ABSTRACT
Temperatures and solar radiation during ripening critically affect grain appearance in rice (Oryza sativa L.). Climatic factors to induce chalky grains were analyzed under the experimental conditions of high-temperature and shading treatment and also under the ambient condition in a high-temperature-prone region of Japan. The frequency of white-back (WB) and basal-white (BW) grains correlated with temperature and solar radiation, whereas that of milky-white (MW) grains was not correlated, suggesting that complicated climatic factors are involved in the formation of MW grains. Further investigation was carried out to identify the parameters that distinguish perfect and MW grains grown in high-temperature versus those grown in low-solar-radiation conditions. As reported previously, the chalk phenotypes in the transverse section of the MW grains were quite different between environments: oval-shaped chalk for MW grains grown in low-solar-radiation condition and center chalk for MW grains grown in high-temperature condition. Grain hardness and amylopectin chain-length distribution did not explain the difference in MW grains between environments. MW grains subjected to high temperatures had a lower protein content without a consistent reduction in the single-grain weight, whereas those from the low-solar-radiation condition had a lower amylose content with a consistent reduction in the single-grain weight, when compared with perfect grains that developed in either environmental condition. Overall, our results suggest that MW grains are formed through different physiological mechanisms with altered starch and protein synthesis under high-temperature and low-solar-radiation conditions.
Abbreviations: BW: basal-white; DAH: days after heading; DP: degree of polymerization; MW; milky-white WB: white-back
Introduction
Rice (Oryza sativa L.) grain quality is an extremely important criterion for deciding whether to release a new variety and for determining its market value. Producing high-quality rice provides an additional opportunity for generating higher export revenues (Fitzgerald et al., Citation2009). Grain quality encompasses a number of parameters including grain shape, amylose content, aroma, and other attributes (Calingacion et al., Citation2014), but grain chalkiness is commonly used as an indicator of market value since a high frequency of chalky grains degrades grain appearance and affects head rice yield (Lyman et al., Citation2013; Zhao et al., Citation2013).
Environmental stresses such as high temperatures and low solar radiation (shaded) during ripening increase the frequency of chalky grains (Morita et al., Citation2016). Depending on the zone where the chalky phenotype is formed in the starchy endosperm, chalky grains are categorized as milky-white (MW), white-back (WB), or basal-white (BW) types (Tashiro & Wardlaw, Citation1991). In common to all types, loosely packed amyloplasts are observed in the chalky zone of endosperm cells, whereas tightly packed amyloplasts occur in the transparent, starchy endosperm cells (Tashiro & Wardlaw, Citation1991). A certain proportion of MW grains degrades the eating quality of steamed rice in Japonica Group cultivars (Kim et al., Citation2000; Wakamatsu et al., Citation2007), whereas WB and BW grains have little effect on eating quality (Wakamatsu et al., Citation2007). The frequency of perfect grains decreases when the average temperature during the first 20 days after heading (DAH) exceeds 26–27 °C (Morita et al., Citation2016; Wakamatsu et al., Citation2007; Wakamatsu et al., Citation2008). A temperature-controlled chamber experiment indicated that high temperatures during both the day and night increase the frequency of chalky grains (Morita et al., Citation2002). Exposure of maturing panicles to high temperatures induces the formation of chalky grains, whereas high-temperature treatments of stems and leaves do not (Morita et al., Citation2004). Reducing the number of source leaves during ripening also increases the frequency of MW grains even under optimal temperatures (Tsukaguchi et al., Citation2011). Under high-temperature stress, wide genetic variation in the frequency of WB and BW grains is observed, although it is difficult to identify varietal differences in MW grains among Japonica Group varieties (Iida et al., Citation2002). Tsukaguchi et al. (Citation2012) reported a different chalk phenotype in the transverse section of MW grains. These results suggest that complicated mechanisms underlie the formation of MW grains.
High temperatures during ripening significantly change starch properties, grain hardness, and the protein content of grain. For instance, high temperatures during ripening also reduce the amylose (Cooper et al., Citation2008; Yamakawa et al., Citation2007) and protein (Lin et al., Citation2010; Yamakawa et al., Citation2007) contents. The frequency of amylopectin chains with degrees of polymerization (DPs) from around 20–27 to over 44 increases in grain produced under high-temperature conditions (Umemoto et al., Citation1999; Yamakawa et al., Citation2007). Chalky grains have a lower hardness value than perfectly translucent grains (Singh et al., Citation2003). Most of these findings result from tests conducted in temperature-controlled chambers from which detailed classifications of the types of chalky grains produced were not reported. Thus, the important parameters related to the production of MW grains under high-temperature field conditions are still a matter of debate. In addition, systematic analyses of these parameters have yet to be conducted to determine how low solar radiation in field conditions influences chalky grain production.
The aim of this study is to identify characteristics of MW grains produced under high-temperature and low-solar-radiation conditions under the field conditions. This study reports the results from two experimental sites examined over multiple years. At one experimental site (Tsukuba), rice plants were treated with high temperature and low solar radiation during ripening using the greenhouse and shading nets, respectively. With regard to the parameters that were different between conditions in Tsukuba site, the reproducibility of the findings was confirmed in high-temperature-prone regions at another experimental site (Mie Prefecture), by evaluating grains produced during three high-temperature years and three low-solar-radiation years from a wide range of climatic conditions that occurred over 16 years (2001–2016). By comparing the perfect grains and MW grains produced under each environmental condition, we attempted to understand whether different physiological responses underpinned the formation of MW grains produced by high-temperature and low-solar-radiation stresses.
Materials and methods
Plant materials
‘Koshihikari’, a Japonica Group rice cultivar, was grown in two locations in Japan – Tsukuba and Mie Prefecture (for details, see the following two sections). Paddy fields were kept flooded except for mid-summer drainage, and chemical sprayings were applied appropriately to keep the plants healthy.
Tsukuba experimental site – high-temperature and shading treatments during ripening
‘Koshihikari’ was grown in the experimental field of the Institute of Crop Science, Tsukuba, Japan (36°01ʹN, 140°06′E), in 2007, 2009, and 2010. Seeds were sown in seedling nursery boxes, and three seedlings were transplanted per hill with a spacing of 30 cm and 15 cm (planting density of 22.2 hills m−2). Nitrogen fertilizer of 4 g N m−2 and 2 g N m−2 was applied as a basal dressing at three days before transplanting and as a top dressing at 21 days before heading, respectively. Phosphorus and potassium (each 4 g m−2) were applied as basal dressings. The canopy was open without treatment until 6 DAH when full heading was complete to avoid stress-induced spikelet sterility at flowering stage. At 6 DAH, rice plants were subjected to high-temperature and shading stress by covering the canopies growing inside greenhouses with a transparent vinyl film (0.1 mm thickness, Achilles, Tokyo, Japan) and a 60% shading net (SL-60, silver color; Taiyou net, Osaka, Japan), respectively. Both sides of the greenhouse were manually opened and closed during the day (07:00–18:00 h) so as not to exceed the lethal temperature inside the greenhouse on the hot days. Both sides of the greenhouse were fully open during 18:00–07:00 h to avoid excessively humid conditions in the canopy during the night. The transparent vinyl film reduced solar radiation by 15%. Temperatures inside the greenhouses were monitored using thermo-sensors and data loggers (SK-L200TH II, Sato Keiryoki Mfg., Tokyo, Japan). The lower half of the canopy was not covered by a shading net to keep the air temperature similar to that of the control condition at the canopy level. Temperatures for the control condition were obtained from the weather station monitored by the Institute of Agro-Environmental Sciences (NIAES), NARO, Tsukuba, Japan, at the same geographical location as the experimental field. High-temperature and shaded treatments were continued for 13 days until 19 DAH; the greenhouse and shading net were removed thereafter. The plot sizes for the control, high-temperature, and shaded treatments were 25 m2 each. Experiments in 2007 had one replication, whereas experiments in 2009 and 2010 had three replications with three independent blocks for each treatment. The dates for 50% heading were 1 August in 2007, 2 August in 2009, and 28 July in 2010. At maturity, 22 plants (corresponding to 0.99 m2) were harvested per replication. The filled grains were sorted with a 1.8-mm mesh sieve and used for the subsequent grain-quality analyses. In 2007, the appearance of filled grains was evaluated by a Rice Grain Quality Inspector (RS-2000X, Shizuoka Seiki, Shizuoka, Japan) without subsequent visual inspection. In 2009 and 2010, the appearance of filled grains was evaluated by a Rice Grain Quality Inspector (model RGQI20A, Satake, Hiroshima, Japan), followed by visual inspection to sort filled grains into perfect grains, MW (including white-cored) grains, WB grains, and BW grains.
Mie Prefecture experimental site – rice growth in a high-temperature-prone region
‘Koshihikari’ was grown in the experimental field of the Mie Prefectural Agricultural Station, Matsusaka, Japan (34°38′N, 136°29′E), from 2001 to 2016. Mie Prefecture is a high-temperature-prone region in Japan. Four seedlings were transplanted per hill with a planting density of 20.8 hills m−2. Nitrogen fertilizer, 3 g N m−2, was applied as a basal dressing, and 2 g N m−2 was applied twice as a top dressing at 18 days and 10 days before heading. Phosphorus and potassium (3 g m−2 each) were applied as basal dressings. The dates for 50% heading were recorded every year, 125 plants (corresponding to 6.0 m2) were harvested, and yield and yield components were determined. Filled grains were sorted with a 1.7-mm mesh sieve. Grain appearance was visually inspected, and the percentages of perfect grains, MW grains, and WB and BW (WB+ BW) grains were determined. Subsequently, center- or oval-type chalky shape was determined by making a transversal section from 30 MW grains per year in the selected three high-temperature and low-solar-radiation years. Temperatures and solar radiation at this geographical location were obtained from the Agro-Meteorological Grid Square Data developed by NIAES/NARC of NARO (Ohno et al., Citation2016).
Photographs of grains
After photographing intact grains, median transverse sections of MW grains grown in the high-temperature and low-solar-radiation conditions were made with a sharp razor blade. The sections were viewed with a stereo microscope (SZX12, Olympus, Tokyo, Japan) and photographed with a digital camera (E-330, Olympus, Tokyo, Japan).
Amylose, protein content, amylopectin chain-length distribution, and grain hardness
Perfect and MW grains (40–100 grains) for each replication were milled to fine powders (MM300, Retsch GmbH, Haan, Germany) after measuring the single-grain weight and moisture content. Polished rice powders were used to determine the apparent amylose content by the iodine absorption method (Juliano, Citation1971, ISOCitation6642-1) in grains harvested from both experimental sites. For the determination of grain protein content by the Dumas combustion method (CitationAACCI Method 46–30.01), polished rice powder was also used to measure the nitrogen content by a nitrogen analyzer (JM-3000CN, J-SCIENCE LAB, Kyoto, Japan) at the Tsukuba site and by a nitrogen analyzer (NCH Analyzer, Sumika Chemical Analysis Service, Tokyo, Japan) at the Mie Prefecture site. Thereafter, values were converted to the protein content by multiplying by a rice-unique coefficient (5.95). Amylopectin chain-length distribution was determined based on the method of Fujita et al. (Citation2001) with slight modification. Grain hardness was measured using a Tensipresser (MyBoy System, Taketomo Electric Inc., Tokyo, Japan). Briefly, a single brown rice grain was placed on the plate of the instrument and the plunger force was set to 2.0 mm/s during compression of the grain. The maximum compression force, a parameter derived from a force–distance curve, was denoted as grain hardness
Statistical analyses
Significance of the means between treatments was tested by analysis of variance (ANOVA) followed by Tukey–Kramer’s test using STATISTIX ver. 9.0 (Analytical Software). Significant differences in the single-grain weight and amylose and protein contents of perfect and MW grains grown in the same treatments were analyzed by a t-test.
Results
Climatic conditions during ripening
Climatic conditions during ripening at the Tsukuba site are shown in . Under control (ambient) conditions, the average temperature throughout the ripening stage was 26.9 oC, 24.0 °C, and 28.5 °C in 2007, 2009, and 2010, respectively. Average daily solar radiation was 18.9 MJ m−2, 16.3 MJ m−2, and 20.6 MJ m−2 in 2007, 2009, and 2010, respectively. The daily average temperature during the high-temperature treatment (6–19 DAH) was 0.5–1.9 °C higher in the high-temperature condition than in the control condition due to the higher daytime temperatures, but not the nighttime temperatures () because the sides of the greenhouse were open at night to avoid excessive humidity conditions (see Tsukuba Experimental Site in the Materials and Methods section).
Table 1. Climatic conditions during ripening and temperature treatment of Tsukuba site.
At the Mie Prefecture site, the daily average temperature and average daily solar radiation for 16 years were 27.6 °C and 18.7 MJ m−2, respectively (Supplementary Table S1). The daily average temperature and average daily solar radiation during ripening ranged from 25.7 °C (2003) to 28.6 °C (2010) and from 15.1 MJ m−2 (2003) to 21.1 MJ m−2 (2008), respectively. Climatic conditions during ripening at the Mie Prefecture site were diverse across all 16 years. Both maximum and minimum air temperatures positively contributed to the daily average temperature, unlike in the Tsukuba site (Supplementary Figure 1A, B).
Grain appearance
Under the control conditions at the Tsukuba site, the frequency of perfect grains was 53.9%, 79.6%, and 42.0% in 2007, 2009, and 2010, respectively (). The frequency of perfect grains was lower by 30.2%, 21.7%, and 20.5% in the high-temperature condition than for the control condition in 2007, 2009, and 2010, respectively, with significant differences in 2009 and 2010. The shaded treatment increased the frequency of perfect grains by 13.8% (2007) and 10.2% (2010) compared to the control condition with a significant difference in 2010. High-temperature treatment significantly increased the frequency of MW grains, whereas the shaded treatment only slightly increased the MW grain frequency compared to the control condition for each year. In 2009, WB grains were at a negligible frequency, whereas approximately 20% of the grains were classified as WB and BW grains in the control condition of 2010. A significant reduction in the frequency of WB grains was observed in the shaded condition compared to the other two conditions in 2010.
Table 2. Grain appearance in 2007, 2009, and 2010 at the Tsukuba site.
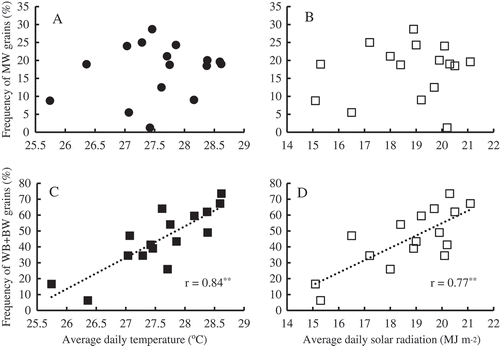
For grain produced at the Mie site, the frequencies of MW grains and WB+ BW grains ranged from 1.2% (2007) to 28.7% (2006) and from 6.3% (2009) to 73.5% (2010), respectively (Supplementary Table S1). The frequency of MW grains was not correlated with the daily average temperature () or the average daily solar radiation () during ripening, whereas the frequency of WB+ BW grains was highly correlated with the daily average temperature () and was correlated with the average daily solar radiation ().
Figure 1. Relationship between the frequency of chalky grains and climatic conditions during ripening at the Mie Prefecture site (2001–2016). A–B: MW grains. C–D: WB+BW grains.
**Significant at the 1% level
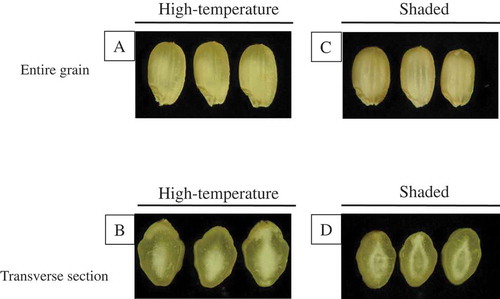
The chalky phenotype of MW grains grown in high-temperature and shaded (low solar radiation) conditions
In 2007 at the Tsukuba site, MW grains were collected from high-temperature and shaded conditions to see whether differences in the chalk phenotype existed. Intact MW grains looked similar among those grown at high-temperature and shaded conditions (). Median transversal sections of MW grains, however, revealed a critical difference in the chalk phenotype. The chalky material occupied the center zone of the starchy endosperm in grain produced in the high-temperature condition (), whereas the chalkiness was observed at the edge of the oval-shaped endosperm, and the inner center zone was transparent in grains produced in the shaded condition ().
Figure 2. Chalky phenotypes in intact grains (A, C) and transverse sections of grains (B, D) grown under high-temperature and shaded conditions at the Tsukuba site (2007).
For grains harvested at the Mie Prefecture site, the frequencies of MW grains collected in 2008 and 2009 were similar at approximately 19–20%, but the climatic conditions of this period varied in terms of the daily average temperature and average daily solar radiation (Supplementary Table S1). Compared to the 16-year average, the daily average temperature was higher in 2008, whereas the average daily solar radiation was lower in 2009 (Supplementary Table S1). The chalk phenotypes of transverse sections of MW grains in 2008 (representative of a high-temperature year, Supplementary Fig. S2A) and 2009 (representative of a low-solar-radiation year, Supplementary Fig. S2B) were similar to those grown in both the high-temperature () and shaded () conditions at the Tsukuba site, but the chalky oval size was much smaller in MW grains from the Mie Prefecture site (Supplementary Fig. S2D) than those from the Tsukuba site ().
Grain hardness between perfect and MW grains
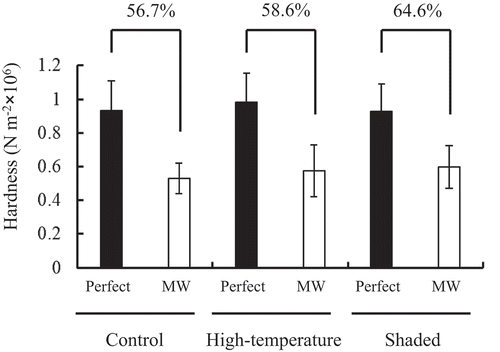
The physical hardness of perfect and MW grains from control, high-temperature, and shaded conditions was examined in 2007 in grain samples produced at the Tsukuba site. The hardness of MW grains was significantly lower than that of perfect grains produced in all conditions. There seemed to be no clear difference in hardness between perfect and MW grains between the treatments ().
Figure 3. Grain hardness of perfect and MW grains grown in control, high-temperature, and shaded conditions at the Tsukuba site (2007). Values are the mean± standard deviation (n = 10 grains). Values above the bridge between perfect grains and MW grains in each condition indicate the ratio of MW to perfect grains.
Amylopectin chain distribution between perfect and MW grains
Differences in amylopectin chain-length distribution between perfect and MW grains from each treatment were investigated for grains produced in 2009 and 2010 at the Tsukuba site (Supplementary Figure S3). MW grains from the control condition in 2009 were not investigated because of their very low frequency (less than 2.0%) (). Under the high-temperature conditions of 2009, the ratio of amylopectin chain lengths was greater in perfect grains for 11≤ DP≤ 20, whereas the ratio was greater in MW grains for 20≤ DP≤ 35 (Supplementary Figure S3A). Under the shaded conditions of 2009, the ratio of amylopectin chain lengths was greater in perfect grains for 9≤ DP≤ 14, whereas the ratio was greater in MW grains for 21≤ DP (Supplementary Figure S3A). These tendencies measured in 2009 were not observed under either the high-temperature or shaded conditions in 2010 (Supplementary Figure S3B). Thus, consistent results were not obtained for the amylopectin chain-length distribution between years.
Comparison of single-grain weight and amylose and protein contents between perfect and MW grains
The single-grain weight and amylose and protein contents were measured using perfect grains and MW grains produced in 2009 and 2010 from each treatment used at the Tsukuba site. Note that the chalk phenotype of the transverse section of MW grains grown in high-temperature and shaded conditions in those years was identical to the transverse section of MW grains in . MW grains from the control condition in 2009 were not investigated because of their very low frequency (less than 2.0%) (). Single-grain weight was lower for MW grains than for perfect grains grown in high-temperature and shaded conditions in both years. The amylose content was significantly lower in MW grains than in perfect grains from the shaded condition, whereas it was higher in MW grains than in perfect grains from the high-temperature condition in both years (). The protein content of MW grains was lower than that of perfect grains grown in the high-temperature condition in both years, whereas the protein content was higher in MW grains than in perfect grains grown in the shaded condition in both years ().
Table 3. Comparison of single-grain weight and amylose and protein contents between perfect and MW grains of Tsukuba site.
For grains produced at the Mie Prefecture site, the single-grain weight and amylose and protein contents were also investigated using perfect grains and MW grains (). From the 16-year environmental dataset, 2002, 2008, and 2010 were selected as representative of high-temperature years, whereas 2003, 2009, and 2014 were selected as representative of low-solar-radiation years. The frequency of MW grains grown during the high-temperature years was consistent at approximately 19–20% (Supplementary Table S1). The daily average solar radiation was slightly lower in 2011 (ranking 14) than in 2014 (ranking 13), but the frequency of MW grains was much higher in 2014 (25.0%) than in 2011 (5.5%) concomitantly with the higher grain yield and spikelet number per area (Supplementary Table S1). We selected the grains from 2014 rather than 2011 for analyzing MW grains. The daily average temperature in the high-temperature years was 0.9 °C higher than that for the 16-year average (Supplementary Table S1). The average daily solar radiation in the low-solar-radiation years was lower by 15.0% than the 16-year average (Supplementary Table S1). Note that WB grains with a slight chalkiness were used as examples of perfect grains in 2008 and 2010 for this measurement since the frequency of perfect grains was only 1.0% (Supplementary Table S1). Single-grain weight was similar or somewhat higher in MW grains than for perfect grains grown in high-temperature years, whereas it was greatly lower in MW grains than perfect grains grown in low-solar-radiation years (). The amylose content was significantly lower in MW grains than in perfect grains grown in the averaged value of low-solar-radiation years, whereas a clear tendency was not observed for grains grown in high-temperature years (). The protein content was significantly lower in MW grains than perfect grains in the averaged value of high-temperature years, whereas the protein content was higher in MW grains than for perfect grains grown in low-solar-radiation years although the differences were not significant (). Ratio of center- and oval-type MW grains was 0.97 and 0.88 on the average of high-temperature and low-solar-radiation years, respectively.
Table 4. Comparison of single-grain weight and amylose and protein contents between perfect and MW grains of Mie Prefecture site.
Discussion
Recent fluctuations in climate, such as high temperatures and low solar radiation, deteriorate rice grain appearance (Morita et al., Citation2016). High temperatures during both the day and night increase the frequency of chalky rice grains (Morita et al., Citation2002). Temperature treatments in a greenhouse at the Tsukuba site were not able to increase sufficiently the night temperature (). Another field experimental site was established in Mie Prefecture, a high-temperature-prone region, where both maximum and minimum temperatures, which are generally obtained during the day and night, respectively, were correlated with the average daily temperature (Supplementary Figure S1). With such an experimental design at two sites and multiyear rice cultivation, we measured the phenotypic, physical, and compositional differences of MW grains grown under high-temperature and low-solar-radiation (shaded) stresses.
At the Tsukuba site, high temperature and the shaded treatment significantly changed the grain appearance (). High temperature treatment during 6–19 DAH increased the daily average temperature by 0.5–1.5 °C (), resulting in a significant reduction in the frequency of perfect grains (). The frequency of MW grains increased under high-temperature conditions both in 2009 and in 2010 (). The shaded treatment significantly increased the frequency of MW grains in 2009, whereas the frequency of MW grains was similar to that grown under control conditions in 2010 (). To discover the key climatic factor(s) for inducing the formation of MW grains, the relationship between temperature and solar radiation and the frequency of MW grains were analyzed under a wide range of climatic conditions that occurred in Mie Prefecture throughout the 16 years (). The data indicated little correlation between climatic conditions and the frequency of MW grains (); in some cases, a lower range of average daily temperature (< 27.0 °C) and a higher range of solar radiation (> 18.0 MJ m−2) resulted in the high frequency (> 15%) of MW grains. In contrast, high frequencies (> 30%) of WB and BW grains were found when the average daily temperature exceeded 27.0 °C (), and high solar radiation also induced high frequencies of WB and BW grains (). High temperatures over 27 °C have been reported to increase the frequency of WB and BW grains (Wakamatsu et al., Citation2007). Furthermore, the simultaneous stresses of intense radiation and high temperatures exacerbate the frequencies of WB and BW grain formation since solar energy promotes increases in panicle temperature (Wakamatsu et al., Citation2008). Our findings agreed with the studies of Wakamatsu et al. (Citation2008). The induction of MW grains, however, could not be explained by temperature and solar radiation. Comparison of data from 2011 and 2014 could be one example. The daily average temperature and solar radiation were similar between these two years, but a great difference in the frequency of MW grains was observed (Supplementary Table S1). It is reported that increased spikelet number per area results in a high frequency of MW grains as well as larger grain yield (Fukushima et al., Citation2017). The high frequency of MW grains in 2014 is possibly due to the greater spikelet number than 2011. These results suggest that a complicated mechanism is involved in the formation of MW grains under field conditions.
High-temperature and low-solar-radiation conditions made a great difference in the chalk phenotype observed in the transverse section of the MW grains: a chalky region formed at the center zone of the starch endosperm in grains grown in high-temperature conditions (; Supplementary Fig. S2B), whereas the chalk phenotype formed in an oval shape in the middle zone of the starchy endosperm under low-solar-radiation condition (; Supplementary Fig. S2D). These differences in chalk phenotype were basically consistent with the observation of Tsukaguchi et al. (Citation2012). Starch accumulation in developing rice endosperm is asynchronous. Active accumulation of starch starts at the center zone and proceeds to the outer zone of the starchy endosperm (Hoshikawa, Citation1968). The chalky phenotype is due to aberrant amyloplast development (Ishimaru et al., Citation2009; Tashiro & Wardlaw, Citation1991; Yamakawa et al., Citation2007). We hypothesize that high-temperature stress and shading stress affected starch accumulation during the early and middle storage phases, respectively. The difference in the degree of low-solar-radiation stress between the Tsukuba site and the Mie Prefecture site might result in the chalky oval size (; Supplementary Fig. S2D).
Furthermore, we attempted to identify the parameter(s) that can distinguish MW grains induced by high-temperature stress with those formed resulting from low-solar-radiation stress. Compositional and physical differences were examined using perfect and MW grains obtained from either the high-temperature or shaded conditions at the Tsukuba site (). Grain hardness and amylopectin chain-length distribution could not explain the differences in the MW grains between high-temperature and shaded conditions (; Supplementary Fig. S3). Single-grain weight was lower in MW grains than in perfect grains under both high-temperature and shaded conditions at the Tsukuba site, whereas that trend was not true in the high-temperature years at the Mie Prefecture site (). The amylose content showed a clear tendency; MW grains had a lower amylose content than perfect grains under the low-solar-radiation conditions at both sites throughout all years examined (). Granule-bound starch synthase (GBSS) is responsible for amylose synthesis in rice (Itoh et al., Citation2003). Reduced activity of GBSS during ripening could be one of the key factors in the formation of MW grains under low-solar-radiation stress. Furthermore, the lower single-grain weight of MW grains compared with perfect grains grown under low-solar-radiation conditions () potentially indicates lower activities of other enzymes related to carbohydrate metabolism, not only GBSS, in MW grains. In contrast, the amylose content of MW grains grown in the high-temperature condition was higher than that for perfect grains at the Tsukuba site, and this tendency was also true for two of three high-temperature years at the Mie Prefecture site (). Very interestingly, MW grains had a lower protein content than perfect grains under the high-temperature conditions at both sites (). This result suggests the involvement of reduced protein synthesis in the formation of MW grains under high-temperature stress. The protein content was higher in MW grains than perfect grains under low-solar-radiation conditions at both sites ().
In this field study, we identified differences in the chalk phenotype of transverse sections of MW grains between grains grown in high-temperature and low-solar-radiation stresses (; Supplementary Fig. S2). Compositional and physical differences were also compared. Such an approach provides direct evidence for the parameter(s) that is (are) substantially involved in the formation of MW grains under high-temperature or low-solar-radiation stress. Our results indicated the involvement of reduced amylose and protein contents in the formation of MW grains under low-solar-radiation and high-temperature stress, respectively. The causal relationship between such compositional differences and different chalky phenotypes in the transverse sections of MW grains is still elusive. The chalky phenotype has been attributed to loosely packed amyloplasts resulting from aberrant starch synthesis (Tashiro & Wardlaw, Citation1991). Therefore, greater attention has been paid to the physiological and molecular changes in carbon metabolism during the formation of MW grains under high-temperature stress. Recent reviews, however, have documented the need to learn how altered inter-relationships between starch and storage protein biosynthesis affect the formation of the chalky phenotype under high-temperature conditions (Sreenivasulu et al., Citation2015), based on the evidence that accumulation of some types of storage proteins is reduced by high-temperature stress (Lin et al., Citation2010; Yamakawa et al., Citation2007). The physiological and molecular mechanisms for how protein and starch syntheses are affected during the formation of MW grains under high-temperature and low-solar-radiation stress, respectively, must be further investigated.
PPS2018_037RP-File005.pptx
Download MS Power Point (858.7 KB)PPS2018_037RP-File004.docx
Download MS Word (19 KB)Acknowledgments
Our thanks go to Dr. M. Yoshimoto (NIAES/NARO) for providing the meteorological dataset for the Tsukuba site. TI thanks Mr. T. Najiwara for his field management.
Disclosure statement
No potential conflict of interest was reported by the authors.
Supplementary material
Supplementary data can be accessed here.
Additional information
Funding
Related Research Data
References
- AACCI method 46-30. (1999). Crude Protein in Cereal.
- Calingacion, M., Laborte, A., Nelson, A., Resurreccion, A., Concepcion, J. C., Daygon, V. D., … & Manful, J. (2014). Diversity of global rice markets and the science required for consumer-targeted rice breeding. PLOS ONE, 9, e85106.
- Cooper, N. T. W., Siebenmorgen, T. J., & Counce, P. A. (2008). Effects of nighttime temperature during kernel development on rice physicochemical properties. Cereal Chemistry , 85, 276–282.
- Fitzgerald, M. A., McCouch, S. R., & Hall, R. D. (2009). Not just a grain of rice: The quest for quality. Trends in Plant Science, 14, 133–139.
- Fujita, N., Hasegawa, H., & Taira, T. (2001). The isolation and characterization of a waxy mutant of diploid wheat (Triticum monococcum L.). Plant Science, 160, 595–602.
- Fukushima, A., Ohta, H., Yokogami, N., & Tsuda, N. (2017). Effects of nitrogen topdressing time on growth, yield, grain appearance and eating quality of the rice varieties bred by Tohoku agricultural research center. Japanese Journal of Crop Science, 86, 7–14. *
- Hoshikawa, K. (1968). Studies on the development of endosperm in rice, 11. Development of starch granules in endosperm tissue. Proceedings of the Crop Science Society of Japan, 37, 207–216. *
- Iida, Y., Yokota, K., Kirihara, T., & Suga, R. (2002). Comparison of the occurrence of kernel damage in rice plants grown in a heated greenhouse and in a paddy field of high-temperature year. Japanese Journal of Crop Science, 71, 174–177. *
- Ishimaru, T., Horigane, A. K., Ida, M., Iwasawa, N., San-Oh, Y. A., Nakazono, M., … Yoshida, M. (2009). Formation of grain chalkiness and changes in water distribution in developing rice caryopses grown under high-temperature stress. Journal of Cereal Science, 50, 166–174.
- ISO 6642-1. (2015). Rice -Determination of amylose content-Part 1: Reference method.
- Itoh, K., Ozaki, H., Okada, K., Hori, H., Takeda, Y., & Mitsui, T. (2003). Introduction of Wx transgenic into rice wx mutants leads to both high- and low-amylose in rice. Plant Cell Physiology, 44, 473–480.
- Juliano, B. O. (1971). A simplified assay for milled rice amylose. Cereal Science Today, 16, 334–338.
- Kim, S. S., Lee, S. E., Kim, O. W., & Kim, D. C. (2000). Physicochemical characteristics of chalky kernels and their effects on sensory quality of cooked rice. Cereal Chemistry, 77, 376–379.
- Lin, C. J., Li, C. Y., Lin, S. K., Yang, F. H., Huang, J. J., Liu, Y. H., & Lur, H. S. (2010). Influence of high temperature during grain filling on the accumulation of storage proteins and grain quality in rice (Oryza sativa L.). Agricultural and Food Chemistry, 58, 10545–10552.
- Lyman, N. B., Jagadish, K. S. V., Nalley, L. L., Dixon, B. L., & Siebenmorgen, T. (2013). Neglecting rice milling yield and quality underestimates economic losses from high-temperature stress. PLOS ONE, 8, e72157.
- Morita, S., Shiratsuchi, H., Takahashi, J., & Fujita, K. (2002). Effect of high temperature on grain ripening in rice plants. -Comparison of the effects of high night temperatures and high day temperatures. Japanese Journal of Crop Science, 71, 102–109. *
- Morita, S., Shiratsuchi, H., Takahashi, J., & Fujita, K. (2004). Effect of high temperature on grain ripening in rice plants. -Analysis of the effects of high night and high day temperatures applied to the panicle and other parts of the plant. Japanese Journal of Crop Science, 73, 77–83. *
- Morita, S., Wada, H., & Matsue, Y. (2016). Countermeasures for heat damage in rice grain quality under climate change. Plant Production Science, 19, 1–11.
- Ohno, H., Sasaki, K., Ohara, G., & Nakazono, K. (2016). Development of grid air temperature and precipitation data compiled from observed, forecasted, and climatic normal data. Climate in Biosphere, 16, 71–79. *
- Singh, N., Sodhi, N. S., Kaur, M., & Saxena, S. K. (2003). Physico-chemical, morphological, thermal, cooking and textural properties of chalky and translucent rice kernels. Food Chemistry, 82, 433–439.
- Sreenivasulu, N., Butardo, V. M., Misra, G., Cuevas, R. P., Anacleto, R., & Kishor, P. B. K. (2015). Designing climate-resilient rice with ideal grain quality suited for high-temperatures. Journal of Experimental Botany, 66, 1737–1748.
- Tashiro, T., & Wardlaw, I. F. (1991). The effect of high temperature on kernel dimensions and the type and occurrence of kernel damage in rice. Australian Journal of Agricultural Research, 42, 485–496.
- Tsukaguchi, T., Ohashi, K., Sakai, H., & Hasegawa, T. (2011). Varietal difference in the occurrence of milky white kernels in response to assimilate supply in rice plants (Oryza sativa L.). Plant Production Science, 14, 111–117.
- Tsukaguchi, T., Yamamura, T., Inoue, H., Nakagawa, H., Murakami, K., & Kita, E. (2012). The response of the occurrence of milky white kernels with different cross-sectional patterns of chalkiness in the endosperm to grain-filling temperature and to assimilate supply in Koshihikari. Japanese Journal of Crop Science, 81, 267–274. *
- Umemoto, T., Nakamura, Y., Satoh, H., & Terashima, K. (1999). Differences in amylopectin structure between two rice varieties in relation to the effects of temperature during grain-filling. Starch, 51, 58–62.
- Wakamatsu, K., Sasaki, O., Uezono, I., & Tanaka, A. (2007). Effects of high air temperature during the ripening period on the grain quality of rice in warm regions of Japan. Japanese Journal of Crop Science, 76, 71–78. *
- Wakamatsu, K., Sasaki, O., Uezono, I., & Tanaka, A. (2008). Effects of the amount of nitrogen application on occurrence of white-back kernels during ripening of rice under high-temperature conditions. Japanese Journal of Crop Science, 77, 424–433. *
- Yamakawa, H., Hirose, T., Kuroda, M., & Yamaguchi, T. (2007). Comprehensive expression profiling of rice grain filling-related genes under high temperature using DNA microarray. Plant Physiology, 144, 258–277.
- Zhao, X., & Fitzgerald, M. (2013). Climate change: Implications for the yield of edible rice. PLOS ONE, 8, e66218.
- *In Japanese with English summary