Rouquier. 1997. Mauvaises herbes des cultures. Paris: Acta.
- Martínez, M., M. Luisa, J.B. Gallego-Fernández, and P.A. Hesp. 2013. Restoration of Coastal Dunes. Berlin Heidelberg: Springer-Verlag.
- Mason, T.J., K. French, and K.G. Russell. 2007. “Moderate impacts of plant invasion and management regimes in coastal hind dune seed banks.” Biological Conservation 134: 428–439.
- Maun, M.A. 2009. The biology of coastal sand dunes. Oxford: Oxford University Press.
- MEA. 2005. Ecosystems and Human Well-being: Synthesis. The Millennium Ecosystem Assessment. Washington, DC: Island Press.
- Molinier, R. ( in collaboration with Martin, P.). 1981. Catalogue des Plantes vasculaires des Bouches-du-Rhône. Marseille: Impr. Municipale.
- Muséum national d’Histoire naturelle. 2003–2013. Inventaire national du Patrimoine naturel. Accessed April 14 2014. http://inpn.mnhn.fr.
- Nordstrom, K.F. 2008. Beach and dune restoration. Cambridge: Cambridge University Press.
- Parker, V.T., and S.T.A. Pickett. 1997. “Why restoration?” In Restoration ecology and sustainable development, edited by K.M. Urbanska, N.R. Webb and P.J. Edwards, 17–32. Cambridge: Cambridge University Press.
- Pye, K., S. Saye, and S. Blott. 2007. Sand dune processes and management for flood and coastal defence. Part 4: Techniques for sand dune management. Joint Defra/EA Flood and Coastal Erosion Risk. Department for Environment, Food and Rural Affairs. London: Department for Environment, Food and Rural Affairs.
- Qiaoling, Y., L. Zhimin, Z. Jiaojun, L. Yongming, W. Hongmei, and J. Deming. 2005. “Structure, pattern and mechanisms of formation of seed banks in sand dune systems in Northeastern Inner Mongolia, China.” Plant and Soil 277: 175–184.
- R Development Core Team 2008. R: A language and environment for statistical computing. R Foundation for Statistical Computing, Vienna, Austria. Accessed April 14 2014. http://www.R-project.org.
- Rhind, P., R. Jones, and L. Jones. 2013. “The impact of dune stabilization on the conservation status of sand dune systems in Wales.” In Restoration of Coastal Dunes, edited by M.L. Martínez, J.B. Gallego-Fernández and P.A. Hesp, 125–143. Berlin Heidelberg: Springer-Verlag.
- Rozé, F., and S. Lemauviel. 2004. “Sand dune restoration in north Brittany (France): a 10 year monitoring study.” Restoration Ecology 12: 29–35.
- SER. 2004. The SER International Primer on Ecological Restoration. [html pages]. Society for Ecological Restoration International Science & Policy Working Group. Accessed October 2013. http://www.ser.org/resources/resources-detail-view/ser-international-primer-on-ecological-restoration.
- Ter Heerdt, G.N.J., G.L. Verweij, R.M. Bekker, and J.P. Bakker. 1996. “An improved method for seed-bank analysis: Seedling emergence after removing the soil by sieving.” Functional Ecology 10: 144–151.
- Thompson, K., J. Bakker, and R. Bekker. 1997. The soil seed banks of North West Europe: methodology, density and longevity. Cambridge: Cambridge University Press.
- van der Valk, A.G. 1975. “The floristic composition and structure of foredune plant communities of cape hatteras national seashore.” Chesapeake Science 16: 115–126.
- Vestergaard, P. 2013. “Natural plant diversity development on a man-made dune system.” In Restoration of Coastal Dunes, edited by M.L. Martínez, J.B. Gallego-Fernández and P.A. Hesp, 49–66. Berlin Heidelberg: Springer-Verlag.
- Willems, J.H., and L.P.M. Bik. 1998. “Restoration of high species density in calcareous grassland: the role of seed rain and soil seed bank.” Applied Vegetation Science 1: 91–100.