
ABSTRACT
Information in visual working memory that is only prospectively relevant can nevertheless guide attention towards memory matching visual input. Previous studies demonstrated that such memory-based attentional biases can be modulated by top-down processing strategies. Here we examined whether attentional capture by memory matching distractors is also modulated by more implicit biases stemming from selection history. Observers performed a visual search task while holding a colour in memory for a subsequent task. Crucially, a coloured distractor in the search display not only matched the memory content half of the time, it also appeared on one location more often than on all other locations. Consistent with statistical learning having a strong impact on attentional priorities, attentional capture by the distractor was attenuated at high probability distractor locations. The additional slowing by memory matching distractors, however, was the same at suppressed and non-suppressed locations. We interpret this finding as evidence that memory-based feature biases are independent from learned spatial biases.
The overwhelming amount of visual information that we encounter everyday by far surpasses the processing capacity of our visual system. Consequently, perceptual processing is highly competitive, a competition that can be influenced by various factors. In addition to stimulus-specific differences which may inherently create a competitive advantage regardless of task-relevance (e.g., a salient colour in a visual scene), top-down selection goals can bias perceptual processing towards relevant visual input (Egeth & Yantis, Citation1997; Theeuwes, Citation2010, Citation2018; Yantis & Egeth, Citation1999). An influential model of attentional selection, the biased competition model, proposes that maintaining task-relevant information within visual working memory (VWM) changes weights in the attentional priority map, which inherently creates a perceptual bias towards matching stimuli (Desimone & Duncan, Citation1995; Zelinsky & Bisley, Citation2015). The idea here is that memory driven sensory pre-activation prior to new visual input creates a competitive advantage for memory matching stimuli over other visual stimuli.
Evidence that VWM is a driving force behind attentional selection stems from dual-task paradigms in which observers perform a perceptual task, while holding information in VWM. In these paradigms the content of VWM is irrelevant to the perceptual task, and often-times even interferes with selection when a distractor in the display matches the memory content. Even though attentional selection of the memory match in such a context is counterproductive, it has been shown that relative to neutral distractors memory matches attract more attention in visual search tasks (e.g., Olivers, Meijer, & Theeuwes, Citation2006b; Soto, Heinke, Humphreys, & Blanco, Citation2005; van Moorselaar, Theeuwes, & Olivers, Citation2014) and attract more eye movements, both in anticipation of (van Loon, Olmos-Solis, & Olivers, Citation2017), and during visual search (Hollingworth & Luck, Citation2009; Hollingworth, Matsukura, & Luck, Citation2013; Olivers et al., Citation2006b; Silvis & Van der Stigchel, Citation2014). Such interference suggests that attentional biases towards memory matching visual input occur automatically.
However, other findings question the automaticity of the interplay between working memory content and attentional selection. It has been shown that attentional capture by memory matching distractors can be modulated by temporal demands of the visual search display (Dalvit & Eimer, Citation2011) and the probability that memory matches coincide with targets vs. distractors (Carlisle & Woodman, Citation2011; Kiyonaga, Egner, & Soto, Citation2012). Also, memory-based attentional capture can be eliminated when the location of the target is cued in advance (Pan & Soto, Citation2010) or when target identity varies on a trial by trial basis (Olivers, Citation2009; but see Bahle, Beck, & Hollingworth, Citation2018). These findings illustrate that the attentional bias towards memory matching visual input is not primarily an automatic phenomenon, but instead can be modulated by strategic factors associated with the attentional state of the observer.
Recently, it was recognized that activity within priority maps is not only controlled by top-down or bottom-up processes but can also be influenced by the “history” of former attentional deployment (Awh, Belopolsky, & Theeuwes, Citation2012). As a specific case, it is proposed that statistical regularities within a visual search display help to shape weights within priority maps such that frequent target and distractor locations become respectively facilitated and suppressed (Jiang, Sha, & Remington, Citation2015; Jiang, Swallow, & Rosenbaum, Citation2013; Wang & Theeuwes, Citation2018a, Citation2018b). To test how statistical regularities modulate distractor filtering, studies have manipulated the configuration of visual search displays such that the singleton distractor was more frequently presented on one specific location (Ferrante et al., Citation2017; Reder, Weber, Shang, & Vanyukov, Citation2003; Wang & Theeuwes, Citation2018a, Citation2018b, Citation2018c). Consistent with an adaptive decreased priority on the frequent distractor location relative to all other locations, these studies not only observed reduced distractor interference but also less efficient target selection at the highly probable distractor location relative to all other locations.
To increase our understanding of the interplay between working memory and attention, here we investigated whether memory-based attentional capture can also be modulated when the attentional state of the observer is not implemented top-down but stems from selection history. For this purpose, we adopted the paradigm used in our previous work to study memory-based attentional capture but manipulated the location of the coloured distractor singleton such that it appeared on one location more often than on all other locations (). If learned spatial biases and memory-based feature biases operate independently we should observe a general reduction of attentional capture by singleton distractors, but the additional slowing by memory-matching distractors should be unaffected. However, if working memory effects on perceptual selection per se are not only modulated by explicit, but also by more implicit biases, we should observe that memory-based attentional capture is reduced or even eliminated when the distractor is presented on the highly probable location.
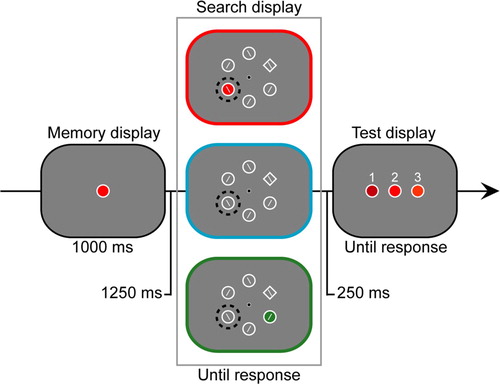
Figure 1. Sequence of events in a trial. All possible distractor conditions, related (top), unrelated (bottom) and no (middle) are displayed. The black dotted circles in the search display indicate the high probability location (counterbalanced across participants). Note that the outlines of the search display boxes correspond to the violins in the violin graphs in .
Experiment 1: memory-based feature biases vs. learned spatial biases
Methods
Participants
A planned number of 18 participants (M = 24, range 19–31, 6 men), recruited via the subject pool of VU University participated in this experiment. Participants provided informed consent approved by the local ethics committee before participating and were compensated with either course credits or monetary reward upon completion of the experiment. All participants had normal or corrected to normal vision.
Apparatus, stimuli, procedure, and design
Stimuli were generated on a dark grey background via OpenSesame version 3.1 (Mathot, Schreij, & Theeuwes, Citation2012) with a Psychopy backend (Peirce, Citation2007) running on a windows 7 PC and presented on a 22-inch screen (1680 × 1050, refreshing at120 Hz) in a dimly lit room. Viewing distance was set at 75 cm and controlled via a chinrest.
The paradigm was modelled after our previous VWM capture experiments (Olivers, Meijer, & Theeuwes, Citation2006a; van Moorselaar et al., Citation2014). Trials started with a 250 ms empty screen followed by a 250 ms black fixation circle with a white outline. The subsequent memory display was presented for 1000 ms, after which a 1250 ms fixation display was shown followed by a search display, until response. The fixation display was then shown again for 250 ms, or, in case of an error during the search, the word “incorrect” displayed in red. Each trial ended with a memory test which was again visible until response ().
The memory displayed contained a single coloured disk (radius 0.9°) with a white outline presented at fixation. The disks colour was selected at random from a pool of five colour categories, each containing nine different exemplars with varying hue and chroma on the basis of the Munsell colour system (Munsell, Citation1929), with the restriction that a colour category was never repeated on subsequent trials.
Each search display contained five distractor disks (radius 0.9°) and one diamond-shaped target (2.6° by 2.6°), all with a white outline and a left or right tilted white bar (width = 3 pixels) in their centre, placed on an imaginary circle (radius 3.9°) centred on fixation. On 2/3 of the trials the grey background of one of the distractor disks was replaced by a colour from the memory colour pool. This resulted in three distractor type conditions. In the related distractor condition the colour of the distractor was identical to the to-be memorized colour on that trial. In the unrelated distractor condition the colour was selected randomly from the other colour categories. In the no distractor condition the search display contained no coloured disks. Although the coloured singleton could appear in all of six stimulus positions, following Benchi Wang and Theeuwes (Citation2018b) it was 8 times more likely to appear on one particular location (61.5%; location counterbalanced) than any of the other locations in the display (7.7%). The target location was selected at random such that across conditions the target and the distractor could be presented on high and low probable locations.
The trial sequence ended with a forced choice recognition task, in which participants had to select the memory matching colour from a horizontal configuration of three coloured disks. The two non-matching disks were of a different colour but selected from the same colour category as the memory colour.
All participants completed 39 practice trials and 10 experimental blocks of 39 trials each. Within an experimental block, each singleton distractor type condition was present 13 times, randomly mixed. In between blocks participants were encouraged to take a short break, while they received feedback on RTs and accuracy in the search task and accuracy in the memory task. Feedback was provided to keep participants engaged with the task and motivate them to keep the amount of errors to a minimum. At the end of the experiment participants were asked whether they noticed that one location contained a distractor with higher probability than other locations, and if so to indicate which location was the high probability location.
Results
Search RTs
Three out of eight participants that said they noticed that one location contained the distractor more often, actually indicated the correct location. The other six participants noticed no systematic modulation (data of the other four participants was not collected), showing that any effects are unlikely to result from conscious control or top-down strategy.
Following our previous memory-based attentional capture experiments, RT analysis only included correct search responses, which led to the exclusion of 2.8% of trials. Next RT data were filtered with a two-step trimming procedure (Olivers et al., Citation2006a; van Moorselaar et al., Citation2014); only trials with an RT between 200 and 5000 ms were selected, which were then trimmed on the basis of a cutoff value of 2.5 standard deviations from the mean RT per participant (2.4%), resulting in an overall loss of 5.1% of trials.
First, we examined how the attentional capture effect was modulated by the probability manipulation ( and ). A repeated measure analysis of variance (rANOVA) limited to trials without a coloured distractor and unrelated distractors presented either on the high or the low probability distractor location showed a main effect, F (2, 34) = 20.18, p < .01. Replicating Wang and Theeuwes (Citation2018a, Citation2018b) planned comparisons demonstrated that, although there was significant attentional capture on both the low (M = 30, SD = 19), t (17) = 6.66, p < .001, and the high probability location (M = 15, SD = 19), t (17) = 3.41, p < .01, RTs were significantly reduced for the high relative to the low probability locations, t (17) = 2.86, p = .011. Consistent with the idea that the high probability location was actively suppressed, in no distractor trials search times were slower when the target was presented at a high probability location (M = 624, SD = 110) than at a low probability location (M = 582, SD = 74, t (17) = 3.39, p < .01), again replicating Wang and Theeuwes (Citation2018a, Citation2018b).
Figure 2. Search RTs Experiment 1 as a function of distractor condition shown separately for the low probability distractor location (left) and the high probability distractor location (right). Blue dotted line shows the average RT in the no distractor condition.
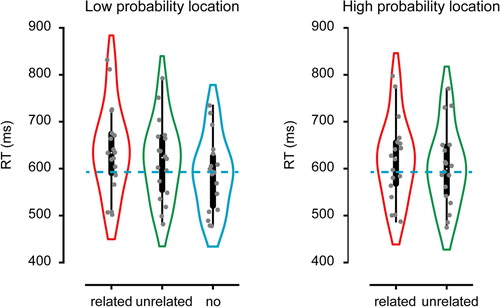
Table 1. Data columns represent mean of average search RTs in ms on correct trials, mean accuracy on search task and mean accuracy on memory task for each condition
Having established that attentional capture by a coloured singleton distractor was reduced when that distractor was presented on a high probability location, we tested whether memory-based attentional capture was also attenuated by the probability manipulation. For this purpose, we entered RTs into a rANOVA with within-subjects factors distractor type (related, unrelated) and distractor location (high probability, low probability). Consistent with our previous findings, we observed a main effect of distractor type reflecting an additional slowing of RTs by memory matching distractors relative to unrelated distractors, F (1, 17) = 15.68, p < .001. Also, overall RTs were significantly reduced when the coloured distractor was presented at the high probability location, F (1, 17) = 36.94, p < .001. Importantly, however, there was no evidence that the additional slowing by a memory matching distractor was attenuated at the high probability location (M = 22, SD = 25 at low probability location; M = 16, SD = 25 at high probability location; F = 0.84, p = .37).Footnote1 Bayesian statistics using JASP (JASP TEAM, Citation2018) showed that the data were 2.27 times more likely under the two main effects model than under the model that added the interaction.
Accuracy
There were no significant effects on accuracy measures, nor on accuracy during search nor during the memory test ().
In conclusion, the data provide no evidence that memory-driven distractor interference is modulated by the learned spatial properties of the display, as the effects of spatial position and of the feature match did not interact. However, as the Bayes factor provides only weak evidence against the null hypothesis (Wetzels et al., Citation2011), it precludes us from drawing firm conclusions. To strengthen our conclusions, we therefore include the relevant findings of a recent large-scale replication study incorporating the same two factors as reported here, in Experiment 2.
Experiment 2: a replication study
To corroborate the findings in Experiment 1, we present data from a larger replication project investigating the reliability of various memory-driven attentional capture phenomena (Fratescu, van Moorselaar, & Mathot, Citation2018), and which included a similar probability manipulation. Other manipulations were included (most notably memory load) but will be reported on elsewhere. Importantly, four times as many participants as in Experiment 1 took part, thus considerably increasing the power with regards to potential null effects.
Methods
The method was the same as in Experiment 1 except for the following changes. Sixty-six participants took part in the experiment under the supervision of 20 students as part of a course. The experiment was approved by ethics review board of the Department of Psychology of the University of Groningen (#17364-O), participation was voluntary and participants did not receive compensation. Students used their own computers for data collection. Students received an explanation of what constitutes a suitable environment for testing (i.e., a calm environment without distractions), but we had no control over the actual testing environments. Participants provided verbal informed consent prior to participation, and were allowed to (and frequently did) abort the experiment whenever they wanted.
Each trial started with a 750 ms black fixation cross. The memory display contained one (load 1) or two (load 2) coloured squares (50 pix in diameter) placed at a random subset of four possible locations centred on the intercardinal axes (200 pix from fixation). Each search display contained seven instead of five white disk-shaped distractors. The radius of the imaginary circle was also 200 pix such that the location of the distractors at the intercardinal locations matched the four possible memory locations. The location of the target was randomly selected, with the restriction that it was never displayed on one of the memory locations of that trial. On 3/4 of the trials the grey background of one of the distractor disks, located at one of the memory locations of that trial, was replaced by a coloured disk. In the related condition the distractor colour could match the colour that was presented on that location on the preceding memory display, or it matched one of the memory colours but now it appeared at the position of one of the other memory colours (In case of load 1, this meant that the distractor was placed on an empty memory location). Although the coloured singleton could appear in all of four possible memory locations, it was six times more likely to appear on one particular location (66.7%; location counterbalanced) than any of the other memory locations (11.1%).
The forced choice recognition task was replaced by a change detection task, in which coloured squares were presented on the memory-locations of that trial. One-half of the trials one of the memory colours had changed to another exemplar from the same colour category. A one-pixel thick line cued one square, and participants were instructed to indicate with a key press whether the cued square changed or was identical to the memory item (“C” for change and “N” for change).
All participants completed 15 practice trials and 10 experimental blocks of 48 trials each. Within an experimental block, each Distractor Condition was present six times for each of the memory loads, randomly mixed, resulting in 30 observations per condition.
Results
Search RTs
Six out of sixty-six participants were excluded because their average filtered RT, search accuracy or memory accuracy deviated more than 2.5 SD from the grand mean.Footnote2 In the remaining 60 participants, exclusion of incorrect searches (2.6%) and filtering with the two-step trimming procedure (2.8%) resulted in an overall loss of 5.4% of trials.
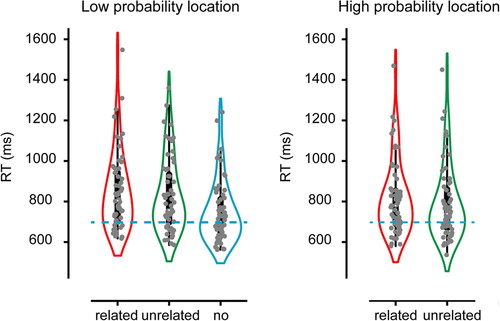
Replicating Experiment 1 and Wang and Theeuwes (Citation2018a, Citation2018b) planned comparisons demonstrated that RTs to neutral distractors were significantly reduced for high relative to low distractor locations, t (59) = 4.75, p < .001. Despite this reduction, however, as in Experiment 1 attentional capture at high probability locations remained reliable (M = 15, SD = 57), t (59) = 5.99, p < .001. Importantly, the rANOVA examining whether memory-based attentional capture was also attenuated by the probability manipulation, again yielded firm main effects of distractor type F (1, 59) = 23.27, p < .001 and distractor location F (1, 59) = 36.43, p < .001, but no interaction (F = 0.71, p = .40). Bayesian analysis confirmed that the data were 3.78 times more likely under the two main effects model than under the model that added the interaction, and this factor increased to 5.29 when the data of both experiments was combined. The replication study thus provided substantial evidence against the hypothesis that memory-based feature and learned spatial biases shape the priority map interactively (Wetzels et al., Citation2011) ().
Figure 3. Search RTs Experiment 2 as a function of distractor condition shown separately for the low probability distractor location (left) and the high probability distractor location (right). Blue dotted line shows the average RT in the no distractor condition.
Discussion
Previous studies have demonstrated that the content of VWM biases attention towards memory matching stimuli, even when that stimulus interferes with the task at hand (e.g., Olivers et al., Citation2006a; Soto et al., Citation2005; van Moorselaar et al., Citation2014). At the same time, a recent line of experiments has shown that statistical regularities across a sequence of visual searches can bias attention away from distractor locations (Ferrante et al., Citation2017; Reder et al., Citation2003; Wang & Theeuwes, Citation2018a, Citation2018b). The present study was designed to examine whether these attentional biases operate independently or whether memory-based attentional capture can be modulated by statistical learning of distractor locations. Consistent with statistical learning having a strong impact on attentional priorities, attentional capture by coloured singletons was attenuated at high probability distractor locations. The additional slowing by memory matching distractors, however, was unaffected by learned spatial suppression. This demonstrates that memory-based feature biases are independent from learned spatial biases.
The learning-based attenuation of attentional capture by neutral distractors is consistent with the idea that through statistical learning weights within the priority map are selectively reduced at the location that frequently contains a distractor (Ferrante et al., Citation2017; Theeuwes, Citation2018; Wang & Theeuwes, Citation2018a, Citation2018b) Consequently, stimuli at that location compete less strongly for attention, which is beneficial when a distractor appears, but, as also observed here, is actually counterproductive when a target happens to be presented at a low priority location. Recently, Stilwell, Bahle, and Vecera (Citation2018) demonstrated that despite such local suppression the visual system continues to extract feature information. In their experimental paradigm feature-based regularities attenuated attentional capture even when the singleton distractor location was completely predictable. While in that study location- and feature-based information cooperatively reduced attentional capture, here we show that feature-based biases also survive learned spatial biases when they interfere rather than facilitate visual search.
The observed spatial suppression is also in line with the “signal suppression” hypothesis advocated by Gaspelin and Luck (Citation2018). In their reasoning all salient stimuli generate an “attend to me signal”, but capture can sometimes be avoided via inhibitory mechanisms. This idea is supported by a growing body of evidence demonstrating that salient distractors that do not capture attention elicit a Pd, an ERP marker of suppression, rather than an N2pc (indexing attentional allocation; e.g., Eimer & Kiss, Citation2008; Sawaki & Luck, Citation2011). Interestingly, Sawaki and Luck (Citation2011) demonstrated that memory-matching probes presented during the delay interval also elicit a Pd, rather than an N2pc, suggesting that an active suppression mechanism can prevent the “automatic” attentional capture by working memory matches. Although we can only speculate about the ERP components underlying our pattern of results, and Sawaki and Luck (Citation2011) provided no insight in how memory-matching probes influenced task performance, the Pd in response to memory-matching probes appears to be at odds with our finding that memory-based attentional biases survive spatial suppression. A potential explanation for this discrepancy might be that memory-based attentional biases are harder to suppress with increased perceptual competition (van Moorselaar et al., Citation2018). While in our experiment the target was embedded within multiple item search display, perceptual competition was minimal in the Sawaki and Luck (Citation2011) experiment as their probe display only contained two lateralized circles (but see van Loon et al., Citation2017). Future research, however, is necessary to establish the exact conditions under which attentional capture by memory matching can be prevented. The present results add to this literature by demonstrating that memory-based attentional biases are not modulated when the attentional state of the observer is learned implicitly rather than implemented top-down.
Acknowledgements
DvM contributed to design, collected the data, performed the analyses, and contributed most of the writing. Both CNLO and JT supervised the project and were involved in design and writing.
Disclosure statement
No potential conflict of interest was reported by the authors.
Additional information
Funding
Notes
1 As suggested by an anonymous reviewer we repeated the analysis after excluding all trials where the distractor colour category matched the colour category of the memoranda (24.8%) or the unrelated distractor from the preceding trial (0.5%). This resulted in the same pattern of results.
2 Note that this trimming procedure is slightly different from the procedure reported in Fratescu et al. (Citation2018). We chose to use the same procedure as reported in our previous work (Olivers et al., Citation2006b; van Moorselaar et al., Citation2014). The reported results, however, were robust to different trimming procedures.
References
- Awh, E., Belopolsky, A. V., & Theeuwes, J. (2012). Top-down versus bottom-up attentional control: A failed theoretical dichotomy. Trends in Cognitive Sciences, 16(8), 437–443. doi: 10.1016/j.tics.2012.06.010
- Bahle, B., Beck, V. M., & Hollingworth, A. (2018). The Architecture of interaction between visual working memory and visual attention. Journal of Experimental Psychology: Human Perception and Performance. doi: 10.1037/xhp0000509
- Carlisle, N. B., & Woodman, G. F. (2011). Automatic and strategic effects in the guidance of attention by working memory representations. Acta Psychologica, 137(2), 217–225. doi: 10.1016/j.actpsy.2010.06.012
- Dalvit, S., & Eimer, M. (2011). Memory-driven attentional capture is modulated by temporal task demands. Visual Cognition, 19(2), 145–153. doi: 10.1080/13506285.2010.543441
- Desimone, R., & Duncan, J. (1995). Neural mechanisms of selective visual attention. Annual Review of Neuroscience. doi: 10.1146/annurev.ne.18.030195.001205
- Egeth, H. E., & Yantis, S. (1997). Visual attention: Control, representation, and time course. Annual Review of Psychology. doi: 10.1146/annurev.psych.48.1.269
- Eimer, M., & Kiss, M. (2008). Involuntary attentional capture is determined by task set: Evidence from event-related brain potentials. Journal of Cognitive Neuroscience. doi: 10.1162/jocn.2008.20099
- Ferrante, O., Patacca, A., Di Caro, V., Della Libera, C., Santandrea, E., & Chelazzi, L. (2017). Altering spatial priority maps via statistical learning of target selection and distractor filtering. Cortex. doi: 10.1016/j.cortex.2017.09.027
- Fratescu, M., van Moorselaar, D., & Mathot, S. (2018). Can you have multiple attentional templates? Large-scale replications of Van Moorselaar, Theeuwes and Olivers (2014) and Hollingworth and Beck (2016). BioRxiv, 474932.
- Gaspelin, N., & Luck, S. J. (2018). The role of inhibition in avoiding distraction by salient stimuli. Trends in Cognitive Sciences, 22(1), 79–92. doi: 10.1016/j.tics.2017.11.001
- Hollingworth, A., & Luck, S. J. (2009). The role of visual working memory (VWM) in the control of gaze during visual search. Attention, Perception, and Psychophysics. doi: 10.3758/APP.71.4.936
- Hollingworth, A., Matsukura, M., & Luck, S. J. (2013). Visual working memory Modulates Rapid Eye movements to Simple Onset targets. Psychological Science, 24(5), 790–796. doi: 10.1177/0956797612459767
- JASP TEAM. (2018). JASP (Version 0.8) [Mac OS]. Retrieved from https://jasp-stats.org/
- Jiang, Y. V., Sha, L. Z., & Remington, R. W. (2015). Modulation of spatial attention by goals, statistical learning, and monetary reward. Attention, Perception, and Psychophysics. doi: 10.3758/s13414-015-0952-z
- Jiang, Y. V., Swallow, K. M., & Rosenbaum, G. M. (2013). Guidance of spatial attention by incidental learning and endogenous cuing. Journal of Experimental Psychology: Human Perception and Performance. doi: 10.1037/a0028022
- Kiyonaga, A., Egner, T., & Soto, D. (2012). Cognitive control over working memory biases of selection. Psychonomic Bulletin and Review, 19(4), 639–646. doi: 10.3758/s13423-012-0253-7
- Mathot, S., Schreij, D., & Theeuwes, J. (2012). Opensesame: An open-source, graphical experiment builder for the social sciences. Behavior Research Methods. doi: 10.3758/s13428-011-0168-7
- Munsell, A. H. (1929). Munsell book of color. Baltimore, MD: Munsell Color Company.
- Olivers, C. N. L. (2009). What drives memory-driven attentional capture? The effects of memory type, display type, and search type. Journal of Experimental Psychology. Human Perception and Performance, 35(5), 1275–1291. doi: 10.1037/a0013896
- Olivers, C. N. L., Meijer, F., & Theeuwes, J. (2006a). Feature-based memory-driven attentional capture: Visual working memory content affects visual attention. Journal of Experimental Psychology. Human Perception and Performance, 32(5), 1243–1265. doi: 10.1037/0096-1523.32.5.1243
- Olivers, C. N. L., Meijer, F., & Theeuwes, J. (2006b). Feature-based memory-driven attentional capture: Visual working memory content affects visual attention. Journal of Experimental Psychology: Human Perception and Performance, 32(5), 1243–1265. doi: 10.1037/0096-1523.32.5.1243
- Pan, Y., & Soto, D. (2010). The modulation of perceptual selection by working memory is dependent on the focus of spatial attention. Vision Research, 50(15), 1437–1444. doi: 10.1016/j.visres.2009.10.016
- Peirce, J. W. (2007). PsychoPy – psychophysics software in python. Journal of Neuroscience Methods. doi: 10.1016/j.jneumeth.2006.11.017
- Reder, L. M., Weber, K., Shang, J., & Vanyukov, P. M. (2003). The adaptive Character of the attentional system: Statistical Sensitivity in a target Localization task. Journal of Experimental Psychology: Human Perception and Performance. doi: 10.1037/0096-1523.29.3.631
- Sawaki, R., & Luck, S. J. (2011). Active suppression of distractors that match the contents of visual working memory. Visual cognition, 19(7), 956–972. doi: 10.1080/13506285.2011.603709
- Silvis, J. D., & Van der Stigchel, S. (2014). How memory mechanisms are a key component in the guidance of our eye movements: Evidence from the global effect. Psychonomic Bulletin and Review. doi: 10.3758/s13423-013-0498-9
- Soto, D., Heinke, D., Humphreys, G. W., & Blanco, M. J. (2005). Early, involuntary top-down guidance of attention from working memory. Journal of Experimental Psychology: Human Perception and Performance, 31(2), 248–261. doi: 10.1037/0096-1523.31.2.248
- Stilwell, B. T., Bahle, B., & Vecera, S. P. (2018). Feature-based statistical regularities of distractors modulate attentional capture. Journal of Experimental Psychology: Human Perception and Performance. doi:10.1037/xhp0000613
- Theeuwes, J. (2010). Top-down and bottom-up control of visual selection. Acta Psychologica. doi: 10.1016/j.actpsy.2010.02.006
- Theeuwes, J. (2018). Visual selection: Usually fast and automatic; seldom slow and volitional. Journal of Cognition. doi: 10.5334/joc.13
- van Loon, A. M., Olmos-Solis, K., & Olivers, C. N. L. (2017). Subtle eye movement metrics reveal task-relevant representations prior to visual search. Journal of Vision. doi: 10.1167/17.6.13
- van Moorselaar, D., Gayet, S., Paffen, C. L. E., Theeuwes, J., Van der Stigchel, S., & Olivers, C. N. L. (2018). Competitive interactions in visual working memory drive access to awareness. Cortex, 102, 6–13. doi: 10.1016/j.cortex.2017.03.026
- van Moorselaar, D., Theeuwes, J., & Olivers, C. N. L. (2014). In competition for the attentional template: Can multiple items within visual working memory guide attention? Journal of Experimental Psychology: Human Perception and Performance, 40(4), 1450–1464. doi: 10.1037/a0036229
- Wang, B., & Theeuwes, J. (2018a). How to inhibit a distractor location? Statistical learning versus active, top-down suppression. Attention, Perception, and Psychophysics. doi: 10.3758/s13414-018-1493-z
- Wang, B., & Theeuwes, J. (2018b). Statistical regularities modulate attentional capture. Journal of Experimental Psychology: Human Perception and Performance. doi: 10.1037/xhp0000472
- Wang, B., & Theeuwes, J. (2018c). Statistical regularities modulate attentional capture independent of search strategy. Attention, Perception, & Psychophysics, 80(7), 1763–1774. doi: 10.3758/s13414-018-1562-3
- Wetzels, R., Matzke, D., Lee, M. D., Rouder, J. N., Iverson, G. J., & Wagenmakers, E.-J. (2011). Statistical evidence in experimental psychology. Perspectives on Psychological Science. doi: 10.1177/1745691611406923
- Yantis, S., & Egeth, H. E. (1999). On the distinction between visual salience and stimulus-driven attentional capture. Journal of Experimental Psychology: Human Perception and Performance. doi: 10.1037/0096-1523.25.3.661
- Zelinsky, G. J., & Bisley, J. W. (2015). The what, where, and why of priority maps and their interactions with visual working memory. Annals of the New York Academy of Sciences. doi: 10.1111/nyas.12606