Abstract
In 2003, severe barrenness due to low temperatures in hybrid maize (Zea mays L.) cultivar occurred in the Konsen region, one of the coldest areas in Japan. Varietal difference in grain set, and abnormal flowering habits and tassel growth were observed in the field in 2003. For the cultivar 39B29, which was affected severely, anthesis occurred several days after silking, and tassel size was reduced. For the cultivar Papirika, which was not affected by low temperature, anthesis began 0–1 d before silking, and tassel size was similar to that in previous years. In a 2011–2013 experiment, three varieties, Papirika, 39B29, and Kwiss were treated at 10 °C for 7 d at 6- to 9-leaf stage. Following the treatment, anthesis was delayed, and tassel length was reduced, in 39B29 and Kwiss, but not in Papirika. The relative tassel length (RTL, ratio of tassel length of experimental plants to that of untreated control plants) was around 60 in 39B29, 80 in Papirika, and 30 in Kwiss on average over 3 years. Based on field observations in 2003 and the pot experiment in 2011–2013, a cultivar whose RTL was 60 or less was deemed sensitive, and a cultivar whose RTL was 80 or more was deemed less sensitive to low-temperature stress during the tassel development stage.
Introduction
The climate of the Konsen region is generally characterized by a cool and foggy summer, and short growing season. Since the Konsen region is one of the main centers of dairy farming in Japan, many farmers cultivate maize (Zea mays L.) as a high-energy forage crop. In this region in 2003, a very cool summer year, severe barrenness occurred in the cultivar widely used in maize fields. One of the reasons for the barrenness in 2003 was due to the abnormal tassel growth (delayed anthesis from silking and reduced tassel length) caused by the extremely low-air temperature and low-solar radiation at the tassel development stage (Hayashi et al., Citation2015). To prevent the occurrence of barrenness associated with low temperature, more information on the varietal difference in the sensitivity to low temperature is needed. However, to the best of my knowledge, the varietal difference in barrenness or tassel growth abnormality in hybrid maize cultivars in relation to low-temperature stress has not been reported.
In maize inbred lines, there are reports evaluating genotypic differences in low-temperature sensitivity to tassel development (Bechoux et al., Citation2000; Tranel et al., Citation2009). The varietal differences in the sensitivity to low-temperature stress during the reproductive growth stage have been reported in rice (Oryza sativa L.) (Satake, Citation1991), soybean (Glycine max (L.) Merr.) (Kurosaki et al., Citation2003), and sorghum (Sorghum bicolor (L.) Moench) (Brooking, Citation1979). If there are differences in low-temperature sensitivity during tassel development among commercial maize hybrids, farmers may be able to mitigate yield reduction by using less-sensitive hybrids in these cool summer regions.
The experiments described herein were intended to investigate differences in flowering characteristics and tassel morphogenesis associated with low-temperature stress between maize varieties. Furthermore, the method for detecting the varietal differences in sensitivity to low-air temperatures is discussed.
Materials and methods
Field experiments
Flowering conditions were observed in 2001, 2002, and 2003 in the test field at the Konsen Agricultural Experiment Station (KAES) located in Nakashibetsu, Hokkaido (43°33′N, 144°59′E, 50 m above sea level). In addition, tassel morphogenesis and ear fertility were observed in 2003 in a farmer’s field near the KAES (approximately 15 km south of KAES). Two cultivars of the same maturity class of 75 RM, i.e. 39B29 (Pioneer Hi-Bred, Iowa, USA, dent × flint single-cross hybrid) and Papirika (NARO Hokkaido Agricultural Research Center, Sapporo, Japan, flint × flint single-cross hybrid) were used in the field experiment.
In the KAES field, the plants were sown on 16 May 2001, 22 May 2002, and 2003 4 rows (4 m long spaced 0.72 m apart, hill distance 0.18 m) with 4 replications. Matured manure (40 ton ha−1) was applied before plowing. Chemical fertilizer consisting of 8 g of N, 21 g of P2O5, and 6 g of K2O m−2 was applied within the rows at sowing time, and of 4 g of N and 4 g of K2O m−2 was applied besides the row when the ninth leaf appeared. The dates of anthesis and silking were recorded when more than 50% of the plants extruded anther and silk, respectively. With these data, the anthesis-silking interval (ASI) was calculated as the number of days from silking to anthesis. In the farmer’s field, maize plants were sown on 25 May 2003 with a vinyl mulch cultivation system (row space 0.76 m apart, hill distance 0.15 m). Fertilizer application and other cultivation methods followed the farmer’s conventional practices. Tassel morphogenesis and ear fertility were visually surveyed using a plot in the field for moderately grown plants (there were no replications).
Pot experiments
Plant materials and culture conditions
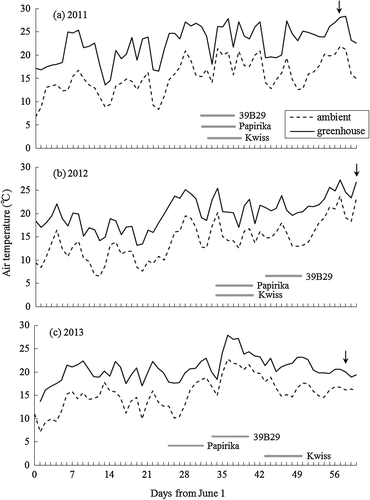
Cultivars 39B29, Papirika, and Kwiss (KWS SAAT AG, Germany, flint × dent single-cross hybrid, 73 RM) were grown in a greenhouse in May or June in 2011, 2012, and 2013. As Kwiss was not cultivated in 2003 in the Konsen region, its sensitivity to cool summer condition is unknown. Growth conditions in pots were the same as those reported previously (Hayashi et al., Citation2015). In brief, plants were grown in a greenhouse at KAES using 1/2000 a Wagner pots. Chemical fertilizers were applied at the time of sowing and after the low-temperature treatment described below. The air temperature in the greenhouse was maintained above 20 °C (Figure ) and each pot was adequately watered.
Figure 1. Daily mean air temperatures in the greenhouse and the ambient temperatures during the pot experiment. Solid line represents greenhouse temperature; dashed line represents ambient temperature. Horizontal bars indicate 8-leaf stage low-temperature treatments. Cultivar 39B29 in 2012 and cultivar Kwiss in 2013 were sown later as compared to other cultivars. Arrows indicate the anthesis date of 39B29 (2011 and 2013) and Kwiss (2012).
Low-temperature treatment
In experiment 1 (Exp1), varietal differences under low-temperature conditions, simulating the field conditions of 2003, were examined. The plants were treated at 13 °C for 15 d or 21 d at the sensitive stage described previously (Hayashi et al., Citation2015) (i.e. 8-leaf stage) by moving the pots from the greenhouse to the air conditioned rooms (Koito Industries, Yokohama, Japan. Floor space: 2.3 m × 2.6 m). In the air conditioned rooms, the solar radiation was reduced to 30% of the greenhouse levels, to replicate the field conditions of the Konsen region in July 2003.
In experiment 2 (Exp2), the low-temperature sensitive stage of the maize varieties was examined using the procedure described by Bechoux et al. (Citation2000). When the plants expanded the sixth, seventh, eighth, or ninth leaf, the pots were treated at 10 °C for 7 d by moving the pots from the greenhouse to the air conditioned rooms. Shading was implemented as described in Exp1 in the air conditioned rooms. For both experiments, three pots were used for each cultivar per treatment, and the other three pots were remained in the greenhouse as control plants. Pots were moved between 0800 and 1000 am, with an interval of more than 10 min in a cool and shaded place to acclimate to the new environment.
The leaf stage (LS) was determined as described previously (Hayashi et al., Citation2015); the number of expanded leaves included the lowermost short true leaf and ended with the uppermost developed leaf, and the leaf just above the uppermost developed leaf was used for determining the decimal number, which was a ratio of the length of the elongated part. Since each plant was assigned to a treatment group just after thinning, and the plant growth differed slightly among pots, the actual LS at the start of the treatment differed from the expected stage. Therefore, the number of observation for each treatment was varied.
Data collection
Dates of anthesis and silking were recorded for each plant. The anthesis day was determined as the day when more than one spikelet in the tassel extruded an anther, and the silking date as the day when at least one pistil extruded from a plant’s ear. After the anthesis period, total tassel length, including the branches bearing spikelets, was measured to evaluate the effect of low temperature on the tassel morphogenesis. Since the total tassel length varied depending on the cultivars and the test year, relative tassel length (RTL) was calculated as the ratio of total tassel length of the treated plants to that of the control plants in each year. The length of the tassel’s central axis at the 7-LS and 8-LS was measured by dissecting two spare plants to identify the tassel growth stage. Climate data were collected from the Automated Meteorological Data Acquisition System at KAES (Japan Meteorological Agency, Citation2014).
Statistical analysis
All statistical processes were performed with R version 2.12.2 (R Foundation for Statistical Computing, Vienna, Austria). ANOVA was conducted to evaluate the effect of low-temperature treatments. For Exp1 data, averages of the treatment for each year were calculated and the test years were assigned as replications. For Exp2 data, the LS at the start of the treatment were rounded to whole numbers to create discrete variables, and the test years were assigned as replications. When significant differences were detected by ANOVA, multiple comparisons were performed with the Tukey’s HSD test. Multiple comparisons were performed with Dunnett’s test, when it was needed, to establish the significance of treatment differences with respect to control values.
Results
Flowering conditions and tassel morphogenesis in fields
In the farmer’s field in 2003, tassels with fewer spikelets (reduced tassel length) and less fertile ears (lack of kernel set) were observed in cultivar 39B29 as described in Hayashi et al. (Citation2015), but not in cultivar Papirika via the visual assessment (Figure ). The ear dry weight and the ratio of ear dry weight to whole crop dry weight was 337 kg 10a−1 and 26.3% in 39B29, and 547 kg 10a−1 and 46.3% in Papirika at the farmer’s field.
Figure 2. Tassel morphogenesis and ear fertility of cultivar Papirika in the farm field in 2003. Photos taken on 16 September 2003. For cultivar 39B29, see Figure 1 and Figure in Hayashi et al. (Citation2015).

In the test field of the KAES in 2001, 2002, and 2003, anthesis tended to occur after silking in 39B29, although maize is a protandrous plant. Especially in 2003, anthesis occurred 7 d after silking in 39B29, but 1 day before in Papirika (Table ). The ASI between 39B29 and Papirika was significantly different (p < 0.05) in the two years, including 2003.
Table 1. Flowering characteristics in the test field of the KAES.
Pot experiments
Experiment 1
Table shows the results of the low-temperature treatment of 13 °C for 15 d and 21 d conducted during 8-LS. In 39B29, anthesis delayed from silking for 3–4 d and tassel lengths were shortened. In Papirika, on the contrary, anthesis occurred before silking, even after the 21 d treatment, and tassel length was more than 70% of the untreated control plants. Kwiss showed similar results as 39B29, but the delay of anthesis from silking and the reduction in tassel length was larger.
Table 2. Effects of the low-temperature treatment (13°C for 15 or 21 days) on the flowering characteristics and tassel morphogenesis.
Experiment 2
Low-temperature treatment (10 °C for 7 d) retarded flowering events for all treated plants compared to control plants (Table ). The delay was greater for anthesis than for silking, especially for the cultivars 39B29 and Kwiss, with the treatments started at 7- to 9-LS. In Papirika, such treatment effects were not observed. In the control plants of all test cultivars, anthesis occurred before silking. Tassel length was also affected by the low-temperature treatment. In 39B29, tassel length tended to be smaller when treated at 8-LS compared to the control (p = 0.058), but not for any other LS (Table ). In Papirika, in contrast, tassel length was shortened compared to the control plants, although it did not decrease remarkably for a specific starting stage of the treatment. In Kwiss, tassel length became significantly shorter when treated at 8-LS compared to the control (p = 0.013).
Table 3. Effects of the low-temperature treatment (10 °C for 7 d) on the flowering characteristics and the total tassel length.
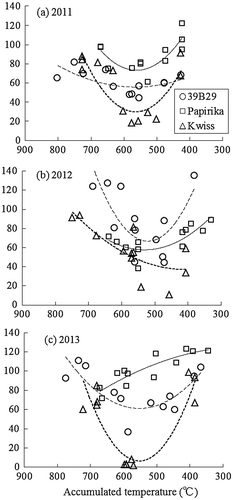
Figure shows the effects of LS at the start of low-temperature treatment on RTL. The RTL was generally reduced by the treatment starting at 7- to 8-LS in 39B29 and Kwiss, but not in Papirika. Focused on the varietal difference, RTL was significantly shorter in Kwiss and tended to be shorter in 39B29 as compared to Papirika in the 8-LS treatment (Table ). For the other LS, the RTL of 39B29 and Kwiss did not differ significantly from Papirika. When the LS in Figure was replaced to the accumulated daily mean air temperature from the starting date of the treatment to the date of anthesis, RTL tended to decrease when the plants were subjected to low temperature at 550 °C before anthesis (Figure ). A temperature of 550 °C was reached about 25 d before anthesis under greenhouse conditions (Figure ).
Figure 3. Relationships between the leaf stage at the start of the low-temperature treatment and the tassel length. RTL was calculated by determining the ratio of the tassel length of treated plants to the mean length of control plants. The regression curves are fitted for each cultivar (coarse broken line, 39B29 (○); solid line, Papirika (□); fine broken line, Kwiss (∆)). The mean lengths of the control plants in 2011, 2012, and 2013 were as follows: 220.7, 198.7, and 271.4 cm in Papirika, 174.4, 118.3, and 102.1 cm in 39B29, and 140.8, 114.8, and 100.2 cm in Kwiss.
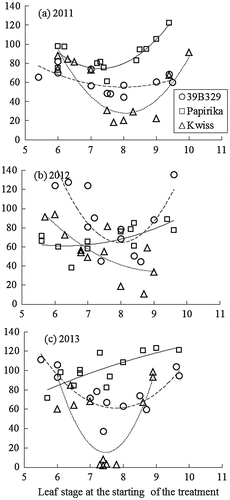
Table 4. RTL in the plants treated at 10 °C for 7 d starting at different leaf stages (LS).
Figure 4. RTL plotted against the accumulated daily mean air temperature from the day that low-temperature treatment began to the day of anthesis. RTLs and the mean tassel lengths of the control plants are the same as in Figure 3. The regression curves are fitted for each cultivar (coarse broken line, 39B29 (○); solid line, Papirika (□); fine broken line, Kwiss (∆)).
The length of the tassel central axis at 8-LS was approximately 140 mm in Papirika, and 40 mm in both 39B29 and Kwiss, while at 7-LS, it was around 50 mm in Papirika, and 10 mm in 39B29 and Kwiss (Table ). ANOVA revealed that the length of the tassel central axis in Papirika in 7-LS did not differ significantly from that of 39B29 and Kwiss in 8-LS.
Table 5. The length of tassel central axis at 7- and 8-leaf stage (LS).
Discussion
Existence of varietal difference
Varietal difference on tassel development due to low temperature is known for some temperate crops. In rice, Satake (Citation1991) showed reduction of the anther length in response to low-temperature treatment was different between test cultivars. In soybean, Kurosaki et al. (Citation2003) tested cold-weather tolerance during the flowering stage using two cultivars of different sensitivities. In sorghum, Brooking (Citation1979) stated that low-temperature treatment imposed at the leptotene stage of microsporogenesis reduced fertility for one genotype. Additionally, in maize inbred lines, the genotypic difference in the sensitivity to low temperature on tassel growth was reported by Tranel et al. (Citation2009) and Bechoux et al. (Citation2000).
In the farmer’s field, 39B29 reduced its ear yield and tassel length as reported in Hayashi et al. (Citation2015), whereas Papirika did not show such abnormalities in a same field. In the test field at the KAES, flowering characteristics in 39B29 were abnormal in 2003 (anthesis delayed from silking) and different from those in Papirika. In addition, the low-temperature treatment simulated the field conditions of 2003 (13 °C for 15 or 21 d starting at 8-LS) showed almost the same delay of anthesis and the same reduction of tassel length as the field conditions in 39B29 and in Kwiss (not cultivated in 2003), but not in Papirika. Therefore, Papirika is seemingly less sensitive to the low-temperature stress than 39B29 and Kwiss.
However, even if Papirika did not show barrenness or impaired tassel development in 2003, it was still unclear if it could overcome the low-temperature stress in 2003. It is possible that Papirika was not at its most sensitive stage during the severe low-temperature period in the field. The fact that the maximum leaf number was fewer in Papirika than in 39B29 and Kwiss (i.e. 12 in Papirika and 14 in 39B29 and Kwiss; data not shown in the Results section), and the flowering events started earlier in Papirika than in 39B29 and Kwiss suggest that Papirika reaches the sensitive stage earlier than the other two cultivars. Therefore, treatments at different tassel development stages will be necessary to clarify the varietal difference in the sensitivity to the low-temperature stress.
In this experiment, the length of the central axis of the tassel in Papirika at 7-LS was similar to that of 39B29 and Kwiss at 8-LS. Since the tassel develops with the elongation of the central axis (Bonnett, Citation1948), Papirika was assumed to be at the same developmental stage in 7-LS with 39B29 and Kwiss in 8-LS, namely the most sensitive stage. The result of the present study demonstrated that Papirika did not show longer ASI or shorter tassel length by the low-temperature treatment between 6-LS and 9-LS. Thus, Papirika can be considered less sensitive to the low-temperature stress at the tassel development stage compared to 39B29 and Kwiss. It means that there is a varietal difference in the effect of the low-temperature stress at the tassel development stage.
Cultivars 39B29 and Kwiss are the hybrids between dent and flint inbreds, and bred far countries from Hokkaido, whereas Papirika is a flint × flint hybrid and is bred in Hokkaido (Koinuma et al., Citation2007). Dent and flint genotypes differ genetically, and the hybrids of dent × flint crosses show significantly higher productivity (greater hybrid vigor) than that of both the dent × dent and flint × flint crosses (Moreno-González et al., Citation2000). Flint genotypes generally have more adaptability to the cool summer regions than do the dent genotypes, as they can grow well in early spring, even in low temperatures (Bhosale et al., Citation2007; Frei, Citation2000; Troyer, Citation2004). Flint genotypes may adapt to cool summer regions as compared to dent genotypes as to show less sensitivity to low-temperature stress during the tassel development stage.
Method to evaluate the varietal difference
Low-temperature treatment needed to be conducted at the several growing stages, because the relationship between LS at the start of the treatment and RTL cannot be stable between the test years (i.e. 2012 in Figure ). Treatment simulating natural condition of 2003 such as 13 °C for 21 d is not suitable to determine the sensitive stage or extent of sensitivity, especially for large numbers of cultivars. It is required to reduce treatment duration, so a 10 °C for 7 d between 6-LS and 9-LS treatment is carried out for Exp2 in the present study (referring to Bechoux et al. (Citation2000), which reported the effect of low temperature on tassel development of maize inbred lines). The results show that the lowest RTL value was around 80 in the plants treated starting at between 7-LS and 8-LS in Papirika, 60 in plants treated at 8-LS in 39B29, and 30 in plants treated at 8-LS in Kwiss on average over 3 test years. The lowest values of the RTL are stable among the test years excluding 2012. Therefore, the sensitivity to low temperature during the tassel development stage for the cultivars at around 75 RM can be evaluated by the following procedure: (a) subject cultivars to low temperature of 10 °C for 7 d from the time when LS is 6–9, (b) repeat for more than 2 years, (c) cultivars with tassel length more than 80% of the untreated control plants are deemed less sensitive, and cultivars with tassel length less than 60% of the untreated control plants are deemed sensitive.
The sensitive stage may differ depending on its maturity class or the maximum leaf number. For the other maturity groups or the cultivars with different maximum leaf numbers, the sensitive stage should be estimated as follows: when the accumulated daily mean air temperature from the start of the treatment to anthesis is 550 °C, or when the length of tassel central axis is about 40 mm.
In rice, deep watering to protect the panicle from low temperatures during the sensitive period (Satake et al., Citation1988) is used in Hokkaido in commercial cultivation to avoid the occurrence of barrenness. However, because maize grows tall, and commercial dairy farming requires large fields, such protective cultivation practices cannot be applied. Adjusting the planting season to move the most sensitive stage of maize plants away from the low-temperature period could prevent damage, but this is limited in the cool summer region due to the short growing season. Thus, selection of a tolerant maize cultivar will be one way to prevent low-temperature-induced barrenness. To develop or introduce new cultivars for the cool summer regions, the sensitivity to low temperature during the tassel development stage must be evaluated using the procedure described above. In addition, farmers in the cool summer regions must be informed of the sensitivity of distributed cultivars to foster sustainable maize cultivation.
| Abbreviations | ||
| ANOVA | = | analysis of variance |
| ASI | = | anthesis-silking interval |
| DAS | = | days after sowing |
| Exp | = | Experiment |
| LS | = | leaf stage |
| RM | = | relative maturity |
| RTL | = | relative tassel length |
Acknowledgments
I express my appreciation to the staff of the Hokkaido Prefectural Nemuro Agricultural Extension Center for their valuable assistance in the survey of farmers’ fields in 2003.
References
- Bhosale, S. U., Rymen, B., Beemster, G. T. S., Melchinger, A. E., & Reif, J. C. (2007). Chilling tolerance of central European maize lines and their factorial crosses. Annals of Botany, 100, 1315–1321.10.1093/aob/mcm215
- Bechoux, N., Bernier, G., & Lejeune, P. (2000). Environmental effects on the early stages of tassel morphogenesis in maize (Zea mays L.). Plant, Cell and Environment, 23, 91–98.10.1046/j.1365-3040.2000.00515.x
- Bonnett, O. T. (1948). Ear and tassel development in maize. Annals of the Missouri Botanical Garden, 35, 260–287.
- Brooking, I. R. (1979). Male sterility in Sorghum bicolor (L.) Moench induced by low night temperature. II. Genotypic differences in sensitivity. Australian Journal of Plant Physiology, 6, 143–147.10.1071/PP9790143
- Frei, O. M. (2000). Changes in yield physiology of corn as a result of breeding in northern Europe. Maydica, 45, 173–183.
- Hayashi, T., Makino, T., Sato, N., & Deguchi, K. (2015). Barrenness and changes in tassel development and flowering habit of hybrid maize associated with low air temperatures. Plant Production Science, 18, 93–98.10.1626/pps.18.93
- Japan Meteorological Agency. (2014). Climatic data retrieval. Retrieved September 4, 2014, from http://www.data.jma.go.jp/obd/stats/etrn/index.php**.
- Koinuma, K., Miura, Y., Miki, K., Enoki, H., Sato, H., Sato, N., … Sawada, Y. (2007). Breeding of an early maturing silage maize cultivar, “Papirika”, with high yielding ability and high ear content. Research Bulletin National Agricultural Research Center for Hokkaido region 186, 1–15*.
- Kurosaki, H., Yumoto, S., & Matsukawa, I. (2003). Pod setting pattern during and after low temperature and the mechanism of cold-weather tolerance at the flowering stage in soybeans. Plant Production Science, 6, 247–254.10.1626/pps.6.247
- Moreno-González, J., Martínes, I., Brichette, I., López, A., & Castro, P. (2000). Breeding potential of European flint and U.S. corn belt dent maize populations for forage use. Crop Science, 40, 1588–1595.10.2135/cropsci2000.4061588x
- Satake, T., Lee, S., & Koike, S. (1988). Male sterility caused by cooling treatment at the young microspore stage in rice plants. XXVIII. Prevention of cool injury with the newly devised water management practices. Effects of the temperature and depth of water before the critical stage. Japanese Journal of Crop Science, 57, 234–241.10.1626/jcs.57.234
- Satake, T. (1991). Male sterility caused by cooling treatment at the young microspore stage in rice plants. XXX. Relation between fertilization and the number of engorged pollen grains among spikelets cooled at different pollen developmental stages. Japanese Journal of Crop Science, 60, 523–528.10.1626/jcs.60.523
- Tranel, D., Knapp, A., & Perdomo, A. (2009). Chilling effects during maize tassel development and the lack of compensational plasticity. Crop Science, 49, 1852–1858.10.2135/cropsci2008.10.0593
- Troyer, A. F. (2004) Persistent and popular germplasm in seventy centuries of corn evolution. In C. W. Smith, et al. (Eds.), Corn: Origin, history, technology, and production (pp. 133–231). Hoboken, NJ: Wiley.