Abstract
Common buckwheat is a self-incompatible, insect-pollinated allogamous plant. This study examined growth and yield of the common buckwheat self-compatible and hybrid lines pollinated with and without flies. Self-compatible ‘IH3’, hybrid ‘IP2/IH3’, and standard self-incompatible ‘Kitawasesoba’ were used in field and pot experiments. Self-compatibility of ‘IH3’ was shown to be of high purity. Approximately 10% segregation of pin plants from ‘IP2/IH3’ was observed. The harvest index of ‘Kitawasesoba’ pollinated without flies was considerably lower than that pollinated with flies in both field and pot experiments. The harvest index values of ‘IH3’ and ‘IP2/IH3’ were hardly affected by the presence or absence of flies. The morphological traits of ‘IH3’ were significantly lower than those of the other two genotypes. Large differences in these traits between ‘IP2/IH3’ and ‘Kitawasesoba’ were not observed in either the field or pot experiments. The seed yield of ‘Kitawasesoba’ pollinated without flies tended to be lower than that pollinated with flies. The seed yield of ‘IP2/IH3’ tended to be higher than that of ‘IH3’. The hybrid line ‘IP2/IH3’ showed a high fertilization rate, which was nearly as high as that of ‘IH3’. Rate of fertilization and percentage of ripe seeds were higher in ‘IH3’ and ‘IP2/IH3’, wherein they were hardly affected by the presence or absence of flies, than in ‘Kitawasesoba’. The ‘IP2/IH3’ hybrid line will be useful for understanding the stable high-yielding ability of self-compatibility common buckwheat.
Classification:
Introduction
The seed yield of common buckwheat (Fagopyrum esculentum Moench) is generally low and unstable with one contributing factor being its complex reproductive system (Ohsawa, Citation1997). Common buckwheat is a self-incompatible, insect-pollinated allogamous plant. The self-incompatibility in common buckwheat is due to its dimorphic flowers, i.e. pin type (long pistil and short stamina) and thrum type (short pistil and long stamina). The fertilization of seeds is usually only accomplished when cross-pollination occurs by insects between pin and thrum plants. However, a few occurrences of seed fertilization through selfing have been observed (Garber & Quisenberry, Citation1927). The genetic difference between pin and thrum plants is due to a single locus S with two alleles S and s that determine the Ss (thrum) and ss (pin) plants (Woo et al., Citation1999). Moreover, seed setting in common buckwheat is dependent on the frequency of flower-visiting insects. Namai (Citation1990) reported that seed setting rates increased by 40 and 100% when the frequencies of flower visits by insects increased one to twofold and fivefold, respectively. In Japan, seed yield per unit area of common buckwheat has gradually decreased year by year over the past two decades. A decrease in insect vector abundance due to the impact of simplified cropland ecosystems, i.e. a decrease in fruit trees and forest around the paddy fields, has been suspected as the reason (Morishita & Suzuki, Citation2012).
From this viewpoint, breeding of self-pollinating common buckwheat has been attempted by introgressing the self-compatibility (Sh) gene of F. homotropicum (Ohnishi, Citation1995), a self-pollinating wild species, into common buckwheat using interspecific hybridization (Campbell, Citation1995). Although a pure line of self-pollinating common buckwheat with a homostyle flower morph has been developed, there is a high possibility of inbreeding depression in this line. Therefore, the self-pollinating common buckwheat variety has been developed only one variety named ‘Buckwheat Norin-PL1’ (Matsui et al., Citation2008). While, Funatsuki et al. (Citation2000) proposed a new method for producing hybrids by applying the self-incompatibility of the pin plant (Japanese patent No. 2958457). This methodology involves inbred development of homostyle (ShSh) and pin (ss) lines. Mukasa et al. (Citation2010) reported that single-cross self-compatible homostyle hybrids (Shs) seeds were obtained using these inbred homostyle and pin lines as pollen and seed parents, respectively. The seed yield of the hybrids showed a 10% increase in yield compared with that of the standard variety ‘Kitawasesoba’ (Mukasa et al., Citation2010). These reports suggest that the development of hybrids is useful in increasing seed yield of common buckwheat. In addition, the outcrossing rate of self-pollinating common buckwheat grown under open pollination conditions has been studied (Mukasa et al., Citation2006). Approximately 10% outcrossing rate was observed in self-compatible common buckwheat (Mukasa, Citation2011). Based on these reports, we hypothesized that the open-pollinated self-compatible hybrid plants may show not only self-pollinating but also certain outcrossing by insect vectors. Therefore, it is important to determine the influence of the presence or absence of flies on the growth and yield of self-compatible hybrid plants. In the present study, we compared the growth and yield of a self-compatible line, hybrid line, and a standard self-incompatible variety as controls in field and pot experiments.
Materials and methods
Plant materials
Three common buckwheat genotypes ‘Kitawasesoba’, ‘IH3’, and ‘IP2/IH3’ were used for field and pot experiments. The self-incompatible variety ‘Kitawasesoba’ is a leading variety in Hokkaido, Japan. A self-compatible line ‘IH3’ was derived from the crossing between a local line ‘Tanno-43’ and material that resulted from backcrossing (four times) of F. homotropicum with the common buckwheat variety ‘Kitayuki’. A self-compatible hybrid line ‘IP2/IH3’ was developed by crossing a pin type (ss) line ‘IP2’ (seed parent) with the self-compatible (ShSh) line ‘H3’ (pollen parent). ‘IH3’ and ‘IP2/IH3’ were developed by the NARO Hokkaido Agricultural Research Center. ‘IP2’ (seed parent of ‘IP2/IH3’) was not used for the present experiment because of its low seed yield, as shown in Table .
Table 1. Seed yield of the plant materials.
Field experiment
The field experiment was conducted at the Memuro Upland Farming Research Station of the National Agriculture and Food Research Organization, Hokkaido Agricultural Research Center (Shinsei, Memuro, Kasai-Gun; 143°03′E 42°55′N). The experimental plot consisted of two ridges, each 1 m long and separated by a distance of 60 cm. The size of each plot was 1.2 m2. On 26 June 2013, the seeds for each genotype were manually planted 5 cm apart in two rows (5 cm separating the rows) on each ridge (one plant per hill). Basal fertilizer of N, P2O5, and K2O at rates of 1.8, 7.2, and 4.2 g m−2, respectively, was applied at a standard level. The experimental plots with and without flies were setup for each genotype. The plots were arranged according to the randomized block design with three replications. In early July 2013, cages (1.8 × 1.2 × 1 m) were placed in all plots and enclosed on all sides with a fine net. The flower morph (pin, thrum, and homostyle) of plants grown in each plot was recorded on 6 August 2013 (flowering time). Thereafter, to avoid cross-pollination, pin and thrum plants grown in plots for ‘IH3’ and ‘IP2/IH3’ were eliminated on 10 August 2013. On the same day, approximately 100 commercial fly pupae were released into each net cage in the plots in which the effect of the presence of the flies was being investigated.
Pot experiment
The pot experiment was conducted in a greenhouse at the Faculty of Bioindustry, Tokyo University of Agriculture (Yasaka, Abashiri; 144°23′E 42°96′N). Eighteen unglazed pots (ϕ 24 cm) filled with commercial garden soil containing 0.374, 1.485, and 0.242 of N, P2O5, and K2O, respectively, were prepared in the greenhouse and randomly arranged with three replications (one pot per plot). On 4 July 2013, 12 seeds for each genotype were planted in each pot. Thereafter on 25 July 2013, a fine net was used to enclose each pot with the help of a strut. The flower morph (pin, thrum, and homostyle) of plants grown in each plot was recorded on 18 August 2013 (flowering time). On the same day, number of plants was decreased to six plants per pot by eliminating the pin and thrum plants grown in pots for ‘IH3’ and ‘IP2/IH3’. For ‘Kitawasesoba’, number of plants was decreased to three pin and three thrum plants per pot (six plants in one pot). On the same day, approximately 30 commercial fly pupae were released into each pot enclosed by fine net. Pots were watered as required.
Measurements
Main stem length, number of nodes on the main stem, and number of primary branches of plants grown in each plot were recorded at maturity for 10 and 3 plants in the field and pot experiments, respectively. Plant samples were harvested and oven-dried at 80 °C for 48 h to measure the above-ground dry weight and seed yield, which was determined by hand-threshing the samples. Yield components, i.e. the number of bloomed florets, fertilization rate, percentage of ripe seeds, and 100-seed weight, were measured according to the methodology of Kasajima and Itoh (Citation2011). Briefly, fertilization rate was determined by dividing the number of fertilized florets by the number of bloomed florets. The percentage of ripe seeds was determined by dividing the number of ripe seeds by the number of fertilized florets. The number of bloomed florets was calculated from the seed yield and other yield components according to Kasajima and Itoh (Citation2011). However, the number of bloomed florets of ‘Kitawasesoba’ in the pot experiment could not be calculated because fertilization rate of some ‘Kitawasesoba’ plants pollinated without flies in the pot showed a value of 0. Yield-related characteristics were also recorded for 10 and 3 plants in the field and pot experiments, respectively. The characteristics evaluated in the present study of pollination with and without flies were compared using two-way analysis of variance. All data analyses were performed using the programs Excel (Microsoft Co., U.S.A.) and Excel statistics (Esmi Co. Ltd., Japan).
Results and discussion
The ratios of thrum to pin to homostyle flower morphs of ‘Kitawasesoba’, ‘IH3’ and ‘IP2/IH3’ in the field and pot experiments before the numbers of plants were eliminated are shown in Figure . In both field and pot experiments, the ratio of pin to thrum plants of ‘Kitawasesoba’ was nearly 1:1. Our data were consistent with those of previous studies that reported the percentage of pin and thrum plants in a population of standard self-incompatible common buckwheat as almost 50% (Eghis, Citation1925; Garber & Quisenberry, Citation1927). In the pot experiment, the percentage of homostyle plants in ‘IH3’ was 100%, whereas in the field experiment, the percentage was nearly 100%. These results showed that the self-compatibility of ‘IH3’ was of high purity. The percentages of homostyle plants of ‘IP2/IH3’ in the field and pot experiments were 84.3 and 87.7%, respectively. On the other hand, the percentages of pin plants of ‘IP2/IH3’ in the field and pot experiments were 9.4 and 12.3%, respectively. The hybrid line ‘IP2/IH3’ was developed by crossing the inbred homostyle with pin lines. Therefore, approximately 10% segregation of pin plants from ‘IP2/IH3’ was confirmed in the present study. However, in the field experiment, 6.3% of thrum plants were observed in ‘IP2/IH3’. While in the pot experiment, no thrum plants were observed in ‘IP2/IH3’. Although the cause of appearance of thrum plant in the field experiment was unknown, the off-type plants, such as pin and thrum plants from IP2/IH3 were eliminated to prevent adverse effects in the present experiment.
Figure 1. Ratio of flower morphs of ‘Kitawasesoba’, ‘IH3’, and ‘IP2/IH3’ in the field (A) and pot experiments (B). ■Homomorphic flower type;
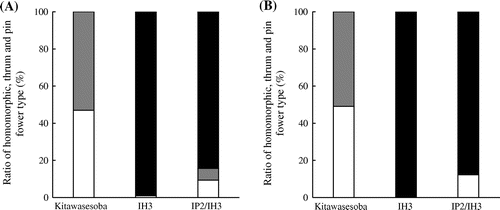
Table shows growth-related traits of each genotype pollinated with and without flies in the field and pot experiments. There were significant differences in harvest index values in the field experiment and number of primary branches in the pot experiment among the treatments. In particular, the harvest index value of ‘Kitawasesoba’ pollinated without flies was considerably lower than that pollinated with flies in both the field and pot experiments. In contrast, the harvest index values of ‘IH3’ and ‘IP2/IH3’ were hardly affected by the presence or absence of flies. There were significant differences in harvest index, main stem length, and number of primary branches among the genotypes in the field experiment, whereas there were significant differences in the numbers of nodes on the main stem and primary branches among the different genotypes in the pot experiment. These morphological traits were significantly lower in ‘IH3’ than in the other two genotypes. It has been demonstrated that the growth of self-compatible common buckwheat plant was inferior to that of open-pollinated standard variety (Matsui et al., Citation2008; Mukasa, Citation2011). In the present study also, the inbreeding depression of ‘IH3’ was observed because of its poor morphological traits such as main stem length. In addition, large differences in these traits were not observed between ‘IP2/IH3’ and ‘Kitawasesoba’ in both the field and pot experiments. It has been revealed that seed yield of hybrid common buckwheat was superior to that of a leading variety (Mukasa et al., Citation2010). These results suggest that the growth of hybrid ‘IP2/IH3’ has practical ability regardless of the presence or absence of flies. The feasibility of common buckwheat hybrids will increase by examining its combining ability in the future.
Table 2. Growth-related traits in each genotype pollinated with and without flies.
Table shows the seed yield and its components in each genotype pollinated with and without flies in the field and pot experiments. No significant differences among the treatments and genotypes in seed yield were observed in the field and pot experiments. However, the seed yield of ‘Kitawasesoba’ pollinated without flies tended to be lower than that pollinated with flies. Fertilization rate of ‘Kitawasesoba’ pollinated without flies was also significantly lower than that pollinated with flies in the pot experiment. In the field experiment, there were significant differences among genotypes in the number of bloomed florets, fertilization rate, and percentage of ripe seeds, whereas a significant difference in fertilization rate existed among genotypes in the pot experiment. The fertilization rates of ‘Kitawasesoba’ were considerably lower than those of the other two genotypes in the field and pot experiments. From these results, the drastic decline in the seed yield of self-incompatible common buckwheat pollinated without flies might be attributed to the decrease in harvest index and fertilization rate. With respect to hybrid line, the seed yield of ‘IP2/IH3’ tended to be higher than that of ‘IH3’; however, this difference was not statistically significant. In addition, we have confirmed that the seed yield of ‘IP2’ (another parent of IP2/IH3) was considerably lower than that of ‘IP2/IH3’ and ‘Kitawasesoba’ (Table ). Mukasa et al. (Citation2010) reported that the seed yield of hybrid common buckwheat was superior to that of an open-pollinated standard variety ‘Kitawasesoba’ by 10%. These findings suggest that increased seed yield of ‘IP2/IH3’ may be relevant to the understanding of heterosis breeding of common buckwheat. The present study also revealed that the hybrid line ‘IP2/IH3’ showed a high fertilization rate of a value similar to that of ‘IH3’. Moreover, ‘IH3’ and ‘IP2/IH3’ showed not only higher fertilization rates but also higher percentages of ripe seeds than ‘Kitawasesoba’. On the other hand, fertilization rate and percentage of ripe seeds in ‘IH3’ and ‘IP2/IH3’ were hardly affected by the presence or absence of flies. The 100-seed weight showed no significant differences among the treatments and genotypes. Therefore, it was clarified that the high seed yield, fertilization rate, and percentage of ripe seeds observed in ‘IP2/IH3’ were not affected by flies. However, further studies will be required to examine whether outcrossing or selfing are promoted by insect vector, because the hybrid line ‘IP2/IH3’ may show certain outcrossing.
Table 3. Seed yield and its components in each genotype pollinated with and without flies.
The present findings indicated that the growth and yield values of ‘IP2/IH3’ were greater than those of ‘IH3’, whereas those of ‘IH3’ and ‘IP2/IH3’ were hardly affected by the presence or absence of flies. Therefore, it was considered that the advantages of ‘IP2/IH3’ will be maintained even in the face of decrease in insect vector abundance. Furthermore, higher growth and yield in ‘IP2/IH3’ was achieved by not only higher fertilization rates but also higher percentages of ripe seeds. The present results indicate that the use of the self-compatible line as a parent for developing the hybrid common buckwheat is practical. The hybrid line ‘IP2/IH3’ will be useful in understanding the stable high-yielding ability of self-compatible common buckwheat.
Disclosure statement
No potential conflict of interest was reported by the authors.
References
- Campbell, C. (1995). Inter-specific hybridization in the Genus Fagopyrum, Proceedings, Sixth International Symposium on Buckwheat, Shinshu, 255–263.
- Eghis, S. A. (1925). Experiments on the drawing up of a method of buckwheat breeding. Bulletin of Applied Botany, Genetics, and Plant Breeding, 14, 235–251.
- Funatsuki, H., Agatsuma, M., & Maruyama, W. (2000) JP. Patent No. 2958457. Tokyo, JP. Ebisu International Patent Office.
- Garber, R. J., & Quisenberry, K. S. (1927). Self-fertilization in buckwheat. Journal of Agricultural Research, 34, 185–190.
- Kasajima, S., & Itoh, H. (2011). Effect of shading during different growth phases on yield parameters of common buckwheat cv. Kitawasesoba in the northern region of Japan. Fagopyrum, 28, 43–46.
- Matsui, K., Tetsuka, T., Hara, T., & Morishita, T. (2008). Breeding and characterization of a new self-compatible common buckwheat parental line, ‘Buckwheat Norin-PL1’. Bulletin of the National Agricultural Research Center for Kyushu Okinawa Region, 49, 11–17*.
- Morishita, T., & Suzuki, T. (2012). Buckwheat varieties and trend of cultivation in Hokkaido, Japan. Research Journal of Food and Agriculture, 35, 9–12**.
- Mukasa, Y., Suzuki, T., & Honda, Y. (2010). A methodology for heterosis breeding of common buckwheat involving the use of the self-compatibility gene derived from Fagopyrum homotropicum. Euphytica, 172, 207–214.10.1007/s10681-009-0047-9
- Mukasa, Y. (2011). Studies on new breeding methodologies and variety developments of two buckwheat species (Fagopyrum esculentum Moench and F. tataricum Gaertn.). Research bulletin of the National Agricultural Research Center for Hokkaido Region, 195, 57–114*.
- Mukasa, Y., Suzuki, T., & Honda, Y. (2006). The degree of outcrossing in open pollinated self-pollinating buckwheat. Report of the Hokkaido Branch, the Japanese Society of Breeding and Hokkaido Branch, the Crop Science Society of Japan, 47, 65–66**.
- Namai, H. (1990). Pollination biology and reproductive ecology for improving genetics and breeding of common buckwheat, Fagopyrum esculentum (1). Fagopyrum, 10, 23–46.
- Ohnishi, O. (1995). Discovery of new Fagopyrum species and its implication for the studies of evolution of Fagopyrum and of the origin of cultivated buckwheat. Proceedings of the 6th International Symposium On Buckwheat, 175–190.
- Ohsawa, R. (1997). Present status and future of buckwheat breeding in Japan. Ikushugaku Saikin no Shinpo, 39, 55–58**.
- Woo, S. H., Adachi, T., Jong, S. K., & Campbell, C. G. (1999). Inheritance of self-compatibility and flower morphology in an inter-specific buckwheat hybrid. Canadian Journal of Plant Science, 79, 483–490.10.4141/P98-117