
Abstract
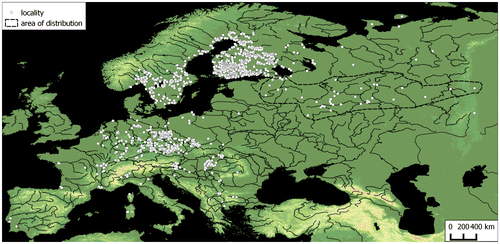
Elatine triandra Schkuhr is the most variable and widespread species within the genus Elatine L.; it has been recorded in all continents, except Antarctica, but it is mainly located in Europe. The study is based on an extensive data set of European literature, herbaria and web data that covers the period 1828–2012. The range of the species in Europe is disjunctive, covering the southern and western parts of the Central European Plain and the southern part of the Fennoscandian Shield. At a smaller scale, the species can also be found along some river valleys. In Central Europe many localities, particularly isolated ones in the northern part of the range, are now only historical. From the data set we determined that E. triandra may be best observed between May and October. We found that species records show a near-significant shift since 1828. Depending on the environmental conditions, individuals of the taxon develop as one of two morphs: terrestrial or aquatic. The aquatic morph is characterized by stems, internodes, lamina and petiole that are twice as long as those of the terrestrial form. Elatine triandra seeds show consistent characteristics, both in terms of morphs and populations. Our studies show that the best diagnostic features, in addition to the construction of flowers, are the size, shape and surface structure of seeds. In Central Europe, E. triandra occurs exclusively in communities classified as Isoëto-Nano-Juncetea.
Introduction
Understanding the distribution patterns and biology of rare and endangered species is essential for the development of nature conservation policies. Many species of the class Isoëto-Nano-Juncetea are rare elements of the European flora, and many are threatened with extinction. From this perspective, species of the genus Elatine L. are especially important. The genus includes about 27 species of small ephemerophytes that appear periodically on bare soil in intermittent water bodies. Their range includes areas with temperate climate on all continents, except Antarctica. The genus Elatine belongs to the family Elatinaceae; however, the origin and taxonomic relations of the family are not fully clarified (Seubert Citation1845; Niedenzu Citation1925; Davis and Chase Citation2004; Popiela and Łysko Citation2011; Popiela et al. Citation2012; Kubitzki Citation2013).
The distribution patterns of the genus Elatine are still relatively unknown. Comprehensive studies have been carried out only on Elatine alsinastrum L., Elatine hexandra (Lapierre) DC., Elatine hydropiper L., Elatine macropoda Gruss. and Elatine hungarica Moesz (e.g. Meusel Citation1978; Hultén and Fries Citation1986; Lampe Citation1996; Popiela and Łysko Citation2010; Popiela et al. Citation2011, 2012; Takács et al. Citation2013). The species of the genus are distributed mostly in the northern hemisphere, predominantly in areas with temperate climate. Some species have wide geographical ranges, such as E. alsinastrum and E. hydropiper (Popiela et al. Citation2012; Popiela, Łysko, and Molnár Citation2013). Others are limited only to Europe, such as E. hexandra (Popiela et al. Citation2011) and E. hungarica (Takács et al. Citation2013), or are known from very restricted areas, such as Elatine paramoana Schmidt-M. & Bernal (Schmidt-Mumm & Bernal Citation1995), Elatine ojibwayensis Garneu (Garneu Citation2006) and Elatine gussonei (Sommier) Brullo Lanfr., Pavone and Ronsisv. (Molnár, Popiela, and Lukács Citation2013).
Elatine triandra Schkuhr [synonyms: Elatine callitrichoides (Nyl.) Kauffm., Elatine chilensis Gray, Elatine inaperta J. Lloyd, Elatine triandra var. callitrichoides Nyl., Elatine orientalis Makino] is the most widespread species within the genus, occurring on all continents, except Antarctica; however, the majority of its recorded locations are in Europe (e.g. Tucker Citation1986). Elatine triandra belongs to the section Triandra Seub. [= Crypta (Nutt.) Seub.], which is characterized by trimerous flowers showing haplostemony (an arrangement of 16 stamens in a single whorl opposite the sepals with equal numbers of anthers and sepals). Elatine triandra is a species of small aquatic and terrestrial plants with a prostrate stem with many secondary branches, rooted at nodes. Leaves are opposite, blade lanceolate-linear to narrowly lanceolate to oblong with distinct hydatodes, and triangular stipules. Flowers are single at the nodes, sessile or rarely very short-stalked, with two to three sepals and usually three petals and three stamens. Capsules are subglobose, somewhat depressed at the apex, seeds cylindrical, greenish- to yellowish-brown with conspicuous pits [2n = 40] (Tucker Citation1986; Uotila Citation2010).
The aims of this paper are: (1) to present the current distribution of E. triandra in Europe; (2) to describe the taxonomic and phenological features of the species; and (3) to indicate the communities in which the species occurs and its conservation status in the central part of the continent.
Material and methods
Research on the distribution of E. triandra was based on analysis of floristic literature, distribution atlases, web sources and examination of specimens deposited in the herbaria listed in Table .
Table 1. Literature, web and herbaria sources used. Herbarium acronyms follow Index Herbariorum (Holmgren and Holmgren Citation1998).
Data on the distribution of the species were gathered, giving priority to the literature over web sources. The data set contains 1959 records (1353 from the literature and herbaria, 606 from the internet), covering the period 1828–2012. Web sources were used only if considered to be reliable (e.g. if they did not deviate from the range or had an author and exact location or were delivered by herbarium servers). Altogether, 427 web data were removed.
The exact time of species observation was recorded. The observation dates were transformed into Julian days. We calculated the relative frequency of species observations within a year; then the species phenology in time was analysed. The median Julian date of observations was plotted against time (1828–2012) and linear regression was computed. All analyses were made using Statistica 7.0 (StatSoft Inc., Tulsa, OK).
The differences in seed and vegetative characteristics between the aquatic and terrestrial morphs of E. triandra were studied in vitro. Seeds of E. triandra were collected from indigenous populations in Hungary: Kisköre (47°29'37.42" N, 20°29'17.75" E). The plants were cultivated in the laboratory of the University of Debrecen. Seeds were sown in 12.5 × 8.5-cm plastic boxes on sterilized (autoclaved for 3 h at 180°C) soil that was continuously wetted. Plantlets were grown in climate-controlled rooms (with 14 h of light/day and 30 μmol m−2 sec−1 light intensity, temperature in light 22 ± 2°C and in darkness 18 ± 2°C).
Two 1-week-old seedlings were transplanted, then kept under aquatic conditions (continuous water cover) or terrestrial conditions (on wet mud) until they reached the fruiting stage. For the morphological study, six characters (length of stem, length of first mature internode, length of first mature lamina, width of first mature lamina, length of petiole, number of nodes) were measured on 50–50 fruiting stems of both aquatic and terrestrial morphs using calipers (0.1-mm accuracy). Four characters [curvature (°), number of pits/lateral row, first dimension (mm), second dimension (mm)] were measured from digital images (Figure ).
Figure 1. The method of seed measurement.
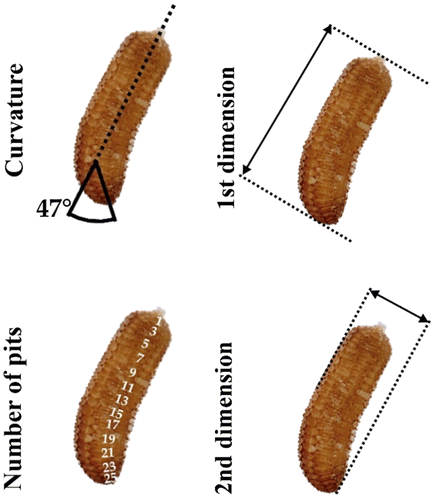
To determinate the variability and diagnostic features of seeds of E. triandra, 50 seeds obtained from each of two Central European populations (a total of 100 seeds) approximately 600 km apart were studied: Poland: Janików (51°34'34.52" N, 14°57'23.35" E), and Hungary: Kisköre (47°29'37.42" N, 20°29'17.75" E). A total of 100 scanning electron microscope images of the seeds were obtained at × 300 magnification using a Zeiss Evo LS10. Characters were measured according to the method described above (Figure ).
The occurrence of E. triandra in plant communities in Central Europe (Poland, southeast Germany, the Czech Republic and Slovakia, Hungary, and northern Italy) was studied using published synoptic tables in which the species had 60% constancy. The data from Poland were obtained from the Polish Vegetation Database (Kącki and Śliwiński Citation2012). This data set was supplemented with unpublished relevés (AMV, Hungary; AP, Poland, Germany; ZK, Poland). A total of 614 published relevés were used. We prepared the synoptic table using the Juice program (Tichý Citation2002) and showed species with constancy > 10%. Vegetation was classified according to the assignment of the authors of the original studies (Ubrizsy Citation1948, Citation1961 after Pietsch Citation1973a; Pignatti Citation1957 after Pietsch Citation1973b; Pietsch Citation1963, Citation1973a, b; Popiela Citation1996; Nowak and Nowak Citation2007; Šumberová and Hrivnák Citation2013). Unpublished data were assigned to particular syntaxonomic units using the Frequency-Positive Fidelity Index (Tichý Citation2005).
Results
In Europe, the main area of E. triandra extends from the north of the Alps to the southern part of the Scandinavian Peninsula and the Finnish Lake District, including the southern and western parts of the Central European Plain (primarily the Saxo-Lusatian Lowlands) and the northern shores of the Baltic Sea. Elatine triandra can also be found in single locations in the southern part of the Odra River valley and in the Elbe and Rhine valleys (Figure ). Larger centres of occurrence are present in the Upper Saône valley, in the southeastern part of the Franconian Jura, in the Bohemian–Moravian Highlands, in the Pannonian Basin and along river valleys in the eastern part of the Alps (Figure ). Small site clusters also occur in the Po valley, in the Maritsa River valley and at the mouth of the Rhône River. In addition, E. triandra has scattered locations: the westernmost sites are at the mouths of the Loire and the Tagus, the most southern locations are in Corsica and Sardinia and in southeastern continental Greece. The northernmost locations are on the northern shores of the Gulf of Bothnia and in the mouth and valley of the Torne River. Elatine triandra is not known from eastern Poland, Ukraine and Belarus and only one location is known in Lithuania. The species is reported from the eastern part of the East European Plain, beyond a large gap in its range.
Figure 2. The distribution of Elatine triandra in central and the eastern part of Western Europe.
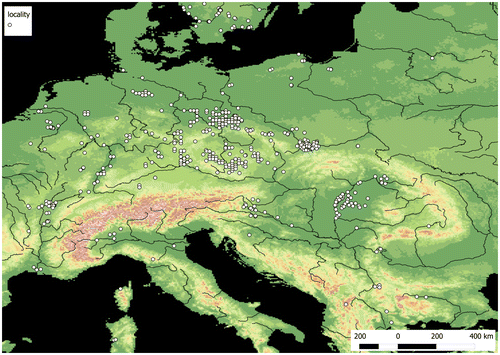
Figure 3. The distribution of Elatine triandra in Europe.
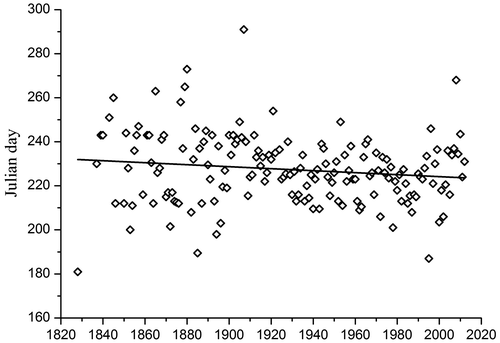
Concerning the phenology of E. triandra, the species can be found between May and October, but the main season is from July to September (Figure ). Linear regression between the median Julian day of observation as the dependent variable and years as the explanatory variable indicated that the first observation of the year of E. triandra has shifted to 8 days earlier (R = −0.142, p = 0.0676) since 1828 (Figure ).
Figure 4. Relative frequency of Elatine triandra collections.
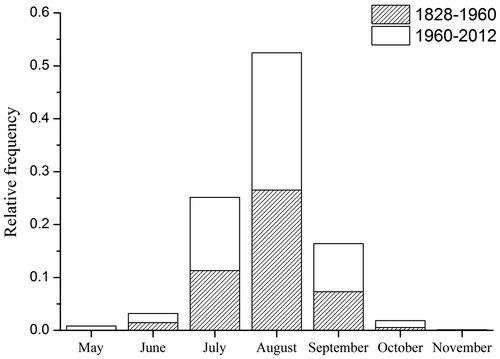
Figure 5. Dates of observation of Elatine triandra in a historical data set. Dots represent the medians of Julian day. Black lines are linear fit (Pearson’s r = –0.1422).
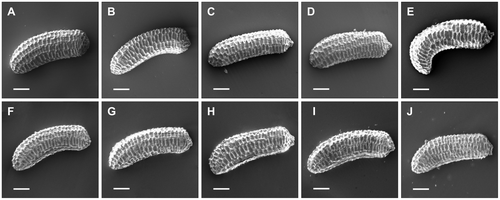
Depending on the environmental conditions, individuals of E. triandra develop as one of two morphs: terrestrial or aquatic. The aquatic morph has stems and internodes twice as long as those of the terrestrial morph, and also lamina and petiola almost doubled in length, but there are no differences in terms of lamina width or the number of nodes (Table , Figure ). Elatine triandra seeds show consistent characteristics for each morph, as well as for populations originating from different parts of its range: length c.0.5 mm, straight or slightly bent in one dimension, and the number of pits in the seed coat is usually > 20 (Tables , Figure ).
Table 2. Comparison of the median value of seed and vegetative characteristics between two different modifications of Elatine triandra.
Figure 6. Morphology of terrestrial and aquatic modifications of Elatine triandra.

Table 3. Values of seed characteristics of the two Elatine triandra populations studied (Hungary, n = 50; Poland, n = 50).
Figure 7. The diversity of seeds of Elatine triandra. Scale bars represent 0.1 mm.
In Central Europe, E. triandra occurs exclusively in communities entering into the Isoëto-Nano-Juncetea class, mainly on the bottoms and in open water of fish ponds and on rice fields (Table ), most often accompanied by Eleocharis acicularis (L.) Roem. & Schult., Lythrum portula (L.) D.A. Webb, Filaginella uliginosa (L.) Opiz and Juncus bufonius L. In the north, important constituents of the associated flora are Eleocharis ovata (Roth) Roem. & Schult., Carex bohemica Schreb. and E. hexandra, and in the south Lindernia procumbens (Krock.) Philcox and E. hungarica. Among the accompanying species, the predominant taxa are associated spatially with the waters and wetlands.
Table 4. Synoptic table of the vegetation groups (1–11) of the Isoëto-Nanojuncetea class characterized by the presence of Elatine triandra with high degrees of constancy (III, IV, V) in the area of Central Europe (the table contains species reaching more than 20% of the presence only).
In Central Europe after 1990, populations of E. triandra were mostly recorded in four areas: the Lusatian Lowland, the Lower Odra and Vistula river valleys, the Bohemian–Moravian Highlands, and the Pannonian Basin (Figure ). Many localities, especially isolated ones in the northern part of the range, are historical now. Therefore, the species has been included in national red books and lists of threatened species such as the IUCN list (IUCN 2014; Table ).
Figure 8. The diversity of Elatine triandra recording over time in Central Europe. 1, after 1990 (in Germany 1980), 2, in the years 1945–1990 (in Germany 1945–1980); 3, before 1945; 4, date unknown. Younger value covers the older one.
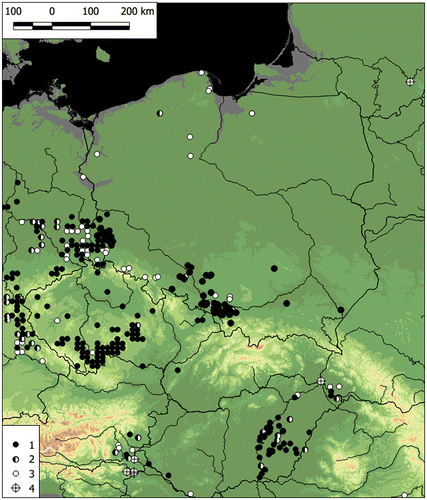
Table 5. Red list categories of Elatine triandra (IUCN, national in Central Europe).
Discussion
The range of E. triandra in Europe is disjunctive. A characteristic pattern of distribution was observed; it occurs in small or large clusters or alone, frequently along river valleys; the distances between locations are frequently as much as several hundred kilometres. In this respect, the distribution resembles that of other species in the same genus. This type of distribution is evidence that colonization of new areas is a slow process, although wet habitats and places that have been recently exposed after earlier floods and are devoid of other plants are not rare habitat types. The method of propagation of E. triandra in Europe is unclear (as it is for other Elatine species) and there is a lack of information about the location of the Pleistocene refuge area. Presumably the distribution range reflects the migration of waterfowl transferring diaspores (seeds and vegetative particles) from one place to another. Although this phenomenon is poorly documented (Kerner Citation1895), we suggest that diaspore transfer of Elatine species via waterfowl is probable. However, this mechanism does not explain the large gap in its distribution range, which includes the commonest places of migrating birds with suitable habitats – e.g. there are no locations with E. triandra in the Warta River Mouth National Park (northwest Poland), which is one of the most important migration places of waterfowl in Central Europe.
The range of E. triandra in the East European Plain seems to be poorly known: the species has been reported in Russian floras without detailed locations, and so determination of the eastern boundaries of its range can be made only provisionally. At present, it is not possible to establish whether the lack of sites is caused by insufficient reconnaissance or because sites are naturally sparse. It seems quite certain that E. triandra is not present in the southern part of the East European Plain, most likely because of the dry, continental climate. The complete lack of occurrence in the western part of the East European Plain is very interesting. If this pattern is natural (not an artefact caused by lack of collection), that would suggest that in Central Europe the species has reached its limit of distribution density.
Land-use activities appear to have a great impact on the distribution range of E. triandra by forming suitable habitats for the species: rice paddies, fisheries, temporarily inundated agricultural fields and grazed shores. The question remains whether these are substitute habitats after anthropogenic destruction of many natural habitats, for instance after river regulation, or whether the formation of new habitats has expanded the range anthropogenically.
The high variability of plant size, leaf shape and size as well as the length of internodes and pedicels (Mason Citation1956) are a characteristic feature of E. triandra and other Elatine species with trimerous flowers showing haplostemony [e.g. Elatine ambigua Wight, Elatine gracilis Mason, Elatine gratioloides Cunn, Elatine oryzetorum Kom., Elatine brachysperma Gray, Elatine fassetiana Steyermark, Elatine ecuadorensis Molau, Elatine lorenziana Hunz., E. paramoana Schmidt-Mumm and Bernal, Elatine peruviana Baehni and J.F. Macbr. and Elatine americana Pursh (Arn.)], (Walters Citation1969; Brinkkemper et al. 2008; Uotila Citation2009, Citation2010; Molnár et al. Citation2013). According to our herbarium studies (made in the Kew Herbarium by the first author) the accuracy of identification of these species is low. It is possible that these taxa represent different morphs within the species E. triandra, and so may be synonyms. Conversely, E. triandra s.s. may not be widespread. Tall plants with long internodes occurring in northern Europe have been distinguished as E. callitrichoides (Uotila Citation2010). In our opinion, a final decision about taxonomic problems of this taxon as well as other taxa from the section Crypta will require collection of plants throughout the world and use of molecular methods.
In terms of the phenology of E. triandra, we detected a historical decrease in the first observation of the year since 1828. It is difficult to establish reasons for this pattern. The observed shift in the appearance of E. triandra is thought to have been caused by changes in precipitation patterns; however, this could not be confirmed from the environmental data set. The shifts may also have been caused by global climate warming and the related acceleration of the growing season, or they may simply be an artefact resulting from collection activity.
Our studies confirm that the best features for identification of E. triandra, in addition to flower construction, are the size, shape and surface structure of seeds. Vegetative characters such as the lengths of the stem, internodes, lamina and petioles are variable and depend on water level. The vegetative values obtained in our experiment deviate from the measures from North Europe (Uotila Citation2010), namely flooded plants are smaller. This can be explained by the fact that under natural conditions in the field the size of the plant depends on environmental factors, most probably on the fertility of the waters.
Elatine triandra can be found on recently exposed soil in dried-out fishing ponds and on river banks and gently sloping shores of water bodies, as well as in rice paddies. It appears mainly in patches of Polygono–Eleocharitetum ovatae of the Eleocharition ovatae alliance (e.g. Moor Citation1937; Brullo and Minissale Citation1998; Šumberová Citation2011; Šumberová and Hrivnák Citation2013; Kącki, Czarniecka, and Swacha Citation2013), Oryzo–Cyperetum difformis (Pignatti Citation1957) and Eleocharito acicularis–Schoenoplectetum supini (Pietsch Citation1973b; Brullo and Minissale Citation1998) associations. In Hungary, the species was identified as an indicator species of rice paddy fields, and it also occurs in continental temporary ponds, mostly in inundated agricultural fields (Lukács, Sramkó and Molnár Citation2013). In the Scandinavian Peninsula, the species is mainly found in mesotrophic and eutrophic lakes, rivers and ponds (Murphy Citation2002; Uotila Citation2010). Our studies show that, in Central Europe, E. triandra occurs exclusively in communities classified as Isoëto-Nano-Juncetea, and is accompanied by 24 character taxa of the class.
As with other ephemeral vegetation, patches with E. triandra develop in different ways depending on the local habitat conditions and dynamic stage. These patches are developed most often on anthropogenic sites at the bottoms and edges of fish ponds and, in the southern part of the area studied, in rice paddies. Elatine triandra occurs in these patches, usually as the terrestrial morph.
Our research has also shown that at present in Central Europe populations of E. triandra are best preserved in western and southern Poland, in southeast Germany, the Czech Republic and eastern Hungary, and are under adequate protective status in these areas. Single isolated populations deserve special attention, because they may be the first to disappear.
Notes on contributors
Agnieszka Popiela’s interests lie in taxonomy of Elatinaceae, chorology of vascular plants and ephemeral vegetation. Contribution: the idea and project coordination, field and/or laboratory research, preparing the manuscript.
Andrzej Łysko’s interests include the use of GIS to analyze the distribution of plants. Contribution: database and maps preparation, field and/or laboratory research, preparing the manuscript.
Attila V. Molnár’s research interest covers the ecology, distribution and taxonomy of Orchidaceae and Elatinaceae. Contribution: field and laboratory research, preparing the manuscript.
Zygmunt Kącki’s interests focus on diversity and ecology of plant communities, and ecoinformatics. Contribution: field research, phytosociological data analyzing, preparing the manuscript.
Balázs András Lukács’s interests lie in ecology and conservation of freshwater vascular plants and vegetation dynamics. Contribution: field and/or laboratory research, data analysis, preparing the manuscript.
Acknowledgements
We wish to thank the staff of the Centre for Molecular Biology, University of Szczecin – Bożena Białecka and Magdalena Bihun for taking the scanning electron micrographs, Zygmunt Dajdok and Arkadiusz Nowak for providing unpublished relevés to the Polish Vegetation Database and the keepers of herbarium for access to the herbarium material. The authors express also their gratitude to the anonymous reviewer for detailed comments that greatly improved the manuscript.
Additional information
Funding
Related Research Data
References
- Adylov, T.A. 1983. Conspectus Florae Asiae Mediae, vol. 7. Moscow – Tashkent: USSR Academy of Sciences/Uzbek Academy of Sciences.
- Benkert, D., F. Fukarek, and H. Korsch, eds. 1996. Verbreitungsatlas der Farn- und Blütenpflanzen Ostdeutschlands [Distribution Atlas of Pteridophytes and Vascular Plants]. Jena: Gustav Fischer Verlag.
- Berten, R., ed. 1993. Limburgse plantenatlas 2. [Atlas of plants of the province of Limburg]. Likona, Belgium: Limburgde Koepel Voor Natuur Studie.
- Bilz, M., S.P. Kell, N. Maxted, and R.V. Lansdown. 2011. European Red List of Vascular Plants. Luxembourg: Publications Office of the European Union.
- Blazenčič, J., and Z. Blazenčič. 1999. “Elatine triandra Schkuhr,”, in The Red Databook Flora of Serbia. 1. Extinct and critically endangered taxa, edited by V. Stefanovič,p. 492. BelgradE: Privo Izdanje.
- Blečič, V. 1972. “Elatinaceae.” in Flore de la République Socialiste de Serbie 3, edited by M. Josifovič, 126–128. Belgrad: Académie Serbe des Sciences et des Arts.
- Brinkkemper, O., E.J. Weeda, S.J.P. Bohncke, and W.J. Kuijper. 2007. “The past and present occurrence of Elatine and implications for palaeoenvironmental reconstructions.” Vegetation History and Archaeobotany 17: 15–24.
- Brullo, S., and P. Minissale. 1998. “Considerazioni sintassonomicha sulla classe Isoëto-Nanojuncetea.” [Taxonomic considerations of the class Isoëto-Nanojuncetea] Itinera Geobotanica 11: 263–290.
- Bugnon, F., J.C. Felzines, R. Goux, J.E. Loiseau, and J.M. Royer. 1993–1998. “Nouvelle Flore de Bourgogne, tome III. Atlas de repartition, clés des groupements végétaux et suppléments aux tomes I et II.“ Dijon: Bulletin Scientifique de Bourgogne, H.S.
- Cesati, V., G. Passerini, and G. Gibelli. 1868–1886. Compendio della flora Italiana. [Compendium of the Italian Flora]. Milano: Fr. Valladi.
- Cirujano S. and M. Velayos. 1993. “Elatine L.”, in Flora Iberica. Plantas vasculares de la Península Iberíca e Islas Baleares [Flora Iberica. Vascular Plants of the Iberic Peninsula and the Balearic Islands], vol. 3, Plumbaginaceae (partim)-Capparaceae, edited by S. Castroviejo et al., 153–156. Madrid: Real Jardín Botánico, Consejo Superior de Investigaciones Científicas.
- Davis, C.C., and M.W. Chase. 2004. “Elatinaceae are sister to Malpighiaceae; Peridiscaceae belong to Saxifragales.al.” American Journal of Botany 91: 262–273.
- Dedkov V.P., and G.V. Grišanov, ed. 2010. Krasna kniga Kaliningraskoj Oblasti. [The red book of the Kaliningrad Region]. Kaliningrad, Russia: Izdatel`stvo RGU im. I. Kanta.
- Desfayes, M. 2008. “Flore vasculaire herbacée des eaux douces et des milieux humides de la Sardaigne.” Flora Mediterranea 18: 247–331.
- Dostál, J. 1989. New flora of Czechoslovakia 1, 2. Prague: Academia.
- Fiori, A., and G. Paleotti. 1896. Flora analitica d’Italia [Analytical Flora of Italy]. Padova, Italy: Tipografia del Seminario.
- Franco, J.A. 1971. Nova Flora de Portugal, vol. 1 [Novel Flora of Portugal]. Lisbon: Edição do Autor.
- Fukarek, F., and H. Henker. 2006. Flora von Mecklenburg-Vorpommern. Farn- und Blütenpflanzen. [Flora of Mecklenburg-Pomerania. Ferns and Flowering Plants]. Jena, Germany: Weissdorn Verlag.
- Fuss, M. 1866. Flora Transsilvaniae exursoria. Hermannstadt, Romania: Societas naturae curiosorum Transsilvanica.
- Garneu, M. 2006. “Elatine ojibwayensis sp. nov., une nouvelle espèce d’Elatinaceae et revue des Elatinaceae du Québec.” Canadian Journal of Botany 84: 1037–1042.
- Gatterer, K., and W. Nezadal ed. 2003. Flora des Regnitzgebietes. Die Farn- und Blütenpflanzen im zentralen Nordbayern, vol. 1. [Flora of the Regnitz (river) area. Ferns and flowering plants of the central part of North Bavaria]. Eching, Germany: IHW Verlag.
- Gerstberger, P., and H. Vollrath. 2007. Flora Nordostbayerns [Flora of North East Bayren]. Bayreuth, Germany: Naturwissenschaftliche Gesellschaft.
- Grulich, V. 2012. “Red List of vascular plants of the Czech Republic: 3rd edition.” Preslia 84: 631–645.
- Haeupler, H., and P. Schönfelder. 1989. Atlas der Farn- und Blütenpflanzen der BRD. 2. [Atlas of ferns and flowering plants of German Federal Republic]. Stuttgart: Ulmer Verlag.
- Hejný, S., and B. Slavík. 1990. Květena České republiky. [Flora of the Czech Republic], vol. 2, 1st ed. Prague: Academia.
- Holmgren, P.K., and N.H. Holmgren. 1998. Index Herbariorum. A global directory of public herbaria and associated staff. New York, NY: New York Botanical Garden.
- Hultén, E., and M. Fries 1986. Atlas of North European vascular plants, north of the tropic of Cancer. 1. Königstein, Germany: Koeltz Scientific Books.
- Jakab, G. 2005. “New data to the flora of South Tiszántúl region II.” Flora Pannonica 3: 91–119.
- Jeanmonod, D., and J. Gamisans. 2007. Flora Corsica. Aix-en-Provence, France: Edisud.
- Jogan, N., ed. 2001. Gradivo za Atlas Flore Slovenije. [Materials for the Atlas of Flora of Slovenia]. Miklavž na Dravskem polju, Slovenia: Center za kartografijo favne in flore.
- Jordanov, D., and D. Peev. 1979. “Elatinaceae Cambees.”, in Flora Reipublicae Popularis Bulgaricae, edited by D. Jordanov, 418–420: Sophia, Bulgaria: Aedibus Academiae Scientiarum Bulgaricae.
- Kaźmierczakowa, R., K. Zarzycki, and Z. Mirek, ed. 2014. Polska Czerwona Księga Roślin. Paprotniki i rośliny kwiatowe. [Polish Red Data Book of Plants. Pteridophytes and flowering plants]. Kraków: Instytut Ochrony Przyrody Polish Academy of Science.
- Kącki, Z.M. Czarniecka, and G. Swacha 2013. “Statistical determination of diagnostic, constant and dominant species of the higher vegetation units of Poland”. Monographiae Botanicae 103:1–267.
- Kącki, Z., and M. Śliwiński. 2012. “The Polish Vegetation Database: structure, resources and development.” Acta Societatis Botanicorum Poloniae 81(2): 75–79.
- Kerner von Marialaun, A. 1895. The Natural History of Plants. Their forms, growth, reproduction, and distribution. 2. London: Blackie and Son.
- Király, G., ed. 2007. Vörös Lista. A magyarországi edényes flóra veszélyeztetett fajai [Red list of the vascular flora of Hungary]. Sopron, Hungaria: Saját kiadás.
- Koch, M., K.G. Berndhardt, and J. Webhofer. 2005. “Alte Teichlandschaften im osterreichischen Waldviertel: Vegetation und Diasporenpotential des Schonauer Teiches bei Zwett. [Old pond landscapes in the Austrian Waldviertel: Vegetation and diasporas potential of Schonauer pond in Zwettl].” Botanische Jahrbücher für Systematik, Pflanzengeschichte und Pflanzengeographie 126: 133–148.
- Kubitzki, K. 2013. Flowering Plants. Eudicots: Malpighiales (The Families and Genera of Vascular Plants). Heidelberg, New York, Dordrecht, London: Springer.
- Kuusk, V., L. Tabaka, and R. Yankyavichene, ed. 1996. Flora of the Baltic countries: compendium of vascular plants. Tartu, Estonia: Eesti Loodusfoto AS.
- Lampe von, M. 1996. “Wuchsform, Wuchsrhythmus und Verbreitung der Arten der Zwergbinsengesellschaften. Growth form, growth rhythm and distribution of the species of ephemeral vegetation.” Dissertationes Botanicae 266: 1–351.
- Lampinen, R., T. Lahti, and M. Heikkinen. 2015. Kasviatlas 2014 [Atlas of plants 2014]. Helsingin Yliopisto, Luonnontieteellinen keskusmuseo, Helsinki. Levinneisyyskartat osoitteessa http://www.luomus.fi/kasviatlas, accessed 2015-23-07.
- De Langhe, J.E., L. Delvosalle, J. Duvigneaud, J. Lambinon, and C. Vanden Berghen, eds. 1983. Flora van België, het Groothertogdom Luxemburg, Noord-Frankrijk en aangrenzende gebieden. [Flora of Belgium, the Grand Duchy of Luxembourg, Northern France and adjacent areas]. Meise, Belgium: Nationale Platentuin van Belgie.
- Lawalrée, A. 1968. “Elatine triandra Schkuhr (Elatinaceae) à Frahan (Belgique).” Bulletin du Jardin Botanique National de Belgique 38: 397–400.
- Ludwig, G., and M. Schnittle, ed. 1996. “Rote Liste gefährdeter Pflanzen Deutschlands. [Red list of endangered plants in Germany].” Schriftenreihe für Vegetationskunde 28: 1–744.
- Lukács, B.A., G. Sramkó, and A. Molnár Vc. 2013. “The plant diversity and conservation value of continental temporary pools.” Biological Conservation 158: 393–400.
- Maglocký, Š., and V. Feráková. 1993. “Red list of ferns and flowering plants (Pteridophyta and Spermatophyta) of the flora of Slovakia (the second draft).” Biológia (Bratislava) 48(4): 361–385.
- Mason, H.L. 1956. “New species of Elatine in California.” Madroño 13: 239–240.
- Maurer, W. 1996. Flora der Steiermark. 1. Farnpflanzen (Pteridophyten) und freikoronblättige Blütenpflanzen (Apetale und Dialypetale). [Flora of Styria. 1. Ferns (Pteridophytes) and free petals flowering plants (Apetale and Dialypetale)]. Eching, Germany: IHW Verlag.
- Meusel, H. ed. 1978. Vergleichende Chorologie der zentraleuropäischen Flora. 2. Textband. [Comparative chorology the central European flora. 2. Text volume]. Jena, Germany: G. Fischer Verlag.
- Micevski, K. 1995. The flora of the Republic of Macedonia 1(3). Skopje, Macedonia: Macedonian Academy of Sciences and Arts.
- Molnár, V.A., and G. Gulyás. 2001. “Data to the knowledge of Nanocyperion-species VII.” Kitaibelia 6: 169–198.
- Molnár, V.A., and N. Pfeiffer. 1999. “Data to the knowledge of Nanocyperion-species II.” Kitaibelia 4: 391–421.
- Molnár, V.A., O. Horváth, J. Tökölyi, and L. Somlyay. 2013. “Typification and seed morphology of Elatine hungarica (Elatinaceae).” Biologia 68: 210–214.
- Molnár, V.A., A. Popiela, and B.A. Lukács 2013. “Elatine gussonei (Sommier) Brullo et al. (Elatinaceae) in Sicily.” Plant Biosystems 148(1): 27–30.
- Moor, M. 1937. “Zur Soziologie der Isoëtalia. [Towards the sociology of the Isoëtalia].” Beiträge zur geobotanischen Landesaufnahme der Schweiz 20: 1–48.
- Muller, S. 1995. “Elatine triandra Schkuhr.”, in Livre Rouge de la flore menacée de France. Tome I: Espèces prioritaires, edited by L. Olivier, J.P. Galland, and H. Maurin. Paris: Collection Patrimoines Naturels (Série Patrimoine Génétique). 20. SPN - IEGB /MNHN, DNP/Ministère de l’Environnement, CBN de Porquerolles. lxxxv + 486 fiches descriptives.
- Muller, S. 2006. Les plantes protégées de Lorraine: distribution, écologie, conservation, biotope. Mèze: Parthénope.
- Murphy, K.J. 2002. “Plant communities and plant diversity in softwater lakes of northern Europe.” Aquatic Botany 73: 287–324.
- Netien, G. 1993. Flore lyonnaise. Lyon: Société Linnéenne de Lyon.
- Niedenzu, F. 1925. “Elatinaceae.” In Die natürlichen Pflanzenfamilien [The Natural Plant Families], edited by A. Engler and K. Prantl, 270–276. Leipzig: W. Engelmann.
- Nikolić, T. 1994. “Index Florae Croaticae 1.” Natura Croatica 3 (Suppl. 2): 1–116.
- Nowak, A. and S. Nowak 2007. “Interesujące zbiorowiska roślinne w dolinie Odry na Śląsku Opolskim. [Interesting plant communities in the valley of the Oder River in Silesia Opole]”, in Przyrodnicze wartości polsko-czeskiego pogranicza [Nature of the Polish-Czech border], edited by J.A. Lis, and M.A. Mazur, 59–81. Opole: Centrum Studiów nad Bioróżnorodnością, Uniwersytet Opolski.
- Peev, D. 1984. “Elatine triandra Schkuhr.”, in Red Data Book of the PR Bulgaria, 1. Plants, edited by V. Velchev, 241. Sofia: Publishing House of the Bulgarian Academy of Sciences.
- Peshkova, G.A. 2006. Flora of Siberia, vol. 10: Geraniaceae – Cornaceae. Enfield, New Hampshire: Science Publishers.
- Piccoli, F., and R. Gerdol. 1981. “Rice-field weed communities in Ferrara Province (northern Italy).” Aquatic Botany 10: 317–328.
- Pietsch, W. 1963. “Vegetationskundliche Studien über die Zwergbinsen- und Strandlingsgesellschaften in der Nieder- und Oberlausitz [Vegetation studies on the ephemeral and beach vegetation in the Netherlands and Oberlausitz].” Abhandlungen und Berichte des Naturkundemuseums Görlitz 38(2): 1–82.
- Pietsch, W. 1973a. “Zur Soziologie und ökologie der Zwergbinsengesellschaften Ungarns (Klasse Isoëto-Nanojuncetea Br.-Bl. et Tx. 1943). [Towards sociology and ecology of ephemeral vegetation in Hungary (class Isoëto-Nanojuncetea Br.-Bl. et Tx. 1943)].” Acta Botanica Hungarica 19(1–4): 269–288.
- Pietsch, W. 1973b. “Beitrag zur Gliederung der europäischen Zwergbinsengesellschaften (Isoëto-Nanojuncetea Br.-Bl. et Tx. 1943). [Contribution to the classification of European ephemeral vegetation (Isoëto-Nanojuncetea Br.-Bl. et Tx. 1943)].” Vegetatio 28: 401–438.
- Pignatti, S. 1957. “La vegetazione della risaie pavesi. [The vegetation of the rice paddies].” Achiva Botanica (Forli) 33: 1–68.
- Pignatti, S. 1982. Flora d'Italia. vol. 3. Bologna: Edagricole.
- Pinke, G., R. Pál, G. Király, and V. Szendrődi. 2006. “Data to the weed flora in Somogy territory (South-Western Hungary).” Botanikai Közlemények 93: 53–68.
- Popiela, A. 1996. “Zbiorowiska z klasy Isoëto-Nanojuncetea na terenie Polski Zachodniej.” Fragmenta Floristica et Geobotanica. Series Polonica 3: 289–310.
- Popiela, A., and A. Łysko. 2010. “The distribution of Elatine macropoda Guss. (Elatinaceae).” Acta Societatis Botanicorum Poloniae 79: 81–86.
- Popiela, A., and A. Łysko. 2011. “The distribution of species of the section Elatinella Seub. (Elatine L., Elatinaceae) in Europe.”, in Geobotanist and taxonomist. A volume dedicated to professor Adam Zajac on the 70th anniversary of his birth, edited by B. Zemanek, 147–159. Cracow: Institute of Botany, Jagiellonian University.
- Popiela, A., A. Łysko, A. Wieczorek, and D. Nalepka. 2011. “The distribution of Elatine hexandra Lapierre (Elatinaceae).” Acta Societatis Botanicorum Poloniae 80: 27–32.
- Popiela, A., A. Łysko, A. Wieczorek, and A.V. Molnár. 2012. “The distribution of Elatine hydropiper L.” Acta Societatis Botanicorum Poloniae 8: 137–143.
- Popiela, A., A. Łysko, and A.V. Molnár. 2013. “Recent distribution of the Euro-Siberian-sub-Mediterranean species Elatine alsinastrum L. (Elatinaceae).” Acta Botanica Croatica 72: 375–386.
- Popiela, A., A. Łysko, M. Janukowicz, and S. Rosadziński 2014. “Elatine triandra Schkuhr.”, in Polska Czerwona Księga Roślin. Paprotniki i Rośliny Kwiatowe. [Polish Red Data Book of Plants. Pteridophytes and flowering plants], edited by R. Kaźmierczakowa. K. Zarzycki, and Z. Mirek. Kraków: Instytut Ochrony Przyrody, Polska Akademia Nauk.
- Ramenskaya, M.L., and V.N. Andreeva. 1982. Manual of the higher plants of Murmansk region and Karelia. Leningrad: Nauka.
- Ranđelović, R., B. Zlatković, D. Dimitrijević, and T. Vlahović. 2010. “Phytogeographical and phytocoenological analysis of the threatened plant taxa in the flora of the Vlasina plateau (SE Serbia).” Biologica Nyssana 1: 1–7.
- Saint-Lager, J.B. 1873. Catalogue des plantes vasculaires de la flore du bassin du Rhône. Lyon: H. Georg.
- Săvulescu, T. ed. 1950. Flora of People Republic of Romania. Bucharest: Editura Academiei Republicii Populare Romîne.
- Schmidt-Mumm, U., and H.Y. Bernal. 1995. “A new species of Elatine (Elatinaceae) from the Colombian paramos in the northern Andes.” Brittonia 47: 27–30.
- Seubert, M. 1845. “Elatinarum monographia.” Nova Acta Academiae Caesareae Leopoldino-Carolinae Germanicae Naturae Curiosorum 21: 34–60.
- Shiskin, B.K. 1961. Flora of Leningrad region. vol. 3. Leningrad: Izdatielstvo Leningradskovo Universiteta.
- Shishkin, B.K. 1974. Flora of the USSR, vol. 15. Jerusalem: Israel Program for Scientific Translations.
- Slavík, B. 1990. Fytokartograficke syntezy ČSR. [Phytogeographical syntheses of the Czechoslovakia]. Vol. 2. Průhonice, Czech Republic: Botanical Institute of Czechoslovak Academy of Science.
- Šumberová, K. 2011. “Vegetace jednoletých vlhkomilnych býlin. [Vegetation annual hygrophilous herbs].”, in Vegetace České republiky. Vol. 3. Vodní a mokřadní vegetace [Vegetation of the Czech Republic, Vol. 3. Aquatic and wetland vegetation], edited by M. Chytrý, 309–346. Praha, Czechoslovakia: Academia.
- Šumberová, K., and R. Hrivnák. 2013. “Formalised classification of the annual herb vegetation of wetlands (Isoëto Nano-Juncetea class) in the Czech Republic and Slovakia (Central Europe).” Phytocoenologia 43 (1–2): 13–40.
- Takács, A., A. Schmotzer, G. Jakab, T. Deli, A. Mesterházy, G. Király, B.A. Lukács, et al. 2013. “Key environmental variables affecting the distribution of Elatine hungarica in the Pannonian Basin.” Preslia 85: 193–207.
- The IUCN Red List of Threatened Species. Version 2014.3. http://www.iucnredlist.org>., accessed on 10 March 2015.
- Tichý, L. 2002. “JUICE, software for vegetation classification.” Journal of Vegetation Science 13: 451–453.
- Tichý, L. 2005. “New similarity indices for the assignment of relevés to the vegetation units of an existing phytosociological classification.” Plant Ecology 179: 67–72.
- Tolmatschev, A.I. 1976. Flora regionis boreali-orientalis territoriae Europaeae URSS. 3. Lelingrad: Nauka.
- Tucker, G.C. 1986. “The genera of Elatinaceae in the southeastern United States.“ Journal of the Arnold Arboretum 67: 471–483.
- Tzvelev, N.N. 1996. “Elatinaceae.” In Flora of Eastern Europe. vol. 9, edited by N.N. Tzvelev, 178–180. Saint Petersburg: Mir i Sem.
- Ubrizsy, G. 1948. “A rizs hazai gyomnövényzete [La végétation des mauvaises herbes dans les cultures de riz en Hongrie] [The vegetation of weeds in rice crops in Hungary].” Acta Agrobotanica Hungarica 1: 1–43.
- Ubrizsy, G. 1961. “Unkrautvegetation der Reiskulturen in Ungarn. [Weed vegetation of rice crops in Hungary.].” Acta Botanica Academiae Scientiarum Hungariae 7: 175–220.
- Uotila, P. 2009. “Lectotypifications in Elatine (Elatinaceae) and some taxonomic remarks.” Annales Botanici Fennici 46: 90–94.
- Uotila, P. 2010. “Elatinaceae.”, in Flora Nordica, vol. 6: Thymelaceae-Apiaceae, edited by B. Jonsell, and T. Karlsson, 62–69. Stockholm: The Swedish Museum of Natural History.
- Walters, T.G. 1969 “Elatine L.“, in Flora Europaea. vol. 2, Rosaceae to Umbelliferae, edited by T.G. Tutin, V.H. Heywood, N.A. Burges, D.M. Moore, D.H. Valentine, S.M. Walters, and D.A. Webb, 295–296. Cambridge: Cambridge University Press.