
ABSTRACT
Self-incompatibility plays an important role in the fertilization of fruit species such as apricot. Apricot (Prunus armeniaca L.) shows gametophytic self-incompatibility, which is controlled by a multi-allelic S-locus. In this study, S-alleles of 77 F1 progenies derived from Paviot, which is one of the French local cultivars, and Kabaasi, one of the most important Turkish dried apricot cultivars, parents were identified by S-RNase intron regions polymerase chain reaction (PCR) amplification and DNA sequencing. The results from the S-allele PCR analysis revealed that the Paviot female parent had an ScS2 genotype and the Kabaasi male parent had S1S9 alleles. Forty-three of the F1 progenies showed self-compatibility allele (Sc) by having either ScS9 or ScS1 alleles. Thirty-four of the F1 progenies were self-incompatible by having either S2S1 or S2S9 alleles. The distributions of detected alleles in F1 progenies were determined as follows: ScS1 31.2%, S1S2 27.3%, ScS9 24.7% and S2S9 16.8%. The results from the study are relevant for the data obtained in apricot breeding programmes in the selection of crossing combinations and in the establishment of commercial orchards.
Introduction
Apricot is one of the most important fruit species due to its commercial importance. Turkey ranks first in the world in apricot production with approximately 676.000 metric tons annually, with the apricot production fluctuating from year to year due to spring frosts.[Citation1] In recent years, molecular techniques are widely used to characterize fruit tree germplasm resources. DNA molecular markers are highly effective and informative in the assessment of the genetic diversity and genetic relationships of fruit trees.[Citation2–6]
Self-incompatibility in Prunus species, including apricot, is controlled by a simple multi-allelic S-locus that rejects its own pollen and has a homomorphic, gametophytic self-incompatibility system.[Citation7] Self-incompatibility alleles allow pollen tube growth in the pistil and access to the ovaries. In cases where the same allele disputes exist on haploid pollen and diploid pistil, the growth of pollen tubes is blocked and a dispute arises due to the S-allele-specific S-RNase gene ribosomal RNA degradation system.[Citation8–11] The effects of the self-incompatibility mechanism force pollination with foreign pollens to occur; and thus the emergence of genetic diversity and an increase in heterozygosity.[Citation12]
Many European apricot cultivars are self-compatible, whereas those in Central Asia and Iran and the Caucasus, including Turkey, are self-incompatible.[Citation13] Prominent apricot cultivars grown in Turkey are located within the Iran–Caucasian eco-geographical group. The apricot cultivars of economic importance in Turkey are found to be, generally, self-incompatible.[Citation2,Citation14–16]
Self-incompatibility plays an important role in the fertilization of fruit species such as apricot. Self-incompatible apricot cultivars need suitable pollinators for fructification.[Citation17,Citation18] A breeding target is to breed self-compatible apricot cultivars having high fruit quality. Therefore, it is imperative to determine the S-alleles of apricot cultivars.[Citation19]
The self-incompatibility mechanism can be determined using conventional methods such as pollination tests and pollen tube growth tests. However, these tests are labour-intensive and time-consuming, and also influenced by environmental factors.[Citation20,Citation21] The use of molecular techniques to reveal S-alleles in genotypes has recently become the most commonly used method that gives the best results, as it allows early selection.[Citation18]
In this study, we report S-alleles of Paviot, which is one of the French local cultivars, and Kabaasi, one of the most important Turkish dried apricot cultivars, by S-RNase intron region polymerase chain reaction (PCR) amplification and DNA sequencing. In addition, the segregation of S-alleles in Paviot × Kabaasi F1 populations is also reported.
Materials and methods
Plant material
The plant materials were provided by the Apricot Research and Application Central Directorate of Agricultural Center of İnönü University. In this study, 77 F1 progenies and their parents (Paviot and Kabaasi) were used. F1 progenies were produced in a project supported by TUBITAK [project no: TOGTAG-3099] in the period 2003–2005. Leaf samples from each plant were stored at 4 °C after lyophilization.
DNA extraction
DNA isolation from leaf samples was performed using the CTAB (cetyl trimethyl ammonium bromide) protocol according to Doyle and Doyle [Citation22] with minor modifications.[Citation23] The concentration of isolated DNA was determined by comparison with λ-DNAs quantified by agarose gel electrophoresis (Thermo Electron Corporation EC135-90, Madison, WI, USA).
S-allele PCR amplification
Primer pairs developed by Romero et al. [Citation24] and Vilanova et al. [Citation25] were used as listed in in order to determine the S-alleles by PCR analysis. Each PCR reaction of 25 µL contained 75 mmol/L of Tris-HCl (pH 8.8), 20 mmol/L of (NH4)2SO4, 2 mmol/L of MgCl2, 0.1% Tween 20, 100 µmol/L of each deoxyribonucleoside triphosphate, 0.2 µmol/L of each primer, 1.0 units of Taq DNA polymerase (Thermo, Waltham, MA, USA) and 50 ng of DNA. For PCR amplification, the samples were pre-denatured at 94 °C for 3 min, followed by 35 cycles in which the samples were denatured for 45 s at 94 °C, annealed for 45 s at 54 °C and extended for 60 s at 72 °C. For the final extension step, the samples were kept at 72 °C for 10 min. The PCR products were separated by electrophoresis in a 3% agarose gel with 0.5X TBE (Tris-borate-ethylenediaminetetraacetic acid) buffer based on band sizes and were visualized under ultraviolet light (Vilber Lourmat Infinity 1100, Collégien, France) by staining with ethidium bromide.
Table 1. The primers used to determine S-alleles of apricots.
DNA sequencing
DNA sequence analysis of PCR products was performed by Sanger sequencing at Medsantek Company (İstanbul, Turkey). The S-alleles of the parents were determined by comparing the sequences using BLAST (basic local alignment search tool) with those available in the NCBI (National Center for Biotechnology Information) databases.
Data analysis
All results were analysed using the SPSS (version 15) statistical analysis package and the data are mean values with standard deviation (±SD) from three replications. Data were analysed by analysis of variance and significant differences between the groups were determined by the multiple comparison procedure according to Duncan.[Citation26] Differences were considered statistically significant at (P < 0.05).
Results and discussion
PCR amplifications using the SrcF–SrcR primer combination detected an allele of 353 bp in Paviot genotype, which corresponds to the self-compatibility Sc allele reported by Vilanova et al.[Citation25] According to Tao et al.,[Citation27] all genotypes without Sc alleles did not fructify, and they were self-incompatible. In addition to the Sc allele, three other alleles were identified: two in Paviot (328 and 267 bp) and one in Kabaasi (373 bp) ().
Figure 1. Electrophoregram of S-alleles of parents and F1 progenies amplified with the SrcF and SrcR primers for the first intron region. P: Paviot, K: Kabaaşı. DNA molecular size marker: Thermo Generuler 50 bp DNA ladder (Waltham, MA, USA).
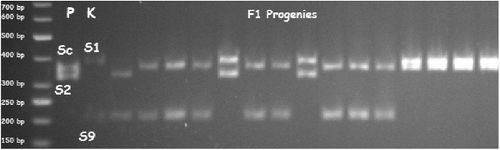
The Paviot and Kabaasi S-allele genotypes showed homology with the allele sequences from Prunus armeniaca with Sc (353 bp), S2 (328 bp), S1 (373 bp) and S9 (267 bp) available in the NCBI database.[Citation23,Citation28] The Paviot genotype was found to have ScS2 alleles and to be self-compatible due to the presence of Sc alleles. It was reported that the Kabaasi genotype has the S9 allele, and it is the most common allele found in the local Turkish apricot varieties such as Adilcevaz 5, Akcadag Gunay, Cataloglu, Cekirge 52, Cologlu, Dortyol 2, Haci Haliloglu, Hasanbey, Ismailaga, Kadıoglu, Kamelya, Kurukabuk, No 2 Zerdali, Ozal, Seftalioglu, Soganci and X3 Zerdali.[Citation2]
Previous selfing studies carried out under specific conditions showed that the Kabaasi cultivar is self-compatible,[Citation16] whereas others have found it to be self-incompatible.[Citation14,Citation15] In this study, the Kabaasi cultivar had S1S9 alleles and was determined to be self-incompatible. Halasz et al. [Citation2] reported that the Kabaasi cultivar had S9S13 alleles, and it was self-incompatible. In that study, the alleles carried by individuals were identified based on the band sizes in electrophoresis gel images. The band sizes were estimated by separating the PCR products by agarose gel electrophoresis; however, it was sometimes very difficult to estimate such bands when the sizes of the alleles were very close to each other. In this study, to overcome this problem, the bands in Kabaasi and Paviot cultivars were bidirectionally sequenced, BLAST searched in NCBI GenBank and in them, the S-alleles were determined as S1S9.
Apricots belonging to the Iran–Caucasian eco-geographical group have been reported to be mostly self-incompatible, those in the European eco-geographical group, mostly self-compatible and 60% of the varieties in Turkey have been found to be self-incompatible.[Citation16,Citation29–31] In this study, the Kabaasi cultivar, which is in the Iran–Caucasian eco-geographical group, was found to be self-incompatible, whereas the Paviot cultivar, which belongs to the European eco-geographical group, was found self-compatible.
Of the 77 F1 progenies tested, 34 samples did not have Sc alleles and were identified to be self-incompatible (). According to Halasz et al.,[Citation2] there was absence of an Sc allele in 44 of 51 apricot genotypes grown in Turkey and they were found to be self-incompatible. Halasz et al. [Citation31] also found that 60% of the varieties grown in Turkey are self-incompatible. In this study, 43 of the F1 progenies had either ScS1 or ScS9 alleles, and they were self-compatible. On the other hand, 34 F1 progenies were found to have S1S2 and S2S9 incompatibility alleles (). Burgos et al. [Citation10] reported that the self-compatibility allele was dominant over the self-incompatibility allele. Similarly, in their study conducted on the self-incompatibility status of apricot F1 populations in, Chen et al. [Citation32] found that the self-compatibility allele was dominant over the self-incompatibility allele; and the S-genotype in Katy, the main individual, was heterozygous (ScS8).
Table 2. S-genotypes of parents and F1 progenies in this study.
Table 3. Fruit characteristics of some F1 progenies.
The distribution of alleles in the F1 progenies was observed to be as follows: ScS1 in 31.2%, S1S2 in 27.3%, ScS9 in 24.7%, and S2S9 in 16.8%. Two alleles were identified in the Paviot genotype with sizes of 353 and 328 bp, and two other alleles were found in the Kabaasi genotype with sizes of 267 and 373 bp. Comparison of the DNA sequences of all the four different alleles with those in GenBank showed that the Sc and S2 alleles were found in the Paviot genotype, whereas S1 and S9 were identified in the Kabaasi genotype. In total, 43 F1 progenies were found to have an Sc allele and thus, to be self-compatible, whereas 34 F1 progenies were self-incompatible due to the absence of an Sc allele.
shows the pomological features of some F1 progenies that had high quality in 2011–2013. F1 progenies Nos. 17, 20, 58, 67 and 72 were found to have high yield and high quality fruit in pomological studies. At the same time, they were determined to be self-compatible, too (). These progenies can be used as parents in breeding programmes or as cultivar candidates in commercial apricot orchards.
Conclusions
In this study, S-allele specific PCR was used to identify the S-alleles of 77 F1 progenies and their parents. The DNA sequences of four alleles were obtained and compared in the NCBI GenBank database. Sc and S2 alleles were found in the Paviot genotype, whereas S1 and S9 alleles were identified in the Kabaasi genotype. It was determined that 43 F1 progenies had the Sc allele; and thus they were self-compatible. On the other hand, 34 F1 progenies had no Sc allele; therefore, they were self-incompatible. The distributions of the detected alleles in the F1 progenies in Paviot (ScS2) and Kabaasi (S1S9) parents were determined as follows: ScS1 31.2%, S1S2 27.3%, ScS9 24.7% and S2S9 16.8%. In breeding studies, the development of new genotypes with the desired characteristics and with known compatibility pattern of parents is of crucial importance in terms of both the required time and the cost of manpower. Therefore, the results from this study are relevant based on the data obtained in apricot breeding programmes in the selection of crossing combinations and in the establishment of commercial orchards.
Acknowledgments
The authors thank Lorenzo Burgos, Nuria Alburquerque and Lydia Bremaud from Centro de Edafologia y Biologia Aplicada del Segura (CEBAS-CSIC), Murcia, Spain, for providing valuable help, support and advice.
Disclosure statement
No potential conflict of interest was reported by the authors.
Additional information
Funding
References
- Asma BM. Every aspect of apricot. 1st ed. Istanbul: Uyum Press; 2011.
- Halasz J, Pedryc A, Ercisli S, et al. S-genotyping supports the genetic relationships between Turkish and Hungarian apricot germplasm. J Amer Soc Hort Sci. 2010;135(5):410–417.
- Kaya T, Balta F, Şensoy S. Fruit quality parameters and molecular analysis of apple germplasm resources from Van Lake Basin, Turkey. Turk J Agric For. 2015;39:864–875.
- Motalebipour EZ, Kafkas S, Ozongun Ş, et al. Construction of dense genetic linkage maps of apple cultivars Kaşel-41 and Williams? Pride by simple sequence repeat markers. Turk J Agric For. 2015;39:967–975.
- Bajpai PK, Warghat AR, Sharma RK, et al. Structure and genetic diversity of natural populations of Morus alba in the Trans-Himalayan Ladakh Region. Biochem Genet. 2014;52:137–152.
- Feng SG, Lu JJ, Gao L, et al. Molecular phylogeny analysis and species identification of Dendrobium (Orchidaceae) in China. Biochem Genet. 2014;52:127–136.
- McCubbin AG, Kao TH. Molecular recognition and response in pollen and pistil interactions. Annu Rev Cell Dev Bio. 2000;16:333–364.
- McClure BA, Ebert PR, Anderson MA, et al. Style self-incompatibility gene products of Nicotiana alata are ribonucleases. Nature. 1989;342:955–957.
- McClure BA, Gray JE, Anderson MA, et al. Self-incompatibility in Nicotiana alata involves degradation of pollen rRNA. Nature. 1990;347:757–760.
- Burgos L, Egea J, Guerriero R, et al. The self-compatibility trait of the main apricot cultivars and new selections from breeding programmes. J Hort Sci Biotech. 1997;72:147–154.
- Roalson EH, McCubbin AG. S-RNases and sexual incompatibility: structure, functions, and evolutionary perspectives. Mol Phylogenet Evol. 2003;29:490–506.
- Donoso JM, Aros D, Meneses C, et al. Identification of S-alleles associated with self-incompatibility in apricots (Prunus armeniaca L.) using molecular markers. J Food Agric Environ. 2009;7(3–4):270–273.
- Kostina LD. Self-fertility studies in apricot. Trud Gos Nikit Botan Sada. 1970;45:7–17. Russian.
- Gulcan R, Misirli A, Saglam H, et al. Properties of Turkish apricot land races. Acta Hort. 2006;701:191–198.
- Misirli A, Saglam H, Gulcan R, et al. Investigation on fertilization biology of important dried apricot cultivars. Acta Hort. 2006;701(1):159–162.
- Paydas S, Eti S, Gulcan R, et al. In vitro investigations on pollen quality, production and self-incompatibility of some apricots varieties in Malatya, Turkey. Acta Hort. 2006;701:75–80.
- Tehrani G, Browns SK. Pollen-incompatibility and self-fertility in sweet cherry. In: Janick J, editor. Plant breeding reviews. Vol. 9. Oxford: Wiley; 1992. p. 367–388.
- Yaegaki H, Shimada T, Moriguchi H, et al. Molecular characterization of S-RNase genes and S-genotypes in the Japanese apricot (Prunus mume Sieb.et Zucc.). Sex Plant Reprod. 2001;3:251–257.
- Chen XS, Li XL, Zhang YM, et al. Advances in apricot germplasm resources evaluation and genetic breeding. J Fruit Sci. 2001;18:178–181.
- Zhang SL, Cao SM, Wu HQ. Self-incompatibility genotypes of fruit trees and their identification methods. J Fruit Sci. 2003;20:358–363.
- Halász J, Hegedûs A. Self-incompatibility in pears (Pyrus communis L., Pyrus serotine Rehd. and Pyrusus suriensis). Rev Int J Hort Sci. 2006;12:87–91.
- Doyle JJ, Doyle JL. A rapid isolation procedure for small quantities of fresh leaf tissue. Phytochem Bull. 1987;19:11–15.
- Kafkas S, Perl-Treves R. Morphological and molecular phylogeny of Pistacia species in Turkey. Theor Appl Genet. 2001;102:908–915.
- Romero C, Vilanova S, Burgos L, et al. Analysis of the S locus structure in Prunus armeniaca L. identification of S-haplotype S-RNase and F-box genes. Plant Mol Biol. 2004;56:145–157.
- Vilanova S, Romero C, Llacer G, et al. Identification of self-incompatibility alleles in apricot by PCR and sequence analysis. J Am Soc Hort Sci. 2002;130:893–898.
- Duncan DB. Multiple range and multiple F tests. Biometrics. 1955;11:1–42.
- Tao R, Habu T, Namba A, et al. Inheritance of Sf-RNase in Japanese apricot (Prunus mume) and its relation to self-compatibility. Theor Appl Genet. 2002;105:222–228.
- Vilanova S, Badenes ML, Burgos L, et al. Self-compatibility of two apricot selections is associated with two pollen-part mutations of different nature. Plant Physiol. 2006;142:629–641.
- Mehlenbacher SA, Cociu V, Hough LF. Apricots, genetic resources of temperate fruit and nut crops. Int Soc Hort Sci. 1990;3:65–107.
- Halasz J, Hegedus A, Herman R, et al. New self-incompatibility alleles in apricot (Prunus armeniaca L.) revealed by stylar ribonuclease assay and S-PCR analysis. Euphytica. 2005;145:57–66.
- Halasz J, Pedryc A, Hegedus A. Origin and dissemination of the pollen-part mutated SC-haplotype that confers self-compatibility in apricot (Prunus armeniaca). New Phytol. 2007;176:793–803.
- Chen X, Wu Y, Chen M, et al. Inheritance and correlation of self-compatibility and other yield components in the apricot hybrid F1 populations. Euphytica. 2006;150:69–74.