
 ?Mathematical formulae have been encoded as MathML and are displayed in this HTML version using MathJax in order to improve their display. Uncheck the box to turn MathJax off. This feature requires Javascript. Click on a formula to zoom.
?Mathematical formulae have been encoded as MathML and are displayed in this HTML version using MathJax in order to improve their display. Uncheck the box to turn MathJax off. This feature requires Javascript. Click on a formula to zoom.ABSTRACT
This research investigated how interactive social contexts shape basic visual attention. It has been shown that social information can modulate inhibition of return effects in joint spatial cueing tasks. We predicted that if perceptions of cooperativeness explain this phenomenon, we would then observe larger inhibition of return effects for more cooperative individuals and in highly cooperative contexts. Experiments 1a and 1b found larger inhibition of return effects and greater perceptions of cooperativeness for female compared to male participants, consistent with the literature on gender stereotypes and the behavioural evidence that females are more cooperative than males. In Experiment 2a and 2b, we experimentally manipulated the cooperativeness of the task, describing it as either a team or an individual game. This time, we found larger inhibition of return effects and greater perceptions of cooperativeness for male participants in the team compared to the individual game. We conclude that construing interactive contexts as cooperative plays an important role in the joint spatial orienting of visual attention, and we propose this as an example of socially distributed cognition.
Humans do not perceive social reality objectively, but rather they construe it subjectively (e.g., Fiske & Taylor, Citation2013). One reason for this is that human cognition is limited in the amount of information it can process. Thus, pre-existing knowledge structures, motivation, and affect guide attention to the most relevant stimuli in the environment, which are then prioritised for information processing (e.g., Fiske & Neuberg, Citation1990). For example, social information can endogenously orient our attention facilitating coordination and cooperation (e.g., Richardson & Gobel, Citation2015). From this perspective, visual attention can be seen as a key mechanism through which social reality is experienced.
Going beyond the traditional view of visual attention as an encoding mechanism, we have recently shown that when looking at and looking with another person, humans can also signal information back into the world (Gobel et al., Citation2015, Citation2017). Such reciprocal social attention is a striking example of interactive cognition, as it receives and signals information from and to others (Richardson & Gobel, Citation2015). Whilst past social attention research has predominantly focused on how characteristics of social stimuli (e.g., an interaction partner) change participants' visual attention, in the present research, we aimed to increase our understanding of social attention by investigating the extent to which interactive social contexts shape basic visual attention.
Social attention
Since the first eye-tracking experiments, it has been shown that people like to look at other people, and in particular their eyes (Yarbus, Citation1965). Moreover, humans are skilled gaze followers (Emery, Citation2000), applying this skill to learn about their environment from the earliest ages (Farroni et al., Citation2000). Looking at another person and following their gaze can reveal a range of useful information about one’s environment. It can help us to connect with others and establish a common ground for joint actions. This social attention has a key function in various aspects of social living, facilitating interpersonal communication, successful cooperation and human interdependence (Richardson & Gobel, Citation2015).
Cognitive scientists have studied the effects of looking at and with co-specifics using tightly controlled experiments in the laboratory. One such experiment is the gaze cueing paradigm, in which a face stimulus is presented in the centre of the computer screen. The face is first presented with closed eyes or with a direct gaze, followed by an averted gaze to imply eye movements. Finally, a target object appears at the looked-at (cued) or at a novel (uncued) location. Time and again findings have yielded faster detection and identification of cued compared to uncued targets (Driver et al., Citation1999; Friesen & Kingstone, Citation1998). Thus, the gaze cueing literature convincingly shows that the attentional focus of another person can act as a strong cue for a person’s own attention (Bayliss & Tipper, Citation2006; Friesen & Kingstone, Citation1998; Frischen et al., Citation2007; Ristic et al., Citation2002).
A different line of research has studied social attention in more interactive contexts using a conspecifics’ movements to detect targets. In these experiments, pairs of naïve participants sit opposite each other and take turns in reaching for a target stimulus. Results show that participants are slower to reach locations that were previously touched by the interaction partner, an inhibitory process called between-subject or social inhibition of return (Welsh et al., Citation2005, Citation2007). Social inhibition of return effects can arise from merely observing shifts in the attention of an interaction partner (Skarratt et al., Citation2010), and they are modulated by social information, such as who the interaction partner is (Nafcha et al., Citation2020). Thus, social inhibition of return is another experimental example of how humans are attuned to the attentional focus of another person.
But in spite of the interactive nature of this dynamic movement paradigm, the social nature of the social inhibition of return effect has been disputed, as it confounds the socialness of the cue with the directionality of the cue (Atkinson et al., Citation2014; Doneva et al., Citation2017). Researchers have speculated that this effect is better explained by shifts in attention to salient cues, which only happen to be social (Cole et al., Citation2019). Similarly, research shows that gaze cueing effects can persist in spite of the gazing agent being obstructed from viewing the target stimulus by a physical barrier (Cole et al., Citation2015), and attributing mental states onto the gazing face, or the acting agent behind the face is neither sufficient nor necessary for automatic shifts of social attention to occur in gaze cueing paradigms (Kingstone et al., Citation2019). Given these limitations of the gaze cueing paradigm and the interpersonal joint reaching paradigm, our own research has investigated social attention using an interactive version of the original spatial cueing paradigm (Posner & Cohen, Citation1984).
Social orienting effects in a joint spatial cueing task
In the classic spatial cueing paradigm, all stimuli are non-social in nature. Participants are exogenously cued to a spatial location. An exogenous intervening cue then disengages attention before participants are instructed to detect a target at the same spatial location or at a novel spatial location. Even though the cue does not predict the target location, it nevertheless influences behaviour (Posner & Cohen, Citation1984). While at shorter cue-target intervals (SOAs < 100 ms), participants are quicker to detect cued targets, the most relevant influence for the present purpose is that at longer cue-target intervals (SOAs > 300 ms) participants are slower to detect cued targets (Klein, Citation2000), a phenomenon known as inhibition of return (IOR) (Posner et al., Citation1985). This inhibition of return effect has been suggested to constitute a key mechanism for the exploration of novel spatial locations (Klein & MacInnes, Citation1999).
In our interactive version of the spatial cueing task, we maintained the exogenous nature of the cueing task since all stimuli were kept non-social in nature, with the only thing that we manipulated being the social meaning of one of the non-social stimuli (Tufft et al., Citation2015). In a novel manipulation, we made participants believe that the exogenous cue in the spatial cueing task represented the gaze location of an interaction partner. Specifically, participants’ attention was cued to a spatial location ostensibly gazed-at by their interaction partner. Their attention was then disengaged from that location by an intervening cue bringing it back to the centre of the screen. Participants were then found to be even slower to detect a target in cued versus the uncued spatial locations when that cue represented the gaze location of an interaction partner compared to when it did not. Thus, we showed that changing beliefs about the social meaning of a non-social cue increased inhibition of return effects (Gobel et al., Citation2018; Gobel & Giesbrecht, Citation2020; Tufft et al., Citation2015). One possible interpretation for why the non-social cue became socially meaningful in the joint spatial cueing task was that it might have been interpreted as a signal emitted by the interaction partner that they were “taking care” of the cued spatial location, reflecting a division of labour, and a notion that draws upon the ideas of distributed cognition (Hutchins, Citation1995).
The role of cooperation in modulating social orienting effects
Because people have a clear understanding that eye movements are meaningful, they can use gaze shifts to signal information to one another (Gobel et al., Citation2015, Citation2017; Jarick & Kingstone, Citation2015). Perceivers can pick this information up and interpret its meaning, for example, that a given spatial location is being "taken care of” in the joint spatial cueing task. If true, then the resulting inhibition of return might free attentional resources for the exploration of novel locations. Yet, to do so successfully, the interacting partners would need to buy into the idea that they are working together on the task and as such interpret the social context as being cooperative. Cooperation establishes common ground, minimizes collective efforts, and avoids potential costs that arise in joint activities (Brennan et al., Citation2008; Clark, Citation1996; Clark & Brennan, Citation1991). Thus, in the case of a joint spatial cueing task, we predicted that greater levels of perceived cooperativeness would result in larger inhibition of return effects, reflecting a division of labour and participants distributing their cognition with others who they interpret as “taking care of” cued (“looked at”) spatial locations.
Social psychological research has shown that participants change how they think about and behave towards an interaction partner depending on the social context (Fiske & Neuberg, Citation1990). In particular, the gender of a person is one of the characteristics that is immediately accessible and that readily activates gender stereotypes. These gender stereotypes describe the beliefs that females are more communal and cooperative, whereas males are more assertive and competitive (Eagly, Citation2009). Moreover, individuals who act outside traditional gender roles are penalized (Rudman, Citation1998). Thus, research has found that females do indeed cooperate more than males, in particular in mix-gender dyads (Balliet et al., Citation2011; Niederle & Vesterlund, Citation2011; Van Vugt et al., Citation2007). If levels of cooperativeness influence the inhibition of return effect, then we might expect to observe larger inhibition of return effects for female compared to male participants. Moreover, by experimentally manipulating the cooperative context of the joint spatial cueing task, we might expect to directly manipulate the magnitude of inhibition of return effects. In the present research, we tested both hypotheses.
The present research
In the present research, we were interested in whether interpreting social contexts as cooperative would influence social attention. We tested this idea using a newly developed online version of the joint spatial cueing task. In this online version, participants signed up to play a game with another person. They briefly “e-met” their playing partner via a video call and saw him wave into the camera. In reality, the interaction partner was always a video recording of a confederate. Next, participants completed a webcam-based eye-tracking calibration procedure, and they saw their playing partner do the same. E-meeting the playing partner and witnessing the eye-tracking calibration procedure were two key elements that we developed for this online version of the joint spatial cueing task. They were crucial in making participants believe that they were interacting with another player and that they could even see where that person was looking on the screen.
Participants then completed the joint spatial cueing task as developed by Tufft and colleagues (Citation2015). They first saw a set of images appear on screen, which they were free to inspect. They then saw the cue (a red dot) appear on top of one of these images, which they were told represented the image their playing partner had just looked at. Participants’ attention was then returned to the centre of screen by an intervening cue. Participants’ task was to respond as quickly as possible to the onset of the target (a blue square) by pressing the space bar (target detection task). At the end of the experiment, we tested participants’ levels of suspiciousness, and removed any participants who did not believe that their playing partner was real or clearly doubted the interactive nature of the game.
Across four experiments, we tested whether cooperativeness would modulate inhibition of return effects in a joint spatial cueing task. Experiment 1a and 1b tested whether more cooperative individuals would show larger inhibition of return effects. We predicted that if females are more cooperative than males, then they would also show larger inhibition of return effects. Experiment 2a and b tested whether highly cooperative interactive contexts would yield larger inhibition of return effects. We predicted that framing the social context as a team game rather than an individual game would result in greater inhibition of return effects.
Experiment 1a
Experiment 1a tested whether more cooperative individuals would show larger inhibition of return effects in a joint spatial cueing task. We predicted that female compared to male participants would show larger inhibition of return effects.
Methods
Participants
The sample size was estimated based on previous joint spatial cueing task studies showing moderate to large effect sizes. A power analysis suggested targeting a sample of at least 75 subjects in order to have 80% power for detection of a medium to large-sized effect when employing the traditional 0.05 criterion for statistical significance.
A total of 78 participants were recruited using Prolific (www.prolific.co) and volunteered to participate in exchange for a payment (average payment of £2.75 for 20 mins work). We aimed to recruit an equal number of males and females, and participants were pre-screened for English nationality, normal-to-corrected vision, a working webcam, and for comfort in taking part in a deception study. Three participants were excluded because they either expressed suspicion about the confederate being another player, or they encountered technical issues with the task. We therefore analysed data from 75 participants (38 females, Mage = 33.8, SDage = 11.1; 37 males, Mage = 32.4, SDage = 11.6).
All participants provided their informed consent and were fully debriefed on the confederate deception following the completion of the experiment. Ethical approval for all experiments was obtained prior to data collection.
Design
We employed a 2 × 2 mixed-factor design with one within-subject factor, target-location (cued vs uncued), and one between-subject factor, participant-gender (male vs female). The main dependent variable of interest was participants’ reaction times to the onset of the target stimulus.
Stimuli and apparatus
The experiment was hosted entirely online using the web-based platform Gorilla (www.gorilla.sc) and set up using the Gorilla Experiment Builder (Anwyl-Irvine et al., Citation2020). Procedures and stimuli were adapted for online data collection (Tufft, Citation2020) from the original joint spatial cueing task (Tufft et al., Citation2015).
To help improve data quality, we instructed participants to maximize their browsers, to hide browser toolbars, to close other programmes such as email clients or other browsers, and to sit comfortably at a table in an adequately lit room. We also restricted recruitment to laptop and desktop devices, Chrome browsers, and internet connections >10 mbps to ensure the smooth and consistent running of the experimental media.
Participants saw a 2 × 2 grid of four photos presented for 1200 ms. There were four sets of four photos used (16 in total) with each set having the same theme (beachscapes, landscapes, flowers, racing cars). All photos were taken at random from the Open Affective Standardized Image Set (OASIS) database (Kurdi et al., Citation2017) and had their saturation adjusted to 60% to ensure the shapes presented on them were clear and highly salient. Next, a red dot would appear in the exact centre of one of the four photos for 300 ms (visual cue). After the cue had disappeared, a yellow star (intervening cue) appeared in the centre of the quadrant for an average of 600 ms (jittered by ±200 ms to ensure target onsets not predictable) in order to exogenously disengage attention from the cue and return it back to the centre of the screen (consistent with studies known to elicit inhibition of return (e.g., Birmingham et al., Citation2007)). This was then followed by the presentation of the blue square (visual target) in the exact centre of one of the four photos. The target remained on screen for up to 2000 ms or until the participant responded with a spacebar press. If no response was given, the trial would end and be followed by the next trial. The trial design is shown in . The sequence and timings used here are in line with previous literature that have demonstrated the reliable generation of socially modulated inhibition of return effects (Gobel et al., Citation2018; Tufft et al., Citation2015).
Figure 1. Joint spatial cuing paradigm trial structure used for all experiments (4 location display shown). Participants were first presented with an array of photos (2 for Exp. 1b, 4 for Exp. 1a, 2a & 2b), following which a red dot cue was presented in the middle of one of the photos. Participants were told that this indicated the photo their playing partner just looked at and as such was imbued with social meaning. After an intervening cue that returned attention back to the centre of the display, participants were instructed to respond as quickly as possible to the appearance of a blue square, which could appear at a cued or uncued location.

In total, participants completed 216 trials, split across 3 blocks of 72 trials. Between each block, participants were reminded that their task was to respond as quickly and as accurately as possible to the blue square and that the red dot represented where their interaction partner had just looked. Of the 216 trials, 24 were catch trials where no blue square was presented to act as an attention check and to ensure response accuracy. When the blue square (visual target) was presented in the same location as the preceding red dot (visual cue), it was classed as a cued target trial; when presented in one of the three other locations, it was classed as an uncued target trial (factor: target-location). As such, 25% of all trials (48 trials) were cued target trials and 75% of all trials (144 trials) were uncued target trials. The difference in reaction times between the cued target and the uncued target trials represented the size of the inhibition of return effect. Red dot cue location and blue square target location were fully counterbalanced across participants to ensure that all cue-target permutations were equally sampled. Photo set type and photo location were pseudo-randomized to ensure an equal number of presentations of each photo at each location across participants.
Procedure
On entering the experiment, participants completed a series of guidance and instruction screens in order to set up the appropriate context for social interactivity and to instruct them on how to perform the task. We first told participants that they would play a simple online game with another randomly selected participant who they would briefly meet via webcam before starting the game. The partner was in fact a pre-recorded video of a male research assistant. The purpose of this was to establish the belief that they would be engaging in a dyadic interaction with another person. To convince participants of this, they completed an actual “online meeting” during which their webcams were turned on, so that they could see themselves on screen, and this was shown alongside a 5 s pre-recorded video of the confederate waving as if meeting the participant for the first time live via webcam. All participants were next assigned a “gaming avatar” (as was the confederate) which displayed their names underneath (confederate was always called “Chris”). These were presented in the top left and right corners of the screen throughout the game to help act as an ongoing cue that there were always two players playing the game live (see ).
Figure 2. Example screenshots showing the procedural set up for the online joint spatial cuing task. Participants first “met” another participant online (a pre-recorded video of a confederate) with their webcam turned on (top left; image depicts authors for illustration). Participants then logged into the game within a shared screen zone, enabling them to view their partner (pre-recorded) doing the same (top right). Next, participants completed a webcam calibration sequence while viewing their partner (pre-recorded) do the same (bottom left). Finally, participants entered the game and completed the joint spatial cuing task believing that the red dot represented their partner’s gaze location (bottom right).
Importantly, we also told participants that during the game we would be tracking their and their partner’s eye movements via their webcams. They were informed that at certain times the eye-trackers would be connected so that they would be able to see on their screens where their playing partner had just looked. Before starting the game and in order to establish this belief, participants completed a real online eye-tracker calibration sequence using actual web-based eye-tracking technology offered by Gorilla (Anwyl-Irvine et al., Citation2020). They then watched a pre-recorded video, which participants believed to be live, of the confederate completing the same sequence. This aimed to demonstrate that we could indeed track their eye movements and those of their partner as well as continuing to bolster the belief that they were connected live to another person.
Next, participants were provided detailed instructions on the game itself. We informed them that in the game, they would view a series of photos arranged in a quadrant, and it was highlighted that they would view the same photos at the same time as their partner on a shared screen. After viewing the quadrant of photos for a couple of seconds, participants were told that a red dot would appear centrally on one of the four photos. Crucially, for the generation of socially modulated inhibition of return effects, they were informed that this red dot used live eye-tracking data to signal which photo their playing partner had just looked at and that their partner would also see on their screen which photo they had just looked at. They were then instructed that their task (along with their playing partner) would be to press the spacebar as soon as they detect the appearance of a blue square that would appear shortly after the red dot and centrally on one of the four photos. Participants were given no further instructions or guidance on how to use the red dot and were told to press the spacebar as quickly and as accurately as possible as reaction times and accuracy would be measured. We did not collect any eye-tracking data at any point, and the position of the red dot and blue square was always computer generated and equiprobable across the four locations.
Upon completion of the game, participants filled out a post-game questionnaire in which they reported their demographics (age, gender, and ethnicity), and answered questions on their perceptions of their own level of cooperativeness (“How cooperative do you think you are?” – Likert sliding scale 1-Not cooperative at all to 7-Very cooperative) and that of their partner (“How cooperative do you think your playing partner is?” – Likert sliding scale 1-Not cooperative at all to 7-Very cooperative). Based on these two questions, a cooperation-index was calculated by summing the ratings of self-cooperativeness and partner-cooperativeness to provide a measure of perceived cooperation in the interaction from the participant’s perspective (M = 10.9, SD = 2.3). Participants also answered several filler questions on their perceptions of their own and their partner’s trustworthiness, performance, and social status (factors not analysed as part of this study).
At the end of the experiment, we checked whether participants were suspicious about any element of the procedure. We asked them to confirm that their webcam had been turned on as well as the webcam of their partner. If they answered “no” to either question, they were invited to offer a reason. Participants were, therefore, able to express any doubts about the confederate being another player without us overtly leading the question. Of the 78 participants recruited, only 1 expressed a clear doubt that the confederate was not real, and only 2 expressed issues about the validity of the eye-tracking procedure. All three were excluded from all analyses.
Results
Participant gender modulates the perceived cooperation of the social interaction
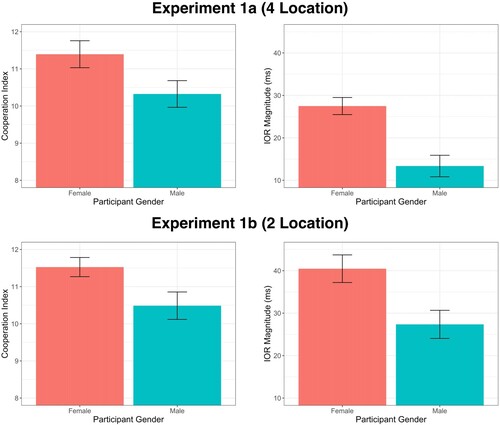
We conducted an independent-samples t test with one between-subjects factor of participant-gender (male vs female) on the cooperation-index measure. Consistent with our hypothesis, we observed a significant main effect of participant-gender, t(72.99) = 2.10, p = .04, d = 0.48, with female participants (N = 38, M = 11.4, SD = 2.2) reporting a higher cooperation-index than male participants (N = 37, M = 10.3, SD = 2.2) (degrees of freedom were corrected to account for unequal variance between groups). Thus, as can be seen in (A), female participants perceived their social interactions to be overall higher in cooperativeness compared to male participants.
Figure 3. (A) (Top Left). Perceived cooperation of the social interaction as a function of participants' gender in Experiment 1a. Error bars represent SEM. (B) (Top Right). Inhibition of return effects as a function of participants’ gender in Experiment 1a. Error bars represent SEM. (C) (Bottom Left). Perceived cooperation of the social interaction as a function of participants’ gender in Experiment 1b. Error bars represent SEM. (D) (Bottom Right). Inhibition of return effects as a function of participants’ gender in Experiment 1b. Error bars represent SEM.
Participant gender modulates the magnitude of inhibition of return effect
We analysed trial reaction time (RT) data for all trials in which cues and targets appeared on screen (no catch trials). We excluded trials on which participants may have anticipated their responses (RT < 175 ms), responded more than three standard deviations above the trial mean RT by trial type (target-location), or did not respond at all (RT > 2000 ms). Thus, we excluded 1.97% of the data.
We tested whether the magnitude of the inhibition of return effect was significantly larger for female participants compared to male participants. We conducted a 2 target-location (cued vs uncued; within-subjects) × 2 participant-gender (male vs female; between subjects) mixed-factorial analysis of variance (ANOVA) on the mean RTs for detecting targets. As predicted, we observed a significant main effect of target-location, F(1, 73) = 160.72, p < .001, , with cued targets (M = 469 ms, SEM = 11 ms) being responded to slower than uncued targets (M = 448 ms, SEM = 10 ms), reflecting an overall inhibition of return effect of 21 ms. The main effect of participant-gender was not significant (F(1, 73) < 1, p = .33,
).
Consistent with our prediction, we also observed a significant two-way interaction between target-location and participant-gender, F(1, 73) = 19.22, p < .001, . Male participants were significantly slower to respond to cued targets (M = 455 ms, SEM = 16 ms) compared to uncued targets (M = 441 ms, SEM = 16 ms) reflecting an inhibition of return effect of 13 ms. This difference was larger for female participants (Mcued = 482 ms, SEMcued = 14 ms; Muncued = 455 ms, SEMuncued = 13 ms), who showed an inhibition of return effect of 27 ms. Put another way, inhibition of return magnitude for female participants (M = 27 ms, SEM = 2 ms) was significantly larger than for male participants (M = 13 ms, SEM = 3 ms), t(68.92) = 4.37, p < .001, d = 1.01. These results are depicted in (B).
Discussion
Consistent with the literature on gender stereotypes, Experiment 1a found that females perceived the interactive context as more cooperative than males, and in line with our prediction, females yielded larger inhibition of return effects than males. Of note, the overall inhibition of return effect in Experiment 1a was 21 ms which is comparable to what Tufft and colleagues (Citation2015) observed in the social condition of the original joint spatial cueing task experiments. Thus, Experiment 1a also provided concurrent validity for the online version of the joint spatial cueing task.
An important limitation of Experiment 1a was the presentation of 25% cued target trials compared to 75% uncued target trials. Despite the location of the target being equiprobable across all four locations, participants may have learned that on any given trial targets were more likely to occur at uncued compared to cued spatial locations (e.g., Friesen et al., Citation2004). Therefore, we cannot rule out that the slowing of responses to cued compared to uncued targets in Experiment 1a may have been a result of target predictability rather than an inhibition of return effect. To address this limitation, in Experiment 1b, we used two cue-target locations, allowing for an equal number of cued and uncued trials.
Experiment 1b
Experiment 1b aimed to replicate the findings of Experiment 1a using a two-location set up to ensure equal numbers of cued and uncued trials. As in Experiment 1a, we predicted that more cooperative individuals (females) would show larger inhibition of return effects.
Methods
Participants
A total of 77 participants were recruited using Prolific (www.prolific.co) and volunteered to participate in exchange for a payment. We used the same recruitment strategy and inclusion criteria as in Experiment 1a. Two participants were excluded because they either expressed clear doubts about the confederate being another player, or they encountered technical issues with the task. We therefore analysed data from 75 participants (38 females, Mage = 32.2, SDage = 12.5; 37 males, Mage = 36.3, SDage = 12.9). All participants provided their informed consent and were fully debriefed on the confederate deception following completion of the experiment.
Design, stimuli, apparatus and procedure
All aspects of design, stimuli, apparatus, and procedure were identical to Experiment 1a except that participants carried out a spatial cueing paradigm with two horizontal cue-target locations. Thus, participants viewed two (rather than four) photos displayed side by side along the horizontal axis with a ratio of picture width to full display width of 1:2.5.
Trial sequences were also identical with participants completing 216 trials, split across 3 blocks of 72 trials. Crucially though, 50 percent of all non-catch trials were now cued target trials (96 trials) and 50 percent were uncued target trials (96 trials), with the remaining 24 trials as catch trials. Location of the target was therefore equiprobable across cued and uncued locations. Photo set type, photo location, red dot location, and blue square location were all fully counterbalanced across participants to ensure that all permutations were equally sampled.
In line with Experiment 1a, a cooperation-index was calculated by summing the ratings of self-cooperativeness and partner-cooperativeness to provide a measure of perceived cooperation in the interaction from the participant’s perspective (M = 11.0, SD = 2.0).
Results
Participant gender modulates the perceived cooperation of the social interaction
We conducted an independent-samples t test with one between-subjects factor of participant-gender (male vs female) on the cooperation-index measure. Consistent with our hypothesis, we observed a significant main effect of participant-gender, t(64.74) = 2.31, p = .02, d = 0.54, with female participants (N = 38, M = 11.5, SD = 1.6) reporting a higher cooperation-index than for male participants (N = 37, M = 10.5, SD = 2.2). Therefore, female participants perceived their social interactions to be overall higher in cooperativeness compared to male participants ((C)).
Participant gender modulates the magnitude of inhibition of return effect
We pre-processed trial reaction time (RT) data in the same way as Experiment 1a, excluding 1.79% of the data, and then tested whether the magnitude of the inhibition of return effect was significantly larger for female participants compared to male participants.
We conducted a 2 target-location (cued vs uncued; within-subjects) × 2 participant-gender (male vs female; between subjects) mixed-factorial ANOVA on the mean RTs for detecting targets. As predicted, we observed a significant main effect of target-location, F(1, 73) = 213.48, p < .001, , with cued targets (M = 511 ms, SEM = 9 ms) being responded to slower than uncued targets (M = 477 ms, SEM = 9 ms), reflecting an overall inhibition of return effect of 34 ms. The main effect of participant-gender was not significant (F(1, 73) = 1.27, p = .26,
).
Consistent with our prediction, we also observed a significant two-way interaction between target-location and participant-gender, F(1, 73) = 7.98, p = .006, . Male participants were significantly slower to respond to cued targets (M = 498 ms, SEM = 12 ms) compared to uncued targets (M = 470 ms, SEM = 13 ms) reflecting an inhibition of return effect of 27ms. Importantly, this difference was larger for female participants (Mcued = 524 ms, SEMcued = 12 ms; Muncued = 483 ms, SEMuncued = 12 ms), who showed an inhibition of return effect of 41 ms. Inhibition of return magnitude for female participants (M = 41 ms, SEM = 3 ms) was significantly larger than for male participants (M = 27 ms, SEM = 3 ms), t(72.93) = 2.82, p = .006, d = 0.65. These results are depicted in (D).
Discussion
Experiment 1b replicated findings of Experiment 1a with female participants showing larger inhibition of return effects compared to male participants. Moreover, female participants continued to report the experimental context as being more cooperative than male participants.
As Experiment 1b employed an equal ratio of cued (50%) compared to uncued trials (50%), these findings rule out the possibility that target predictability may have accounted for findings in Experiment 1a. They are consistent with other recent findings showing social modulation of inhibition of return effects in a joint spatial cueing task when using equiprobable cue-target trials (Gobel & Giesbrecht, Citation2020).
Interestingly, the overall size of the IOR effect in Experiment 1a (21 ms) was smaller than in Experiment 1b (34 ms). This intriguing result is consistent with literature that has shown a decrease in inhibition of return magnitude with increasing cue-target locations (Birmingham et al., Citation2007; Pratt et al., Citation1998). This gradient in inhibition of return magnitude is not believed to be a function of changes in target predictability or limits on processing capacity, but rather a dilution in the intensity of inhibitory tagging that comes from spreading responses across the visual field (Birmingham et al., Citation2007).
Experiment 2a
Experiments 1a and 1b showed that more cooperative individuals (females) yielded larger inhibition of return effects in the joint spatial cueing task as well as perceiving the interactive context as more cooperative. Experiment 2a aimed to directly manipulate the cooperativeness of the social context in order to test whether highly cooperative social contexts would elicit larger inhibition of return effects in a joint spatial cueing task. To test this hypothesis, for half of all participants, we introduced the joint spatial cueing task as a social but individual game (playing alongside an online partner) and for the other half of all participants as a team game (playing together with an online partner). We based this manipulation on the social psychological literature linking identification with groups to greater cooperative behaviour.
Groups do better and survive for longer when their members cooperate with one another (Tyler & Blader, Citation2001). Individuals, in turn, are motivated to become and stay a member of a group, as groups play a crucial role for their social identity or collective self (Sedikides & Brewer, Citation2001). Specifically, social identity theory postulates two important functions that groups play for individuals: groups help to shape individuals’ definition of who they are, and groups help to evaluate individuals’ self-worth (Tajfel & Turner, Citation2001). Thus, the more individuals identify with their group, the more they cooperate within it (Tyler & Blader, Citation2001).
We aimed to activate participants’ identification with their group, in this case, the dyadic game they would play with the interaction partner, by highlighting that they would be part of a team. We reasoned that to the extent that participants would identify as part of a team, they would be more motivated to work together to achieve a joint goal, that is to win the game. In other words, we predicted that playing the game as a team would represent a high-cooperative social context. In contrast, we predicted that playing the game individually would represent a low-cooperative social context. It should be noted that the social presence of the other game player remained consistent across both conditions, with the only difference being the participant’s belief about the individual-ness versus the team-ness of the game. In line with our view that construing a joint spatial cuing task as a cooperative game would invite opportunities for a division of labour, where participants interpret their partner as “taking care of” the cued (“looked at”) location, we predicted inhibition of return effects to be larger in the team compared to the individual game condition.
Methods
Participants
As per Experiment 1, the sample size was estimated based on previous studies using the joint spatial cueing task showing moderate to large effect sizes. A power analysis suggested targeting a sample of at least 136 subjects in order to have 80% power for detection of a medium-sized effect when employing the traditional 0.05 criterion for statistical significance. In Experiment 2, we were more conservative in our sample size estimation since evidence from Experiment 1 suggested slightly more variance in online data compared to previous lab-based data that may contribute to smaller than expected effect sizes.
A total of 146 participants were recruited using Prolific (www.prolific.co). We used the same recruitment strategy and inclusion criteria as before. Three participants were excluded because they either expressed clear doubt about the confederate being another player or they encountered technical issues with the task. We, therefore, analysed data from 143 participants (73 females of which 36 were in the individual condition and 37 were in the team condition, Mage = 33.3, SDage = 11.6; 70 males of which 36 were in the Individual Condition and 34 were in the Team Condition, Mage = 33.2, SDage = 11.7). All participants provided their informed consent and were fully debriefed on the confederate deception following completion of the experiment.
Design
We employed a 2 target-location (cued target vs uncued target) × 2 participant-gender (male vs female) × 2 game-context (team vs individual) mixed-factor design with the first factor being within-subjects, and latter two factors being between-subjects. The main dependent variable of interest was participants’ reaction times to the onset of the target stimulus.
Stimuli, apparatus & procedure
Experiment 2 was also hosted entirely online using the web-based platform Gorilla (www.gorilla.sc), set up using the Gorilla Experiment Builder (Anwyl-Irvine et al., Citation2020), and with the same restrictions on device type, browser type, and internet speed. Participants also completed the same guidance, instruction, and trial procedures as in Experiment 1 (using 4 cue-target locations) except for one key difference – whether they were told that the game was played as a team with their partner or as an individual alongside their partner (factor: game-context).
In the team game condition, participants were told that they would be working “with another participant as a team” and that the aim was to “maximise the team score”, which would be calculated from the average of their combined (participant and confederate) speed and accuracy in detecting the blue square. Furthermore, the confederate “Chris” was always referred to as the participant’s “teammate” rather than their “playing partner”. All other information about the game remained the same as in Experiment 1, and the red dot continued to represent where their playing partner looked.
In contrast, in the individual game condition, participants were told that they would be playing “alongside another participant as an individual” and that the aim was to maximise their “individual score”, which would be calculated based on their own speed and accuracy in detecting the blue square. They were also told that their score would be compared against all other players that would complete the game (i.e., not exclusively compared against the confederate). And rather than their “playing partner,” “Chris” was always referred to as their “assigned competitor”. As in the Team game condition, all other information about the game remained the same as in Experiment 1.
In reality, we did not compute any scores in either of the game-context conditions. All social, contextual and perceptual information remained consistent across all conditions with the only difference being whether the participant believed they were playing with someone else as part of a team game or alongside someone else as an individual game.
On completion of the game, participants filled out the same post-game questionnaire as Experiment 1. Based on the two questions on cooperativeness, a cooperation-index was calculated by summing the ratings of self-cooperativeness and partner-cooperativeness to provide a measure of perceived cooperation in the interaction from the participant’s perspective (M = 11.1, SD = 1.8). Participants responded to two further questions which aimed to check if the game-context manipulation was successful in shifting participants’ perceptions about the game itself (“In the game, how much did you feel like a team playing with your playing partner?” / “In the game, how much did you feel like an individual playing alongside your playing partner?” – Likert sliding scale 1-Not at all to 7-Very).
Results
Manipulation check: perceived team-ness vs perceived individual-ness of the game
To test whether our manipulation was successful in generating a sense of team-ness or individual-ness during the game, we conducted a 2 game-context (team vs individual) × 2 participant-gender (male vs female) ANOVA. We did this independently for the team and the individual manipulation check items.
For perceived team-ness, we observed a significant main effect of game-context, F(1, 139) = 16.92, p < .001, , such that mean ratings of experiencing the game as a team were significantly higher in the team game condition (N = 71, M = 4.1, SEM = 0.2) than in the individual game condition (N = 72, M = 3.0, SEM = 0.2), consistent with the team game condition generating a greater sense of team-ness. Notably, the main effect of participant-gender and the interaction of participant-gender and game-context did not reach levels of significance (all Fs < 1, all ps > .35).
For perceived individual-ness, this result was partially mirrored with a significant main effect of game-context, F(1, 139) = 33.49, p < .001, . Participants reported to experience the individual game condition (N = 72, M = 5.2, SEM = 0.2) more as an individual than the team game condition (N = 71, M = 3.6, SEM = 0.2), consistent with the individual game condition generating a sense of individual-ness. Interestingly, there was also a main effect of participant-gender, F(1, 139) = 8.42, p = .004,
, such that male participants (N = 70, M = 4.8, SEM = 0.2) reported to experience the game context more as an individual compared to female participants (N = 73, M = 4.0, SEM = 0.2). However, there was no evidence that this differed by the type of game played as the interaction of participant-gender and game-context was not significant, F(1, 139) = 0.04, p = .84,
.
In sum, our game-context manipulation was successful in inducing a sense of team-ness in the team game condition and a sense of individual-ness in the individual game condition for both male and female participants.
Game context modulates the perceived cooperativeness of the social interaction
To test whether participants experienced greater cooperativeness in the team compared to the individual game conditions, we submitted the cooperation-index to a 2 game-context (team vs individual) × 2 participant-gender (male vs female) ANOVA. We observed a significant main effect of game-context, F(1, 139) = 4.75, p = .03, , such that mean cooperation-index scores were higher in the team game condition (M = 11.4, SEM = 0.2) compared to the individual game condition (M = 10.8, SEM = 0.2), consistent with the manipulation raising perceptions of cooperativeness in the team condition.
Interestingly and as can be seen in (A), this was further qualified by a significant two-way interaction between game-context and participant-gender, F(1, 139) = 4.09, p = .045, . Post hoc model contrasts of estimated marginal means with Bonferroni adjusted α levels revealed that male participants rated the cooperativeness of the dyads significantly lower in the individual game condition (M = 10.2, SEM = 0.3) compared to the team game condition (M = 11.5, SEM = 0.2), t(139) = 2.94, p = .008, d = 0.70. However, there was no evidence for any difference in perceived cooperativeness for female participants between the team and individual game conditions (Mteam = 11.3, SEMteam = 0.3; Mindividual = 11.3, SEMindividual = 0.3; t(139) = .11, p = 1, d = 0.03). The main effect of participant-gender did not reach significance, F(1, 139) = 2.34, p = .13,
.
Figure 4. (A) (Top Left). Perceived cooperation of the social interaction as a function of game context and participants’ gender in Experiment 2a. Error bars represent SEM. (B) (Top Right). Inhibition of return effects as a function of game context and participants’ gender in Experiment 2a. Error bars represent SEM. (C) (Bottom Left). Perceived cooperation of the social interaction as a function of game context and participants’ gender in Experiment 2b. Error bars represent SEM. Figure (D) (Bottom Right). Inhibition of return effects as a function of game context and participants’ gender in Experiment 2b. Error bars represent SEM.
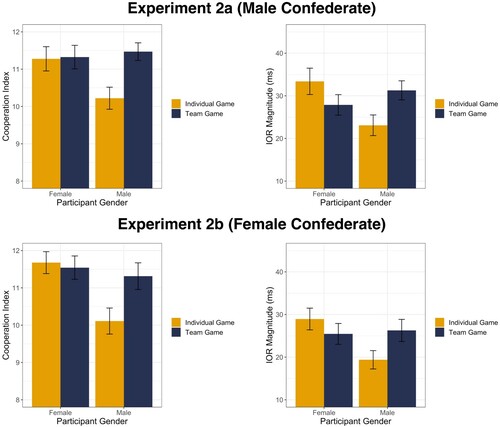
In sum, game-context modulated the perceived cooperativeness of the dyadic pairs, but for male participants only. Male participants reported higher perceived cooperativeness of the dyad in the Team game condition compared to the individual game condition. In contrast, there was no such difference for female participants, who rated both game conditions consistently high in perceived cooperativeness.
Game context modulates the magnitude of the inhibition of return effect
We pre-processed trial reaction time (RT) data in the same way as Experiment 1, excluding 1.82% of the data, and then tested whether inhibition of return effects would be greater in the team game condition compared to the individual game condition.
We conducted a 2 target-location (cued vs uncued; within-subject) × 2 game-context (team vs individual; between-subject) × 2 participant-gender (male vs female; between-subject) mixed-factorial ANOVA for mean reaction times to detecting the target. We observed a significant main effect of target-location, F(1, 139) = 506.97, p < .001, , with mean reaction times for detecting cued targets (M = 444 ms, SEM = 6 ms) slower than uncued targets (M = 415 ms, SEM = 6 ms) reflecting an overall inhibition of return effect of 29ms. The main effect of game-context was also significant, F(1, 139) = 4.36, p = .04,
, with mean reaction times in the team game condition (M = 441 ms, SEM = 9 ms) slower than in the individual game condition (M = 418 ms, SEM = 7 ms). Notably, the two-way interaction between target-location and game-context was not significant, F(1, 139) = 0.27, p = .60,
. However, the three-way interaction between target-location, game-context and participant-gender was significant, F(1, 139) = 7.13, p = .008,
. To further unpack, we analysed the target-location by game-context interaction for male and female participants separately.
For male participants, the target-location by game-context interaction was significant, F(1, 68) = 6.14, p = .02, . Male participants were slower to detect cued targets (M = 457 ms, SEM = 12 ms) than uncued targets (M = 426 ms, SEM = 12 ms) in the team game condition, reflecting an inhibition of return effect of 31 ms. Crucially, a post hoc comparison of mean inhibition of return by game-context shows this difference was significantly reduced to 23 ms in the individual game condition (Mcued = 421 ms, SEMcued = 10 ms; Muncued = 398 ms, SEMuncued = 9 ms), t(67.82) = 2.49, p = .02, d = 0.59.
In contrast, for female participants, the target-location by game-context interaction was not significant, F(1, 71) = 2.00, p = .16, . Female participants were slower to detect cued targets (M = 455 ms, SEM = 13 ms) than uncued targets (M = 427 ms, SEM = 12 ms) in the team game condition, reflecting an inhibition of return effect of 28 ms. This difference was not significantly larger for the individual game condition (Mcued = 442 ms, SEMcued = 12 ms; Muncued = 409 ms, SEMuncued = 11 ms; IOR = 33 ms), t(66.38) = 1.41, p = .16, d = 0.33. No other main effects or interactions reached significance (target-location x participant-gender, F(1, 139) = 1.80, p = .18,
, all other Fs < 1, all ps > .45).
In sum, game-context modulated the magnitude of inhibition of return for male participants only. Male participants showed a larger inhibition of return effect in the team game condition compared to the individual game condition. In contrast, there was no such difference in inhibition of return effect for female participants. These results are shown in (B).
Discussion
Experiment 2a yielded a series of important findings. Unexpectedly, participants in the team game condition compared to the individual game condition did not differ in the magnitude of inhibition of return overall. Instead, analyses showed that this was only the case for male participants, who showed a significantly larger inhibition of return effect in the team compared to the individual game condition. Why was this not the case for female participants? One possibility is that females were more cooperative to begin with. Indeed, the existing literature demonstrates that when being exposed to social dilemmas framed as an individual task, females exhibit much less self-serving behaviours than males (Van Vugt et al., Citation2007).
Another important finding from Experiment 2a was that participants’ psychological experience of the team compared to the individual game condition differed significantly, with participants reporting the former as the more collaborative social context. Interestingly, here again, we observed important gender differences. Male participants reported that they experienced the team game condition as the more cooperative social context compared to the individual game condition. This was not the case for female participants, who experienced the team and the individual game conditions as equally high-cooperative social contexts. These psychological interpretations of the social context help to further understand the unexpected gender difference in the magnitude of the inhibition of return effect.
One limitation of Experiment 2a, and indeed Experiments 1a and 1b, was that the e-meeting of the alleged interacting player was a closely choreographed video recording of a male research assistant. We chose to only use one male player identity, so that every participant would be made to believe they interacted with the exact same person represented by the exact identical stimulus in terms of physical attractiveness, facial expression, dress, and background. As we were interested in the effect of the participants and their interpretation of the social context, we thought it would be prudent to control for the interacting player’s identity, as social psychological research shows how people change their self-perceptions and interpretations of the social situation contingent on the person they interact with (Balliet et al., Citation2011; Fiske et al., Citation2002; Sinclair et al., Citation2005). However, a remaining question is whether the reported effects are generalizable to a female interaction partner. We conducted Experiment 2b to answer this question.
Experiment 2b
Experiment 2b tested whether the results from Experiment 2a would replicate when the interaction partner was female. We predicted that highly cooperative social contexts would elicit larger inhibition of return effects in a joint spatial cueing task.
Methods
Participants
A total of 153 participants were recruited using Prolific (www.prolific.co). We used the same recruitment strategy and inclusion criteria as before. Thirteen participants were excluded because they either expressed clear doubt about the confederate being another player (12), or they encountered technical issues with the task (1). The larger number of exclusions reflected the change in confederate and relative differences in believability of the pre-recording. We therefore analysed data from 140 participants (69 females with 34 in the Individual Condition and 35 in the Team Condition, Mage = 31.3, SDage = 11.5; 71 males with 36 in the Individual Condition and 35 in the Team Condition, Mage = 37.6, SDage = 13.5). All participants provided their informed consent and were fully debriefed on the confederate deception following completion of the experiment.
Design, stimuli, apparatus and procedure
All aspects of design, stimuli, apparatus, and procedure were identical to Experiment 2a, except for the gender of the confederate. This time, the game-playing partner was a pre-recorded video of a female research assistant. As before, participants completed an “online meeting” during which their webcams were turned on, so that they could see themselves on screen, and this was shown alongside a 5 s pre-recorded video of the confederate meeting the participant for the first time live via webcam. The female confederate was instructed to wave in the same manner as the male confederate and continued to use the gender-neutral name “Chris”.
Again, a cooperation-index was calculated by summing the ratings of self-cooperativeness and partner-cooperativeness to provide a measure of perceived cooperation in the interaction from the participant’s perspective (M = 11.1, SD = 2.0).
Results
Manipulation check: perceived team-ness vs perceived individual-ness of the game
To test whether our manipulation was successful in generating a sense of team-ness or individual-ness during the game, we conducted a 2 game-context (team vs individual) × 2 participant-gender (male vs female) ANOVA. We did this independently for the team and the individual manipulation check items.
For perceived team-ness, we observed a significant main effect of game-context, F(1, 136) = 10.77, p = .001, , such that mean ratings of experiencing the game as a team were significantly higher in the team game condition (N = 70, M = 4.3, SEM = 0.2) than in the individual game condition (N = 70, M = 3.3, SEM = 0.2), consistent with the team game condition generating a greater sense of team-ness. The main effect of participant-gender (F(1, 136) = 2.53, p = .11,
) and the interaction of participant-gender and game-context did not reach levels of significance (F(1, 136) = 0.33, p = .56,
).
For perceived individual-ness, there was a significant main effect of game-context, F(1, 136) = 47.46, p < .001, . Participants reported to experience the individual game condition (N = 70, M = 5.2, SEM = 0.2) more as an individual than the team game condition (N = 70, M = 3.3, SEM = 0.2), consistent with the individual game condition generating a sense of individual-ness. There was no main effect of participant-gender, F(1, 136) = 0.03, p = .86,
, however, the interaction of participant-gender and game-context approached significance, F(1, 136) = 3.76, p = .06,
(Males: Mteam = 3.1, SDteam = 0.3, Mindividual = 5.5, SDindividual = 0.2; Females: Mteam = 3.6, SDteam = 0.3, Mindividual = 4.9, SDindividual = 0.3).
In sum, our game-context manipulation was successful in inducing a sense of team-ness in the team game condition and a sense of individual-ness in the individual game condition for both male and female participants.
Game context modulates the perceived cooperativeness of the social interaction
To test whether participants experienced greater cooperativeness in the team compared to the individual game conditions, we submitted the cooperation-index to a 2 game-context (team vs individual) × 2 participant-gender (male vs female) ANOVA. The main effect of game-context was not significant, F(1, 136) = 2.62, p = .11, , although directionally mean cooperation-index scores were higher in the team game condition (M = 11.4, SEM = 0.2) compared to the individual game condition (M = 10.9, SEM = 0.3). The main effect of participant-gender did reach significance, F(1, 136) = 7.37, p = .007,
, with overall perceptions of cooperativeness greater for females (M = 11.6, SD = 0.2) than males (M = 10.7, SD = 0.3).
Replicating Experiment 2a, there was a significant two-way interaction between game-context and participant-gender, F(1, 136) = 4.09, p = .045, . Post-hoc model contrasts of estimated marginal means with Bonferroni adjusted α levels revealed that male participants rated the cooperativeness of the dyads significantly lower in the individual game condition (M = 10.1, SEM = 0.3) compared to the team game condition (M = 11.3, SEM = 0.4), t(136) = 2.59, p = .02, d = 0.62. However, there was no evidence for any difference in perceived cooperativeness for female participants between the team and individual game conditions (Mteam = 11.5, SEMteam = 0.3; Mindividual = 11.7, SEMindividual = 0.3; t(136) = .28, p = 1, d = 0.07).
In sum, Experiment 2b replicated Experiment 2a in that game-context modulated the perceived cooperativeness of the dyadic pairs, but for male participants only. Male participants reported higher perceived cooperativeness of the dyad in the Team game condition compared to the individual game condition with no such difference for female participants, who rated both game conditions as highly cooperative ((C)).
Game context modulates the magnitude of the inhibition of return effect
We pre-processed trial reaction time (RT) data in the same way as Experiment 1, excluding 1.92% of the data, and then tested whether inhibition of return effects would be greater in the team game condition compared to the individual game condition.
We conducted a 2 target-location (cued vs uncued; within-subject) × 2 game-context (team vs individual; between-subject) × 2 participant-gender (male vs female; between-subject) mixed-factorial ANOVA for mean reaction times to detecting the target. We observed a significant main effect of target-location, F(1, 136) = 423.18, p < .001, , with mean reaction times for detecting cued targets (M = 478 ms, SEM = 6 ms) slower than uncued targets (M = 453 ms, SEM = 6 ms) reflecting an overall inhibition of return effect of 25ms.
Again, the two-way interaction between target-location and game-context was not significant, F(1, 136) = 0.49, p = .48, . However, in line with Experiment 2a, the three-way interaction between target-location, game-context and participant-gender was significant, F(1, 136) = 4.55, p = .04,
. To further unpack, we analysed the target-location by game-context interaction for male and female participants separately.
For male participants, the target-location by game-context interaction was significant, F(1, 69) = 4.21, p = .04, . Male participants were slower to detect cued targets (M = 482 ms, SEM = 13 ms) than uncued targets (M = 456 ms, SEM = 13 ms) in the team game condition, reflecting an inhibition of return effect of 26ms. Crucially, a post-hoc comparison of mean IOR by game-context shows this difference was significantly reduced to 19 ms in the individual game condition (Mcued = 450 ms, SEMcued = 9 ms; Muncued = 431 ms, SEMuncued = 9 ms), t(66.43) = 2.05, p = .045, d = 0.49.
In contrast, for female participants, the target-location by game-context interaction was not significant, F(1, 67) = 0.98, p = .33, . Female participants were slower to detect cued targets (M = 486 ms, SEM = 13 ms) than uncued targets (M = 461 ms, SEM = 13 ms) in the team game condition, reflecting an inhibition of return effect of 25ms. This was not significantly different from the individual game condition (Mcued = 495 ms, SEMcued = 14 ms; Muncued = 466 ms, SEMuncued = 13 ms; IOR = 29ms), t(66.80) = 0.99, p = .33, d = 0.24. No other main effects or interactions reached significance.
In sum, game-context modulated the magnitude of inhibition of return for male participants only, in line with findings from Experiment 2a and extending them to a female confederate ((D)).
Discussion
Experiment 2b showed that male participants yielded a significantly larger inhibition of return effect in the team compared to the individual game condition, but this was not the case for female participants. These findings not only replicate results from Experiment 2a but also generalize them further, as a female research assistant was used as game playing partner (i.e., confederate). This suggests that effects did not differ between same-gender and mix-gender dyads. Furthermore, in a separate analysis, we were able to rule out that different target angles across uncued target trials, resulting from a four-location set up, systematically influenced our findings in Experiment 2a and 2b (see supplementary materials).
General discussion
Summary of the results
Past research using a joint spatial cueing task has shown that social orienting effects are shaped by the partner with whom participants interact with (Gobel et al., Citation2018; Gobel & Giesbrecht, Citation2020; Tufft et al., Citation2015). The present research tested whether who the participants were and how they interpreted the social context would also modulate inhibition of return effects in a joint spatial cueing task. We found that being a more cooperative participant (Experiment 1a and 1b) and interacting within a high-cooperative social context (Experiment 2a and 2b) increased the magnitude of the inhibition of return effects (see for summary of mean RTs).
Experiment 1a and 1b found that female compared to male participants showed larger inhibition of return effects, and they interpreted the task as more cooperative. This finding is consistent with the literature on gender stereotypes and the behavioural evidence of females overall being more cooperative than men (Balliet et al., Citation2011; Eagly, Citation2009). Importantly, these results hold whether we used four spatial locations with 25% cued trials (Experiment 1a) or two spatial locations with 50% cued trials (Experiment 1b). Thus, the percentage of cued trials did not affect the social modulation of inhibition of return effects.
Experiment 2a and 2b found that introducing the joint spatial cueing task as a team game rather than an individual game, and directly manipulating the cooperativeness of the context, increased inhibition of return effects for male participants, who perceived team games as a more cooperative context. This finding is consistent with the social identity literature suggesting that highly identified group members cooperate more (Tyler & Blader, Citation2001). Experiment 2a and Experiment 2b further showed that these results were robust for interacting with a same gender or an opposite gender playing partner.
Thus, this series of experiments consistently demonstrate how perceptions of cooperativeness within the interactive context of a joint spatial cuing task play an important role in the modulation of inhibition of return effects. One possible interpretation of these findings is that when perceiving the interaction context to be a cooperative one, participants view their interaction partner as “taking care” of looked-at spatial locations, highlighting a division of labour, and enabling them to focus their attentional resources on the detection of targets in novel spatial locations. This view points to a socially attuned and distributed cognitive system that facilitates social living.
Theoretical implications
Our findings make a series of contributions towards a better understanding of interactive cognition. Firstly, social attention establishes common ground (Clark & Brennan, Citation1991), underlies joint action (Sebanz et al., Citation2006), and might facilitate the joint exploration of novel spatial locations (Gobel et al., Citation2018). But past social attention research has mostly focused on the role that interaction partners play for guiding visual attention. For example, research shows that who the interaction partner is prioritizes social orienting effects as early as 150 ms after the cue onset (Gobel & Giesbrecht, Citation2020). The present research adds to this literature, highlighting that who the participant is might be equally important in guiding visual attention in interpersonal contexts.
Secondly, our findings add to previous research suggesting that the creation of a positive and cooperative interaction context is essential for interactive cognition effects, such as a joint Simon effect, to occur (Hommel et al., Citation2009). We found that high-cooperative contexts increased inhibition of return effects. Other research, however, was unable to document any change in social inhibition of return effects when participants carried out a joint reaching task either as a cooperation or a competition (Atkinson et al., Citation2018), interacted with a similar or dissimilar interaction partner (Dalmaso et al., Citation2021), or competed versus cooperated in a joint perspective-taking task (Surtees et al., Citation2021). One possibility is that if the interaction partner is rendered highly salient in two experiments conditions, participants co-represented their partner’s task in both experimental conditions, resulting in a null finding. In Experiment 2a and 2b, we carefully removed the social relevance of the interaction partner in the individual game condition, while, crucially, maintaining both their social presence and the imbued social meaning of the cue. This was achieved by explaining to participants that they would play alongside a partner in trying to win the game against all other participating players. In male participants, we observed that this manipulation resulted in a significant lower perception of cooperativeness and a significantly lower inhibition of return effect in comparison to playing the game as a team.
Finally, the present findings suggest that construing the social context as cooperative might play an important role for socially distributed cognition to emerge (Hutchins, Citation1995; Tufft & Richardson, Citation2020). For example, when participants collaboratively search for visual targets, they use information about each other’s gaze location to improve their dyadic performance (Brennan et al., Citation2008). We propose that the present research adds to this literature in that when participants perceived social contexts to be highly cooperative, thelarger inhibition of return effects seen reflect a division of labour. We think that future research on interactive cognition needs to adopt a holistic perspective in order to better understand how it is dynamically shaped by who enters the situation (i.e., personality and dispositions), who they interact with (i.e., social characteristics of the interaction partner), and how they interpret the social context (i.e., social situation). Only if research addresses all three pillars of social behaviour will it be able to comprehensively assess cognition during interpersonal interactions.
Limitations and future studies
Our research is not without limitations. Conducting psychophysical research online is a novel frontier (Tufft, Citation2020). For example, as we conducted our research online, we had less experimental control over the size or the salience of our stimuli, as participants used various computers with different screen resolutions, and looked at them from different distances. This may have rendered our measure noisier than ideal. However, it should be noted that the overall inhibition of return effect in the present research (21 ms) was very similar to the magnitude of the inhibition of return effect that was originally observed in joint spatial cueing tasked tested in the controlled environment of the laboratory (Tufft et al., Citation2015). Crucially, if the online nature of our data collection increased the noise of our measure, it did so for both male and female participants (Experiment 1a & 1b), as well as in both the team and individual game conditions (Experiment 2a & 2b). Thus, we do not think that online data collection would have systematically influenced our results.
In the present study, we only used two-player identities as confederates. The e-meeting of the alleged interacting player was a closely choreographed video recording of a man (Experiment 1a, 1b, and 2a) or a woman (Experiment 2b). This was done so that every participant would be made to believe they interacted with the exact same person represented by the exact identical stimulus in terms of physical attractiveness, facial expression, dress, and background. As we were interested in the effect of the participants and their interpretation of the social context, we thought it would be prudent to control for the interacting player’s identity, as social psychological research shows how people change their self-perceptions and interpretations of the social situation contingent on the person they interact with (Balliet et al., Citation2011; Fiske et al., Citation2002; Sinclair et al., Citation2005). For example, the social attention literature suggests that group membership modulates attentional facilitation and social inhibition of return effects (Liuzza et al., Citation2011; Nafcha et al., Citation2020; Pavan et al., Citation2011). Thus, future research could test whether the here reported effects are generalizable to social contexts when the interaction partner holds a different group membership.
It is important to note that the precise nature and the specific mechanisms underlying the inhibition of return effect remain disputed (Dukewich & Klein, Citation2015; Klein, Citation2000; Lupiáñez et al., Citation2013). It is widely acknowledged that the inhibition of return effect in the original spatial cueing paradigm is exogenous and reflexive in nature (Klein, Citation2000; Posner et al., Citation1985; Tipper & Kingstone, Citation2005). The original spatial cueing paradigm uses, as does the here reported joint version of the spatial cueing paradigm, peripheral cues to exogenously engage attention and intervening cues to exogenously disengaging attention from them. However, this does not rule out the possibility that endogenous attention may interact with the exogenous nature of inhibition of return. Indeed, research has shown that goal-directed processes can modulate inhibition of return effects (Birmingham et al., Citation2007; Tipper & Kingstone, Citation2005). For example, Bucker and Theeuwes (Citation2014) showed that while the initial orienting effect is purely stimulus driven, attentional reorienting and inhibitory processes following the initial capture of attention are susceptible to modulation by motivation (Bucker & Theeuwes, Citation2014).
While it goes beyond the current research to pinpoint what stages of information processing and through what mechanism cooperative contexts modulate inhibition of return in the joint spatial cueing task, our findings add to the long list of studies demonstrating endogenous modulation of inhibition of return effects (Birmingham et al., Citation2007; Bucker & Theeuwes, Citation2014; Jones et al., Citation2002; Lupiáñez et al., Citation2001; Pratt et al., Citation1998; Tipper & Kingstone, Citation2005). Our findings are consistent with previous research showing that beliefs about the social meaning of the cue alone suffice to modulate inhibition of return effects (Gobel et al., Citation2018; Gobel & Giesbrecht, Citation2020; Liu et al., Citation2021; Tufft et al., Citation2015). Future research is needed to investigate the specific mechanism underlying such social modulation of inhibition of return effects, for example, whether it is more strategic and effortful or implicit and effortless in nature. The importance of the current findings lies in demonstrating the robust role that gender and perceived cooperation play in the social modulation of inhibition of return.
Table 1. Mean RTs in ms (with SEM) for Experiment 1 and 2.
Conclusion
Humans do not perceive social reality objectively, but rather they construe it subjectively. In the present research, we tested how the subjective interpretation of social contexts shapes visual attention. We found that when the social context of interacting with another person in a joint spatial cueing task was interpreted as cooperative, then inhibition of return effects increased. In Experiment 1a and 1b, these effects emerged for females, known to be cooperative in disposition, and in Experiment 2a and 2b, they emerged for males when they were led to interpret the social context as a team game. Our findings suggest that subjective interpretations of social contexts as highly cooperative play an important role in the joint orienting of visual attention, and speaks to behavioural coordination and distributed cognition. We suggest that future research on interactive cognition needs to address how social attention is dynamically guided by who enters the situation, who they interact with, and how they construe the interaction.
Data availability statement
The data and analysis code that support the findings of this study are openly available in the OSF at http://doi.org/10.17605/OSF.IO/72RMF.
Disclosure statement
No potential conflict of interest was reported by the author(s).
References
- Anwyl-Irvine, A. L., Massonnié, J., Flitton, A., Kirkham, N., & Evershed, J. K. (2020). Gorilla in our midst: An online behavioral experiment builder. Behavior Research Methods, 52(1), 388–407. https://doi.org/https://doi.org/10.3758/s13428-019-01237-x
- Atkinson, M. A., Millett, A. C., Doneva, S. P., Simpson, A., & Cole, G. G. (2018). How social is social inhibition of return? Attention, Perception, & Psychophysics, 80(8), 1892–1903. https://doi.org/https://doi.org/10.3758/s13414-018-1546-3
- Atkinson, M. A., Simpson, A., Skarratt, P. A., & Cole, G. G. (2014). Is social inhibition of return due to action co-representation? Acta Psychologica, 150, 85–93. https://doi.org/https://doi.org/10.1016/j.actpsy.2014.04.003
- Balliet, D., Li, N. P., Macfarlan, S. J., & Van Vugt, M. (2011). Sex differences in cooperation: A meta-analytic review of social dilemmas. Psychological Bulletin, 137(6), 881–909. https://doi.org/https://doi.org/10.1037/a0025354
- Bayliss, A. P., & Tipper, S. P. (2006). Predictive gaze cues and personality judgments: Should eye trust you? Psychological Science, 17(6), 514–520. https://doi.org/https://doi.org/10.1111/j.1467-9280.2006.01737.x
- Birmingham, E., Visser, T. A. W., Snyder, J. J., & Kingstone, A. (2007). Inhibition of return: Unraveling a paradox. Psychonomic Bulletin and Review, 14(5), 957–963. https://doi.org/https://doi.org/10.3758/BF03194128
- Brennan, S. E., Chen, X., Dickinson, C. A., Neider, M. B., & Zelinsky, G. J. (2008). Coordinating cognition: The costs and benefits of shared gaze during collaborative search. Cognition, 106(3), 1465–1477. https://doi.org/https://doi.org/10.1016/j.cognition.2007.05.012
- Bucker, B., & Theeuwes, J. (2014). The effect of reward on orienting and reorienting in exogenous cuing. Cognitive, Affective, & Behavioral Neuroscience, 14(2), 635–646. https://doi.org/https://doi.org/10.3758/s13415-014-0278-7
- Clark, H. H. (1996). Using language. Cambridge University Press.
- Clark, H. H., & Brennan, S. E. (1991). Grounding in communication. In Perspectives on socially shared cognition (pp. 127–149). American Psychological Association. https://doi.org/https://doi.org/10.1037/10096-006
- Cole, G. G., Smith, D. T., & Atkinson, M. A. (2015). Mental state attribution and the gaze cueing effect. Attention, Perception & Psychophysics, 77(4), 1105–1115. https://doi.org/https://doi.org/10.3758/s13414-014-0780-6
- Cole, G. G., Welsh, T. N., & Skarratt, P. A. (2019). The role of transients in action observation. Attention, Perception, and Psychophysics, 81(7), 2177–2191. https://doi.org/https://doi.org/10.3758/s13414-019-01740-5
- Dalmaso, M., Castelli, L., Scatturin, P., & Galfano, G. (2021). Can attitude similarity shape social inhibition of return? Visual Cognition, May. https://doi.org/https://doi.org/10.1080/13506285.2021.1922566
- Doneva, S. P., Atkinson, M. A., Skarratt, P. A., & Cole, G. G. (2017). Action or attention in social inhibition of return? Psychological Research, 81(1), 43–54. https://doi.org/https://doi.org/10.1007/s00426-015-0738-x
- Driver, J., Davis, G., Ricciardelli, P., Kidd, P., Maxwell, E., & Baron-Cohen, S. (1999). Gaze perception triggers reflexive visuospatial orienting. Visual Cognition, 6(5), 509–540. https://doi.org/https://doi.org/10.1080/135062899394920
- Dukewich, K. R., & Klein, R. M. (2015). Inhibition of return: A phenomenon in search of a definition and a theoretical framework. Attention, Perception, & Psychophysics, 77(5), 1647–1658. https://doi.org/https://doi.org/10.3758/s13414-015-0835-3
- Eagly, A. H. (2009). The his and hers of prosocial behavior: An examination of the social psychology of gender. American Psychologist, 64(8), 644–658. https://doi.org/https://doi.org/10.1037/0003-066X.64.8.644
- Emery, N. J. (2000). The eyes have it: The neuroethology, function and evolution of social gaze. Neuroscience and Biobehavioral Reviews, 24(6), 581–604. https://doi.org/https://doi.org/10.1016/S0149-7634(00)00025-7
- Farroni, T., Johnson, M. H., Brockbank, M., & Simion, F. (2000). Infants’ use of gaze direction to cue attention: The importance of perceived motion. Visual Cognition, 7(6), 705–718. https://doi.org/https://doi.org/10.1080/13506280050144399
- Fiske, S. T., Cuddy, A. J. C., Glick, P., & Xu, J. (2002). A model of (often mixed) stereotype content: Competence and warmth respectively follow from perceived status and competition. Journal of Personality and Social Psychology, 82(6), 878–902. https://doi.org/https://doi.org/10.1037/0022-3514.82.6.878
- Fiske, S. T., & Neuberg, S. L. (1990). A continuum of impression formation, from category-based to individuating processes: Influences of information and motivation on attention and interpretation. Advances in Experimental Social Psychology, 23, 1–74. https://doi.org/https://doi.org/10.1016/S0065-2601(08)60317-2
- Fiske, S. T., & Taylor, S. E. (2013). Social cognition: From brains to culture. Sage.
- Friesen, C. K., & Kingstone, A. (1998). The eyes have it! Reflexive orienting is triggered by nonpredictive gaze. Psychonomic Bulletin & Review, 5(3), 490–495. https://doi.org/https://doi.org/10.3758/BF03208827
- Friesen, C. K., Ristic, J., & Kingstone, A. (2004). Attentional effects of counterpredictive gaze and arrow cues. Journal of Experimental Psychology: Human Perception and Performance, 30(2), 319–329. https://doi.org/https://doi.org/10.1037/0096-1523.30.2.319
- Frischen, A., Bayliss, A. P., & Tipper, S. P. (2007). Gaze cueing of attention: Visual attention, social cognition, and individual differences. Psychological Bulletin, 133(4), 694–724. https://doi.org/https://doi.org/10.1037/0033-2909.133.4.694
- Gobel, M. S., Chen, A., & Richardson, D. C. (2017). How different cultures look at faces depends on the interpersonal context. Canadian Journal of Experimental Psychology/Revue Canadienne de Psychologie Expérimentale, 71(3), 258–264. https://doi.org/https://doi.org/10.1037/cep0000119
- Gobel, M. S., & Giesbrecht, B. (2020). Social information rapidly prioritizes overt but not covert attention in a joint spatial cueing task. Acta Psychologica, 211, 103188. https://doi.org/https://doi.org/10.1016/j.actpsy.2020.103188
- Gobel, M. S., Kim, H. S., & Richardson, D. C. (2015). The dual function of social gaze. Cognition, 136, 359–364. https://doi.org/https://doi.org/10.1016/j.cognition.2014.11.040
- Gobel, M. S., Tufft, M. R., & Richardson, D. C. (2018). Social beliefs and visual attention: How the social relevance of a cue influences spatial orienting. Cognitive Science, 42, 161–185. https://doi.org/https://doi.org/10.1111/cogs.12529
- Hommel, B., Colzato, L. S., & Van Den Wildenberg, W. P. M. (2009). How social are task representations? Psychological Science, 20(7), 794–798. https://doi.org/https://doi.org/10.1111/j.1467-9280.2009.02367.x
- Hutchins, E. (1995). Cognition in the wild. MIT Press.
- Jarick, M., & Kingstone, A. (2015). The duality of gaze: Eyes extract and signal social information during sustained cooperative and competitive dyadic gaze. Frontiers in Psychology, 6(September), 1423. https://doi.org/https://doi.org/10.3389/fpsyg.2015.01423
- Jones, M. R., Moynihan, H., MacKenzie, N., & Puente, J. (2002). Temporal aspects of stimulus-driven attending in dynamic arrays. Psychological Science, 13(4), 313–319. https://doi.org/https://doi.org/10.1111/1467-9280.00458
- Kingstone, A., Kachkovski, G., Vasilyev, D., Kuk, M., & Welsh, T. N. (2019). Mental attribution is not sufficient or necessary to trigger attentional orienting to gaze. Cognition, 189(August 2018), 35–40. https://doi.org/https://doi.org/10.1016/j.cognition.2019.03.010
- Klein, R. M. (2000). Inhibition of return. Trends in Cognitive Sciences, 4(4), 138–147. https://doi.org/https://doi.org/10.1016/S1364-6613(00)01452-2
- Klein, R. M., & MacInnes, W. J. (1999). Inhibition of return is a foraging facilitator in visual search. Psychological Science, 10(4), 346–352. https://doi.org/https://doi.org/10.1111/1467-9280.00166
- Kurdi, B., Lozano, S., & Banaji, M. R. (2017). Introducing the Open Affective Standardized Image Set (OASIS). Behavior Research Methods, 49(2), 457–470. https://doi.org/https://doi.org/10.3758/s13428-016-0715-3
- Liu, X., Qian, Q., Wang, L., Wang, A., & Zhang, M. (2021). Spatial inhibition of return affected by self-prioritization effect in three-dimensional space. Perception, 50(3), 231–248. https://doi.org/https://doi.org/10.1177/0301006621992940
- Liuzza, M. T., Cazzato, V., Vecchione, M., Crostella, F., Caprara, G. V., Aglioti, S. M. & Lauwereyns J. (2011). Follow my eyes: The gaze of politicians reflexively captures the gaze of ingroup voters. PloS One, 6(9), e25117. https://doi.org/https://doi.org/10.1371/journal.pone.0025117
- Lupiáñez, J., Martín-arévalo, E., & Chica, A. B. (2013). Is Inhibition of Return due to attentional disengagement or to a detection cost ? The Detection Cost Theory of IOR. September 1997, 221–252.
- Lupiáñez, J., Milliken, B., Solano, C., Weaver, B., & Tipper, S. P. (2001). On the strategic modulation of the time course of facilitation and inhibition of return. The Quarterly Journal of Experimental Psychology Section A, 54(3), 753–773. https://doi.org/https://doi.org/10.1080/713755990
- Nafcha, O., Morshed-Sakran, A., Shamay-Tsoory, S., & Gabay, S. (2020). The effect of co-actor group membership on the social inhibition of return effect. Acta Psychologica, 208(February), 103119. https://doi.org/https://doi.org/10.1016/j.actpsy.2020.103119
- Niederle, M., & Vesterlund, L. (2011). Gender and competition. Annual Review of Economics, 3(1), 601–630. https://doi.org/https://doi.org/10.1146/annurev-economics-111809-125122
- Pavan, G., Dalmaso, M., Galfano, G., Castelli, L.& Sirigu A. (2011). Racial group membership is associated to gaze-mediated orienting in Italy. PloS One, 6(10), e25608. https://doi.org/https://doi.org/10.1371/journal.pone.0025608
- Posner, M. I., & Cohen, Y. (1984). Components of visual orienting. Attention and Performance X: Control of Language Processes, 3(4), 531–556. https://doi.org/https://doi.org/10.1162/jocn.1991.3.4.335
- Posner, M. I., Rafal, R. D., Choate, L. S., & Vaughan, J. (1985). Inhibition of return: Neural basis and function. Cognitive Neuropsychology, 2(3), 211–228. https://doi.org/https://doi.org/10.1080/02643298508252866
- Pratt, J., Adam, J. J., & McAuliffe, J. (1998). The spatial relationship between cues and targets mediates inhibition of return. Canadian Journal of Experimental Psychology/Revue Canadienne de Psychologie Expérimentale, 52(4), 213–216. https://doi.org/https://doi.org/10.1037/h0087294
- Richardson, D. C., & Gobel, M. S. (2015). Social attention. In J. M. Fawcett, E. F. Risko, & A. Kingstone (Eds.), The handbook of attention (pp. 349–367). The MIT Press.
- Ristic, J., Friesen, C. K., & Kingstone, A. (2002). Are eyes special? It depends on how you look at it. Psychonomic Bulletin & Review, 9(3), 507–513. https://doi.org/https://doi.org/10.3758/BF03196306
- Rudman, L. A. (1998). Self-promotion as a risk factor for women: The costs and benefits of counterstereotypical impression management. Journal of Personality and Social Psychology, 74(3), 629–645. https://doi.org/https://doi.org/10.1037/0022-3514.74.3.629
- Sebanz, N., Bekkering, H., & Knoblich, G. (2006). Joint action: Bodies and minds moving together. Trends in Cognitive Sciences, 10(2), 70–76. https://doi.org/https://doi.org/10.1016/j.tics.2005.12.009
- Sedikides, C., & Brewer, M. B. (2001). Individual self, relational self, collective self. Psychology Press.
- Sinclair, S., Lowery, B. S., Hardin, C. D., & Colangelo, A. (2005). Social tuning of automatic racial attitudes: The role of affiliative motivation. Journal of Personality and Social Psychology, 89(4), 583–592. https://doi.org/https://doi.org/10.1037/0022-3514.89.4.583
- Skarratt, P. A., Cole, G. G., & Kingstone, A. (2010). Social inhibition of return. Acta Psychologica, 134(1), 48–54. https://doi.org/https://doi.org/10.1016/j.actpsy.2009.12.003
- Surtees, A. D. R., Lewis-Keen, A., An, J., & Evison, M. (2021). When do I see it your way: Individual, interpersonal and contextual influences on spontaneous perspective taking. Manuscript under review.
- Tajfel, H., & Turner, J. C. (2001). An integrative theory of intergroup conflict. In M. A. Hogg & D. Abrams (Eds.), Key readings in social psychology. Intergroup relations: Essential readings (pp. 94–109). Psychology Press.
- Tipper, C., & Kingstone, A. (2005). Is inhibition of return a reflexive effect? Cognition, 97(3), B55–B62. https://doi.org/https://doi.org/10.1016/j.cognition.2005.02.003
- Tufft, M. R. (2020). All about generating social context online [Webinar]. Behavioural Science Online. https://beonlineconference.com/all-about-generating-social-context-online
- Tufft, M. R., Gobel, M. S., & Richardson, D. C. (2015). Social eye cue : How knowledge of another person ‘s attention changes your own. In D. C. Noelle, R. Dale, A. S. Warlaumont, J. Yoshimi, T. Matlock, C. D. Jennings, & P. P. Maglio (Eds.), Proceedings of the 37th Annual Meeting of the Cognitive Science Society (pp. 2440–2445). Cognitive Science Society.
- Tufft, M. R., & Richardson, D. C. (2020). Social offloading: Just working together is enough to remove semantic interference. In S. Denison, M. Mack, Y. Xu, & B. C. Armstrong (Eds.), Proceedings of the 42nd Annual Conference of the Cognitive Science Society (pp. 859–865). Cognitive Science Society.
- Tyler, T. R., & Blader, S. L. (2001). Identity and cooperative behavior in groups. Group Processes & Intergroup Relations, 4(3), 207–226. https://doi.org/https://doi.org/10.1177/1368430201004003003
- Van Vugt, M., De Cremer, D., & Janssen, D. P. (2007). Gender differences in cooperation and competition: The male-warrior hypothesis: Research report. Psychological Science, 18(1), 19–23. https://doi.org/https://doi.org/10.1111/j.1467-9280.2007.01842.x
- Welsh, T. N., Elliott, D., Anson, J. G., Dhillon, V., Weeks, D. J., Lyons, J. L., & Chua, R. (2005). Does Joe influence Fred’s action? Inhibition of return across different nervous systems. Neuroscience Letters, 385(2), 99–104. https://doi.org/https://doi.org/10.1016/j.neulet.2005.05.013
- Welsh, T. N., Lyons, J., Weeks, D. J., Anson, J. G., Chua, R., Mendoza, J., & Elliott, D. (2007). Within- and between-nervous-system inhibition of return: Observation is as good as performance. Psychonomic Bulletin & Review, 14(5), 950–956. https://doi.org/https://doi.org/10.3758/BF03194127
- Yarbus, A. L. (1965). Eye movements and vision. Plenum Press.