Abstract
This study aimed to characterize the physiological and morphological traits that are associated with adaptation to unflooded soil conditions in rice. Four indica rice cultivars (Puluik Arang, Badari Dhan, Shwe Nang Gyi, and Ratul), which were previously identified as highly or less adaptable to unflooded soil conditions, were grown under flooded and unflooded (soil water potential; -0.10 MPa) soil conditions. Water uptake was measured every day for three weeks, and then the leaf water potential, the stomatal conductance, the dry matter weight, shoot and root morphological traits were measured. Puluik Arang and Badari Dhan exhibited greater leaf area expansion and higher maintenance of root development under the unflooded condition than that by other cultivars. The leaf water potential and stomatal conductance of fully expanded highest leaf in Puluik Arang and Badari Dhan were not affected by unflooded soil regime. Leaf area and root morphological traits were significantly correlated with water uptake regardless of soil moisture regimes. These results suggested that Puluik Arang and Badari Dhan exhibited great water uptake capacity through physiological and morphological adaptation of shoot and root traits to unflooded condition, resulting in great biomass productivity under the condition.
The global population is increasing rapidly; therefore, stable production and a reliable food supply are major concerns. The competition for fresh water by agricultural and urban/industrial applications threatens the sustainability of food production (Bouman et al., Citation2006; Peng et al., Citation2009). Rice can adapt to a wide range of hydrological conditions; however, its productivity under water deficit is low compared to that in irrigated lowland ecosystems (Maclean et al., Citation2002). To improve yield potential under water deficit, many studies have investigated water saving technology, and/or genetic improvement for drought resistance (Serraj et al., Citation2011; Tuong et al., Citation2005; Zhang et al., Citation2009).
The identification of the physiological traits responsible for greater biomass or yield production may facilitate genetic improvements in productivity in rice grown in water limited conditions. Because water uptake is strongly linked to biomass production, improved water uptake capacity is one of the most important targets for yield improvement under water deficit conditions (Blum, Citation2009; Kobata et al., Citation1996; Nguyen et al., Citation1997). Water uptake is regulated by balancing of water uptake through the root system and transpiration by shoot (Cosgrove & Holbrook, Citation2010). Leaf expansion and stomatal behavior strongly regulate the amount of transpiration and photosynthetic production through radiation capture and gas exchange (Katsura et al., Citation2010; Maruyama & Tajima, Citation1990). The regulation of transpiration from shoot have been well investigated, but relatively little is known about the mechanisms of water uptake by root system although the contribution of root system to plant water balance has been recognized (Kamoshita, Citation2011).
We previously examined the genotypic variation in biomass production under different soil moisture conditions using a rice diversity research set of germplasm (RDRS) developed by the National Institute of Agrobiological Science (NIAS) (Matsunami et al., Citation2012). The RDRS comprises 69 cultivars and harbors records of 91% of the alleles identified in the representative 332 accessions selected from a global collection of rice cultivars (Kojima et al., Citation2005). The RDRS responded in different ways to soil moisture conditions, i.e. half of the cultivars had higher biomass production under flooded condition, whereas others maintained or increased their biomass production under unflooded but wet soil moisture conditions. The results suggested that cultivars were adapted either to flooded or unflooded soil moisture conditions, although the details of the morphological and physiological traits that underlie the variable adaptability to soil moisture were not elucidated in our previous study.
Water uptake under water limited conditions is thought to be due to plant morphological development (e.g. leaf expansion, root elongation) and physiological traits (e.g. stomatal behavior, hydraulic conductivity). In this study, therefore, genotypic variation of water uptake under unflooded condition was investigated in terms of shoot and root morphological development and physiological traits, such as stomatal conductance and leaf water potential (LWP). It was hypothesized that a combination of morphological and physiological traits of shoot and root would explain genotypic difference in water uptake capacity under unflooded condition.
Materials and Methods
1. Materials
We used four indica rice (Oryza sativa L.) cultivars selected from the NIAS global rice core collection (Kojima et al., Citation2005): Puluik Arang, Badari Dhan, Shwe Nang Gyi, and Ratul. The four cultivars had comparable biomass production under a flooded regime, but different responses to unflooded condition; Puluik Arang and Badari Dhan had greater biomass production under unflooded regime than under flooded condition, whereas biomass production decreased in Shwe Nang Gyi and Ratul under unflooded regime (Matsunami et al., Citation2012).
2. Culture details and soil moisture regimes
Seeds were sown into a seedling tray, and grown in a growth chamber under a 12 h light/12 h dark photoperiod (450 μmol s−1 m−2 photosynthetic photon flux density during the light period), with day/night temperatures of 25/20 °C and a relative humidity of 70%. Two-week-old seedlings were transplanted to 1/10,000-a Wagner pots (127 φmm, 198 mm height; ICM, Japan), with one plant per pot. After transplanting, the plants were subjected to flooded condition (43% [w/w], soil water potentials of -0.02 MPa), and unflooded condition (33%, -0.10 MPa). The reduction in the weight of each pot was measured every day, and the soil moisture level was maintained by adding water. Water uptake was estimated from the daily reduction in pot weight, minus the amount of evaporation from the soil surface, which was determined by measuring the weight change in the pot without a plant. The soil moisture regimes started after transplanting and continued for three weeks. Further details and soil properties were described previously (Matsunami et al., Citation2012).
3. Measurements
At 19 days after treatment, the stomatal conductance of the fully expanded highest leaf of the main stem was measured using a leaf porometer (SC-1 Leaf Porometer, DECAGON., U.S.A.). Measurements were conducted at 3 h after lights on.The LWP of the fully expanded highest leaf was determined 3 h after lights on at 20 days after treatment. The leaf disks were sampled and immediately placed in a C-52-SF sample chamber (WESCOR, U. S. A.), and the samples were allowed to equilibrate in the chamber. Then the chambers were connected to the dew point microvoltmeter (HR-33T, WESCOR, U.S.A.) to measure the LWP.
Shoot and root biomass production and the morphological traits were evaluated by harvesting all of the tissues at three weeks after treatment. The leaf area was measured with an automatic area meter (AAM-9, Hayashidenko, Japan), and the shoot samples were then dried at 80 °C for more than three days and weighed. Root samples were preserved in FAA solution (50% ethanol: acetic acid: formalin = 8:1:1) and stained with 0.1% (w/w) Coomassie Brilliant Blue. The root segments were scanned with an image scanner (GT-9800, Epson, Japan) at 400 dpi, 8-bit grayscale. To determine root length, we used the image analysis system “Root Length 1.8 win” (Kimura et al., Citation1999). After investigating the root morphological traits, the root samples were dried at 80 °C for more than three days and weighed.
Statistical analyses were conducted to determine the effects of the cultivar/water regimes on the traits using JMP 8 Statistical Discovery software (SAS Institute, U.S.A).
Results
1. Shoot and root biomass production and morphological development
Table shows the shoot dry weight (DW) and morphological traits of the four rice cultivars grown under flooded and unflooded soil conditions. There were no significant differences in the shoot DW among cultivars under the flooded condition (2.28 – 2.73 g plant−1). The shoot DW under the unflooded condition was significantly heavier in Puluik Arang (3.09 g plant−1) than Shwe Nang Gyi (2.32 g plant−1) and Ratul (1.89 g plant−1). The responses of shoot DW to flooded and unflooded conditions were differed by cultivars; Puluik Arang and Badari Dhan had 13%–18% heavier shoot DW under the unflooded condition than the flooded condition, whereas Shwe Nang Gyi and Ratul had lighter shoot DW under the unflooded condition. The tiller number in Ratul (9.7 tillers plant−1) under the flooded condition was largest, and Puluik Arang (6.7 tillers plant−1) and Badari Dhan (7.3 tillers plant−1) showed significantly fewer number of tillers compared with that of Ratul. On the other hand, unflooded condition enhanced tiller development in Puluik Arang and Badari Dhan; the two cultivars had 18%–23% more number of tillers under the unflooded condition than the flooded condition. The tiller number under unflooded condition in Ratul (6.7 tillers plant−1) was significantly decreased under the unflooded condition than the flooded condition, and the number was smallest among the cultivars. The leaf age was similar among cultivars under the flooded condition. The leaf age in Puluik Arang and Badari Dhan was not affected by soil moisture regime, whereas the leaf age in Shwe Nang Gyi and Ratul delayed with soil moisture deficit. There were no significant differences in the leaf area among the cultivars under the flooded condition (341–429 cm2 plant−1). The leaf area under the unflooded condition was significantly larger in Puluik Arang (497 cm2 plant−1) than Shwe Nang Gyi (396 cm2 plant−1) and Ratul (321 cm2 plant−1). The responses of leaf area to flooded and unflooded conditions were differed with cultivars; Puluik Arang and Badari Dhan had 29%–34% larger leaf area under the unflooded condition than the flooded condition, whereas Shwe Nang Gyi and Ratul had smaller leaf area under the unflooded condition.
Table 1. Shoot dry weight and shoot morphological traits of four rice cultivars subjected to flooded and unflooded conditions.
Response of root DW to soil moisture conditions was similar to shoot DW (Table ). There were no significant differences in the root DW among the cultivars under the flooded condition (0.38–0.44 g plant−1). Under the unflooded condition, Puluik Arang exhibited heaviest root DW (0.49 g plant−1), and it was significantly heavier than the root DW in Ratul (0.33 g plant−1). Puluik Arang and Badari Dhan had about 10% heavier root DW under the unflooded condition than the flooded condition. Root DW of Shwe Nang Gyi was not affected under soil moisture condition. Ratul showed 20% less root DW under the unflooded condition than that under the flooded. The crown root number under the flooded condition were similar among Puluik Arang, Shwe Nang Gy, and Ratul (80–84 roots plant−1), but Badari Dhan (68 root plant−1) had fewer number of crown root although the difference was not significant. The crown root number was restricted in all cultivars with soil moisture deficit, especially Shwe Nang Gyi and Ratul had significantly fewer root under the unflooded condition than flooded condition. There were no significant cultivar difference in the total root length and root surface area under the flooded conditions; the root length and root surface area were 242–306 m plant−1and 1,115–1,343 cm2 plant−1, respectively. The root length was significantly shorter under the unflooded condition than the flooded condition, regardless of the cultivar (116–193 m plant−1). The root surface area were also restricted with unflooded regime although the degree of reduction were varied among the cultivars; Puluik Arang and Badari Dhan had 18–20%, Shwe Nang Gyi had 33% and Ratul had 44% less surface area compared with that under the flooded condition. Puluik Arang (1,076 cm2 plant−1) exhibited significantly larger root surface area than that in Ratul (622 cm2 plant−1) under the unflooded condition.
Table 2. Root dry weight and root morphological traits of four rice cultivars subjected to flooded and unflooded conditions.
2. Leaf water potential and stomatal conductance
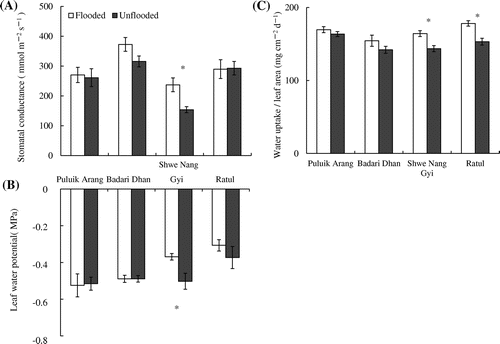
The stomatal conductance of fully expanded highest leaf were varied among cultivars; the stomatal conductance was highest in Badari Dhan, lowest in Shwe Nang Gyi and Puluik Arang and Ratul were intermediate (Figure A). The stomatal conductance in Shwe Nang Gyi was significantly decreased by unflooded regime. Similar to stomatal conductance, the LWP of Shwe Nang Gyi was significantly lowered under the unflooded condition (Figure B). The daily amount of water uptake per unit leaf area was significantly decreased by unflooded regime in Shwe Nang Gyi and Ratul, whereas the amount of water uptake per leaf area were not affected by soil moisture regime in Puluik Arang and Badari Dhan (Figure C).
Figure 1. Stomatal conductance (A), LWP (B) of the fully expanded highest leaf and water uptake per unit leaf area (C) grown under different soil moisture regimes. Water uptake / leaf area were calculated dividing the daily amount of water uptake by the leaf area measured at 3 weeks after treatment (n = 6). The stomatal conductance (n = 5) and the LWP (n = 3) were measured of the fully expanded highest leaf, using a different plant. Vertical bars indicate the standard error. * indicate significant difference between flooded and unflooded conditions at p < 0.05 according to t-test.
3. Water uptake and its relationship with morphological traits
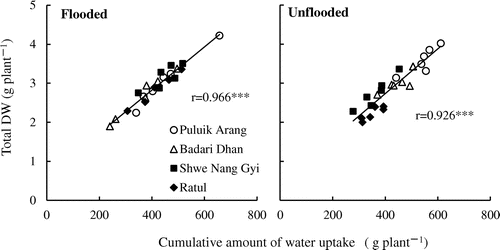
The cumulative amount of water uptake during treatment was comparable between the cultivars under the flooded regime (Figure ). The interactions between water regime and cultivar (W × C) were significant for the amount of water uptake; under unflooded condition, Puluik Arang and Badari Dhan absorbed more water than did under the flooded condition, whereas Shwe Nang Gyi and Ratul absorbed less water compared with that under the flooded condition. The cumulative amount of water uptake was strongly correlated with the biomass production under both soil moisture regime; the correlation coefficients were 0.966 under the flooded and 0.926 under the unflooded condition (Figure ).
Figure 2. Cumulative amount water uptake of four rice cultivars under flooded and unflooded conditions. Vertical bars indicate the standard error of six replicates of the amount of water-uptake from different plants. Bars followed by different letters are significantly different according to Tukey’s test at p < 0.05. Levels of significance: ** significant at p < 0.01, n.s. = not significant.
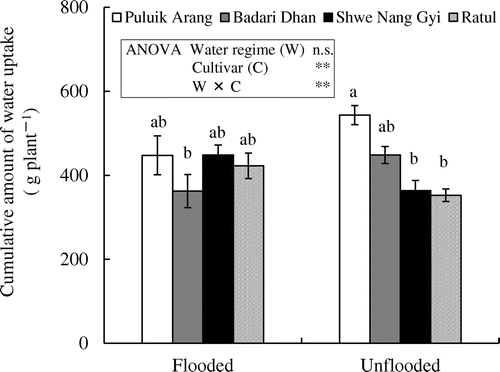
Figure3. Relationships between the cumulative amount of water uptake and total DW among four rice cultivars. Diagonal lines indicate regressions for all values of each soil water regimes. Levels of significance: *** significant at p < 0.001.
Leaf area, root length, and root surface area were significantly correlated with the daily amount of water uptake at 3 weeks after treatment (Table ). The correlation coefficient between water uptake and leaf area were higher than that between root morphological traits regardless of water regime. The correlation coefficient between water uptake and root morphological traits was higher under the unflooded condition than that under the flooded condition.
Table 3. Correlation coefficients between the daily amount of water uptake and shoot and root morphological traits.
4. Discussion
We assessed the effects of soil moisture on water uptake and the physiological and morphological traits of rice cultivars adapted to flooded or unflooded conditions. Water uptake reflected cultivar biomass production under different soil moisture regimes; e.g. Puluik Arang, which exhibited the greatest water uptake capacity under unflooded condition showed greater biomass production under the unflooded condition (Table , Figures and ). Therefore, the cultivars that had a greater water uptake capacity also exhibited higher biomass production, and there existed genotypic variation in the adaptation to soil moisture conditions.
Water uptake consists of two components, passive water uptake and active water uptake; the former, which consist mostly amount of water uptake, is regulated by transpiration from leaf (Cosgrove and Holbrook, Citation2010). Therefore, leaf expansion and physiological traits are important to discuss about water uptake ability. In regard to the leaf expansion, Puluik Arang and Badari Dhan, which increased or maintained tiller emergence and leaf age under unflooded condition, had about 30% larger leaf area under the unflooded condition than under the flooded (Table ). The LWP and stomatal conductance of Puluik Arang and Badari Dhan were not affected under soil moisture conditions (Figure ). The LWP and stomatal conductance of Ratul were also comparable between the two soil moisture regimes. The maintenance of LWP and stomatal conductance in Ratul was probably due to the reduction in aboveground plant size under the unflooded condition. The reduction of LWP and stomatal conductance in Shwe Nang Gyi were largest compared with the other cultivars, suggesting that Shwe Nang Gyi could not uptake adequate amount of water to maintain the leaf water status. Although we did not measure the transpiration flux at steady state, it is estimated that Puluik Arang and Badari Dhan had smaller whole plant hydraulic resistance (R) under unflooded condition than that under the flooded according to the following equation: R = (Ψsoil − Ψleaf)/T, where T is the transpiration (≒water uptake) per unit leaf area, Ψsoil, the soil water potential and Ψleaf, the LWP (Hirasawa & Ishihara, Citation1991). The maintenance of LWP and water uptake per leaf area in Puluik Arang and Badari Dhan under unflooded condition might be due to the adjustment of hydraulic conductivity.
The positive and strong correlation between water uptake and leaf area were observed regardless of soil moisture regime (Table ), thus, the leaf area expansion is linked to the genotypic variation in water uptake under the unflooded condition in this study. Stable and rapid leaf expansion is also reported as an important trait for rice grown under water limited conditions in terms of radiation capturing, weed competitiveness, and nitrogen uptake ability (Dingkuhn et al., Citation1999; Katsura et al., Citation2010, Okami et al., Citation2011).
On the other hand, under environmentally stressed condition around rhizosphere, water uptake decreases if the water uptake by root cannot keep up with the transpiration demand. Genotypic difference in water uptake is due in part to root architecture and root hydraulic conductivity. In terms of root architecture, deep rooting, root angle, and plasticity in root development are well recognized as important traits for enhancing rice productivity under water limited conditions (Araki & Iijima, Citation1998; Kano-Nakata et al., Citation2011; Uga et al., Citation2013). Recent studies have revealed that aquaporin influences root hydraulic conductivity; ex. when roots are subjected to low temperature or treated with respiratory inhibitor, root hydraulic conductivity is decreased due to reduction of aquaporin activity (Murai-Hatano et al., Citation2008; Tournaire-Roux et al., Citation2003). These investigations showed the importance of genetic improvement in root function for better water uptake under stressed conditions. In this study, root morphological traits, such as crown root number, root length, and root surface area, were restricted under the unflooded condition compared with that under the flooded condition, regardless of the cultivar. Similar results were reported in other studies, where the elongation of total root length was strongly regulated under the soil moisture condition and was generally restricted, even under mild water deficit or saturated soil conditions (Kano-Nakata et al., Citation2011; Kato & Okami, Citation2010). When compared among cultivars, Puluik Arang and Badari Dhan, which exhibited great water uptake under the unflooded, maintained the number of crown root, root length, and root surface area at levels higher than that by Shwe Nang Gyi and Ratul under the unflooded condition. Although the correlation coefficients were higher in the relation between leaf area and water uptake, there were also significant relationship between root morphological traits and water uptake (Table ). Especially under the unflooded condition, the correlation between root morphological traits and water uptake were stronger than those under the flooded condition. Therefore, the root development contributed to biomass productivity through enhanced water uptake under unflooded condition. It is needed to investigate whether the cultivars with adaptation to unflooded condition possessed greater root function such as water uptake capacity per unit root length and/or root hydraulic conductivity.
In conclusion, the two cultivars Puluik Arang and Badari Dhan, which increased biomass productivity under unflooded condition, had larger number of tillers under the unflooded condition compared with that under the flooded regime, resulted in a wider leaf area expansion. In terms of root traits, Puluik Arang and Badari Dhan maintained root length and surface area at higher levels compared with Shwe Nang Gyi and Ratul. Therefore, genotypic difference in water uptake capacity was due in part to vigor leaf expansion and root development under unflooded condition. In addition, the great water uptake capacity in Puluik Arang and Badari Dhan under unflooded condition would contribute to the maintenance of leaf water states, such as stomatal conductance and LWP. These results suggested that the cultivars with adaptation to unflooded condition possessed favorable physiological and morphological traits of shoot and root which were associated with water uptake ability and thus contributed great biomass productivity under unflooded condition.
In this study, we found that Puluik Arang exhibited greatest water uptake under unflooded condition, and possessed great physiological and morphological traits that associated with water uptake. Thus, Puluik Arang has the possibility to be a good material for the investigation to understand the rice adaptability to mild soil moisture deficit. Further investigations such as identification of physiological and morphological plasticity responding to soil moisture conditions and the balance of shoot and root development for better water uptake and water use efficiency are necessary.
Disclosure statement
No potential conflict of interest was reported by the authors.
Funding
This research was financially supported in part by a Grant-in-Aid for the JSPS (the Japanese Society for the Promotion of Science) fellowship awarded to M. M.
Acknowledgments
Special thanks go to the late Dr. S. Masaki (Akita Prefectural Agricultural Experiment Station) for his helpful advice on the implementation of this study.
References
- Araki, H., & Iijima, M. (1998). Rooting nodes of deep roots in rice and maize grown in a long tube. Plant Production Science, 1, 242–247. 10.1626/pps.1.242
- Blum, A. (2009). Effective use of water (EUW) and not water use efficiency (WUE) is the target of crop yield improvement under drought stress. Field Crops Research, 112, 119–123. 10.1016/j.fcr.2009.03.009
- Bouman, B. A. M., Humphreys, E., Tuong, T. P., & Barker, R. (2006). Rice and water. Advances Agronomie, 92, 187–237.
- Cosgrove, D., & Holbrook, M. (2010). Water balance of plants. In L. Taiz & E. Zeiger (Eds.), Plant physiology (5th ed., pp. 85–105). Massachusetts, Sinauer Associate.
- Dingkuhn, M., Johnson, D. E., Sow, A., & Audebert, A. Y. (1999). Relationships between upland rice canopy characteristics and weed competitiveness. Field Crops Research, 61, 79–95. 10.1016/S0378-4290(98)00152-X
- Hirasawa, T., & Ishihara, K. (1991). On resistance to water transport in crop plants for estimating water uptake ability under intense transpiration. Japanese journal of crop science, 60, 174–183. 10.1626/jcs.60.174
- Kamoshita, A. 2011. Current status of research on improvement of drought resistance in rice (Oryza sativa L.). Japanese Journal of Crop Science, 80, 1-12.10.1626/jcs.80.1
- Kano-Nakata, M., Inukai, Y., Wade, L. J., Siopongco, J. D. L. C., & Yamauchi, A. (2011). Root development, water uptake, and shoot dry matter production under water deficit conditions in two CSSLs of rice: Functional roles of root plasticity. Plant Production Science, 14, 307–317. 10.1626/pps.14.307
- Kato, Y., & Okami, M. (2010). Root growth dynamics and stomatal behaviour of rice (Oryza sativa L.) grown under aerobic and flooded conditions. Field Crops Research, 117, 9–17.10.1016/j.fcr.2009.12.003
- Katsura, K., Okami, M., Mizunuma, H., & Kato, Y. (2010). Radiation use efficiency, N accumulation and biomass production of high-yielding rice in aerobic culture. Field Crops Research, 117, 81–89.10.1016/j.fcr.2010.02.006
- Kimura, K., Kikuchi, S., & Yamasaki, S. (1999). Accurate root length measurement by image analysis. Plant Soil, 216, 117–127. 10.1023/A:1004778925316
- Kobata T., Okuno T., & Yamamoto T. (1996). Contributions of capacity for soil water extraction and water use efficiency to maintenance of dry matter production in rice subjected to drought. Japanese of Journal of Crop Science, 64, 652-662.
- Kojima, Y., Ebana, K., Fukuoka, S., Nagamine, T., & Kawase, M. (2005). Development of an RFLP-based rice diversity research set of Germplasm. Breeding Science, 55, 431–440. 10.1270/jsbbs.55.431
- Maclean, J. L., Dawe, D. C., Hardy, B., & Hettel, G. P. (2002). Rice Almanac (3rd ed.) pp. 1–253. IRRI, IRRIWARDA, CIAT, Manila, FAO Press.
- Matsunami, M., Matsunami, T., Ogawa, A., Toyofuku, K., Kodama, I., & Kokubun, M. (2012). Genotypic variation in biomass production at the early vegetative stage among rice cultivars subjected to deficient soil moisture regimes and its association with water uptake capacity. Plant Production Science, 15, 82–91. 10.1626/pps.15.82
- Murai-Hatano, M., Kuwagata, T., Sakurai, J., Nonami, H., Ahamed, A., Nagasuga, K., … Okada, M. (2008). Effect of low root temperature on hydraulic conductivity of rice plants and the possible role of aquaporins. Plant and Cell Physiology, 49, 1294–1305. 10.1093/pcp/pcn104
- Maruyama, S., & Tajima, K. (1990). Leaf conductance in japonica and indica rice varieties. I. Size, frequency and aperture of stomata. Japanese journal of crop science, 59, 801–808. 10.1626/jcs.59.801
- Nguyen, H. T., Babu, R. C., & Blum, A. (1997). Breeding for drought resistance in rice: Physiology and molecular genetics considerations. Crop Science, 37, 1426–1434. 10.2135/cropsci1997.0011183X003700050002x
- Okami, M., Kato, Y., & Yamagishi, J. (2011). Role of early vigor in adaptation of rice to water-saving aerobic culture: Effects of nitrogen utilization and leaf growth. Field Crops Research, 124, 124–131. 10.1016/j.fcr.2011.06.013
- Peng, S., Tang, Q., & Zou, Y. (2009). Current status and challenges of rice production in China. Plant Production Science, 12, 3–8. 10.1626/pps.12.3
- Serraj, R., McNally, K.L., Slamet-Loedin, I., Kohli, A., Haefele, S.M., Atlin, G., & Kumar, A. (2011). Drought resistance improvement in rice: An integrated genetic and resource management strategy. Plant Production Science, 14, 47–55.
- Tournaire-Roux, C., Sutka, M., Javot, H., Gout, E., Gerbeau, P., Luu, D.T., … Maurel, C. (2003). Cytosolic pH regulates root water transport during anoxic stress through gating of aquaporins. Nature, 425, 393–397. 10.1038/nature01853
- Tuong, T. P., Bouman, B. A. M., & Mortimer, M. (2005). More rice, less water-integrated approaches for increasing water productivity in irrigated rice-based systems in Asia. Plant Production Science, 8, 231–241. 10.1626/pps.8.231
- Uga, Y., Sugimoto, K., Ogawa, S., Rane, J., Ishitani, M., Hara, N., … Yano, M. (2013). Control of root system architecture by DEEPER ROOTING 1 increases rice yield under drought conditions. Nature Genetics, 45, 1097–1102. 10.1038/ng.2725
- Zhang, H., Xue, Y., Wang, Z., Yang, J., & Zhang, J. (2009). An alternate wetting and moderate soil drying regime improves root and shoot growth in rice. Crop Science, 49, 2246–2260. 10.2135/cropsci2009.02.0099