Abstract
Field experiments were carried out in 2014 and 2015 to characterize the associated attributes responsible for dry matter accumulation in high-yielding soybean. We attempted to create a high-yielding environment by introducing narrow-row planting at two planting densities, using a new cultivar ‘Hatsusayaka’ and a current leading cultivar ‘Sachiyutaka’ in an upland experimental field. Dry matter accumulation was assessed in terms of light interception and radiation use efficiency (RUE). Growth analysis was performed to evaluate the crop growth rate (CGR) and the contribution of the net assimilation rate (NAR) and mean leaf area index to CGR. Maximum soybean yields of 590 and 658 g m−2 were obtained for Hatsusayaka and Sachiyutaka, respectively, in 2015 at the high planting density, with the corresponding maximum aboveground dry matter equaling 1463 and 1331 g m−2 and maximum LAI equaling 8.5 and 7.6. Although cumulative intercepted solar radiation was lower than in previous studies, early canopy closure at around the beginning of the flowering stage and very high RUE (1.54 and 1.68 g MJ−2 for Hatsusayaka and Sachiyutaka, respectively) contributed to the high dry matter accumulation. In contrast to the high yield in 2015, continuous excess soil moisture in early August 2014 may have inhibited nodule nitrogen fixation and decreased the nitrogen content, resulting in an extremely high specific leaf area and low leaf greenness, which agrees well with the low NAR during the corresponding period.
Classification:
Abbreviations:
- AGDM: aboveground dry matter
- CGR: crop growth rate
- CIPAR: cumulative intercepted photosynthetically active radiation
- CISR: cumulative intercepted solar radiation
- DAS: days after sowing
- FIPAR: fraction of intercepted photosynthetically active radiation
- LAI: leaf area index
- mLAI: mean leaf area index
- NAR: net assimilation rate
- PAR: photosynthetically active radiation
- RUE: radiation use efficiency; SLA: specific leaf area
Introduction
Soybean (Glycine max [L.] Merrill) products are an essential part of Japan’s traditional dietary culture and are a UNESCO-registered intangible cultural heritage. However, soybean production in Japan (.2 Mt in 2013) was less than 7% of the national demand (MAFF; http://www.maff.go.jp/j/tokei/). The average soybean yield in Japan in 2014, at 169 g m−2 (MAFF, http://www.maff.go.jp/j/tokei/), was only 58% of the values reported in two major soybean-producing nations, the United States and Brazil (FAOSTAT, http://faostat.fao.org/). Japan’s low yield may be attributed to disadvantageous climate characteristics for soybean cultivation, such as a rainy season at the time of sowing, drought stress after the rainy season, and typhoons (Fatichin et al., Citation2013; Matsuo et al., Citation2016). Excess soil moisture due to poor drainage in fields converted from paddy rice frequently inhibits the emergence and growth of soybean, which is a major constraint in Japanese soybean production because more than 80% of the soybean crop is cultivated in converted paddy fields (Matsuo et al., Citation2013; Shimada et al., Citation2012).
On the other hand, a previous report showed that high yield (>500 g m−2) could be obtained in a converted paddy field (Nakaseko et al., Citation1984; Shimada et al., Citation1990; Spaeth et al., Citation1987). These results suggested that soybean’s yield potential in Japan, which represents the yield in the absence of limitations caused by water and nutrient availability, diseases, and weeds, (Evans & Fischer, Citation1999), is potentially high if these and other constraints can be removed. The potential yield of soybean was estimated to reach 800 g m−2 (Specht et al., Citation1999). Very high soybean yields, with values close to the estimated potential yield [795 g m−2; Van Roekel and Purcell (Citation2014)] or higher [920 g m−2; Isoda et al. (Citation2006)] have been reported. However, because there have been so few reports on the yield potential of soybean in Japan, it is necessary to explore the current yield potential under an optimal growth environment using modern cultivars.
Genetic improvement of soybean yield is frequently associated with increased dry matter accumulation rather than with an increased harvest index (Edwards et al., Citation2005; Kumudini et al., Citation2001). Therefore, characterizing the ecophysiological factors responsible for dry matter accumulation is essential for understanding the high-yielding conditions for soybean. Hence, we developed the present study to focus on light interception and utilization by soybean plants because both are key constituents for dry matter accumulation.
Ball et al. (Citation2000) argued that having a short duration before the full canopy cover is achieved (>90% light interception) may increase yield because it shortens the time required after emergence to begin linear biomass accumulation, which Goudriaan and Monteith (Citation1990) is considered to be lost time. Lee et al. (Citation2008) showed that achieving complete ground cover by the R1 [stage in the classification of Fehr and Caviness (Citation1977)] or shortly thereafter is required to achieve maximum yield. However, the standard spatial arrangement of soybean plants in Japan is 10–20 plants m−2, with .60–.80 m between rows, .07–.20 m between plants within the rows, and tilling between the rows. This suggests that planting at narrow row spacing (ca. .3 m between rows) would reduce the time required to achieve full canopy cover compared with planting at the conventional distance between rows (.6–.8 m). This idea was supported by an examination of the effects of the distance between rows at a constant planting density (15 plants m−2); Andrade et al. (Citation2002) showed that seed yield at higher light intensity (about 90%) at R1 in narrowly spaced rows (.19 m) was about 10% greater than that at lower light intensity (about 73%) at R1 in widely spaced rows (.57 m). Equidistant plant spacing in narrow-row planting is beneficial because it minimizes competition between plants and increases light interception starting from the early growth stages (Ball et al., Citation2000; Egli, Citation1994; Nakano et al., Citation2001; Shibles & Weber, Citation1966; Weber et al., Citation1966). Recently, narrow-row planting has been increasing in popularity in Japan due to the potential to improve yield and decrease labor (Furuhata et al., Citation2008; Uchikawa et al., Citation2009).
A key constituent of dry matter accumulation is the plant’s radiation use efficiency (RUE), which is defined as the amount of biomass produced per unit of intercepted solar radiation (Monteith, Citation1977). In their review of RUE, Sinclair and Muchow (Citation1999) reported that RUE for soybean ranged from .66 to 1.26 g MJ−1 and averaged 1.02 g MJ−1. Recently, very high RUE (1.89 g MJ−1) was reported for high-yielding soybean in Missouri, U.S.A. (Van Roekel & Purcell, Citation2014). In Japan, the previously reported high RUE were 1.40 g MJ−1 (Shiraiwa et al., Citation1994), 1.26 g MJ−1 (Nakaseko & Goto, Citation1983), and 1.15 g MJ−1 (Sinclair & Shiraiwa, Citation1993). Because these values are far from the value of 1.89 g MJ−1, it would be useful to re-evaluate the RUE that can be achieved in Japan under an optimal growth environment using modern cultivars.
Kawasaki et al. (Citation2016) demonstrated that Japanese soybean cultivars released from 1971 to 1986 had a lower yield potential than the American cultivars released from 1986 to 2007. This suggested that slow adoption of modern, high-yielding cultivars may be one of the reasons for the low soybean yield in Japan (Hajika, Citation2013). The current leading soybean cultivars in southwestern Japan, Fukuyutaka and Sachiyutaka, were released in 1980 and 2001, respectively. Hatsusayaka, a new soybean cultivar released in 2011, has productivity equal to or higher than that of Sachiyutaka (Saruta et al., Citation2012). Sachiyutaka and Hatsusayaka can be used in narrow-row planting because of their high lodging resistance (Okabe et al., Citation2009), whereas Fukuyutaka is susceptible to lodging and therefore adapts poorly to narrow-row-spacing planting (Furuhata et al., Citation2008). Thus, it would be worthwhile to evaluate the yield potential of Hatsusayaka in an environment capable of providing optimal yield.
In this study, we attempted to create a high-yielding environment for soybean yield in an upland experimental field by means of narrow-row planting at two planting densities to assess the current yield potential of modern cultivars Hatsusayaka and Sachiyutaka. Field experiments were performed in successive years, and an exceptionally high yield and RUE were obtained in the second year. Our objective was to elucidate the dry matter accumulation that can produce the exceptionally high yield and RUE. The dry matter accumulation was characterized as a function of the meteorological environment, light interception, and RUE. Growth analysis was performed to evaluate the crop growth rate (CGR) and the contribution of the net assimilation rate (NAR) and mean leaf area index (mLAI) to CGR. The empirical data obtained from this study will help to characterize the associated attributes for dry matter accumulation that can produce exceptionally high soybean yield in Japan. Although the data that allowed a comparison between cultivars were limited to a single year, the results for Sachiyutaka provide reference values for comparison with other cultivars, as well as to evaluate the effects of planting density on yield and dry matter accumulation.
Materials and methods
Experimental site, cultivars, and field management
Two field experiments were conducted at the Faculty of Agriculture, Kagawa University (34°16′N, 134°7′E), in 2014 and 2015, using two determinate soybean cultivars (Hatsusayaka in both years and Sachiyutaka in 2015) and two planting densities (12.8 and 18.5 plants m−2). The experimental field had a well-drained loam soil.
We established four 1.8 m × 18.0 m ridges for Hatsusayaka in 2014 and six 1.5 m × 18.0 m ridges for both cultivars in 2015, with the ridges running from north to south. Each ridge consisted of six rows in 2014 and five rows in 2015, separated by .3 m in both years. Seeds were sown on 18 July 2014 and 13 July 2015 at .09 m between seeds using a hand-powered sowing machine (HS-120; Mukai Kogyo, Inc., Osaka, Japan). At the V1 stage, seedlings were thinned to a spacing of .18 and .27 m between plants, which corresponded to planting densities of 18.5 plants m−2 (hereafter, D18) and 12.8 plants m−2 (hereafter, D12), respectively. The treatments were arranged in a randomized block design with six replications and areas of 1.8 m × 5.0 m (2014) and 1.5 m × 5.0 m (2015) for each plot.
The field was fertilized with compound fertilizer before sowing at rates of 20, 56, and 56 kg ha−1 of N, P2O5, and K2O, respectively. A herbicide mixture of benthiocarb, pendimethalin, and linuron (Clearturn; Kumiai Chemical Industry Co., Ltd., Tokyo, Japan) was sprayed after sowing to prevent weeds. Plants were sprinkled water several times by a water hose attached with a spray nozzle during the early vegetative stage when the soil moisture seemed deficient.
Measurements
Aboveground material from four plants per plot (for all destructive samples in 2014 and the first two destructive samples in 2015) or from three plants per plot (for the third to the sixth destructive samples in 2015) were collected at 2-week intervals: at 12, 26, 40, 54, 68, and 82 days after sowing (DAS) in 2014 and at 17, 31, 45, 59, 73, and 87 DAS in 2015. We did not sample plants in the border rows so as to avoid edge effects. Leaf greenness of the topmost fully expanded leaf was measured with a SPAD meter (SPAD-502; Konica Minolta Inc., Tokyo, Japan). All leaf blades were then removed and spread in a single layer on a large copy stand, except for dead and yellowed parts, and images were taken with a digital camera. The leaf area per plot was measured from the digital images by means of image analysis using LIA32 software (http://www.agr.nagoya-u.ac.jp/~shinkan/LIA32/).
The dry weight of the plant parts was determined after oven-drying at 80 °C for more than 72 h. Specific leaf area (SLA, cm2 g−1) was calculated by dividing the total leaf area by the total leaf dry weight per plot. The dry weight and leaf area of each plot were converted into values per unit ground area to provide the aboveground dry matter (AGDM, g m−2) and leaf area index (LAI, m2 m−2) based on the number of plants sampled and the planting density. The changes in AGDM (W2 and W1) and LAI (L2 and L1) at two sampling times (t2 and t1) were used to calculate CGR, NAR, and mLAI, as follows:
Air temperature, daily total solar radiation, and daily precipitation were measured at the meteorological station of the Faculty of Agriculture, Kagawa University, which is located adjacent to the experimental field. The 13-year mean values from 2001 to 2013 at the meteorological station were compared with the values in 2014 and 2015.
The fraction of photosynthetically active radiation (PAR) intercepted by the plant canopy (FIPAR) in each plot was measured weekly using the SunScan Canopy Analysis System (Delta-T Devices, Cambridge, UK). This system measures PAR above and below the canopy simultaneously, with a sunshine sensor placed at the center of the experimental field higher than the canopy height and a 1-m probe that contains 64 PAR sensors inserted into the canopy at the ground level, perpendicular to the row direction. The 1-m probe was connected with the sunshine sensor using a 25-m cable and was attached to a handheld computer to store the data. The mean of five measurements per plot was used to calculate FIPAR according to Purcell (Citation2000) and Purcell et al. (Citation2002):
Daily total intercepted solar radiation was calculated based on the daily total solar radiation measured at the meteorological station, multiplied by the FIPAR value measured with the SunScan instrument. FIPAR for days on which no measurements were taken was estimated by means of linear interpolation between the two closest measurements. Cumulative intercepted solar radiation (CISR, MJ m−2) was calculated by summing the daily intercepted solar radiation after emergence of the seedlings in each plot. RUE was determined as the slope of a simple linear regression between AGDM and CISR for each plot using all of the destructive sample data (i.e. six times) during the periods from 12 to 82 DAS in 2014 and from 17 to 87 DAS in 2015.
At maturity, 20 plants from each plot were destructively sampled; this occurred on 27 October 2014 (102 DAS) and 26 October 2015 (106 DAS). Yield (the ‘fine’ seed weight, adjusted to a 15% moisture content) and the yield components were obtained from the number and dry weight of the main stems, branches, nodes, pods (total and fertile), and seeds. Seeds were classified into fine and nonconforming (which includes seeds that were damaged by pests and diseases, that split, or that showed other obvious defects) using a 7.3-mm sieve, and weighed. The number of seeds was calculated from the seed weight divided by the 100-seed weight. Yield, the number of seeds, and the 100-seed weight excluded the non-conforming seeds. Yield data were converted into values per unit ground area based on the number of plants sampled (20 plants) and the planting density.
Statistical analysis
Data for Hatsusayaka were analyzed by means of a two-way ANOVA using the model for a randomized block design with six replications to evaluate the effects of planting density, year, and their interaction on all measured variables. All factors except density were considered to be random effects. For Sachiyutaka, the effects of planting density on all measured variables were evaluated using the t-test. Regression analysis was carried out to quantify the relationships between AGDM and CISR, between CGR and NAR, and between CGR and mLAI. Statistical analysis was performed by JMP statistical software (SAS Institute Japan, Tokyo, Japan).
Results
Plant development and meteorological conditions
Table presents the calendar dates and resulting DAS for CISR at stages R1, R5, and R6. Hatsusayaka began flowering (R1) on 20 August 2014 (33 DAS) and Sachiyutaka and Hatsusayaka began flowering on 17 and 18 August 2015, respectively (35 and 36 DAS). In both years, there was no significant difference in R1 between the two densities. The plants began to set seed (R5) on 18 September 2014 (62 DAS) and 9 September 2015 (58 DAS) and reached the full seed stage (R6) on 22 September 2014 (66 DAS) and 24 September 2015 (71 DAS) in both cultivars and at both densities. The effects of year and density on CISR in Hatsusayaka were both significant, and that of density on CISR was significant in Sachiyutaka. CISR values at stages R1 and R6 were greater in 2015 than in 2014, whereas those at stage R5 were greater in 2014 than in 2015 at both densities. CISR was significantly smaller in D12 than in D18 in all years and cultivars.
Table 1. Dates (month/day and DAS) and CISR values at the R1, R5, and R6 stages for the Hatsusayaka and Sachiyutaka soybean cultivars grown at normal (D12) and dense (D18) densities in 2014 and 2015.
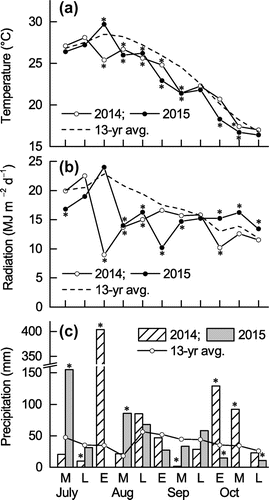
Figure shows the changes in mean temperature, daily total radiation, and daily total precipitation at 10-day intervals during the 2014 and 2015 experimental periods and compares these values with the 13-year means (2001–2013) at the meteorological station. In both years, there were many cloudy and rainy days throughout the experimental period. Almost all mean temperatures from mid-August (R1) to mid-October (R7) in both years were significantly lower than the 13-year mean (Figure (a)). Hereafter, all significant differences in Figure are based on the 95% confidence interval. Daily total solar radiation was significantly lower than the 13-year mean from early August (the vegetative stage) to early October (stage R6 or R7) in 2014, particularly in early August (vegetative stage), when the value was 69% lower than the mean. Daily total solar radiation was significantly lower from mid-August (R1) to mid-September (R5 or R6) in 2015, particularly in early September (R5), when the value was 42% lower than the mean. Daily total solar radiation during October 2014 (R6 or R7 to R8) tended to be lower than the 13-year mean (the difference was only significant in early October), but during October 2015 (R6 or R7 to R8) it was significantly higher than the mean. Precipitation in 2014 was significantly lower than the 13-year mean in late July (during the early vegetative stage) and in mid-September (R5), and it was significantly higher than the mean in early August (vegetative stage) and from early to mid-October (R6 or R7). In 2015, precipitation in mid-July (the early vegetative stage) and mid-August (R5) was significantly higher than the 13-year mean, but precipitation was significantly lower than the mean from mid- to late October (R7 to R8).
Figure 1. Changes in (a) mean temperature, (b) daily total solar radiation, and (c) precipitation at 10-day intervals (i.e. an average of three times per month) to represent the early (E), middle (M), and late (L) parts of the month during the experimental periods in 2014 and 2015. Averages represent the 13-year mean from 2001 to 2013 at the meteorological station adjacent to the study site. Asterisks (*) indicate values outside of the 95% confidence interval for the 13-year mean (i.e. significantly higher or lower values).
Yield and yield components
Seed yields ranged from 381 to 658 g m−2 across all cultivars, years, and densities (Table ). The effects of year and density on seed yield and all yield components of Hatsusayaka were significant, except for the effect of year on the number of fertile pods and the effects of year and density for the 100-seed weight and harvest index. There was no significant year × density interaction for any of the variables. For Hatsusayaka in 2015, seed yield, number of seeds, number of nodes, and AGDM were about 30% higher than the corresponding values in 2014. For Hatsusayaka in D18, seed yield, number of seeds, number of fertile pods, and AGDM were about 17–20% higher than those in D12, and the numbers of nodes and branches were about 30% higher than those in D12. For Hatsusayaka, lodging angles were higher in 2014 than in 2015 at both densities and were higher in D18 than in D12 in both years and cultivars. The effects of year and density on lodging angle were both significant, but in Sachiyutaka, the effect of density was not significant. In Sachiyutaka, the D18 yield components did not differ significantly from the D12 values, except for a significantly higher number of fertile pods, nodes, and branches. The magnitude of the increase in seed yield and other components in Sachiyutaka in D18 compared with that in D12 was similar to that in Hatsusayaka, with one exception: for the number of branches, the increase at D18 in Sachiyutaka was half (15%) of the increase in Hatsusayaka (30%). Although the effect of cultivar on yield was not evaluated statistically because the Sachiyutaka data were limited to a single year, the mean values of seed yield and the 100-seed weight were 12 and 14% higher in Sachiyutaka, and the numbers of fertile pods, nodes, and branches were about 15% lower in Sachiyutaka.
Table 2. Yield, yield components, AGDM, harvest index, and lodging angle at maturity for the soybean cultivars Hatsusayaka and Sachiyutaka grown at normal (D12) and dense (D18) densities in 2014 and 2015.
AGDM, LAI, SLA, and SPAD value
Figure shows the changes of AGDM and LAI for the two cultivars. In Hatsusayaka, AGDM and LAI were higher in 2015 than in 2014 throughout the growth period at both densities and were higher in D18 than in D12 in both years and for both cultivars. In both cultivars, maximum AGDM was achieved at 82 DAS in 2014 and 87 DAS in 2015 (8 October in both years), and maximum LAI was achieved at 68 DAS in 2014 (24 September) and 58 DAS in 2015 (9 September), except that maximum LAI in Sachiyutaka in D18 was at 45 DAS in 2015 (27 August). Maximum AGDM ranged from 867 to 1463 g m−2, and maximum LAI ranged from 4.3 to 8.5 m2 m−2 (Table ). For Hatsusayaka, the effects of year and density on maximum AGDM and maximum LAI were both significant. The mean values of maximum AGDM and LAI for the two densities in 2015 were 40 and 60% larger than those in 2014, and the mean values of maximum AGDM and LAI for the two years were about 23% larger in D18 than in D12. For Hatsusayaka, the effects of year and density on the dates of 80% and 90% FIPAR and the CISR values on those dates were significant (the dates were earlier and CISR was greater in D18), but there was no significant year × density interaction for either parameter. For Sachiyutaka in 2015, neither the date nor the CISR value differed significantly between the two densities.
Figure 2. Changes of (a, b) AGDM and (c, d) LAI for the soybean cultivars (H, Hatsusayaka; S, Sachiyutaka) at normal (D12) and dense (D18) densities in 2014 and 2015. Values are means ± S.E. (n = 6).
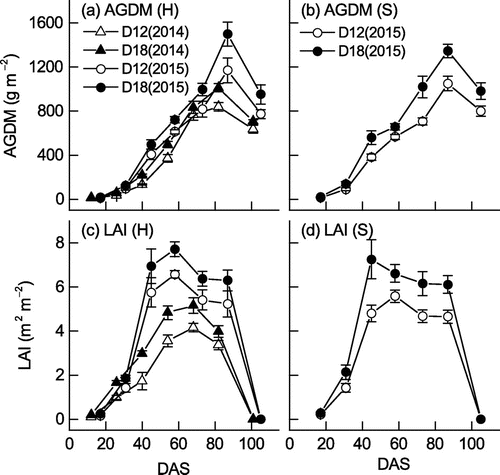
Table 3. Maximum AGDM, maximum LAI, CISR at the last destructive samples (82 DAS in 2014 and 87 DAS in 2015), RUE, and the date (DAS) and CISR values when FIPAR reached 80% and 90%.
For Hatsusayaka, very high values of SLA (570 cm2 g−1 in D12 and 597 cm2 g−1 in D18) were observed at 26 DAS in 2014 (Figure ). Except for those extremes, SLA ranged from 172 to 310 cm2 g−1 across all cultivars, years, and densities. SLA values in 2014 were lower than those in 2015 after 40 DAS. In 2015, the changes in SLA were similar for Hatsusayaka and Sachiyutaka, except for a higher SLA in Hatsusayaka than in Sachiyutaka at 45 DAS. SLA of D18 at 68 DAS in 2014 was higher than that in D12.
Figure 3. Changes in SLA of the soybean cultivars (a) Hatsusayaka and (b) Sachiyutaka grown at normal (D12) and dense (D18) densities in 2014 and 2015. Values are means ± S.E. (n = 6).
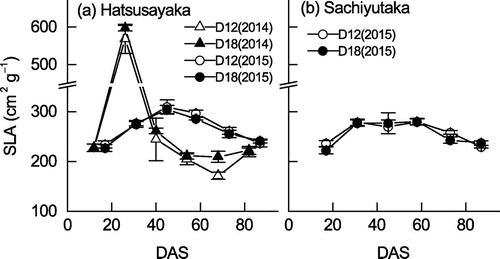
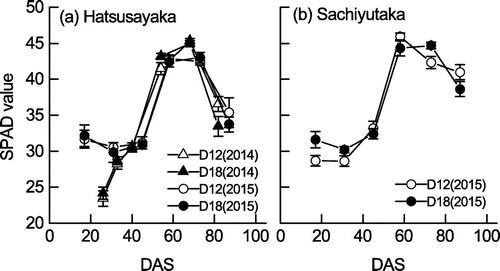
Figure shows the changes in SPAD values. In Hatsusayaka, these changes were similar in 2014 and 2015. In 2014, the mean SPAD value at 26 DAS (13 August) was 23.8 at both densities, which was approximately 20% lower than the mean between 17 DAS (30 July) and 31 DAS (13 August) for both densities in 2015. In 2015, the mean SPAD value for the two densities was 31.9 until 45 DAS (27 August); thereafter, it increased to 42.7, then decreased to 34.6 at 87 DAS (8 October). There were only small differences in SPAD values between planting densities. In 2015, we noted only small differences between cultivars until 80 DAS, but SPAD values at 87 DAS were about 15% higher in Sachiyutaka than in Hatsusayaka.
Figure 4. Changes in SPAD values of the soybean cultivars (a) Hatsusayaka and (b) Sachiyutaka grown at normal (D12) and dense (D18) densities in 2014 and 2015. Values are means ± S.E. (n = 6).
Growth analysis
Figure shows the changes in CGR, NAR, and mLAI. CGR values in D18 tended to be higher than those in D12 throughout most of the study period, but the differences between densities were only significant during the first (12 to 26 DAS) and second (26 to 40 DAS) periods for Hatsusayaka 2014 and the fourth period (54 to 68 DAS) for Sachiyutaka. In 2014, the mean CGR for the two densities in Hatsusayaka during the first period was 2.5 g m−2 d−1, then it increased to a peak (26.2 g m−2 d−1) during the fourth period and decreased to 8.9 g m−2 d−1 during the last period (68 to 82 DAS). For Hatsusayaka in 2015, the mean CGR for the two densities during the first period (17 to 31 DAS) was 6.9 g m−2 d−1; thereafter, it increased greatly to 24.4 g m−2 d−1 during the second period (31 to 45 DAS), decreased further, to 16.6 g m−2 d−1, during the third period (45 to 58 DAS), and then increased greatly again, to 30.6 g m−2 d−1, during the last period (73 to 87 DAS).
Figure 5. Changes of (a, b) CGR, (c, d) NAR, and (e, f) mLAI of the two soybean cultivars (H, Hatsusayaka; S, Sachiyutaka) at normal (D12) and dense (D18) densities in 2014 and 2015. Values are means ± S.E. (n = 6).
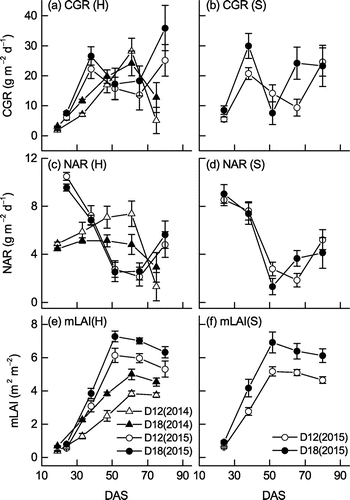
There was no significant differences in NAR between the two densities in both years and both cultivars, except for a significantly higher NAR for Hatsusayaka in D12 than in D18 during the first period (12 to 26 DAS) in 2014. The mean NAR of the two densities for Hatsusayaka during the first period (12 to 26 DAS) in 2014 was 53% lower than that in 2015 (Fig. c). Thereafter, NAR in Hatsusayaka 2014 remained under 5.1 g m−2 d−1 (D18) or increased to 7.4 g m−2 d−1 (D12) until the fourth period (54 to 68 DAS), then decreased greatly during the last period. In contrast to the low NAR values for Hatsusayaka in the first period in 2014, the mean NAR for the two densities in 2015 was largest (10.0 g m−2 d−1) during the first period (17 to 31 DAS), then decreased greatly, to 2.7 g m−2 d−1, during the third period (45 to 58 DAS) and increased to 5.2 g m−2 d−1 during the last period (73 to 85 DAS).
For Hatsusayaka, mLAI values in 2015 were higher than those in 2014 throughout the growth period (Figure (e)). The peak mLAI in 2015 occurred during the third period (45 to 58 DAS) in both cultivars, whereas the peak for Hatsusayaka in 2014 occurred during the fourth period (54 to 68 DAS). For Hatsusayaka in 2014 and Sachiyutaka in 2015, mLAI values in D18 were significantly higher than in D12 throughout the study period. In contrast, mLAI values for Hatsusayaka in 2015 were significantly higher in D18 than in D12 during the first (17 to 31 DAS) and fourth (58 to 73 DAS) periods.
The changes of growth parameters in Sachiyutaka in 2015 were similar to those in Hatsusayaka for the most part (Figure ), but we observed some greater differences between cultivars in D18 than in D12. For instance, the highest CGR in D18 in Sachiyutaka occurred during the second period, versus the last period in Hatsusayaka.
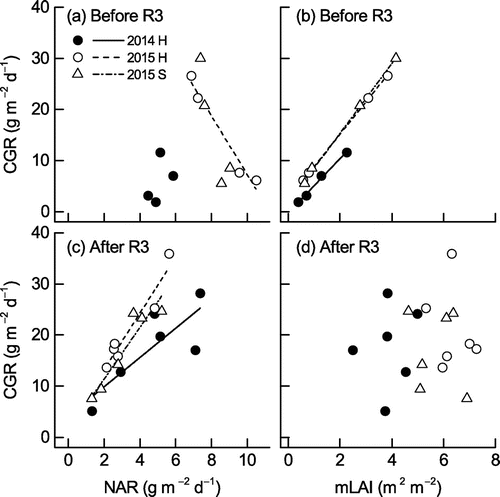
Figure shows the relationships between CGR and NAR and between CGR and mLAI before R3 (Figure (a) and (b)) and after R3 (Figure (c) and (d)). There was a statistically significant relationship between CGR and NAR in Hatsusayaka in 2015 before R3, and CGR before R3 was also significantly related to mLAI in both years and both cultivars. On the other hand, CGR after R3 was significantly related to NAR in both years and both cultivars but was unrelated to mLAI after R3.
Figure 6. Linear regressions for the relationships between CGR and (a, c) NAR and (b d) mLAI before (a, b) and after (c, d) stage R3 for the two soybean cultivars (H, Hatsusayaka; S, Sachiyutaka) in 2014 and 2015. Linear regression equations are presented only for the following statistically significant relationships. Coefficients of determination (R2) are (a) Hatsusayaka 2015: R2 = .968 (p < .05), (b) Hatsusayaka 2014: R2 = .994 (p < .001), Hatsusayaka 2015: R2 = 1.000 (p < .001), Sachiyutaka 2015: R2 = .998 (p < .001), (c) Hatsusayaka 2014: R2 = .671 (p < .05), Hatsusayaka 2015: R2 = .920 (p < .001), Sachiyutaka 2015: R2 = .889 (p < .001). (d) No significant relationships.
Radiation use efficiency
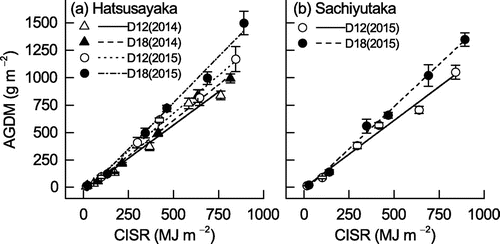
Linear regression analysis was performed for the relationship between AGDM and CISR to calculate RUE (i.e. the slope of the regression) using all AGDM data from the first to the last destructive samples (82 and 87 DAS in 2014 and 2015, respectively, for all cultivars and densities) and the CISR on the corresponding date. (CISR values from the last destructive samples are shown in Table .) There were strong and significant linear relationships between AGDM and CISR in both years, both cultivars, and both densities (Fig. ). The mean RUE calculated in each replicate in both years, cultivars, and densities ranged from 1.21 to 1.68 g MJ−1 and the coefficients of determination for the linear regressions calculated in each plot were all statistically significant (.817 < R2 < .998, .05 < p < .001). The effects of year and density on RUE in Hatsusayaka were statistically significant, and the effect of density on RUE in Sachiyutaka was significant. In Hatsusayaka, the mean RUE of the two densities in 2015 was 21% higher than that in 2014, and the mean RUE of D18 was 16% higher than that of D12 in both years. In Sachiyutaka, RUE was 25% higher in D18 than in D12.
Figure 7. Linear regression relationships between AGDM and CISR for the two soybean cultivars for (a) Hatsusayaka and (b) Sachiyutaka grown at normal (D12) and dense (D18) densities in 2014 and 2015. Values are means ± S.E. (n = 6). Equations and coefficients of determination (R2) are (a) Hatsusayaka: D12 in 2014: y = −34.8 + 1.207x, R2 = .978 (p < .001), D18 in 2014: y = −35.3 + 1.295x, R2 = .995 (p < .001), D12 in 2015: y = −15.4 + 1.381x, R2 = .993 (p < .001), D18 in 2015: y = −66.6 + 1.678x, R2 = .990 (p < .001), (b) Sachiyutaka: D12 in 2015: y = −3.7 + 1.229x, R2 = .986 (p < .001), D18 in 2015: y = −33.5 + 1.541x, R2 = .995 (p < .001).
Discussion
Specht et al. (Citation1999) estimated that soybean’s yield potential could reach 800 g m−2 if the estimated seed yield limit for corn was 2250 g m−2 and the ratio of corn yield to soybean yield was 2.8. In fact, very high soybean yields that approach or exceed this estimated potential yield have been reported: 795 g m−2 (Van Roekel & Purcell, Citation2014) and 920 g m−2 (Isoda et al., Citation2006). The maximum soybean yields in the current study (590 and 658 g m−2 in Hatsusayaka and Sachiyutaka, respectively; Table ) were 18 and 26% lower than that estimated potential yield but were equivalent to or higher than the previously reported high yields in Japan, which ranged from 528 to 649 g m−2 (Nakaseko et al., Citation1984; Shimada et al., Citation1990; Spaeth et al., Citation1987).
One reason for the high yield in the current study is the use of an upland field for the experiment. This is because most of the soybean in Japan is cultivated in converted paddy fields, in which excess soil moisture frequently inhibits plant growth (Matsuo et al., Citation2013; Shimada et al., Citation2012). However, because the abovementioned high soybean yields in Japan (Nakaseko et al., Citation1984; Shimada et al., Citation1990; Spaeth et al., Citation1987) were obtained in converted paddy fields, the field itself is not a limiting factor for yield if soil moisture conditions are adequate. High soil fertility may also contribute to the potential soybean yield in a converted paddy (Nakaseko et al., Citation1984). Hence, the reasons for the high yield in the current study should be explored in terms of climate conditions and dry matter production as a function of light interception and RUE.
Nakaseko et al. (Citation1984) estimated that an LAI of 8.4 is necessary to achieve 1200 kg m−2 of aboveground biomass in soybean based on a linear regression equation for the relationship between AGDM and maximum LAI derived from high-yield experiments. Another high-yield experiment in Japan showed that LAI must be greater than 7.2 to achieve a yield higher than 500 kg m−2 (Shimada et al., Citation1990). Shimada et al. showed that a maximum LAI of 9.1 and nearly 1500 g m−2 of AGDM produced 596 g m−2 of maximum yield. Table shows that the maximum AGDM values in the present study for Hatsusayaka and Sachiyutaka (1463 and 1331 g m−2, respectively) and the maximum LAI values (8.5 and 7.6, respectively) exceed the AGDM and LAI that appear to be required to attain high yield.
The significant relationships between CGR and mLAI before stage R3 and between CGR and NAR after R3 suggest that the differences in CGR between the two densities in each cultivar and each year can be attributed to the differences in mLAI before R3 and in NAR after R3, respectively (Figure ). Significant linear regressions between CGR and mLAI in the early reproductive growth stages and between CGR and NAR in the later reproductive growth stages were consistent with a previous report (Isoda et al., Citation2010). On the other hand, the higher AGDM in Hatsusayaka in 2015 than in 2014 throughout the growth period (Figure ) can be attributed to high CGR during the first period (12 to 26 DAS), second period (26 to 40 DAS), and last period (68 to 82 DAS), which was sustained by high NAR during the corresponding periods and the very rapid increase of LAI that occurred around R1 (26 to 40 DAS, in mid-August; Figure ). Slow CGR and low LAI during an early growth stage in 2014 under low temperatures, low solar radiation, and excess soil water agree with a previous report in which cooler temperatures delayed canopy formation (De Bruin & Pedersen, Citation2009) and a previous report of inhibition of leaf area development by excess soil moisture during an early growth stage that reduced biomass accumulation by decreasing intercepted solar radiation and RUE (Bajgain et al., Citation2015).
In contrast to the high AGDM, LAI, and seed yield of Hatsusayaka in 2015, this cultivar had substantially lower values in 2014 (Tables and , Figure ). Those differences between years can be attributed to differences in the weather conditions (Figure ). Many cloudy and rainy days occurred throughout the growth period in both years, and the temperature and daily total solar radiation from mid-August to mid-September (about R1 to R5) were lower than the 13-year mean. In 2014, western Japan received record-high precipitation, accompanied by low temperatures and short hours of sunshine from the summer to early fall, due to the frequent heavy rain caused by a stationary front and typhoons. The temperatures and solar radiation in early August (the mid- to late vegetative growth stages) in 2014 were particularly low, and this combined with the extremely high precipitation to decrease CGR from the first growth period (12 to 26 DAS) to the second growth period (26 to 40 DAS) (Figures and ). On the other hand, the temperatures and solar radiation were higher than the 13-year mean during the same periods in 2015. There was enough precipitation in mid- to late July to compensate for a lack of rain in early August. These climate conditions may have increased CGR during this period compared with the same period in 2014. Furthermore, the low CGR during the final period in 2014 contrasted with high CGR during the same period in 2015. In October 2014 (about R6 to R8), the mean temperature and daily total solar radiation tended to be lower and precipitation tended to be higher than the 13-year mean, whereas in October 2015, mean temperature and precipitation tended to be lower than the 13-year mean, but daily total solar radiation was higher. These climate differences can explain the differences in CGR during the final period.
Extremely high SLA and low SPAD values until mid-August in 2014 (Figures and ) suggest a possible nitrogen deficiency for the plants. Because low oxygen in the soil due to excess soil moisture can inhibit nodule nitrogen fixation (Shimada et al., Citation2012), the continuous excess soil moisture caused by the record-high precipitation in 2014 may have inhibited nodule nitrogen fixation, thereby decreasing the leaf chlorophyll content and producing low SPAD values. Previously reported significant relationships between specific leaf weight (the reciprocal of SLA) and specific leaf nitrogen content (Shiraiwa et al., Citation1994) support the hypothesis of a nitrogen deficiency in 2014 in the current study, since a higher SLA would result in lower leaf nitrogen content, all else being equal. These hypotheses are supported by the low NAR observed during the first and second growth periods (12 to 40 DAS) in 2014 (Figure ).
Because RUE is the slope of the regression for the relationship between plant dry weight and cumulative intercepted radiation (Sinclair & Muchow, Citation1999), the strong and significant linear relationships found in the present study (Table , Figure ) indicate constant RUE throughout the growth period. The maximum RUE obtained in the current study (1.68 g MJ−1 for Hatsusayaka in 2015) was 17 to 32% higher than previously reported high RUE values in Japan: 1.40 g MJ−1 (Shiraiwa et al., Citation1994), 1.26 g MJ−1 (Nakaseko & Goto, Citation1983), and 1.15 g MJ−1 (Sinclair & Shiraiwa, Citation1993). For comparison, very high RUE values (1.46 to 1.89 g MJ−1) were determined for high-yielding soybean in Missouri, U.S.A. (Van Roekel & Purcell, Citation2014). The high RUE in the current study could have resulted from the vigorous growth from the vegetative stage to the R6 stage despite the relatively low solar radiation during this period (Fig. ). The solar radiation from emergence to maturity totaled 1429 and 1623 MJ m−2 in 2014 and 2015, respectively (Table ), which were 20 to 29% lower than the mean total solar radiation captured from emergence to the R7 stage by nine soybean cultivars studied in Arkansas, U.S.A. (Kawasaki et al., Citation2016). In terms of the cumulative intercepted photosynthetically active radiation (CIPAR) from emergence to the R6 stage, 605 MJ m−2 of CIPAR were required to reach 90% of asymptotic yield (Edwards et al., Citation2005), and CIPAR ranged from 382 to 400 MJ m−2 for new and old soybean cultivars at a planting density of 14.8 m−2 (De Bruin & Pedersen, Citation2009). In contrast, CISR from emergence to the R6 stage in our experiment ranged from 555 to 665 MJ m−2 (= 278–332 MJ m−2 in PAR; Table ), and these values were substantially lower than those in the previous studies.
Table 4. Daily total solar radiation during the experiments in 2014 and 2015, and the 13-year means (2001–2013) at the meteorological station adjacent to the study site.
Achieving complete ground cover by the R1 stage or shortly thereafter is required to achieve maximum yield (Andrade et al., Citation2002; Ball et al., Citation2000; Board, Citation2000; Lee et al., Citation2008; Purcell et al., Citation2002). The light interception in our experiment reached 80% about 1 week after R1 in 2014 and around R1 in 2015, and reached 90% at 10 d after R1 in 2014 and 2–6 d after R1 in 2015 (Tables and ). The dates when both 80 and 90% of FIPAR were achieved in Hatsusayaka were earlier in 2015 than in 2014 and in D18 than in D12. These results suggested that achieving early coverage of the ground in 2015 and in D18 contributed to the greater dry matter accumulation than in 2014 and in D12.
Regardless of early canopy closure and saturation of radiation interception in the current study, the lower incident solar radiation in this region (Table ) decreased CISR (Tables and ). These results suggested that low solar radiation was the major cause of lower CISR, and that achieving vigorous growth despite the lower CISR resulted in high RUE. Furthermore, because increasing diffuse radiation coupled with decreasing irradiance can increase RUE (Sinclair et al., Citation1992), an inherently higher fraction of diffuse radiation in Japan can explain this growth (Sinclair & Shiraiwa, Citation1993). In fact, there were many cloudy and rainy days from mid-August to mid-September in both years, and radiation from R1 to R5 was lower than average (Figure ). In addition, Nakaseko and Goto (Citation1983) found that soybean RUE decreases curvilinearly with increasing light intensity as a result of photosynthetic light saturation. This supports our observation of high RUE in the current study despite the low light intensity.
The significantly higher RUE in D18 than in D12 in both years and both cultivars (Table , Figure ) suggested that RUE in dense stands is higher than that in normal stands. Similar results were reported by Bajgain et al. (Citation2015). On the other hand, Shibles and Weber (Citation1965, Citation1966) demonstrated constant RUE across a wide range of plant population, and Purcell et al. (Citation2002) found that RUE decreased with increasing density. The reason for the inconsistencies between the current and previous studies would be attributed to the narrow range of planting density (12.8 and 18.5 plants m−2 in current study in comparison to the wide range (7 to 135 plants m−2) in Purcell et al. (Citation2002). Considering the RUE decrease with increasing planting densities was about .002 g MJ plant−1 (Purcell et al., Citation2002), the difference of planting density (ca. 6 plants m−2) in this study is too narrow to assess the difference of RUE having the standard error about .08 (Table ). The experiment with a wide range of planting density would be necessary to assess the effect of population density on RUE.
Conclusions
Soybean seed yield as high as 590 and 658 g m−2 was obtained for Hatsusayaka and Sachiyutaka in 2015 in the dense planting. These high yields were associated with conditions that produced the maximum AGDM and LAI. High AGDM throughout the 2015 growth period could be attributed to the high CGR during the early and late growth periods, sustained by high NAR, and by the rapid increase of LAI during the early growth period. This rapid growth was promoted by favorable temperature, solar radiation, and precipitation at key stages. Although CISR in the current study was lower than that in other studies, early canopy closure at around R1 and high RUE promoted high AGDM matter accumulation. The present yield and dry matter accumulation data support the hypothesis that dense planting offers advantages over normal planting in narrow-row planting. In contrast to the high yield achieved in 2015, the low yield and dry matter accumulation in Hatsusayaka in 2014 can be attributed to record-high precipitation of this year, low temperatures, and decreased solar radiation due to frequent heavy rains caused by a stationary rain front and typhoons. Continuous excess soil moisture in early August 2014 may have inhibited nodule nitrogen fixation and thereby decreased the leaf nitrogen content, which was revealed by high SLA and low SPAD values; this agrees well with the low NAR values during the corresponding periods.
Disclosure statement
No potential conflict of interest was reported by the authors.
Acknowledgments
The authors thank the Western Region Agricultural Research Center, National Agriculture and Food Research Organization, for providing the Hatsusayaka seeds and anonymous reviewers for valuable comments.
References
- Andrade, F. H., Calviño, P., Cirilo, A., & Barbieri, P. (2002). Yield responses to narrow rows depend on increased radiation interception. Agronomy Journal, 94, 975–980.10.2134/agronj2002.0975
- Bajgain, R., Kawasaki, Y., Akamatsu, Y., Tanaka, Y., Kawamura, H., Katsura, K., & Shiraiwa, T. (2015). Biomass production and yield of soybean grown under converted paddy fields with excess water during the early growth stage. Field Crops Research, 180, 221–227.10.1016/j.fcr.2015.06.010
- Ball, R. A., Purcell, L. C., & Vories, E. D. (2000). Optimizing soybean plant population for a short-season production system in the Southern USA. Crop Science, 40, 757–764.10.2135/cropsci2000.403757x
- Board, J. (2000). Light interception efficiency and light quality affect yield compensation of soybean at low plant populations. Crop Science, 40, 1285–1294.10.2135/cropsci2000.4051285x
- De Bruin, J. L., & Pedersen, P. (2009). New and old soybean cultivar responses to plant density and intercepted light. Crop Science, 49, 2225–2232.10.2135/cropsci2009.02.0063
- Edwards, J. T., Purcell, L. C., & Karcher, D. E. (2005). Soybean yield and biomass responses to increasing plant population among diverse maturity groups: II. Light Interception and Utilization. Crop Science, 45, 1778–1785.
- Egli, D. B. (1994). Mechanisms responsible for soybean yield response to equidistant planting patterns. Agronomy Journal, 86, 1046–1049.10.2134/agronj1994.00021962008600060021x
- Evans, L. T., & Fischer, R. A. (1999). Yield potential: Its definition, measurement, and significance. Crop Science, 39, 1544–1551.10.2135/cropsci1999.3961544x
- Fatichin, Zheng, S.-H., Narasaki, K., & Arima, S. (2013). Genotypic adaptation of soybean to late sowing in Southwestern Japan. Plant Production Science, 16, 123–130.10.1626/pps.16.123
- Fehr, W. R., & Caviness, C. E. (1977). Stages of soybean development (pp. 1–12). Ames, IA: Cooperative Extension Service, Agriculture and Home Economics Experiment Station, Iowa State University of Science and Technology.
- Furuhata, M., Morita, H., & Yamashita, H. (2008). Performances of dry matter and seed production under narrow-row-dense-planting culture of soybean cultivar, Sachiyutaka, in South-Western Japan. Japanese Journal of Crop Science, 77, 409–417***.
- Goudriaan, J., & Monteith, J. L. (1990). A mathematical function for crop growth based on light interception and leaf area expansion. Annals of Botany, 66, 695–702.10.1093/oxfordjournals.aob.a088084
- Hajika, M. (2013). Future direction and technology of soybean breeding. Agriculture and Horticulture, 88, 513–526*.
- Isoda, A., Mao, H., Li, Z., & Wang, P. (2010). Growth of high-yielding soybeans and its relation to air temperature in Xinjiang, China. Plant Production Science, 13, 209–217.10.1626/pps.13.209
- Isoda, A., Mori, M., Matsumoto, S., Li, Z., & Wang, P. (2006). High yielding performance of soybean in Northern Xinjiang, China. Plant Production Science, 9, 401–407.10.1626/pps.9.401
- Kawasaki, Y., Tanaka, Y., Katsura, K., Purcell, L. C., & Shiraiwa, T. (2016). Yield and dry matter productivity of Japanese and US soybean cultivars. Plant Production Science, 19, 257–266.10.1080/1343943X.2015.1133235
- Kumudini, S., Hume, D. J., & Chu, G. (2001). Genetic improvement in short season soybeans: I. Dry matter accumulation, partitioning, and leaf area duration. Crop Science, 41, 391–398.10.2135/cropsci2001.412391x
- Lee, C. D., Egli, D. B., & TeKrony, D. M. (2008). Soybean response to plant population at early and late planting dates in the mid-south. Agronomy Journal, 100, 971–976.10.2134/agronj2007.0210
- Matsuo, N., Fukami, K., & Tsuchiya, S. (2016). Effects of early planting and cultivars on the yield and agronomic traits of soybeans grown in southwestern Japan. Plant Production Science, 19, 370–380. doi:10.1080/1343943X.2016.11554171-11
- Matsuo, N., Takahashi, M., Nakano, H., Fukami, K., Tsuchiya, S., Morita, S., … Tasaka, K. (2013). Growth and yield responses of two soybean cultivars grown under controlled groundwater level in southwestern Japan. Plant Production Science, 16, 84–94.10.1626/pps.16.84
- Monteith, J. L. (1977). Climate and the efficiency of crop production in Britain. Philosophical transactions of the Royal Society of London Ser B, 281, 277–294.10.1098/rstb.1977.0140
- Nakano, H., Komoto, K., & Ishida, K. (2001). Effect of planting pattern on development and growth of the branch from each node on the main stem in soybean plants. Japanese Journal of Crop Science, 70, 40–46***.
- Nakaseko, K., & Goto, K. (1983). Comparative studies on dry matter production, plant type and productivity in Soybean, Azuki bean and Kidney bean: VII. An analysis of the productivity among the three crops on the basis of radiation absorption and its efficiency for dry matter accumulation. Japanese Journal of Crop Science, 52, 49–58***.
- Nakaseko, K., Nomura, H., Gotoh, K., Ohuma, T., Yoshikatu, A., & Shu, K. (1984). Dry matter accumulation and plant type of the high yielding soybean grown under converted rice paddy fields. Japanese Journal of Crop Science, 53, 510–518***.
- Okabe, A., Takada, Y., & Saruta, M. (2009). Selection of soybean cultivars for narrow- row spacing and non-molding cultivation. Japanese Journal of Crop Science, 78, 85–86**.
- Purcell, L. C. (2000). Soybean canopy coverage and light interception measurements using digital imagery. Crop Science, 40, 834–837.10.2135/cropsci2000.403834x
- Purcell, L. C., Ball, R. A., Reaper, J. D., & Vories, E. D. (2002). Radiation use efficiency and biomass production in soybean at different plant population densities. Crop Science, 42, 172–177.10.2135/cropsci2002.0172
- Saruta, M., Takada, Y., Matsunaga, R., Hajika, M., Takahashi, M., & Komatsu, K. (2012). A new soybean cultivar ‘Hatsusayaka’, with tolerance of delayed leaf senescence and suitability for tofu processing. Bulletin of NARO Western Region Agricultural Research Center, 11, 81–99***.
- Shibles, R. M., & Weber, C. R. (1965). Leaf area, solar radiation interception and dry matter production by soybeans. Crop Science, 5, 575–577.10.2135/cropsci1965.0011183X000500060027x
- Shibles, R. M., & Weber, C. R. (1966). Interception of solar radiation and dry matter production by various soybean planting patterns. Crop Science, 6, 55–59.10.2135/cropsci1966.0011183X000600010017x
- Shimada, S., Hamaguchi, H., Kim, Y., Matsuura, K., Kato, M., Kokuryu, T., Tazawa, J., & Fujimori, S. (2012). Effects of water table control by farm-oriented enhancing aquatic system on photosynthesis, nodule nitrogen fixation, and yield of soybeans. Plant Production Science, 15, 132–143.10.1626/pps.15.132
- Shimada, S., Hirokawa, F., & Myyagawa, T. (1990). Effects of planting date and planting density on a high yielding soybean cultivar grown at drained paddy field in sanyo district. Japanese Journal of Crop Science, 59, 257–264***.
- Shiraiwa, T., Hashikawa, U., Taka, S., & Sakai, A. (1994). Effects of canopy light distribution characteristics and leaf nitrogen content on efficiency of radiation use in dry matter accumulation of soybean cultivars. Japanese Journal of Crop Science, 63, 1–8***.
- Sinclair, T. R., & Muchow, R. C. (1999). Radiation use efficiency. Advances in Agronomy, 65, 215–265.10.1016/S0065-2113(08)60914-1
- Sinclair, T. R., & Shiraiwa, T. (1993). Soybean radiation-use efficiency as influenced by nonuniform specific leaf nitrogen distribution and diffuse radiation. Crop Science, 33, 808–812.10.2135/cropsci1993.0011183X003300040036x
- Sinclair, T. R., Shiraiwa, T., & Hammer, G. L. (1992). Variation in crop radiation-use efficiency with increased diffuse radiation. Crop Science, 32, 1281–1284.10.2135/cropsci1992.0011183X003200050043x
- Spaeth, S. C., Sinclair, T. R., Ohnuma, T., & Konno, S. (1987). Temperature, radiation, and duration dependence of high soybean yields: Measurement and simulation. Field Crops Research, 16, 297–307.10.1016/0378-4290(87)90068-2
- Specht, J. E., Hume, D. J., & Kumudini, S. V. (1999). Soybean yield potential – A genetic and physiological perspective. Crop Science, 39, 1560–1570.10.2135/cropsci1999.3961560x
- Uchikawa, O., Tanaka, K., Miyazaki, M., & Matue, Y. (2009). Effects of planting pattern on growth, yield and nitrogen fixation activity of soybean cropped with late planting and non-intertillage cultivation method in Northern Kyusyu. Japanese Journal of Crop Science, 78, 163–169***.
- Van Roekel, R. J., & Purcell, L. C. (2014). Soybean biomass and nitrogen accumulation rates and radiation use efficiency in a maximum yield environment. Crop Science, 54, 1189–1196.10.2135/cropsci2013.08.0546
- Weber, C. R., Shibles, R. M., & Byth, D. E. (1966). Effect of plant population and row spacing on soybean development and production. Agronomy Journal, 58, 99–102.10.2134/agronj1966.00021962005800010034x