ABSTRACT
Future agricultural management programs should consider the responsiveness of crops to increasing atmospheric CO2 concentrations ([CO2]) and temperatures. Eddo is an important tuber crop widely grown in Asia, but its response to [CO2] and temperatures remains largely unknown. We investigated the responses of eddo and rice plants to elevated [CO2] (ambient [CO2] + 200 µmol mol−1) at low and high temperatures during the early-to-intermediate growth stage in temperature gradient chambers in 2018 and 2019. The dry weights (DWs) of the eddo leaf blades, petioles, roots, and above-ground parts as well as the DWs of the rice leaf blades, leaf sheaths and stems, above-ground parts, and whole plants significantly increased under elevated [CO2]. The number of leaves, leaf area, and SPAD value of eddo significantly increased under elevated [CO2], whereas the leaf area and SPAD value of rice did not. The net photosynthetic rate of eddo and rice tended to increase under elevated [CO2]. The increasing ratio of the total DW in elevated vs ambient [CO2] was significantly higher for eddo than for rice. The result indicates that elevated [CO2] increases the eddo biomass more than the rice biomass, possibly because of the increasing eddo leaf area and SPAD value induced by elevated [CO2]. The total DW of rice increased significantly in response to the high air temperature under elevated [CO2] conditions. However, the effect of [CO2] on the total DW of eddo was similar at both temperatures, reflecting the non-significant interaction between [CO2] and temperature.
Abbreviations: [CO2]: atmospheric carbon dioxide concentration, DW: dry weight, FACE: free-air CO2 enrichment, gs: stomatal conductance, PN: net photosynthetic rate, TGC: temperature gradient chamber
1. Introduction
The atmospheric carbon dioxide concentration ([CO2]) has increased from a stable 280 µmol mol−1 before the industrial revolution to 409 µmol mol−1 in 2019 (NOAA, Citation2020). The [CO2] has increased by an average of 1.3 µmol mol−1 per year over the past 40 years (1960–2000), and recent estimates indicate that the average [CO2] has risen by 2.2 µmol mol−1 per year from 2001 to 2019 (NOAA, Citation2020). Thus, various studies have investigated the effects of elevated [CO2] on plants in open-top chambers, growth chambers, greenhouses, and fields with free-air CO2 enrichment (FACE) systems. Many of these studies revealed that elevated [CO2] conditions positively affect plant growth and yield (Kimball, Citation1983; Kimball et al., Citation2002; Long et al., Citation2004). Kimball (Citation2016) reported that it is possible to increase the biomass and yield of C3 species and that the average yield of some C3 grain crops can increase by approximately 19% using a FACE system.
Photosynthesis can be accelerated by elevated [CO2] because the carboxylation rate of RuBisCo increases and the oxygenation of ribulose-1,5-bisphosphate is competitively inhibited (Drake et al., Citation1997). An initial increase in photosynthesis can subsequently be partially or fully down-regulated during a long-term exposure to elevated [CO2] (Stitt, Citation1991). Photosynthetic regulation partly depends on the balance between the substrate for photosynthesis (such as CO2) and the sink capacity (Paul & Foyer, Citation2001). Plants with large vegetative sinks are stimulated by elevated [CO2] more than plants with shorter-lived reproductive sinks (Arp, Citation1991; Miglietta et al., Citation1998; De Temmerman et al., Citation2007). Potato has a large carbohydrate sink in the form of tubers that have apoplastic phloem for loading sucrose, which leads to an increased yield under elevated [CO2] (Farrar & Williams, Citation1991; Schapendonk et al., Citation2000; De Temmerman et al., Citation2002). Thus, there is a hypothesis that sink constraints limit the CO2 fertilization effect, and root and tuber crops may have a more significant response to elevated [CO2] than other plant types because their sink size is often unlimited.
Carbon dioxide is a significant contributor to global warming. The global surface temperature will likely rise by 1.5°C between 2030 and 2052 even if stringent measures are imposed to limit greenhouse gas emissions (IPCC, Citation2018). Therefore, it is imperative that the effects of increasing [CO2] and temperature on future crop growth and agricultural productivity are investigated. Previous studies have focused on the combined effects of elevated [CO2] and temperature on many major crop species such as rice (Baker et al., Citation1992; Cheng et al., Citation2009; Jing et al., Citation2016; Nakagawa & Horie, Citation2000; Roy et al., Citation2012; Shimono et al., Citation2008), maize (Mina et al., Citation2019; Ruiz-Vera et al., Citation2015), wheat (Cai et al., Citation2016; Ottman et al., Citation2012), soybean (Palacios et al., Citation2019), cassava (Imai et al., Citation1984), and peanut (Bannayan et al., Citation2009; Newman et al., Citation2005). In rice, doubling the [CO2] increases the whole plant dry weight (DW) more at high air temperatures [33/26°C (day/night)] than at low air temperatures [28/21°C (day/night)] during the vegetative growth stage (Imai et al., Citation1985). However, a recent review of the responses of various crops to [CO2] revealed both positive and negative interactions between [CO2] and temperature on crop yield and growth in FACE systems (Kimball, Citation2016). Kimball (Citation2016) suggested that increasing the temperature alone can stimulate or slow plant growth depending on whether a plant is currently exposed to an environmental temperature that is below or above its optimum temperature for growth.
Eddo [Colocasia esculenta (L.) Schott var. antiquorum Hubbard & Rehder] is a root and tuber crop belonging to the taro family (Araceae). Taro is cultivated in various regions worldwide, including in Africa, Asia, and Oceania. Its corms are highly valuable as a staple food. Thus, the demand for this crop is likely to continue to increase as the global human population increases. In 2018, taro production was about 10.6 million tons worldwide, and the harvested area was about 1.7 million ha (FAOSTAT, Citation2020). Eddo is the most widely cultivated taro crop in Japan. In 2018, eddo production in Japan was 144,800 tons, with a mean yield of 1,260 kg 10a−1 (e-STAT, Citation2020). More specifically, the eddo production in 2018 in Iwate (experimental site of this study), Yamagata, and Fukushima prefectures in the Tohoku region of northern Japan was 4,496 tons (3.1% of the total production in Japan), with a mean yield of 803 kg 10a−1 (e-STAT, Citation2020). The minimum temperature needed for the eddo seed corms to germinate is 15°C and the optimum temperature for growth is 25–30°C, whereas eddo plants cease to grow at temperatures below 5°C (Hoshikawa, Citation1980). Cultivating eddo in northern Japan can be difficult because of the relatively low mean air temperature and short cultivation period at suitable temperatures.
Eddo corms are detectable from the early vegetative stage. Eddo has an unlimited sink capacity in the corms, whereas the underground parts of cereals, such as rice, do not. Accordingly, the corms may positively contribute to eddo responses to elevated [CO2]. Increased temperatures at the experimental site of this study in northern Japan should enhance eddo production and result in better responses to [CO2] compared with the corresponding responses in rice. In this study, we hypothesized that eddo may respond to elevated [CO2] better than rice and that elevated [CO2] positively affects eddo growth and biomass during the early-to-intermediate vegetative growth stage, especially at temperatures greater than the ambient temperature at our experimental site. To test these hypotheses, we used two different types of crops, eddo and rice, and investigated the responses to elevated [CO2] at different air temperatures during the early-to-intermediate vegetative growth stage. We used temperature gradient chambers (TGCs) to examine the effects of elevated [CO2] on eddo and rice plant growth, biomass, the soil plant analysis development (SPAD) value, stomatal conductance (gs), and the net photosynthetic rate (PN) of leaf blades at two different air temperatures.
2. Materials and methods
2.1. Study sites and treatment conditions
We evaluated the effects of elevated [CO2] at two different air temperatures in two TGCs from June 30 to 4 August 2018 and from June 15 to 20 July 2019 at the Tohoku Agriculture Research Center, The National Agriculture and Food Research Organization (NARO) (39°74′N, 141°13′E) in Morioka, Japan.
The TGCs were sunlit greenhouses (6 m wide × 30 m long × 3 m high). Fans were used to ensure the air in the TGCs flowed continuously from the inlet to the exhaust fans. A temperature gradient was continuously maintained along the longitudinal axis by cooling the air with an air conditioner at the inlet end and/or by warming the air with solar radiation or a supplemental heat input (an air heater and ducts) at the outlet end. The [CO2] in one TGC was kept at ambient levels, whereas that in the other TGC was kept at about 200 µmol mol−1 above the ambient levels by injecting pure CO2 (the [CO2] exceeded 99.95%; HOKURYO Co., Ltd., Iwate, Japan) at the air inlet between 03:30 and 19:30. The CO2 emission rate was set proportional to the air exhaust rate. An infrared gas analyzer (LI-820; LI-COR, Lincoln, NE, USA) was used to monitor the [CO2] based on the air collected from about 2 m above the ground in the middle of each TGC. The average daytime (04:00 to 20:00) [CO2] over the treatment period was 416 ± 12 μmol mol−1 (data are presented here and subsequently as the mean ± standard deviation) in 2018 and 415 ± 8 μmol mol−1 in 2019 in the ambient [CO2] TGC, whereas it was 616 ± 13 μmol mol−1 in 2018 and 609 ± 8 μmol mol−1 in 2019 in the elevated [CO2] TGC.
The air temperature and relative humidity were monitored at each location using Pt 100 resistance hygrothermographs with an aspirated double-tube radiation-shield. Five hygrothermographs were set at approximately even intervals along the longitudinal axis of each TGC, and the height of the hygrothermographs was gradually increased so that it was consistent with the maximum plant height of rice. Two plots were set along an air temperature gradient in each TGC. The air temperature and relative humidity of each plot were calculated based on their gradients in each TGC. In the ambient [CO2] TGC, the low air temperature plot (approximately the same as the ambient air temperature) had an average air temperature of 23.4°C ± 2.5°C in 2018 and 19.0°C ± 1.6°C in 2019, whereas the high air temperature plot had an average air temperature of 27.7°C ± 2.6°C in 2018 and 23.1°C ± 1.7°C in 2019. In the elevated [CO2] TGC, the low air temperature plot had an average air temperature of 23.5°C ± 2.6°C in 2018 and 19.1°C ± 1.6°C in 2019, whereas the high air temperature plot had an average air temperature of 27.9°C ± 2.7°C in 2018 and 23.2°C ± 1.7°C in 2019. The natural air temperatures outside of the TGCs during the treatment periods were 24.2°C ± 3.4°C in 2018 and 19.7°C ± 2.5°C in 2019.
In the ambient [CO2] chamber, the relative humidity was 76% ± 8% and 77% ± 8% in the low air temperature plot and 40% ± 4% and 49% ± 8% in the high air temperature plot in 2018 and 2019, respectively. In the elevated [CO2] chamber, the relative humidity was 70% ± 8% and 75% ± 11% in the low air temperature plot and 45% ± 5% and 48% ± 5% in the high air temperature plot in 2018 and 2019, respectively. Solar radiation was measured using sunlight sensors (LP02; TAIYO KEIKI, Tokyo, Japan), which were positioned approximately 5 m above the ground beside the TGCs. Total light transmission of outer cover films (F-CLEANTM clear; AGC Green-Tech Co., Ltd., Tokyo, Japan) for TGCs is 94%. The average daytime solar radiation during the treatment period was 193 Wm−2 day−1 in 2018 and 181 Wm−2 day−1 in 2019. The [CO2], air temperature, relative humidity, and solar radiation were recorded at 5 s intervals and the averages for 1 min intervals were calculated by a data logger (CR 1000; Campbell Sci. Inc., Logan, UT, USA.).
Plants are generally more sensitive to air temperature changes than to relative humidity changes. Thus, the four treatments in this study were as follows:
A[CO2]LT: ambient [CO2] and low air temperature;
E[CO2]LT: elevated [CO2] and low air temperature;
A[CO2]HT: ambient [CO2] and high air temperature;
E[CO2]HT: elevated [CO2] and high air temperature.
2.2 Plant materials and cultural practices
Eddo cultivar Aichiwase, which is one of the major cultivars in Japan, was analyzed in this study because of a lack of widely cultivated cultivars in the Tohoku region. Additionally, we were interested in how common eddo varieties may respond to future conditions in northern Japan resulting from global warming (i.e., elevated [CO2] and high temperatures). Rice cultivar Hitomebore, which is widely cultivated in northern Japan, was also analyzed in this study. Eddo seed corms of a uniform size were selected, with an average fresh weight per corm of 71 ± 6 g and 78 ± 7 g in 2018 and 2019, respectively. The selected seed corms were sterilized in 0.5% (v/v) sodium hypochlorite solution for 5 min and then washed with water for 30 min. The corms were sown in small plastic pots (13 cm height × 12 cm internal diameter) filled with commercial soil (containing 0.32 g L−1 nitrogen, 0.21 g L−1 phosphorus, and 0.30 g L−1 potassium) (Ikubyobaiyodo; TAKII & Co., Ltd., Kyoto, Japan). The seed corms were covered by 3–4 cm of soil on May 25 for the 2018 experiment and May 16 for the 2019 experiment. Rice seeds were soaked for 10 min in 20% (v/v) sodium chloride solution to eliminate unfilled seeds and then sterilized with a fungicide containing Procloraz (SPORTAK®; Nissan Chemicals Co. Ltd., Saitama, Japan). The seeds were submerged in water at 20°C for 3 days and then at 30°C for 1 day. The germinated seeds were sown in a seedling tray on June 21 for the 2018 experiment and June 6 for the 2019 experiment. The eddo and rice seedlings were grown in a greenhouse with opened windows at Hirosaki University (40°59′N, 140°47′E), Hirosaki, Japan.
Treatments were initiated in the TGCs when the plants reached the early vegetative growth stage and lasted for 5 weeks, at which point the plants were in the intermediate vegetative growth stage. Just before the treatments in the TGCs, two young eddo plants at the 3- to 4-leaf stage in 2018 and at the 2- to 2.5-leaf stage in 2019 were transplanted into a plastic pot (35 cm height × 32 cm internal diameter) filled with the same soil used in the seedling period. The lateral buds had not yet extended when the plants were transplanted. The corms were covered by about 6 cm of soil. The plants were suitably watered three to five times every week during the treatment period. At 9 days after sowing, the rice seedlings (the plant age in leaf number was three- to three point two) were transplanted into 1/5000a Wagner’s pots (19 cm height × 16 cm internal diameter) filled with commercial soil (containing 1.7 g L−1 nitrogen, 4.4 g L−1 phosphorus, and 1.8 g L−1 potassium) (Agrobaido; Kanumasangyo Co. Ltd., Tochigi, Japan), with two seedlings per pot, before starting the treatments in the TGCs. The rice seedlings were flooded during the treatment period. For both eddo and rice plants, each pot was rotated and their positions were changed weekly to minimize the effects of environmental differences.
2.3 Growth and dry weight measurements
Regarding eddo, an apical bud germinates from a seed corm and develops into the main plant part. Lateral buds also germinate and develop into the lateral plant parts. Therefore, an eddo plant consists of a main plant part and some lateral plant parts. In this study, 9–10 eddo plants were analyzed per treatment. The number of leaves and number of lateral plants in an eddo plant were individually counted, and then the plant height was measured 1 day before the final treatment day. The same eddo plants were collected from pots on the final treatment day, after which the roots were carefully washed with tap water to remove soil. The leaf blades, petioles, corms, and roots were collected separately for each treatment. The eddo leaf blades and a scale were photographed with a digital camera. The leaf area was measured using the Image J software (1.52a; Wayne Rasband, National Institutes of Health, MD, USA). Eddo plants were dried at 80°C for 6 days before measuring the DW. Additionally, 10 rice plants were analyzed per treatment. The number of leaves on the main stem part as well as the plant height and number of tillers were determined 1 day before the final treatment day. Rice plants were sampled at the same time and in the same manner as the eddo plants. The rice leaf blade area was measured using an automatic leaf area meter (AAM-9; Hayashi Denko Co. Ltd., Tokyo, Japan). Analyses were completed with the 9–10 eddo plants and the 10 rice plants collected for each treatment.
2.4 SPAD, stomatal conductance, and net photosynthetic rate measurements
For each treatment, we randomly selected five eddo and rice plants from among the analyzed plants described above to measure the gs and PN at the central part of the youngest fully expanded leaves on the eddo main plant and rice main stem at 2–3 days before the final treatment day in 2018 and 2019. The measurements were conducted between 07:00 and 14:00 using a portable photosynthesis measurement system (LI-6400; LI-COR, Lincoln, NE, USA). The [CO2] in the air entering the leaf chamber cuvette was adjusted to 400 and 600 μmol mol−1 for the ambient and elevated [CO2] plots, respectively. The relative humidity of the air entering the cuvette was adjusted to about 60%. In the cuvette, the photosynthetic photon flux density was set to 1,500 μmol m−2 s−1 using the internal blue and red LED light source and the block temperature was adjusted to 27°C. After the gs and PN were measured, the SPAD value was determined using a SPAD meter (SPAD-502, Minolta, Tokyo, Japan). The measurements were completed using a single leaf blade from the main parts of five individual eddo and rice plants per treatment.
2.5 Statistical analysis
We conducted an analysis of variance (ANOVA) with a split-plot design to determine the significance of the effects of [CO2] and temperature on eddo and rice growth. The main plot was [CO2] and the subplot was air temperature, with years (2018 and 2019) treated as replications (random effect). The ANOVA model was constructed by CO2 (C), Temperature (T), Year (Y), C × T, C × Y and C × T × Y for . The natural logarithms of the mean value ratios in elevated vs ambient [CO2] for the total DW and PN were used for the ANOVA with a model constructed by Crop (Cr), Temperature (T), Year (Y), Cr × T, Cr × Y, Cr × T × Y for . Statistical significance was indicated for P < 0.01 and P < 0.05, and the actual P values were presented herein as 0.05 ≤ P < 0.1. All statistical analyses were conducted using the general linear model procedure of the SPSS statistical software (version 24.0; IBM, New York, NY, USA).
Table 1. Effects of elevated CO2 concentration ([CO2]) on eddo and rice plant growth at two different air temperatures
Table 2. Effects of elevated CO2 concentration ([CO2]) on the dry weight (DW) of eddo at two different air temperatures
Table 3. Effects of elevated CO2 concentration ([CO2]) on the dry weight (DW) of rice at two different air temperatures
Table 4. Effects of elevated CO2 concentration ([CO2]) on the mean value ratios of the total dry weight (DW) and net photosynthetic rate in elevated vs ambient [CO2] for eddo and rice at two different air temperatures
Table 5. Effects of elevated CO2 concentration ([CO2]) on the SPAD value, stomatal conductance (gs), and net photosynthetic rate (PN) of a single leaf blade from eddo and rice plants at two different air temperatures
3. Results
3.1 Growth parameters
An analysis of eddo plants revealed that the number of leaves, leaf area, and plant height increased significantly (P < 0.05) with elevated [CO2] (). The number of leaves, leaf area, and the number of lateral plants in eddo also significantly (P < 0.05) increased with high temperature (). Regarding the rice plants, the number of leaves (P < 0.01) was significantly increased by elevated [CO2] and the number of leaves and the number of tillers were significantly (P < 0.05) increased by high temperature (). ANOVA results of the combination of [CO2] and temperature showed that the number of lateral plants in eddo under elevated [CO2] was significantly (P < 0.05) increased by high temperature (). In rice, there were no significant [CO2] × temperature interactions for any of the examined growth parameters ().
3.2 Dry weights
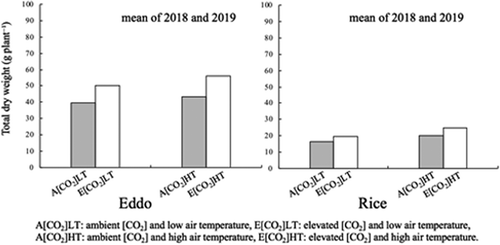
The DWs of the eddo leaf blades, petioles, and above-ground parts increased significantly (P < 0.05) with elevated [CO2] (). The ANOVA results also revealed the root DW and total DW of eddo plants tended to increase under elevated [CO2], with P values of 0.070 and 0.080, respectively (). In rice, the DWs of the leaf blades, leaf sheaths and stems (shoots without leaf blades), above-ground parts, and whole plants increased significantly (P < 0.05) with elevated [CO2] (). Additionally, the DW of eddo corms and the DWs of rice leaf blades, leaf sheaths and stems, above-ground parts, and whole plants increased significantly (P < 0.05) in response to the high air temperature (). Effects of elevated [CO2] on all DW parameters of eddo are similar at low and high temperatures, reflecting the non-significant interaction between [CO2] and temperature (). In rice, the total DW in response to elevated [CO2] was significantly increased (P < 0.01) by the exposure to the high air temperature (). The mean value ratios in elevated vs ambient [CO2] at low and high air temperatures were 1.26 and 1.30 for eddo and 1.17 and 1.26 for rice, respectively (). The ratio was significantly higher (P < 0.05) for eddo than for rice at both temperatures ().
3.3 SPAD value, stomatal conductance, and net photosynthetic rate
The SPAD values increased significantly (P < 0.01) when eddo plants were exposed to elevated [CO2] (). An exposure to high air temperature did not increase the SPAD values under elevated [CO2] conditions (). In rice, [CO2] and temperature alone or combined had no significant effects on the SPAD values (). For both eddo and rice, the highest mean SPAD values were obtained following the E[CO2]HT treatment (). Elevated [CO2] and temperature alone and combined had no significant effect on the gs of eddo or rice plants; however, the mean gs tended to decrease in response to elevated [CO2] at the low and high air temperatures in both 2018 and 2019 ().
There were no significant increases in PN under elevated [CO2] conditions for eddo and rice, with P values of 0.055 and 0.056, respectively (). The high air temperature treatment had a significant positive effect (P < 0.05) on the PN, but only for eddo. The PN of eddo and rice under elevated [CO2] conditions was not significantly increased by the high air temperature (). The mean ratios of the PN in elevated [CO2] vs ambient [CO2] at low and high air temperatures were respectively 1.32 and 1.38 for eddo and 1.28 and 1.18 for rice (). There were no significant differences in the ratios between eddo and rice and between the temperature treatments ().
4. Discussion
Previous studies proved that elevated [CO2] substantially affects plant growth and biomass. Saminathan et al. (Citation2019) reported that for sweet potato, the number of storage roots, the leaf, root, and stem DWs, and the total plant biomass increase significantly in response to elevated [CO2]. Similarly, in an earlier investigation by Aien et al. (Citation2014), an exposure to elevated [CO2] significantly increased the number of tubers per plant and the total dry matter production of two potato cultivars. Sakai et al. (Citation2019) conducted FACE experiments, in which the yields of five Japanese rice cultivars increased although the increasing rate varied depending on the cultivar. In our study, many parameters related to the growth and DW of eddo increased significantly when plants were exposed to elevated [CO2] (). Therefore, to the best of our knowledge, this is the first report of the positive effects of elevated [CO2] on eddo growth and biomass during the early-to-intermediate vegetative growth stage. The total DW of rice plants increased significantly (P < 0.01) in response to elevated [CO2] (), whereas the total DW of eddo plants tended to increase (P = 0.080) under elevated [CO2] conditions (). However, we also determined that the increasing ratio of the total DW in elevated vs ambient [CO2] was higher for eddo than for rice (). The result suggests that elevated [CO2] increases the biomass of eddo plants more than the biomass of rice plants under the analyzed experimental conditions. This finding may support the hypothesis that plants with a large sink capacity, such as root and tuber crops, are more responsive to elevated [CO2] than plants with a limited sink capacity. The number of leaves and the leaf area of eddo plants increased significantly following an exposure to elevated [CO2], whereas the leaf area of rice plants did not significantly increase in response to elevated [CO2] (). Thus, the leaf area as well as the number of leaves may contribute to the higher increasing rate of biomass in eddo under elevated [CO2] conditions.
Various results have been reported for the combined effects of elevated [CO2] and temperature on biomass and/or yield of rice, wheat, maize, soybean, and C3 and C4 grasses (Kimball, Citation2016). In cassava, a root and tuber crop, the number of leaves, leaf area, and stem length as well as the leaf blade, petiole, stem, root, and whole plant DWs under elevated [CO2] conditions are significantly higher at high air temperatures [33/26°C (day/night)] than at low air temperatures [28/21°C (day/night)] (Imai et al., Citation1984). In the current study, the mean values for the analyzed growth and biomass parameters for eddo and rice under elevated [CO2] conditions were higher at the high air temperature than at the low air temperature, regardless of significance (). Additionally, under elevated [CO2], the total DW of rice increased significantly at the high air temperature (). In contrast, under elevated [CO2], the eddo DW was not significantly increased by the exposure to the high air temperature (). This may be explained by the notable difference in plant growth between 2018 and 2019, which was primarily due to the difference in the average air temperature and solar radiation during the treatment period between the 2 years. Accordingly, these differences between years likely affected the ANOVA results. Additional investigations will need to be conducted to verify the hypothesis that the positive effects of elevated [CO2] on the eddo biomass are enhanced by warm conditions at our experimental site.
The leaf chlorophyll contents of garlic (Allium sativum L., cv. White Roppen) reportedly increase significantly with increasing CO2 concentrations (Naznin et al., Citation2015). SPAD value is one of the indicators of chlorophyll content. In an earlier study on rice, the simultaneous increase in [CO2] and air temperature led to a 14% increase in the SPAD value, relative to the average value in the open field (Figueiredo et al., Citation2015). In the current study, the SPAD value significantly increased in response to elevated [CO2] for eddo, but not for rice (). Uprety et al. (Citation2002) reported that an exposure to high CO2 levels decreases the gs, which affects the photosynthetic rate. Similarly, the gs of eddo and rice exhibited a decreasing trend under elevated [CO2] conditions at the low and high air temperatures, although the changes were not significant (). Regarding eddo, we propose that the increase in the chlorophyll content may be one of the factors influencing the tendency of PN (P = 0.055) to increase under elevated [CO2] conditions at both temperatures ().
In this study, the positive effects of elevated [CO2] on eddo plants during the early-to-intermediate vegetative growth stage were revealed by analyzing plant growth, biomass, SPAD values, and PN (a positive tendency). The actual mean values of corm DW and the number of lateral plants in eddo in 2018 and 2019 were higher under elevated [CO2] than under ambient [CO2] and both values were the highest in elevated [CO2] conditions under the high air temperature, although not significantly so (). Regarding eddo, increases in the number of lateral plant parts are associated with increases in the number of corms. The tendency for eddo plants to respond positively to elevated [CO2] in the early-to-intermediate vegetative growth stage might contribute to the final eddo yield. However, Sage et al. (Citation1989) indicated that a long-term exposure to elevated [CO2] affects the CO2 response of photosynthesis and that the patterns of acclimation vary among species in the following three ways: (a) the initial CO2 response is unaffected, but the photosynthetic rate at high CO2 levels increases (Solanum tuberosum); (b) the initial CO2 response decreases, but the CO2-saturated photosynthetic rate is only slightly affected (Chenopodium album and Phaseolus vulgaris); (c) both the initial CO2 response and the CO2-saturated photosynthetic rate decrease (Brassica oleracea and Solanum melongena). Future studies on the effects of a long-term exposure to elevated [CO2] on eddo plants may lead to a more comprehensive characterization of the acclimation of photosynthesis and plant yield. Additionally, some environmental conditions (i.e. soil temperature, and difference in temperature between day and night) in pot experiments of TGCs and field experiments are possibly different. FACE tests with customary cultivation are also needed in the future. This study firstly provided information related with elevated [CO2] response of eddo at two different air temperatures with using TGCs.
5. Conclusions
In this study, we revealed that some biomass parameters of eddo plants significantly increase in response to elevated [CO2] at low and high air temperatures. In rice, the biomass also significantly increases under elevated [CO2] conditions at both temperatures. The data presented herein indicate that the number of leaves, leaf area, and SPAD value of eddo significantly increase in response to elevated [CO2], whereas the leaf area and SPAD value of rice do not. Moreover, the PN tends to increase under elevated [CO2] in eddo and rice. To further compare eddo and rice, we investigated the increasing ratio of the total DW and PN, which are key parameters, in elevated vs ambient [CO2]. The increasing ratio of total DW was significantly greater for eddo than for rice. The result implies that an exposure to elevated [CO2] increases the eddo biomass more than the rice biomass. This finding can support the hypothesis that plants with a large sink capacity are more responsive to elevated [CO2]. Considered together, these findings suggest that the leaf area and SPAD value (chlorophyll content) contribute to the biomass increases of eddo plants in response to elevated [CO2]. Furthermore, under elevated [CO2] conditions, the total DW of rice increased significantly at the high air temperature. In contrast, the effect of [CO2] on the total DW of eddo was similar at low and high temperatures under our experimental conditions.
Acknowledgments
We would like to thank Yusei Tsushima, Center for instrumental analysis, Hirosaki University, Japan, for providing technical assistance with scanning electron microscopy and Kafumi Segawa, NARO TARC, for assistance with plant cultivation at TGCs. We thank Edanz Group (https://en-author-services.edanzgroup.com/ac) for editing a draft of this manuscript.
Disclosure statement
The authors have no conflicts of interest to declare.
Correction Statement
This article has been republished with minor changes. These changes do not impact the academic content of the article.
References
- Aien, A., Pal, M., Khetarpal, S., & Pandey, S. K. (2014). Impact of elevated atmospheric CO2 concentration on the growth and yield in two potato cultivars. Journal of Agriculture Science and Technology, 16(7), 1661–1670. http://journals.modares.ac.ir/article-23-4377-en.html
- Arp, W. J. (1991). Effects of source-sink relations on photosynthetic acclimation to elevated CO2. Plant, Cell & Environment, 14(8), 869–875. https://doi.org/10.1111/j.1365-3040.1991.tb01450.x
- Baker, J. T., Allen, L. H., Jr., & Boote, K. J. (1992). Temperature effects on rice at elevated CO2 concentration. Journal of Experimental Botany, 43(7), 959–964. https://doi.org/10.1093/jxb/43.7.959
- Bannayan, M., Soler, C. M. T., Garcia, A. G. y., Guerra, L. C., & Hoogenboom, G. (2009). Interactive effects of elevated [CO2] and temperature on growth and development of a short- and long-season peanut cultivar. Climatic Change, 93(3–4), 389–406. https://doi.org/10.1007/s10584-008-9510-1
- Cai, C., Yin, X., He, S., Jiang, W., Si, C., Struik, P. C., Luo, W., Li, G., Xie, Y., Xiong, Y., & Pan, G. (2016). Responses of wheat and rice to factorial combinations of ambient and elevated CO2 and temperature in FACE experiments. Global Change Biology, 22(2), 856–874. https://doi.org/10.1111/gcb.13065
- Cheng, W., Sakai, H., Yagi, K., & Hasegawa, T. (2009). Interactions of elevated [CO2] and night temperature on rice growth and yield. Agricultural and Forest Meteorology, 149(1), 51–58. https://doi.org/10.1016/j.agrformet.2008.07.006
- De Temmerman, L., Vandermeiren, K., & Van Oijen, M. (2007). Chapter 19 - Response to the Environment: Carbon Dioxide. In D. Vreugdenhil, C. Gebhardt, D. K. L. Mackerron, & H. A. Rossc (Eds.), Potato Biology and Biotechnology (pp. 395–413). ELSEVIER. https://doi.org/10.1016/B978-044451018-1/50061-0
- De Temmerman, L., Wolf, J., Colls, J., Bindi, M., Fangmeier, A., Finnan, J., Ojanperä, K., & Pleijel, H. (2002). Effect of climatic conditions on tuber yield (Solanum tuberosum L.) in the European ‘CHIP’ experiments. European Journal of Agronomy, 17(4), 243–255. https://doi.org/10.1016/S1161-0301(02)00064-3
- Drake, B. G., Gonzàlez-Meler, M. A., & Long, S. P. (1997). MORE EFFICIENT PLANTS: A consequence of rising atmospheric CO2? Annual Review of Plant Biology, 48(1), 609–639. https://doi.org/10.1146/annurev.arplant.48.1.609
- e-STAT. (2020, May 2). Official Statistics of Japan. https://www.e-stat.go.jp.
- FAOSTAT. (2020, May 2). Food and Agriculture Organization of the United Nations. http://www.fao.org/faostat.
- Farrar, J. F., & Williams, M. L. (1991). The effects of increased atmospheric carbon dioxide and temperature on carbon partitioning, source-sink relations and respiration. Plant, Cell & Environment, 14(8), 819–830. https://doi.org/10.1111/j.1365-3040.1991.tb01445.x
- Figueiredo, N., Carranca, C., Trindade, H., Pereira, J., Goufo, P., Coutinho, J., Marques, P., Maricato, R., & de Varennes, A. (2015). Elevated carbon dioxide and temperature effects on rice yield, leaf greenness, and phenological stages duration. Paddy and Water Environment, 13(4), 313–324. https://doi.org/10.1007/s10333-014-0447-x
- Hoshikawa, K. (1980). Food Crop-New Compiled edition. Yokendo.
- Imai, K., Coleman, D. F., & Yanagisawa, T. (1984). Elevated atmospheric partial pressure of carbon dioxide and dry matter production of cassava (Manihot esculenta Crantz). Japanese Journal of Crop Science, 53(4), 479–485. https://doi.org/10.1626/jcs.53.479
- Imai, K., Coleman, D. F., & Yanagisawa, T. (1985). Increase in atmospheric partial pressure of carbon dioxide and growth and yield of rice (Oryza sativa L.). Japanese Journal of Crop Science, 54(4), 413–418. https://doi.org/10.1626/jcs.54.413
- IPCC. (2018). Summary for Policymakers. In: Global Warming of 1.5°C. An IPCC Special Report on the impacts of global warming of 1.5°C above pre-industrial levels and related global greenhouse gas emission pathways, in the context of strengthening the global response to the threat of climate change, sustainable development, and efforts to eradicate poverty [Masson-Delmotte, V., P. Zhai, H.-O. Pörtner, D. Roberts, J. Skea, P.R. Shukla, A. Pirani, W. Moufouma-Okia, C. Péan, R. Pidcock, S. Connors, J.B.R. Matthews, Y. Chen, X. Zhou, M.I. Gomis, E. Lonnoy, T. Maycock, M. Tignor, and T. Waterfield (eds.)] (pp. 32). World Meteorological Organization.
- Jing, L., Wang, J., Shen, S., Wang, Y., Zhu, J., Wang, Y., & Yang, L. (2016). The impact of elevated CO2 and temperature on grain quality of rice grown under open-air field conditions. Journal of the Science of Food and Agriculture, 96(11), 3658–3667. https://doi.org/10.1002/jsfa.7545
- Kimball, B. A. (1983). Carbon dioxide and agricultural yield: An assemblage and analysis of 430 prior observations. Agronomy Journal, 75(5), 779–788. https://doi.org/10.2134/agronj1983.00021962007500050014x
- Kimball, B. A. (2016). Crop responses to elevated CO2 and interactions with H2O, N, and temperature. Current Opinion in Plant Biology, 31, 36–43. https://doi.org/10.1016/j.pbi.2016.03.006
- Kimball, B. A., Kobayashi, K., & Bindi, M. (2002). Responses of agricultural crops to free-air CO2 enrichment. In D. L. Sparks (Ed.), Advances in Agronomy (Vol. 77, pp. 293–368). ELSEVIER. https://doi.org/10.1016/S0065-2113(02)77017-X.
- Long, S. P., Ainsworth, E. A., Rogers, A., & Ort, D. R. (2004). Rising atmospheric carbon dioxide: Plants FACE the future. Annual Review of Plant Biology, 55(1), 591–628. https://doi.org/10.1146/annurev.arplant.55.031903.141610
- Miglietta, F., Magliulo, V., Bindi, M., Cerio, L., Vaccari, F. P., Loduca, V., & Peressotti, A. (1998). Free Air CO2 Enrichment of potato (Solanum tuberosum L.): Development, growth and yield. Global Change Biology, 4(2), 163–172. https://doi.org/10.1046/j.1365-2486.1998.00120.x
- Mina, U., Kumar, R., Gogoi, R., Bhatia, A., Harit, R. C., Singh, D., Kumar, A., & Kumar, A. (2019). Effect of elevated temperature and carbon dioxide on maize genotypes health index. Ecological Indicators, 105, 292–302. https://doi.org/10.1016/j.ecolind.2017.08.060
- Nakagawa, H., & Horie, T. (2000). Rice responses to elevated CO2 and temperature. Global Environmental Research, 3(2), 101–114. http://www.airies.or.jp/journal_03-2eng.html
- Naznin, M. T., Kitaya, Y., Shibuya, T., Endo, R., Hirai, H., & Lefsrud, M. G. (2015). Ground based study on culturing garlic as a source of vegetable food and medicine in space-growth and ajoene accumulation in garlic plants cultured with different CO2 regimes. Biological Sciences in Space, 29, 1–7. https://doi.org/10.2187/bss.29.1
- Newman, Y. C., Sollenberger, L. E., Boote, K. J., Allen Jr., L. H., Vu, J. C. V., & Hall, M. B. (2005). Temperature and carbon dioxide effect on nutritive value of rhizome peanut herbage. Crop Science, 45(1), 316–321. https://doi.org/10.2135/cropsci2005.0316
- NOAA. (2020). National oceanic & atmospheric administration- Earth system research laboratory. Trends in atmospheric carbon dioxide. https://www.esrl.noaa.gov/gmd/ccgg/trends
- Ottman, M. J., Kimball, B. A., White, J. W., & Wall, G. W. (2012). Wheat growth response to increased temperature from varied planting dates and supplemental infrared heating. Agronomy Journal, 104(1), 7–16. https://doi.org/10.2134/agronj2011.0212
- Palacios, C. J., Grandis, A., Carvalho, V. J., Salatino, A., & Buckeridge, M. S. (2019). Isolated and combined effects of elevated CO2 and high temperature on the whole-plant biomass and the chemical composition of soybean seeds. Food Chemistry, 275, 610–617. https://doi.org/10.1016/j.foodchem.2018.09.052
- Paul, M. J., & Foyer, C. H. (2001). Sink regulation of photosynthesis. Journal of Experimental Botany, 52(360), 1383–1400. https://doi.org/10.1093/jexbot/52.360.1383
- Roy, K. S., Bhattacharyya, P., Neogi, S., Rao, K. S., & Adhya, T. K. (2012). Combined effect of elevated CO2 and temperature on dry matter production, net assimilation rate, C and N allocations in tropical rice (Oryza sativa L.). Field Crops Research, 139, 71–79. https://doi.org/10.1016/j.fcr.2012.10.011
- Ruiz-Vera, U. M., Siebers, M. H., Drag, D. W., Ort, D. R., & Bernacchi, C. J. (2015). Canopy warming caused photosynthetic acclimation and reduced seed yield in maize grown at ambient and elevated [CO2]. Global Change Biology, 21(11), 4237–4249. https://doi.org/10.1111/gcb.13013
- Sage, R. F., Sharkey, T. D., & Seemann, J. R. (1989). Acclimation of photosynthesis to elevated CO2 in five C3 species. Plant Physiology, 89(2), 590–596. https://doi.org/10.1104/pp.89.2.590
- Sakai, H., Tokida, T., Usui, Y., Nakamura, H., & Hasegawa, T. (2019). Yield responses to elevated CO2 concentration among Japanese rice cultivars released since 1882. Plant Production Science, 22(3), 352–366. https://doi.org/org/10.1080/1343943X.2019.1626255
- Saminathan, T., Alvarado, A., Lopez, C., Shinde, S., Gajanayake, B., Abburi, V. L., Vajja, V. G., Jagadeeswaran, G., Reddy, K. R., Nimmakayala, P., & Reddy, U. K. (2019). Elevated carbon dioxide and drought modulate physiology and storage-root development in sweet potato by regulating microRNAs. Functional & Integrative Genomics, 19(1), 171–190. https://doi.org/10.1007/s10142-018-0635-7
- Schapendonk, A. H. C. M., Oijen, M. V., Dijkstra, P., Pot, C. S., Jordi, W. J. R. M., & Stoopen, G. M. (2000). Effects of elevated CO2 concentration on photosynthetic acclimation and productivity of two potato cultivars grown in open-top chambers. Functional Plant Biology, 27(2), 1119–1130. https://doi.org/10.1071/PP99205
- Shimono, H., Okada, M., Yamakawa, Y., Nakamura, H., Kobayashi, K., & Hasegawa, T. (2008). Rice yield enhancement by elevated CO2 is reduced in cool weather. Global Change Biology, 14(2), 276–284. https://doi.org/10.1111/j.1365-2486.2007.01498.x
- Stitt, M. (1991). Rising CO2 levels and their potential significance for carbon flow in photosynthetic cells. Plant, Cell & Environment, 14(8), 741–762. https://doi.org/10.1111/j.1365-3040.1991.tb01440.x
- Uprety, D. C., Dwivedi, N., Jain, V., & Mohan, R. (2002). Effect of elevated carbon dioxide concentration on the stomatal parameters of rice cultivars. Photosynthetica, 40(2), 315–319. https://doi.org/10.1023/A:1021322513770