Abstract
In relay intercropping systems, late-planted crops often grow under the shade of the canopy of early-planted tall crops and then transfer to full sunlight after the harvest of the early-planted crops. In order to know the effects of recovery growth of the late-planted soya bean in maize–soya bean relay intercropping, a field experiment was carried out to observe architectural, morphological, physiological and anatomical traits of soya bean plants related to shade and subsequent removal in intercropping before and after maize harvest, respectively. During shade period, soya bean biomass was severely reduced, and stem elongation was stimulated. Typical features of shade grown leaves were found, such as lower LMA (leaf mass per unit area), thinner thickness, higher chlorophyll content, lower chlorophyll a:b ratio. Whole-plant leaf area analysis found that soya bean increased leaf area ratio by adjusting leaf morphology rather than by dry mass allocation. After maize harvest, leaf area and leaf mass increased rapidly, contributing to compensation growth in intercropped soya bean. Meanwhile, physiological and anatomical traits of leaf went back to similar levels as grown in sole cropping. However, stem morphological traits were irreversible after removal of shade. Finally, no difference on seed weight per plant of soya bean was observed between relay intercropping and sole cropping. Based on these findings, we speculated the recovery growth might be the direct determining factor on pod formation in soya bean, and improvement on the capacity of recovery growth could increase yield of relay intercropped soya bean.
Intercropping is a sustainable agricultural practice widely used in Asian, African and Latin American countries to enhance food security and to use natural resources more efficiently. In intercropping situations, two or more crops are grown simultaneously in the same field during a part or the entire growing season (Francis, Citation1989; Willey, Citation1979). Relay intercropping is one type of intercropping (Duval, Citation2005; Nelson et al., Citation2011; Zhang et al., Citation2008b); many studies described the high productivity of relay strip intercropping (INT) and resource use efficiency levels (Andrade et al., Citation2012; Bedoussac & Justes, Citation2010; Echarte et al., Citation2011; Gao et al., Citation2009; Mushagalusa et al., Citation2008; Zhang et al., Citation2008b). In INT, the component crops are planted in different strips, and the late-planted crop is sown after the early-planted crop has reached its reproductive stage but before its harvest (Francis, Citation1989). The unique of relay intercropping is that the late-planted crop experiences two very different periods: the coexisting period and the solo-existing period. Due to the optimized planting dates of component crops, relay strip intercropping improves light interception based on the total photosynthetically active radiation (PAR) available per year within the window for favourable growing conditions (Spiertz, Citation2012).
During the coexisting period in INT, light is the most frequently limiting resource if water and nutrient requirements of crops are satisfied (Francis, Citation1989), and light is frequently the most important factor related to over yielding by crops mixtures that exhibit temporal complementarity and high efficiency (Malézieux et al., Citation2009). It is clear that shade impact only takes place during the coexisting period. Shade is ubiquitous in nature and all plants are shaded to some degree during their lifetime (Valladares & Niinemets, Citation2008). Under shade, plants often exhibit a remarkable adjustment, such as thinner leaf, higher chlorophyll (Chl) content per unit leaf mass with a increment of Chl b. (Valladares & Niinemets, Citation2008). Although most agronomic studies reported the increased total yield or biomass of intercropping, there were some studies out the reduced yield and biomass of the component crop in intercropping in paralleled with those in sole cropping (SOL). For example, the biomass and nutrient accumulation in intercropped soya bean (Glycine max) was significantly smaller than sole soya bean, and the rates of dry matter accumulation in the intercropped maize were significantly lower than sole maize (Li et al., Citation2001). And shading reduced total dry matter of beans by 67% at the end of the growing season when intercropped with maize, resulting in yield losses (Tsubo & Walker, Citation2004). Another study also found the increase in radiation productivity for maize–soya bean intercrop was the result of an increase in radiation use efficiency and of a minor but significant increase in radiation capture efficiency, and the yield attained by intercropping was mainly limited by a poor production of the soya bean component (Coll et al., Citation2012). Recent study on maize–soya bean relay intercropping revealed that the intercropped soya bean just captured 13.8% of PAR at the vegetative stage compared with those grown under SOL (Liu et al., Citation2015). And the planting geometries of maize–soya bean relay intercropping directly affect soya bean yield, and the distance between two component crops had a dominant function under narrower bandwidth of intercropping, and appropriate increase of the distance could be used to achieve high yields (Yang et al., Citation2015). These results suggest the shade impact on component crop is the main focus on determining total yield of intercropping.
After the harvest of early-planted crop, the late-planted crop recovers from shade to full sunlight condition. Previous report pointed out that the dry matter accumulation increased significantly, and this recovery is one factor contributing to yield advantage of intercropping (Li et al., Citation2001). In fact, the overlapped growing seasons of component crops in relay intercropping might be the key of over yielding. However, most studies only evaluated the yield of component crop in relay intercropping at the final harvests, the mechanisms on recover from shade of late-planted crop at agronomic and physiological levels were still not fully understood.
Soya bean is an important oil and protein crop in the world, and also one of the major crops planted in intercropping. Maize (Zea mays)–soya bean relay strip intercropping is one of the typical intercropping patterns around the world. In maize–soya bean relay strip intercropping, maize is superior component crop, the detrimental effects of soya bean over maize was ignorable (Echarte et al., Citation2011). The increase of total yield of intercropping was associated with the increase in intercropped soya bean yield when planted with tall component crops in relay intercropping (Andrade et al., Citation2012). Increases of soya bean yield without losses of maize yield are proper approaches improving overall yield of maize–soya bean relay intercropping. Our previous studies focused on the shade on morphological traits and lodging resistance of soya bean seedlings in this pattern (Liu et al., Citation2015; Yang et al., Citation2014). The period of recovery growth of soya bean after maize harvest lasts around two months, which cannot be neglected on yield formation. To better understand the shade response and recovery growth characteristics of soya bean in maize–soya bean relay intercropping, biomass, photosynthesis, leaf morph-anatomic traits of soya bean during coexisting period and the successive solo-existing periods were investigated. The purposes of this study were to test whether the recovery growth compensates the negative impact of shade for relay intercropped soya bean or not.
Materials and methods
Field site and experimental design
The experiment was conducted during the maize–soya bean relay strip intercropping growing season from March to October in 2011, at the Teaching and Experimental Farm of Sichuan Agricultural University, Ya’an (29°59′N, 103°00′E). The soil of the experimental field is a purple clay loam (pH 7.5), and at the beginning of experiment, total N, P, K, available N, P, K, and organic matter were 2.79 g kg−1, .383 g kg−1, 12.89 g kg−1, 168.6 mg kg−1, 81.3 mg kg−1, 140.1 mg kg−1, and 4.32%, respectively. The most widely used local indeterminate soya bean variety Gongxuan 1 was used in this experiment. Soya bean was planted in two cropping treatments: INT and SOL. Field experiment was completely randomized designed with 3 replications, totally containing 6 plots with 6 m length by 4 m width per plot. In SOL treatment, soya bean was planted as solid rows with .5 m row spacing. In INT treatment, soya bean and maize were planted as alternating strips, every soya bean strip was relay intercropped between maize strips. Each plot contained two maize and two soya bean strips, and each soya bean strip and maize strip consisted of 2 soya bean and 2 maize rows, respectively. Strip spacing (distance between maize and soya bean rows), soya bean row spacing and maize row spacing were all .5 m. In INT treatment, soya bean density was reduced by one half of SOL density, where individual plant of soya bean had the same growing space as in SOL condition. One soya bean strip was used for destructive measurement, and the other soya bean strip was used for yield determination at maturity. All strips were oriented north–south. Irrigation, weeding, fertilizers and other field practices were kept on the same level across all treatments. Maize (var. Chuandan 418) was sown in the seedbed on March 28th, and transplanted into field on April 9th within .4 m intra-row spacing and 2 plants per hole. Soya bean was drilled on June 11th within .35 m intra-row spacing and 2 plants per hole. The planting arrangement of two cropping patterns can be found in Figure . Maize was harvested on August 9th 2011, and soya bean was harvested more than two months later on October 25th, 2011.
Figure 1. Planting arrangements of relay strip intercropping (a) and sole cropping (b). Explanation: open circles (○) and closed circles (●) represent soya bean and maize holes, respectively. Each hole contained 2 plants.

Sampling and measurements
Sampling and measurements were performed three times during the experiment: July 16th, August 9th and August 26th, representing before, at and after the harvest of maize, respectively, and the growth stages of soya bean were V5, R2 and R4 (Fehr et al., Citation1971), respectively. In each plot, three soya bean plants from three holes were sampled for measurements of biomass and morphological traits, and other five plants were tagged for photosynthesis measurements and destructive sampling for Chl, anatomical structure determination.
Architecture and morphological traits of plant
Plants were cut at the ground level, and the aboveground parts were brought back to laboratory to measure biomass and morphological traits. Plants were separated into leaf and non-leaf organs. Leaves were scanned by flatbed scanner (CanoScan LiDE 200, Canon Inc., Japan), and then leaf area was measured using ImageJ 1.45s. Plant height (mainstem length) and stem diameter (first internode) were also determined. After the leaf and non-leaf organs (including stem and petiole) were oven-dried at 75 °C for 72 h, aboveground biomass, fraction of leaf mass (fL), leaf area ratio (LAR) and leaf area index (LAI) were then calculated. LAR was calculated by dividing leaf area by aboveground biomass per plant. LAI was calculated by multiplying plant density with leaf area per plant.
Physiological and anatomical traits of leaf
Photosynthetic rate was measured on the middle leaflets of the 3rd leaf from top on five tagged plants in each plot. Photosynthesis were measured with the portable photosynthesis system (Li-6400XT, Li-Cor Inc., USA) equipped with LED Light Source (6400-02B). Photosynthetic rate was measured under 1,000 μmol m−2 s−1 light intensity, 500 ml min−1 air flow rate, 380 μmol mol−1 sample CO2 concentrations, 30 °C and 50–75% relative humidity.
After net photosynthetic rate measurements, the trifoliolate leaves were sampled to laboratory. Two middle segments (5 mm × 8 mm) without midrib from each middle leaflet were cut out and fixed in FAA solution for later paraffin section observation. Total leaf thickness, palisade and spongy mesophyll thickness were quantified by ImageJ 1.45s.
Four leaf discs (diameter = 1 cm) were punched from each middle leaflet. Two discs were oven-dried at 65 °C to constant weight to determine dry weight and calculate leaf mass per unit area. The other two discs were extracted in 80% aqueous acetone solvent to determine total Chl content and the ratio of Chl a:b (Lichtenthaler, Citation1987). Remained leaves were oven-dried and milled into fine powder, and nitrogen content was determined after digesting by segmented flow analysis (FUTURA II, Alliance Instrument, France). Chl and nitrogen contents per unit area were determined then.
Yield
At harvest time, soya bean plants in six continuous holes in two rows (each row contained three holes) were manually harvested at R7 stage (Fehr et al., Citation1971). Plant density at maturity, seed weight per plant, pod number per plant, seed number per pod, 100-seed weight and soya bean yield per unit area were determined.
Statistics analysis
Mean values of variables from each plot was used for data statistics. Differences between cropping patterns were analysed by ANOVA in SPSS software (SPSS, Chicago, USA). All measured and calculated traits were set as dependent variable; cropping treatment was set as fixed factor.
Results
Morphological traits and growth
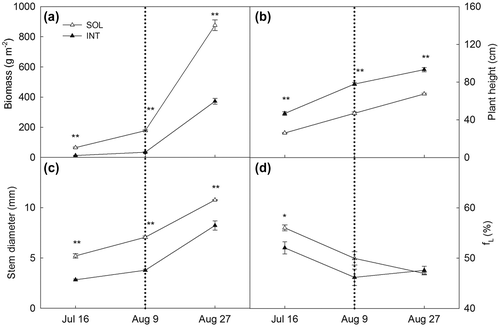
Reduced aboveground biomass was found in INT throughout the experiment (Figure (a). Biomass of soya bean was significantly reduced in INT throughout the experiment. But soya bean accumulated biomass rapidly after the time of maize harvest in both INT and SOL treatments. Soya bean expressed significant plant height elongation in INT treatment during coexisting period (Figure (b), while stem diameter reduced dramatically (Figure (c)). After the maize harvest, although plant height and stem diameter continued to increase in both treatments, the differences between two treatments were still significant. Fraction of leaf mass of INT was lower than SOL at the first sampling time during coexisting period, while fL of both treatments went to the same levels since the maize harvest (Figure (d)).
Figure 2. Biomass (a), plant height (b), stem diameter(c), fraction of leaf mass (d) of soya bean planted in sole cropping (SOL) and relay strip intercropping (INT). Explanation: fL: fraction of leaf mass; In all figures in this article, the vertical dash line indicates the harvest day of maize. ** means significant level at .01, and * means significant level at .05.
Morphological and anatomical traits of leaf
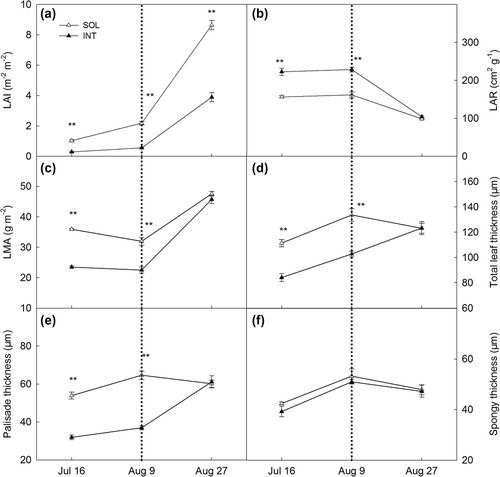
LAI in INT soya bean were significantly lower than SOL plants throughout the experiment. Although LAI increased rapidly after the time of maize harvest, the difference between the two treatments trended to be larger (Figure (a)). By contrast, significantly higher LAR was observed during coexisting period, but LAR of both treatments declined to similar levels after the maize harvest (Figure (b)). LMA of soya bean leaves grown in INT over the shade period were slightly above 20 g m−2(Figure (c)), significantly lower than that grown in SOL. After the maize harvest, LMA in INT started to increase rapidly. At the third sampling time, no difference between INT and SOL was observed. Coincided with the LMA results, soya bean grown under shade condition in INT showed significant thinner leaves than SOL counterparts during the coexisting period (Figure (d)). And after the maize harvest, total leaf thickness in INT treatment increased dramatically, reaching the level as the control SOL leaves. Analogous variations were found on palisade thicknesses (Figure (e)). While for spongy tissue, no difference was observed throughout the experiment (Figure (f)). Cross-sectional photograph of leaf can be found at the supplemental files.
Figure 3. LAI (a), LAR (b), leaf mass per unit area (c), total leaf thickness (d), palisade thickness (e) and spongy thickness (f) of soya bean planted in sole cropping (SOL) and relay strip intercropping (INT). Explanation: LAI: Leaf area index, LAR: leaf area ratio, LMA: leaf mass per unit area.
Photosynthetic traits of leaf
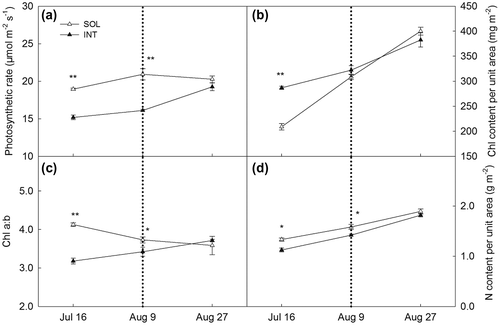
The photosynthetic rates measured under controlled PAR 1,000 μmol m−2 s−1 are shown in Figure (a). Significant lower photosynthetic rates of INT treatment were found during coexisting period. But no difference was found after the removal of shade. Chl contents per unit area were significantly higher in INT grown leaves at the first sampling time (Figure (b)). From the removal of maize plant, leaves grown in both treatments expressed similar Chl contents. Meanwhile, Chl a:b showed significant difference before harvest of maize (Figure (c)), inferring the favour on Chl b in this shade situation. After the maize harvest, Chl a:b between two treatments had no difference. Nitrogen contents showed only higher values under shade period before maize harvest (Figure (d)).
Figure 4. Photosynthetic rate (a), chlorophyll content per unit area (b), chlorophyll a:b ratio (c), nitrogen content per unit area (d) of soya bean leaves planted in sole cropping (SOL) and relay strip intercropping (INT). Explanation: Chl: chlorophyll, N: nitrogen.
Yield
For yield and its components, no significant differences on seed weight per plant, pod number per plant, seed number per pod and 100-seed weight between INT and SOL were found (Table ). However, the plant density at maturity of INT at the harvest time was significantly lower than SOL, which led significant lower yield of whole field.
Table 1. Yield and yield components of soya bean plant and in sole cropping and relay strip intercropping.
Discussion
Responses to shade of soya bean leaves during coexisting period
Under low light condition, plants growth is usually limited due to shortage ATP and energy supplied by photosynthesis (Niinemets, Citation2010; Valladares & Niinemets, Citation2008). Our results of reduced biomass (Figure ) of soya bean during coexisting period in maize–soya bean relay intercropping were similar to other intercropping studies (Bedoussac & Justes, Citation2011; Lithourgidis et al., Citation2011; Zhang et al., Citation2011). Response to shade is associated with a great deal of features, and most of these features are involved in leaf physiology, biochemistry, anatomy and morphology (Valladares & Niinemets, Citation2008). As photosynthesis plays an important role in leaf physiology for carbon gain (Givnish, Citation1988; Valladares & Niinemets, Citation2008), photosynthetic rates and related features of soya bean leaves were investigated. Lower photosynthetic rates of INT grown soya bean were found in this study under the measuring light intensity of 1,000 μmol m−2 s−1 (Figure ). Thus, we speculate that the reduced biomass was mainly caused by the low light availability during coexisting period.
Leaf Chl content is well established as a common reference to reflect shade response (Niinemets, Citation2010; Valladares & Niinemets, Citation2008). The effect of light intensity on the leaf Chl a:b ratio is one of the most characteristic differences between sun and shade leaves (Boardman, Citation1977). Decreased Chl a:b ratio can be explained by more and broader grana stacks in thylakoid, and most photosystem II bound Chl b locate in grana stacks (Boardman, Citation1977; Anderson, Citation1986). Increased total Chl content per unit dry mass is the result of thinner leaves and the relatively invariable Chl content per unit area (Niinemets, Citation2010). Our LMA and anatomical results provide evidence of the thinner leaves in INT treatments during coexisting period (Figure ). Thinner leaves coincided with previous studies that shade grown leaves especially had reduced thickness of palisade tissue (Terashima et al., Citation2006; Terashima et al., Citation2011; Terashima et al., Citation2001). Shade grown leaves are thought to be a consequence of less requirement of mesophyll surface to secure the area of CO2 diffusion under lower supplies of ATP and NADPH (Terashima et al., Citation2006). Therefore, thinner leaves usually had lower photosynthetic capacity, because carboxylation of photosynthesis takes place in chloroplast, and most chloroplasts locate in palisade tissue, thinner layer of palisade led to smaller surface area of mesophyll, which constrained CO2 diffusion from intercellular cavities into chloroplast (Terashima et al., Citation2006; Terashima et al., Citation2011).
Besides leaf physiological and anatomical features, plants also have acclimation traits of architecture and morphology at whole-plant level. In this study, we found very significant stem elongation of soya bean under shade during coexisting period (Figure ), which resulted in higher plant height and lower stem diameter. Meanwhile, we used fL, LAI and LAR to reflect shade responses at architectural level. It is clear that INT grown soya bean before removal of maize shading expressed slighter increase on fL, than control plants (Figure ). By contrast, LAR of shade grown plants increased by over 40% than control plants (Figure ). Our study confirmed that shaded soya bean increase LAR by leaf morphology far more than by allocation (Poorter et al., Citation2012).
Shade avoidance is a strategy plants evolved in response to shade. For shade avoidance, plants often increase stem and hypocotyl elongation at the expense of leaf growth, and the elongated hypocotyl is often seen as a indicator of shade avoidance (Ballaré, Citation1999; Casal, Citation2012; Franklin & Whitelam, Citation2005; Smith, Citation2000; Vandenbussche et al., Citation2005). Overall, the stronger elongated stem and altered architectural confirmed that soya bean dealt with shading light in relay intercropping via shade avoidance in this study. Therefore, architectural and morphological traits might be more variable than physiological traits for soya bean, a typical sun favouring crop, when grown under shade condition. Selection standard of soya bean cultivar with better adaptation to shade of relay strip intercropping could be based on the architectural and morphological traits of seedling.
Recovery from shade and yield formation
Environmental changing on temporal scale is the unique interesting feature of relay intercropping, shade impact only exists at soya bean early growing stage. Many studies have reported the compensation or recovery growth of the later-planted crops in relay intercrop (Li et al., Citation2001; Zhang et al., Citation2008a). In this study, the difference results of investigated features between coexisting and recovery periods provide further evidence for temporal variations of responses to light conditions in relay intercropped soya bean. After the growth recovery, all investigated leaf physiological features displayed no difference between INT and SOL conditions, such as photosynthetic rate, Chl content and anatomical structures. It is probably that the soya bean, at least the cultivar Gongxuan 1, in our study had strong recovery capacity from shade in relay intercropping. Meanwhile, as the rapid recovery growth and biomass accumulation after maize harvest, biomass partition to leaf and leaf area (Figure ) increased to similar levels with the SOL plants. Therefore, these observations indicated that the recovery growth was mainly contributed by rapid increment of leaf area. By contrast, for morphological and architectural traits, only plant height and stem diameter still expressed irreversible difference between two cropping patterns after removal of shade. Combined with these results together, we speculate that the coexisting and the successive solo-existing periods have different effects on soya bean growth, and the recovery capacity, especially on leaf growing capacity can eliminate shade impact during coexisting period.
In maize–soya bean relay intercropping in this study, the initial flowering stage (R1) of soya bean (Fehr et al., Citation1971) was around the harvesting time of maize, thus, shade condition did not occur during soya bean reproductive stages. Previous studies pointed out that shade during vegetative growth reduced soya bean biomass at initial flowering stage, but had no effect on crop growth rate during flowering and podset (R1 to R5) or seeds per square metre if light interception after R1 was not reduced (Jiang & Egli, Citation1995). Our studies indirectly supported this mechanism that after the harvest of maize, shade effect was removed, then, soya bean growth (Figure ) and yield per plant (Table ) was not reduced in relay intercropping.
Moreover, the harvest of maize left empty strips in field, which might increase light availability and growing space for soya bean and consequently induced recovery growth in INT treatment. Thus, our findings could be accounted for by assuming that recovery growth at the late stage after harvest of maize in relay intercropping may be the key factors for yield formation of soya bean. The light condition caused by empty strips in field was similar to light enrichment treatment in previous study, in which the enriched light could increase soya bean pod number (Liu et al., Citation2006). Hence, the pod formation of relay intercropped soya bean might be profited from the empty strips after maize harvest. Unfortunately, as the first investigation on recovery growth of soya bean in relay intercropping, only one sampling after maize harvest was conducted in this study. At this time, soya bean was at around R4 stage growth, and there was about two months from R4 to maturity. Whether the recovery growth increased seed filling after maize harvest or not needs to be studied in the future.
Conclusion
During the early coexisting shade period in maize–soya bean relay strip intercropping, soya bean expressed typically stem elongation and leaf anatomical and physiological variations. After the harvest of maize, physiological and anatomical traits of leaf, and leaf area and dry mass fraction of whole plant recovered to similar levels as control plants grown in sole cropping. However, morphological traits of stem were irreversible after the removal of shade. Finally, no yield loss of individual plant was observed in intercropping because of recovery growth effect after maize harvest. Hence, this study suggested that recovery growth of leaf might be significantly positive factor for yield formation of soya bean in intercropping. Improvement on recovery capacities after removal of shade might be a new focus for increasing the productivity of relay intercropped soya bean.
Supplementary Data
The supplemental file for this article can be accessed at https://figshare.com/s/1471f2ac79e3f700a60f.
Disclosure statement
No potential conflict of interest was reported by the authors.
Funding
This work was supported by National Natural Science Foundation of China [grant number 31171476], [grant number 31071373]; National Program on Key Basic Research Project [2011CB100402]; the China Agriculture Research System [CARS-04-PS19].
TPPS_1128095_supp.jpg
Download JPEG Image (97.8 KB){kind=link}
References
- Anderson, J. M. (1986). Photoregulation of the composition, function, and structure of thylakoid membranes. Annual Review of Plant Physiology, 37, 93–136.10.1146/annurev.pp.37.060186.000521
- Andrade, J. F., et al. (2012). Sunflower–soybean intercrop productivity under different water conditions and sowing managements. Agronomy Journal, 104, 1049–1055.10.2134/agronj2012.0051
- Ballaré, C. L. (1999). Keeping up with the neighbours: Phytochrome sensing and other signalling mechanisms. Trends in Plant Science, 4, 97–102.10.1016/S1360-1385(99)01383-7
- Bedoussac, L., & Justes, E. (2010). Dynamic analysis of competition and complementarity for light and N use to understand the yield and the protein content of a durum wheat-winter pea intercrop. Plant and Soil, 330, 37–54.10.1007/s11104-010-0303-8
- Bedoussac, L., & Justes, E. (2011). A comparison of commonly used indices for evaluating species interactions and intercrop efficiency: Application to durum wheat–winter pea intercrops. Field Crops Research, 124, 25–36.10.1016/j.fcr.2011.05.025
- Boardman, N. K. (1977). Comparative photosynthesis of sun and shade plants. Annual Review of Plant Physiology, 28, 355–377.10.1146/annurev.pp.28.060177.002035
- Casal J. J. (2012). Shade avoidance. The Arabidopsis Book, 10, e0157.
- Coll, L., et al. (2012). Capture and use of water and radiation in summer intercrops in the south-east Pampas of Argentina. Field Crops Research, 134, 105–113.10.1016/j.fcr.2012.05.005
- Duval, J. R. (2005). Relay-intercropping does not reduce strawberry yield in an annual-hill production system. HortTechnology, 15, 907–908.
- Echarte, L., et al. (2011). Yield response to plant density of maize and sunflower intercropped with soybean. Field Crops Research, 121, 423–429.10.1016/j.fcr.2011.01.011
- Fehr, W. R., et al. (1971). Stage of development descriptions for soybeans, Glycine max (L.) Merrill. Crop Science, 11, 929–931.10.2135/cropsci1971.0011183X001100060051x
- Francis, C. A. (1989). Biological efficiencies in multiple-cropping systems. Advances in Agronomy, 42, 1–42.10.1016/S0065-2113(08)60522-2
- Franklin, K. A., & Whitelam, G. C. (2005). Phytochromes and shade-avoidance responses in plants. Annals of Botany, 96, 169–175.10.1093/aob/mci165
- Gao, Y., et al. (2009). Crop coefficient and water-use efficiency of winter wheat/spring maize strip intercropping. Field Crops Research, 111, 65–73.10.1016/j.fcr.2008.10.007
- Givnish, T. J. (1988). Adaptation to sun and shade: A whole-plant perspective. Functional Plant Biology, 15, 63–92.
- Jiang, H., & Egli, D. B. (1995). Soybean seed number and crop growth rate during flowering. Agronomy Journal, 87, 264–267.10.2134/agronj1995.00021962008700020020x
- Li, L., et al. (2001). Wheat/maize or wheat/soybean strip intercropping: II. Recovery or compensation of maize and soybean after wheat harvesting. Field Crops Research, 71, 173–181.10.1016/S0378-4290(01)00157-5
- Lichtenthaler, H. K. (1987). Chlorophylls and carotenoids: Pigments of photosynthetic biomembranes. Methods in Enzymology, 148, 350–382.10.1016/0076-6879(87)48036-1
- Lithourgidis, A. S., et al. (2011). Dry matter yield, nitrogen content, and competition in pea-cereal intercropping systems. European Journal of Agronomy, 34, 287–294.10.1016/j.eja.2011.02.007
- Liu, W., et al. (2015). Evaluation of soybean (Glycine max) stem vining in maize-soybean relay strip intercropping system. Plant Production Science, 18, 69–75.10.1626/pps.18.69
- Liu, X., et al. (2006). Yield and yield components responses of old and new soybean cultivars to source-sink manipulation under light enrichment. Plant, Soil and Environment, 52, 150–158.
- Malézieux, E., et al. (2009). Mixing plant species in cropping systems: Concepts, tools and models. A review. Agronomy for Sustainable Development, 29, 43–62.10.1051/agro:2007057
- Mushagalusa, G. N., et al. (2008). Shoot and root competition in potato/maize intercropping: Effects on growth and yield. Environmental and Experimental Botany, 64, 180–188.10.1016/j.envexpbot.2008.05.008
- Nelson, K. A., et al. (2011). Harvest aid application timing affects wheat and relay intercropped soybean yield. Agronomy Journal, 103, 851–855.10.2134/agronj2010.0384
- Niinemets, Ü. (2010). A review of light interception in plant stands from leaf to canopy in different plant functional types and in species with varying shade tolerance. Ecological Research, 25, 693–714.10.1007/s11284-010-0712-4
- Poorter, H., et al. (2012). Biomass allocation to leaves, stems and roots: Meta-analyses of interspecific variation and environmental control. New Phytologist, 193, 30–50.10.1111/j.1469-8137.2011.03952.x
- Smith, H. (2000). Phytochromes and light signal perception by plants – an emerging synthesis. Nature, 407, 585–591.10.1038/35036500
- Spiertz, H. (2012). Avenues to meet food security. The role of agronomy on solving complexity in food production and resource use. European Journal of Agronomy, 43, 1–8.10.1016/j.eja.2012.04.004
- Terashima, I., et al. (2006). Irradiance and phenotype: Comparative eco-development of sun and shade leaves in relation to photosynthetic CO2 diffusion. Journal of Experimental Botany, 57, 343–354.
- Terashima, I., et al. (2011). Leaf functional anatomy in relation to photosynthesis. Plant Physiology, 155, 108–116.10.1104/pp.110.165472
- Terashima, I., et al. (2001). Why are sun leaves thicker than shade leaves? – Consideration based on analyses of CO2 diffusion in the leaf. Journal of Plant Research, 114, 93–105.10.1007/PL00013972
- Tsubo, M., & Walker, S. (2004). Shade effects on Phaseolus vulgaris L. intercropped with Zea mays L. under well-watered conditions. Journal of Agronomy and Crop Science, 190, 168–176.10.1111/jac.2004.190.issue-3
- Valladares, F., et al. (2007). Ecological limits to plant phenotypic plasticity. New Phytologist, 176, 749–763.10.1111/nph.2007.176.issue-4
- Valladares, F., & Niinemets, Ü. (2008). Shade tolerance, a key plant feature of complex nature and consequences. Annual Review of Ecology, Evolution, and Systematics, 39, 237–257.10.1146/annurev.ecolsys.39.110707.173506
- Vandenbussche, F., et al. (2005). Reaching out of the shade. Current Opinion in Plant Biology, 8, 462–468.10.1016/j.pbi.2005.07.007
- Willey, R. W. (1979). Intercropping: Its importance and research needs. I. Competition and yield advantages. Field Crop Abstracts, 32, 1–10.
- Yang, F., et al. (2014). Growth of soybean seedlings in relay strip intercropping systems in relation to light quantity and red: Far-red ratio. Field Crops Research, 155, 245–253.10.1016/j.fcr.2013.08.011
- Yang, F., et al. (2015). Yield response to different planting geometries in maize-soybean relay strip intercropping systems. Agronomy Journal, 107, 296–304.10.2134/agronj14.0263
- Zhang, G., et al. (2011). Interspecific competitiveness affects the total biomass yield in an alfalfa and corn intercropping system. Field Crops Research, 124, 66–73.10.1016/j.fcr.2011.06.006
- Zhang, L., et al. (2008a). Nitrogen economy in relay intercropping systems of wheat and cotton. Plant and Soil, 303, 55–68.10.1007/s11104-007-9442-y
- Zhang, L., et al. (2008b). Light interception and utilization in relay intercrops of wheat and cotton. Field Crops Research, 107, 29–42.10.1016/j.fcr.2007.12.014