ABSTRACT
To elucidate the effects of maturity and the stem growth habit on the planting density-dependent branching plasticity of soybean cultivars, we studied the branch traits of 12 cultivars or lines planted at different densities (8.3, 16.7, and 22.2 plants m−2) in Sapporo (2012) and Ebetsu (2013). The 12 cultivars and lines consisted of three determinate cultivars from Hokkaido, three indeterminate cultivars from the northern US, and near-isogenic lines with the backgrounds of Canadian, US, and Japanese cultivars exhibiting diverse stem growth habits. We investigated the relationship between the maturity or stem growth habit and branching plasticity, which was calculated based on the ratios of different branch traits under sparse and dense planting conditions. The use of the ratios of the total branch length and the number of nodes per branch under sparse and dense planting conditions as a measure of branching plasticity revealed varietal differences across years. For the determinate and indeterminate cultivars in both years, branching plasticity was positively correlated with the number of days until stage R5 (onset of seed filling), which is when branches cease to elongate. Comparisons of Japanese and US cultivars and near-isogenic lines for the Dt1 gene (mediating the stem growth habit) indicated that the branching plasticity of indeterminate cultivars and lines is greater than that of determinate cultivars, with a large variation among backgrounds and cultivars. The results of this study imply that branching plasticity is greater in late-maturing soybean cultivars. Moreover, the indeterminate growth habit substantially enhances branching plasticity.
GRAPHICAL ABSTRACT

Introduction
A previous study evaluated the effects of planting density on the branching of Japanese and US soybean cultivars from 2009 to 2012 (Agudamu et al., Citation2016). The data revealed varietal differences in the branching response to planting density, and a branching plasticity score (ratio of the total branch length under sparse and dense planting conditions) was proposed as an indicator of the magnitude of the branching response to various planting densities. Soybean varieties with high branching plasticity are able to maintain stable yields, even when the number of main stem nodes per unit area is low because they produce more branches to compensate for the limited production of main stem nodes.
Excluding soybean cultivars grown in Hokkaido, 80% of the soybeans cultivated in Japan are produced in fields converted from rice paddies (Kato et al., Citation2013; Ministry of Agriculture, Forestry and Fisheries Minister’s Secretariat Statistics Division, Citation2009), which are often poorly drained. In most of the soybean fields converted from rice paddies, ridging is applied during the cultivation period to prevent lodging due to typhoons and to prevent flood damage. The ridging of soybean fields is completed by setting the row spacing to 60–90 cm (Shimada, Citation2011). Such low planting densities (10 or fewer plants m−2) decreases the number of nodes per unit area, which decreases the seed yield (Takahashi, Citation2011). In southwestern Japan, soybean planting tends to coincide with the rainy period (Okabe, Citation2011; Takahashi, Citation2011). The shortened branching period associated with this late planting often results in a suboptimal number of nodes and decreased yield (Okabe, Citation2011; Shimada et al., Citation1990; Takahashi, Citation2011). In northern Japan, the detrimental effects of cold weather on soybean cultivation include poor vigor, plant damage, and delayed maturation (Tsuchiya, Citation1986). Regarding the poor vigor due to cold weather, low temperatures during the early vegetative growth stage suppress branch elongation, which contributes to the low node number and decreased yield (Sanbuichi & Tsuchiya, Citation1976). Problems with seeder machines can also result in missing plants, leading to patches with low node number per unit area.
In environments, such as those described above, where a suboptimal number of nodes is due to the conditions during the vegetative growth period, yield can be stabilized by conferring branching plasticity to the main cultivars, which will enable compensatory branch elongation during the mid-vegetative growth stage. Conferring high branching plasticity to the main soybean cultivars grown in different regions of Japan will reduce the yield instability associated with the fluctuations in the node number per unit area, ultimately increasing seed yield. The degree of branching plasticity depends on the duration of the branch elongation period. Thus, the branching plasticity of late-maturing varieties is generally higher than that of early-maturing varieties. According to Agudamu and Shiraiwa (Citation2015), because the branch elongation period continues through stage R5 in indeterminate cultivars and through stage R3 in determinate cultivars, the branching plasticity is expected to be higher in indeterminate cultivars than in determinate cultivars.
To the best of our knowledge, there are no published studies regarding the relationship between branching plasticity and stem growth habit or maturity that examined multiple cultivars and inter-annual variability. However, cultivars adapted to sparse planting densities have been reviewed in terms of the flexibility in the planting date and for decreasing seed costs by lowering the sowing rate (Rigsby & Board, Citation2003). Lueschen and Hicks (Citation1977) and Herbert and Litchfield (Citation1982) reported the varietal differences in responses to planting densities among indeterminate US soybean cultivars. The ability to compensate for potential yield losses by increasing branching is important for the stability of soybean yield at various plant densities. Moreover, branching is determined by two components, total dry matter weight and the ratio of the branch dry matter weight to total weight (Carpenter & Board, Citation1997a), which affect the varietal differences in branching (Board, Citation2000; Carpenter & Board, Citation1997b). According to Board and Kahlon (Citation2013), there is a large varietal difference in the branching of late determinate cultivars under sparse planting conditions. However, the effects of maturity and the stem growth habit on the branching due to planting densities have not been fully characterized.
The objective of this study was to clarify the effects of maturity and the stem growth habit on the branching plasticity of soybean cultivars grown at various plant densities. We investigated the relationship between branching plasticity and the maturity or stem growth habit of 12 cultivars or lines cultivated with various plant densities. Experiments were conducted at the National Agricultural Research Center for Hokkaido Region (NARCH) in Sapporo (2012) and at Rakuno Gakuen University (RGU) in Ebetsu (2013).
Materials and methods
Cultivars and lines
The first experiment was completed in 2012 at NARCH, whereas the second experiment was completed in 2013 at RGU. Three Hokkaido soybean cultivars (‘Toyomusume’, ‘Toyoharuka’, and ‘Yuzuru’) and three northern US soybean cultivars (‘Jack’, ‘Athow’, and ‘LD00-3309ʹ) were analyzed in both experiments (). The three Hokkaido cultivars are determinate types, whereas the three US cultivars are indeterminate types. Additionally, the following three pairs of near-isogenic lines (NILs) related to stem growth habit were tested in both experiments: ‘Harosoy-dt1ʹ and ‘Harosoy-Dt1ʹ (Canadian cultivar), ‘Williams-dt1ʹ and ‘Williams-Dt1ʹ (US cultivar), and ‘ST-dt1ʹ and ‘ST-Dt1ʹ [cross between US and Japanese cultivars (‘Stressland’ × ‘Tachinagaha’, ST)] (–). The notations ‘-dt1ʹ and ‘-Dt1ʹ indicate determinate and indeterminate types, respectively.
Table 1. Precipitation, daily average air temperature and solar radiation during the growing seasons.
Table 2. Seed yield and the related characteristics in Hokkaido and nothern US cultivars.
Table 3. Effect of stem growth habit of Canadian cultivar ‘Harosoy’, northern U.S. cultivar ‘Williams’ and the ST line (‘Stressland’ × ‘Tachinagaha’) on seed yield and yield-related traits in near-isogenic lines.
Table 4. Branching plasticity values based on the ratios of branch traits of Hokkaido and northern US cultivars under sparse and dense planting conditions.
Table 5. Effects of the stem growth habit on branching plasticity values based on the ratios of branch traits of near-isogenic lines under sparse and dense planting conditions.
‘Toyomusume’ is known for its stable and high yields, whereas ‘Toyoharuka’ is cultivated in Hokkaido because of its lodging resistance. Both of these cultivars have similar maturity dates [maturity group (MG) I]. ‘Yuzuru’ is cultivated in southern Hokkaido and produces large seeds. ‘Jack’ is a soybean cultivar traditionally cultivated in the midwestern states of the US. ‘Yuzuru’ and ‘Jack’ were selected because of their similar maturity (MG II). ‘Athow’ and ‘LD00-3309ʹ are new high-yielding cultivars grown in the midwestern states of the US (MG III and IV, respectively).
Planting densities, field management practices, and measurements
For both experiments, 12 soybean cultivars and lines were grown at three plant densities (60 cm row width × 7.5, 10, and 20 cm intra-row spacing). For experiments evaluating the effects of planting density, ensuring the range of densities is appropriate is critical. Thus, we considered the results of studies by Lee et al. (Citation2008) and Parvez et al. (Citation1989), which indicated that the greatest variation in soybean yield occurs over a planting density range of 8–25 plants m−2. Therefore, we applied planting densities that fall within this range.
When the first trifoliolate leaves emerged, the seedlings were thinned to one plant per hill. Seedlings were transplanted from adjacent hills to vacant hills to maintain the completeness of the plant community. A fertilizer containing 2 g m−2 N (as ammonium sulfate), 12 g m−2 P2O5 (as calcium superphosphate), and 8 g m−2 of K2O (as potassium sulfate) was applied according to standard practices in Hokkaido. In both experiments, weeds were removed manually up until the flowering stage. Seeds were treated with thiamethoxam to protect plants from pests (e.g., seed-corn flies, aphids, and cutworms). Additionally, metalaxyl and fludioxonil emulsions were applied in late July and early August on an as-required basis to protect the plants from Japanese beetles and aphids.
To evaluate the soybean yield, 60 medium-sized plants were manually harvested at maturity (stage R8) from each plot. The harvested plants were air-dried for 3 or 4 weeks, after which the nodes and pods on the main stems and branches were counted. Following a manual threshing, the number of seeds on the main stems and branches as well as the 100-seed weight were recorded. Seed yield was adjusted to 130 g kg−1 moisture content.
The total branch length per plant was measured for 20 plants. The total branch length per branch elongation period up to stage R5 and the average branch length (total branch length per branch number) were calculated to compensate for the differences in the maturity and branch numbers of soybean cultivars, respectively. Specifically, the branch elongation period was longer and the branch lengths were greater for the late-maturing cultivars than for the early-maturing cultivars and indeterminate types (i.e., US cultivars), which continue to develop branches until stage R5 (Agudamu & Shiraiwa, Citation2015; Hanway & Weber, Citation1971).
Statistical analyses
A split-plot design was used in both experiments. The plant densities were distributed in the plots, whereas the determinate cultivars (Hokkaido) and indeterminate cultivars (US) were included in the subplots (10.5 m2). The experiments were conducted with three replicates. An analysis of variance (ANOVA) was completed to evaluate the differences in the effects of treatments on the seed yield.
Branching plasticity was evaluated based on the branch performance traits, seed yield, pod number, node number, and total length under sparse planting conditions relative to the same characteristics under dense planting conditions (i.e., ratios) as described by Agudamu et al. (Citation2016). We hereafter refer to these ratios as branching plasticity values. In addition to the ANOVA, Pearson correlation coefficients for the variables were also calculated and the Tukey-Kramer test was used for the multiple comparisons of the means for the treatments. The homogeneity of error variances was verified by the F max test.
Results
Climatic conditions
The temperature and rainfall during the experiments were higher than the corresponding average annual values (). Specifically, the mean temperatures were 1.6°C higher than average in 2012 and 1.2°C higher than average in 2013. Rainfall was 72 mm higher than average in 2012 and 126 mm higher than average in 2013. The period from June to August in 2012, which corresponds to the vegetative growth stage and the first half of the reproductive growth stage, was marked by continuous light rain and slightly higher than average temperatures. In September, which corresponds to the latter half of the reproductive growth stage, the temperature and rainfall increased substantially. In 2013, the rainfall was similar to that of an average year in June and July, which corresponds to the vegetative growth stage, but increased considerably in August and September, which corresponds to the entire reproductive growth stage. There was almost no difference in the amount of solar radiation between the 2 years. An analysis of the effects of rainfall during the branch elongation period (June to August) revealed that in 2012, branch elongation was suppressed because of decreased rainfall. In contrast, in 2013, branch elongation was promoted because of increased temperatures and rainfall.
Development as well as the seed yield and related traits in Hokkaido and northern US cultivars
The seed yield and related characteristics in the Hokkaido and northern US cultivars are presented in . In 2012 and 2013, stage R8 (i.e., maturity) occurred 8–22 days later in the US cultivars than in the Hokkaido cultivars (). Additionally, the number of days until the beginning of the podding stage (R5) for the US cultivars (when the branches of the indeterminate cultivars stopped growing) was 11–15 days longer than that for the Hokkaido cultivars.
The total seed yield and branch seed yield of the US cultivars were higher than the corresponding yields of the Hokkaido cultivars under the standard planting (60 cm × 10 cm) and sparse planting (60 cm × 20 cm) conditions in both years. The difference between the Japanese and US cultivars regarding the total seed yield and branch seed yield increased as the planting density decreased. Additionally, the ANOVA results revealed significant differences between the Japanese and US varieties and planting densities as well as a significant interaction between the cultivar and planting density.
The number of pods per branch, the number of nodes per branch, the total branch length, and the number of branches were greater for the US cultivars than for the Hokkaido cultivars at all planting densities in both years. The difference between the Japanese and US cultivars regarding the number of pods per branch and the number of nodes per branch increased as the planting density decreased. The ANOVA indicated there were significant differences between the Japanese and US cultivars and planting densities as well as a significant interaction between the cultivar and planting density.
The average branch length (i.e., total branch length divided by the number of branches) and the standardized branch length [i.e., total branch length divided by the number of days until the beginning of the podding stage (R5)] were greater for the US cultivars than for the Hokkaido cultivars under the standard (60 cm × 10 cm) and sparse (60 cm × 20 cm) planting densities. The differences between the Japanese and US varieties in terms of the average branch length and the total branch length with standardized maturity increased as the planting density decreased. The results of the ANOVA revealed significant differences between the Japanese and US cultivars and planting densities as well as a significant interaction between the cultivar and planting density.
Development as well as the seed yield and related traits in the NILs for stem growth habit
The effects of the stem growth habit of the Canadian cultivar ‘Harosoy’, the northern US cultivar ‘Williams’, and the ST line (‘Stressland’ × ‘Tachinagaha’) on the seed yield and yield-related traits in the NILs are presented in . The maturity stage of the indeterminate NILs with the ‘Harosoy’, ‘Williams’, and ST backgrounds occurred 2–10 days later than that of the determinate NILs (). The number of days until the beginning of the podding stage (R5) in the indeterminate NILs was 10 days longer than that in the determinate NILs in 2012 and 2013.
Similar total seed yields were obtained for the indeterminate NILs with the ‘Harosoy’ and ‘William’ backgrounds. However, the seed yield resulting from the sparse planting (60 cm × 20 cm) conditions was lower than that of the other planting densities for the determinate NILs. Moreover, the total seed yield of the indeterminate and determinate NILs with the ST background decreased with decreasing plant densities.
The branch seed yield clearly increased with decreasing plant densities for the indeterminate NILs with the ‘Harosoy’ and ‘William’ backgrounds. The indeterminate NIL with the ST background also exhibited an increasing trend. However, for all three backgrounds, there was no clear increase associated with decreasing plant densities in the determinate NILs. On the basis of an ANOVA, the total seed yield and branch seed yield were calculated based on the interactions among the stem growth habit, the variety, stem growth, and planting density. These interactions were also significant.
The extent of the increase in the number of pods per branch and the number of nodes per branch associated with sparse planting was larger for the indeterminate NILs than for the determinate NILs with the ‘Harosoy’ and ‘William’ backgrounds. The same tendency was observed for the NILs with the ST background, but it was not necessarily significant. Therefore, the number of pods per branch and the number of nodes per branch were determined based on the interactions among stem growth habit and planting density. These interactions were significant, as were the interactions among the cultivar, stem growth, and planting density.
The increase in the total branch length with decreasing plant densities was greater for the indeterminate NILs than for the determinate NILs in all three backgrounds (‘Harosoy’, ‘Williams’, and ST). The significance of the interactions among the cultivar, stem growth, and planting density was confirmed. The increase in the branch number with decreasing plant densities was also greater for the indeterminate NILs than for the determinate NILs in all three backgrounds. However, the ANOVA results revealed a significant interaction only with the stem growth habit.
Branching plasticity values
The branching plasticity values (i.e., ratios of the branch yield, the number of pods per branch, the number of nodes per branch, and the total branch length under sparse and dense planting conditions) were compared for the Hokkaido and northern US cultivars. In both years, the values were greater for the indeterminate US cultivars than for the determinate Japanese cultivars (). A comparison of the determinate Japanese cultivars with the same growth habit indicated the ratios of the number of pods per branch, the number of nodes per branch, and the total branch length under sparse and dense planting conditions were greatest for ‘Yuzuru’ and smallest for ‘Toyomusume’ in both years. A similar trend in the ratio of the branch yield was observed in 2013, but the differences between cultivars were smaller in 2012.
Among the indeterminate US cultivars, the ratios of the total branch length and pod number under sparse and dense planting conditions were highest for ‘LD00-3309ʹ in both years. Although the ratio of the number of pods per branch was higher for ‘LD00-3309ʹ than for the other cultivars in 2013, the same ratio was higher for ‘Athow’ and ‘Jack’ than for ‘LD003309ʹ in 2012. Similarly, the ratios of the branch yield were higher for ‘Athow’ and ‘Jack’ than for ‘LD00-3309ʹ in both years.
The overall ANOVA revealed the significant main effects of the cultivar for all branch traits. The effect of the stem growth habit on the branching plasticity values was examined in the NILs. In the NILs with the ‘Harosoy’ background, the ratios of the branch yield, the number of pods per branch, the number of nodes per branch, and the total branch length under sparse and dense planting conditions were higher for the indeterminate lines than for determinate lines.
In the NILs with the ‘Williams’ background, the ratios of the number of nodes per branch and the total branch length were higher for the indeterminate lines than for the determinate lines. Although the ratios of the branch yield and the number of pods per branch were higher for the indeterminate lines than for determinate lines in 2012, no differences between lines were observed in 2013 ().
Regarding the NILs with the ST background, with the exception of the ratio of the branch yield in 2013, the ratios of the branch yield, the number of pods per branch, the number of nodes per branch, and the total branch length under sparse and dense planting conditions were greater for the indeterminate lines than for the determinate lines in both years.
Relationships between the branching plasticity value and maturity and the stem growth habit
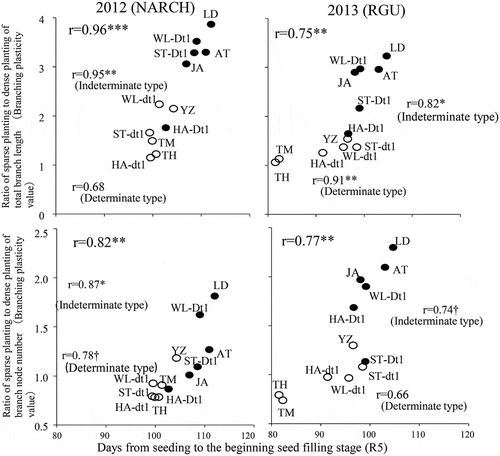
In 2012 and 2013, the ratio of the total branch length was positively correlated with maturity (r = 0.96**) and the stem growth habit (r = 0.75**) when both the determinate and indeterminate days to R5 were included (). When only the indeterminate cultivars were included, positive correlations between the same variables were observed in 2012 (r = 0.95** and r = 0.82*). For the determinate cultivars, although a positive correlation was observed in 2013 (r = 0.91**), no significant correlations were observed in 2012. In 2012 and 2013, another measure of branching plasticity, the ratio of the number of nodes per branch, was positively correlated with days to R5 (r = 0.82**) and the stem growth habit (r = 0.77**) when both determinate and indeterminate cultivars were included (). When only the indeterminate cultivars were considered, there were positive correlations between the same variables in both years (r = 0.87*, r = 0.74†). For the determinate cultivars, although a positive correlation was observed in 2012 (r = 0. 78†), no significant correlations were observed in 2013.
Figure 1. Relationship between the number of days from seeding to the beginning of the seed filling stage (R5) and the branching plasticity values for the total branch length and the number of nodes per branch.
presents the relationship between the ratio of the total branch length under sparse and dense planting conditions (branching plasticity value) in 2012 (NARCH) and 2013 (RGU). This branching plasticity value was positively correlated (r = 0.94**) in 2012 and 2013 when both determinate and indeterminate cultivars were included. When analyzed individually, the branching plasticity values of the determinate and indeterminate cultivars were also correlated between years (r = 0.78†, r = 0.82**).
Figure 2. Relationship between the branching plasticity values for the total branch length and the number of nodes per branch in 2012 (NARCH) and 2013 (RGU).
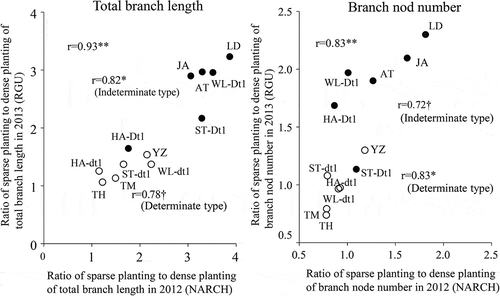
Regarding the total branch length, the ratio of the number of nodes per branch was positively correlated when all cultivars were included. When analyzed separately, the branching plasticity values of the determinate and indeterminate cultivars were also correlated between years (r = 0.83*, r = 0.72†).
Discussion
A positive correlation was observed between the number of days to stage R5 (onset of seed filling) and branching plasticity in terms of the total branch length, both when all cultivars were pooled and when cultivars with different stem growth habits were analyzed separately (). Thus, there is a strong relationship between maturity and branching plasticity, with early- and late-maturing cultivars exhibiting low and high branching plasticity, respectively. Additionally, on the basis of the positive correlation of the total branch length between 2 years (), we believe that although the total branch length for a given cultivar or line may vary between years, the differences between cultivars and lines are stable.
Considering the relationship between the total branch length and the stem growth habit, the values for this branching plasticity measure were greater for the indeterminate US cultivars than for the determinate Japanese cultivars (). Regarding the NILs with different stem growth habits, for all cultivars tested, the branch plasticity of the indeterminate lines was clearly higher than that of the determinate lines (). Therefore, we conclude that the indeterminate growth habit substantially enhances branching plasticity.
The results of earlier investigations suggested that Dt1 expression delays maturation (Chang et al., Citation1982; Green et al., Citation1976). However, branching plasticity in terms of branch length, when normalized by dividing by the number of days to stage R5 (onset of seed filling) to exclude the effect of maturity, was still clearly higher for the indeterminate cultivars than for the determinate cultivars. These observations imply that Dt1 can separately enhance branching plasticity as well as delay maturation.
Agudamu et al. (Citation2016) reported that varietal differences in the planting density dependence of branching plasticity can be evaluated based on the ratio of the total branch length under sparse and dense planting conditions. The results presented herein, which indicate that the ratio of the number of nodes per branch under sparse and dense planting conditions similarly exhibits varietal differences ( and ). Moreover, these differences are stable across years (). Accordingly, we conclude that the number of nodes per branch can be used as an indicator of branching plasticity. As in the case of total branch length, the number of nodes per branch decreases and increases with early and late maturation, respectively (), confirming that the indeterminate growth habit due to Dt1 enhances branching plasticity (, ). In contrast, regarding the use of the ratios of the branch yield and the number of pods per branch under sparse and dense planting conditions as measures of branching plasticity, in late-maturing cultivars, such as ‘LD00-3309ʹ and ‘William’, which mature in late October onwards, low temperatures cause poor seed filling and, consequently, greater numbers of immature pods and decreased yields, ultimately resulting in unstable branching plasticity. This trend was especially evident in 2013, when the average air temperature in September was 1.9°C lower than in 2012. Branch responses to planting density are basically mediated by the following two processes: the branch morphogenesis during the vegetative growth stage and the anthesis and fertilization during the reproductive growth stage upon completion of the branch morphogenesis. With respect to the branch traits evaluated in this study, the total branch length and the number of nodes per branch are associated with the former process, whereas the branch yield and the number of pods per branch are related to the latter process. Accordingly, this study also demonstrates that the latter traits are important when examining the relationship between branching plasticity and yield performance, whereas the former traits are appropriate for examining the varietal differences in branch morphogenesis.
When we determined that the annual differences in the planting density dependence of branching plasticity are related to branch morphogenesis, we observed that, overall, branching plasticity in terms of the total branch length was higher in 2012 than in 2013. We speculate this was because the period from late June to early August, when branches form and vigorously elongate, tended to be drier in 2012 than in 2013, which suppressed the branch elongation in the densely planted plots. The temperatures also tended to be lower, which delayed the onset of anthesis and prolonged the branch elongation period in the sparsely planted plots, thereby increasing the ratio of the total branch length under sparse and dense planting.
A comparison of the magnitude of the varietal differences in the branching plasticity measures for the two growth habit types revealed greater varietal differences among the indeterminate cultivars than among the determinate cultivars ( and , ). It is unclear whether this is attributable solely to the effect of the Dt1 gene combined with maturity or whether a genetic region other than those related to the stem growth habit and maturity is involved.
Board and Kahlon (Citation2013) analyzed 41 late-maturing cultivars (MG V and VI) in the southern US soybean-producing area and concluded that the cultivars adapted to sparse planting conditions can be selected based on the normalized branch weight, which is calculated by dividing the branch dry matter weight at stage R5 (onset of seed filling) by the number of days until stage R5. There is little decrease in the yield of soybean cultivars with a large normalized branch weight under sparse planting conditions. That is, there are varietal differences that are not related to the maturity of cultivars adapted to sparse planting conditions. These findings suggest that there are large varietal differences in the branching plasticity among the late-maturing determinate cultivars. However, in the current study, the varietal differences in the branching plasticity among the determinate cultivars was very small when compared with the varietal differences among the indeterminate cultivars. This discrepancy may be related to the fact that the determinate cultivars tested in this study were early-maturing cultivars (below MG II). To clarify this issue, the existence of genetic regions related to branching plasticity other than those related to the stem growth habit and maturity will need to be confirmed by performing genetic analyses of recombinant inbred lines generated by crossing cultivars with high and low branching plasticity. Additionally, reference cultivars with high, low, and intermediate branching plasticity must be identified from among the cultivars with the same stem growth habit and maturity.
Even if no genetic regions other than those related to maturity and stem growth habit are found to contribute to branching plasticity, combined analyses of the main genes that substantially influence maturity e1/E1, e2/E2 (Bernard, Citation1971), e3/E3 (Buzzell, Citation1971), and e4/E4 (Buzzell & Voldeng, Citation1980) and the genes related to the stem growth habit dt1/Dt1 (Bernard, Citation1972) are important for identifying cultivars with high branching plasticity. Such cultivars may be useful as a breeding line to develop new varieties adapted to cold climates.
This study demonstrated that the branching plasticity is greater in late-maturing soybean cultivars. Additionally, the indeterminate growth habit conferred by Dt1 substantially enhances branching plasticity.
Conclusions
On the basis of comparisons of Japanese and US cultivars and NILs for the Dt1 gene (indeterminate growth habit), it is evident that the branching plasticity of indeterminate cultivars and lines is greater than that of determinate cultivars and that the varietal differences are considerable. The branching plasticity in response to the planting density of soybean cultivars increases with delayed maturation. Furthermore, the indeterminate growth habit substantially enhances branching plasticity.
PPS2019_032RP-File009.xlsx
Download MS Excel (21.3 KB)Acknowledgments
This research was supported in part by a grant from the Ministry of Agriculture, Forestry, and Fisheries of Japan (Genomics-based Technology for Agricultural Improvement, SFC1002). The US soybean cultivars were provided by the National Plant Germplasm System, USDA-ARS, USA. The Hokkaido soybean cultivars were provided by the Agricultural Experiment Station of the Hokkaido Research Organization. We thank the students of the Crop Science Laboratory, Department of Sustainable Agriculture, Rakuno Gakuen University for assisting with the yield survey and sampling. We also thank Edanz Group (https://en-author-services.edanzgroup.com/) for editing a draft of this manuscript.
Disclosure Statement
No potential conflict of interest was reported by the authors.
Supplemental Material
Supplemental data for this article can be accessed here.
Related Research Data
References
- Agudamu, Y. T., & Shiraiwa, T. (2016). Branch development responses to planting density and yield stability in soybean cultivars. Plant Production Science, 19(3), 331–339. https://doi.org/10.1080/1343943X.2016.1157443
- Agudamu, Yoshihira. T., & Shiraiwa, T. (2015). Effect of stem growth habit on soybean yield stability to planting density: Varietal difference in branching plasticity, and the factorial analysis from the point of view of branch development and light-intercepting characteristics. Research of One Health, 2015, In Japanese with English title. 17–38. https://gra.rakuno.ac.jp/wp-content/themes/aplan/pdf/roh_gihei.pdf
- Bernard, R. L. (1971). Two major genes for time of flowering and maturity in soybeans. Crop Science, 11(2), 242–244. https://doi.org/10.2135/cropsci1971.0011183X001100020022x
- Bernard, R. L. (1972). Two genes affecting stem termination in soybeans. Crop Science, 12(2), 235–239. https://doi.org/10.2135/cropsci1972.0011183X001200020028x
- Board, J. E. (2000). Light interception efficiency and light quality affect yield compensation of soybean at low plant populations. Crop Science, 40(5), 1285–1294. https://doi.org/10.2135/cropsci2000.4051285x
- Board, J. E., & Kahlon, C. S. (2013). Morphological responses to low plant population differ between soybean genotypes. Crop Science, 53(3), 1109–1119. https://doi.org/10.2135/cropsci2012.04.0255
- Buzzell, R. I. (1971). Inheritance of a soybean flowering response to fluorescent daylength. conditions. Canadian Journal of Genetics and Cytology, 13(4), 703–707. https://doi.org/10.1139/g71-100
- Buzzell, R. I., & Voldeng, L., H. D. (1980). Inheritance of insensitivity to long daylength. Soybean Genetics Newsletter, 7, 26–29. https://lib.dr.iastate.edu/soybeangenetics/vol7/iss1/13/
- Carpenter, A. C., & Board, J. E. (1997a). Branch seed yield components controlling soybean yield stability across plant populations. Crop Science, 37(3), 885–891. https://doi.org/10.2135/cropsci1997.0011183X003700030031x
- Carpenter, A. C., & Board, J. E. (1997b). Growth dynamic factors controlling soybean yield stability across plant populations. Crop Science, 37(5), 1520–1526. https://doi.org/10.2135/cropsci1997.0011183X003700050018x
- Chang, J. F., Green, D. E., & Shibles, R. (1982). Yield and agronomic performance of semi-determinate and indeterminate soybean stem types. Crop Science, 22(1), 97–101. https://doi.org/10.2135/cropsci1982.0011183X002200010022x
- Green, D. E., Burlamaqui, P. F., & Shibles, R. (1976). Performance of randomly selected soybean lines with semideterminate and indeterminate growth habits. Crop Science, 17(2), 335–339. https://doi.org/10.2135/cropsci1977.0011183X001700020026x
- Hanway, J. J., & Weber, C. R. (1971). Dry matter accumulation in soybean plants as influenced by N, P, and K fertilizer. Agronomy Journal, 63(2), 263–266. https://doi.org/10.2134/agronj1971.00021962006300020020x
- Herbert, S. G., & Litchfield, G. V. (1982). Partitioning soybean seed yield components. Crop Science, 22(5), 1074–1079. https://doi.org/10.2135/cropsci1982.0011183X002200050044x
- Kato, M., Minamida, K., Tojo, M., Kokuryu, T., Hamaguchi, H., & Shimada, S. (2013). Association of Pythium and Phytophthora with preemergence seedling damping-off of soybean growth in field converted from a paddy field in Japan. Plant Production Science, 16(1), 95–104. https://doi.org/10.1626/pps.16.95
- Lee, C. D., Egli, D. B., & Tekrony, D. M. (2008). Soybean response to plant population at early and late planting dates in the mid-South. Agronomy Journal, 100(4), 971–976. https://doi.org/10.2134/agronj2007.0210
- Lueschen, W. E., & Hicks, D. R. (1977). Influence of plant population on field performance of three soybean cultivars. Agronomy Journal, 69(3), 390–393. https://doi.org/10.2134/agronj1977.00021962006900030015x
- Ministry of Agriculture, Forestry and Fisheries Minister’s Secretariat Statistics Division. (2009). Field crop statistics (Food crops, forage crops, craft crops). In Japanese, The 91st Statistical Yearbook of Ministry of Agriculture, Forestry and Fisheries. https://www.maff.go.jp/e/data/stat/91th/
- Okabe, A. .(2011). [Soybean cultivation in Kinki, Chugoku, and Shikoku], M. Kokubun Ed., Cultivation and uses in soybean. 76–80. Tokyo: Asakura. In Japanese.
- Parvez, A. Q., Gardner, F. P., & Boote, K. J. (1989). Determinate- and indeterminate-type soybean cultivar responses to pattern, density, and planting date. Crop Science, 29(1), 150–157. https://doi.org/10.2135/cropsci1989.0011183X002900010034x
- Rigsby, B., & Board, J. E. (2003). Identification of soybean cultivars that yield well at low plant populations. Crop Science, 43(1), 234–239. https://doi.org/10.2135/cropsci2003.2340
- Sanbuichi, T., & Tsuchiya, T. (1976). Growth Of Early Stages in Different Leaflet Shapes Of Soybeans. Hokkaido British Japanese Journal of Crop Science, 16, 25. In Japanese with English title. https://doi.org/10.20751/hdanwakai.16.0_25
- Shimada, S. (2011). Soybean cultivation management technology. In M. Kokubun (Ed.), Cultivation and uses in soybean (pp. 45–59). In Japanese.
- Shimada, S., Hirokawa, F., & Miyakawa, T. Effects of planting date and planting density on a high yielding soybean cultivar grown at drained paddy field in Sanyo district. (1990). Japanese Journal of crop science, 59(2), 257–264. In Japanese with English title. https://doi.org/10.1626/jcs.59.257
- Takahashi, M. (2011). Soybean cultivation in Kyushu. In M. Kokubun (Ed.), Cultivation and uses in soybean (pp. 81–86). In Japanese. Asakura Publishing Co., Ltd
- Tsuchiya, T. (1986). Culture method. K. Sunada Ed., Soybean cultivation in Hokkaido region (64–71). Agricultural Technique Extension Association. In Japanese.