
ABSTRACT
This paper takes a critical approach to understanding the social and cultural ‘work’ of natural heritage conservation, focussing specifically on ex-situ biodiversity cryopreservation practices. Drawing on ethnographic fieldwork with the Frozen Ark, a UK-based ‘frozen zoo’ aiming to preserve the DNA of endangered animal species, the paper situates the development of non-human animal biobanks in relation to current anxieties regarding the anticipated loss of biodiversity. These developments are seeding new global futures by driving advances in technologies, techniques and practices of cloning, de-extinction, re-wilding and potential species re-introduction. While this provides impetus to rethink the nature of ‘nature’ itself, as something which is actively made by such conservation practices, we also aim to make a contribution to the development of a series of critical concepts for analysis of ex-situ and in-situ natural heritage preservation practices, which further illuminates their roles in building distinctive futures, through discussion of the relationship between conservation proxies, biobanking and biocapitals. We suggest that questions of value and the role of future making in relation to heritage cannot be disassociated from an analysis of economic issues, and, therefore, the paper is framed within a broader discussion of the place of ex-situ biodiversity cryopreservation in the late capitalist global economy.
Introduction
The urge to preserve for future generations things that humans consider to be under direct or indirect ‘threat’ has a long tradition in the context of both ‘natural’ and ‘cultural’ heritage practices (e.g. Holdgate Citation1999; Lowenthal Citation1985; Sodikoff Citation2012), a tradition in which biodiversity conservation – as the dominant concept by which natural heritage values have been articulated, enumerated and understood since the 1980s in relation to processes of species extinction – plays an important role (e.g. see Meskell Citation2012; Sepkoski Citation2016; Bargheer Citation2016). Much like the urge to preserve objects, places and traditions of shared cultural significance for a yet-indeterminate time-to-come, efforts to freeze biomaterials from threatened species for the potential use in (as yet) unknown future scenarios are often rooted in fundamentally anthropocentric rationales, driven by the conception that ‘biodiversity’ constitutes a crucial piece in the puzzle of sustained human survival (Vidal and Dias Citation2016, 9 and 13). The conservation of non-human animal species and their natural habitats as an ‘insurance’ measure in the present is, however, not only directed at ‘what-could-become’ – an imminent present – but is equally, if not primarily, directed towards actively intervening in the production of more or less distant futures. Within this context, biobanks are future-making institutions (Harrison Citation2017) that have the potential to create new ecological, social and economic realities. Indeed, such biobanks are ‘live archives’ and actively intervene within the assumed entropic relationship between diversity loss and time (e.g. Radin Citation2015; Chrulew Citation2017; 293; Harrison Citation2017), producing multiple dormant possibilities or, what Joanna Radin has called ‘latent futures’ (Citation2013).
Drawing on ongoing ethnographic researchFootnote1 with the Frozen Ark (a UK-based, international biobanking initiative which aims to preserve the DNA of endangered animal species) and with affiliated conservation biologists at Cardiff University, this article identifies and observes a contemporary shift in the meaning of the practice of freezing genetic material of non-human animals. From an initial ‘heroic’ narrative that cast such biobanks in a static, dormant role – isolated arks to carry endangered DNA into an uncertain future (Doyle Citation1997; Watson and Holt Citation2001; Bowkett Citation2009; Chrulew Citation2017) – we detect a recent shift to a more active function which acknowledges their potential for reanimation of genetic material in future de-extinction programmes. We suggest that the role of such institutions has transformed from repository to speculative reinvestment: the ‘arks’ that stored and safeguarded genetic samples for survival within an endangerment narrative (c.f. Turner Citation2007) have altered to become ‘investment banks’ where genetic materials can be actively reworked and revived to build new futures (see also Bowker Citation2005; Heatherington Citation2012; Chrulew Citation2017). Where the new forms of biocapital generated with such repositories seem to reflect reformulations of late capitalist values (e.g. Doyle Citation1997; Shukin Citation2009; Thacker Citation2010), in this paper we consider the ways in which a critical perspective on the operations of these enterprises might help us to bring new insights to bear on the latent possibilities contained within these reservoirs of frozen DNA. Further, we explore the ways in which concepts derived from this work might be applied to comparative analysis of ex- and in-situ natural and cultural preservation practices as part of comparative studies which aim to illuminate their roles in building distinctive futures (see also Harrison Citation2015, Citation2017; Harrison et al. Citation2016). Within this context we are particularly interested in those that emerge from our examination of the relationship between extinctions, proxies and forms of biocapital, which in turn draws on Latour’s (Citation1987, Citation1999) discussion of the role of immutable and combinable mobiles in the history of the sciences. Given the significant role which the study of values has assumed within critical cultural heritage studies, we suggest that the production of these new forms of biosocial values constitutes an important subject for critical analysis of contemporary biodiversity and natural heritage conservation.
Biodiversity and the crisis of the sixth mass extinction
The notion that the Earth is currently in the midst of the ‘Sixth Mass Extinction’, resulting in what has been described as a ‘biodiversity crisis’ (Singh Citation2002) – a global loss of diversity in flora and fauna – is now in wide public circulation, driving academic research, public policy, and directing the course of nature conservation efforts globally. Mass extinctions are defined by biologists as the disappearance of three-quarters of the Earth’s species in a geologically short time span (Barnosky et al. Citation2011, 51). Although it has been observed that species have regularly gone extinct throughout the millennia through non-anthropogenic (or at least, primarily non-anthropogenic) processes – processes which Darwin famously saw as a slow and gradual but inevitable and progressive, driven by natural selection and the ‘survival of the fittest’ (e.g. see Sepkoski Citation2016) – human influence in the form of over-hunting and habitat depletion are today understood to be playing a central role in the current ‘chronic disaster’ (Westley Citation1997) of animal extinction rates (Fletcher Citation2008). Anthropogenic climate change is seen to constitute a key threat multiplier to these factors (Cahill et al. Citation2012). Paleobiologists warn that no matter on what scale these anthropogenic extinctions take place, ‘biodiversity’ will need millions of years to recover (Kirchner and Weil Citation2000, 177).
But what does the term ‘biodiversity’ mean in this context? Since its introduction as a shorthand for ‘biological diversity’ by conservation biologists Walter G. Rosen and E. O. Wilson in 1986 (Wilson Citation1988), the concept of ‘biodiversity’ has come to stand in for a range of different conservation targets, from individual species to landscapes and ecosystems, and the primary means by which the values of ‘natural’ heritage are articulated. Generally used to refer to and measure all categories of biological diversity and its abundance, including everything from ‘alleles, to populations, to species, to communities, to eco-systems’ (Sarkar Citation1999; 405; see also CBD Citation1992), biodiversity has also acted as a more ‘scientific’ synonym for nature. Generally, biological diversity, the ‘web of life’ (CBD Citation2000), is considered to be something intrinsically good, the protection of which is paramount for the survival of humanity:
Biological resources are the pillars upon which we build civilizations…The loss of biodiversity threatens our food supplies, opportunities for recreation and tourism, and sources of wood, medicines and energy. It also interferes with essential ecological functions (www.cbd.int/convention)
Still, the meaning of the term ‘biodiversity’ remains ambiguous, its definition continuously re-moulded to fit the agenda and situation of the stakeholder(s) involved (Swingland Citation2001), but always framed in relation to broader narratives of extinction and loss. In the process, biodiversity has become a powerful concept employed across the natural and social sciences, as well as governmental and corporate agencies, to justify and frame interventions in, and actions upon the ‘natural’ world (Bowker Citation2005). In the field of ecology and conservation biology, the concept of biodiversity is conventionally approached as something that is of universal value and in need of protection (Perrings, Folke, and Mäler Citation1992), and, like cultural heritage (Harrison Citation2016) as part of what Vidal and Dias term the ‘endangerment sensibility’, a network of ‘scientific practices [that] belong in a complex of knowledge, values, affects and interests characterised by a particularly acute perception that some organisms and things are under “threat”, and by a purposeful responsiveness to such a predicament’ (Vidal and Dias Citation2016, 2).
Classifying biological diversity as endangered involves a conscious decision to put it into a specific category that carries within itself connotations of loss, depletion and forgetting – negative or ‘anti-values par excellence’ (Vidal and Dias Citation2016, 1) – that force one to see biodiversity as potentially entropic, and thus in need of saving (c.f. Harrison Citation2017; see also Sepkoski Citation2016). To this way of thinking, biodiversity becomes something which is always inherently necessarily diminishing and thus followed by the persistent ‘shadow of extinction’ (Van Dooren Citation2014, 8), and for which humans act as saviour of the victims of their own misdoings. Here, we draw on Van Dooren’s definition of extinction, as a slow unravelling of entangled lives that does not represent a single event but the unmaking of an inter-linked ecological and social web of human and non-human species (Van Dooren Citation2014; on the emerging field of extinction studies see also Bird Rose Citation2011; Sodikoff Citation2012; Heise Citation2016; Bird Rose, Van Dooren, and Chrulew Citation2017). The important point to note is the way in which biodiversity conservation as a concept designates itself simultaneously as a field of enumeration – in the sense in which it involves bureaucratic practices of identifying, counting and accounting for certain conservation targets in natural heritage management – and intervention, in the sense in which it justifies and predicates certain actions on those conservation targets and/or the field(s) in which those targets are contained (see also Bowker Citation2000). There are clear parallels here with observations made of cultural heritage as defined by endangerment and ‘risk’ (e.g. Harrison Citation2013; Rico Citation2015, Citation2016]; see also Bennett et al. Citation2017).
The predicted loss of two-thirds of the world’s vertebrae population by 2020 (WWF Citation2016) and a similarly bleak outlook for invertebrate species, has intensified biodiversity conservation efforts globally. These take the form of both in-situ conservation programmes (e.g. through the designation of protected areas) and ex-situ captive breeding programmes (e.g. in zoos and aquaria). More recently, these ex-situ conservation efforts have accelerated, as a result of the DNA ‘revolution’, through the development of organised archives of non-human animal biomaterials which aim to document and preserve genetic information on the biology, ecology and evolutionary history of threatened mammals, birds and reptiles in the form of viable cells and DNA preparations, before it is irretrievably lost (Corley‐Smith and Brandhorst Citation1999; Watson and Holt Citation2001; Friese Citation2013; Costa and Bruford Citation2018). Genetic resource banking – the freezing of plant and animal genetic material for ex-situ storage and its use in research within a present-day and potential future context – has emerged as a response to what has been understood to be a contemporary extinction crisis, and in many cases cryobanks have come to be seen as the only and last resort for recording and storing biological material from endangered species for potential future retrieval.
Making and mobilising frozen futures: cryopreservation and biodiversity conservation
The practice of freezing and storing of biological material (including blood, germplasm, embryos, tissues and somatic cells of non-human animals) in genetic resource banks for the advancement of human medicine and the development of agro-industries is not a new development in scientific research (see further discussion in Radin Citation2015, Citation2017; Radin and Kowal Citation2017). Nonetheless, it is only recently that ex situ cryogenic practices have become a leading and driving force in biodiversity research within the context of endangered species conservation (see Gemeinholzer et al. Citation2011; Wildt et al. Citation1997; see also Howard et al. Citation2016; Wisely et al. Citation2015; on the ferret biobank) – with biobank facilities such as the Smithsonian, the San Diego Frozen Zoo® and the genetic repository at the National History Museum in London collecting blood, tissues, cell cultures, eggs, spermatozoa and embryos specifically for conservation purposes. According to its website, the Zoological Society of San Diego (ZSSD) Center for Reproduction of Endangered Species (CRES) now stores ‘the largest and most diverse collection of its kind in the world’ with over ‘10,000 living cell cultures, oocytes, sperm, and embryos representing nearly 1,000 taxa, including one extinct species, the po’ouli’.Footnote2 Since its foundation in 1975, the San Diego Frozen Zoo® has become an irreplaceable and continuously expanding source of biological information for significant scientific advancements in fields such as conservation, medicine, assisted reproduction, evolutionary biology, physiology and wildlife medicine (Chemnick, Houck, and Ryder Citation2009).
In a paper on the Frozen Zoo® concept published in 1984, Benirschke advocates that ‘biologists at zoological gardens have a unique opportunity – if not an obligation – to preserve materials for scientific study. At a time when biomedical capabilities are expanding rapidly, we find ourselves in the position that biological resources are dwindling rapidly. Many forms of life are at the point where extinction is imminent, yet the animal or plant has not become understood in any of its major biological ways’ (1984, 325, our emphases). Benirschke’s words convey an urgency to not only save dwindling genetic material for scientific study in the present but to safeguard it for an undetermined future in which humans will be in a better position to extract from it as-yet uncovered information (see also Morgan and Macdonald, this issue). (They also provide the key to understanding the role of such facilities in contributing to the growth of new forms of biocapital, as we will discuss later in the paper). Here, cryobanks become the harbourers of ‘time-travelling resource[s]’ (Radin Citation2017), which are both enactors of, and produce templates for ‘futures in the making’ (Adam and Groves Citation2007; Turner Citation2007). Genetic resources of endangered animals, for example, have enabled developments in reproductive technologies to maintain genetic diversity which have already produced promising ‘real-life’ results in a number of conservation programmes (e.g. Howard et al. Citation1992; Wildt et al. Citation1997). As such, frozen zoos and other non-human biobanks are driving ongoing research into cloning, de-extinction and re-introduction of endangered and once extinct species (see further discussion in O’Connor Citation2015; Shapiro Citation2015; Pilcher Citation2016). These developments are likely to have significant impacts on what we might now, in the light of the recognition of the Anthropocene epoch, term the ‘Human Planet’ (Lewis and Maslin Citation2018).
The Frozen Ark
The Frozen Ark Project was originally set up in 2004 at the University of Nottingham by the late geneticist Professor Bryan Clarke FRS and his wife, immunologist Dr Ann Clarke, as well as developmental biologist Dame Anne McLaren, in an effort to preserve the genetic resources of threatened wild species kept in zoos and aquaria around the world before they could go extinct (see Clarke Citation2009; Costa and Bruford Citation2018), assuming this extinction was a given. From the 1960s, Bryan dedicated much of his time to work on speciation in the Partula land snail, native to the volcanic islands of French Polynesia. It was around the same time that Lissachatina fulica, the giant African land snail was brought to the island of Tahiti for breeding purposes as a culinary delicacy – a decision with serious consequences. The snail spread to neighbouring islands and, in the absence of any natural predators, bred at such an alarming rate that the government decided to introduce the carnivorous Florida Rosy wolfsnail (Euglandina rosea) to the island to control the quickly escalating agricultural pest. However, instead of eating its targeted prey, the new snail ravaged the Partula snail population and within 15 years, the latter had disappeared from most of the islands (Clarke, Murray, and Johnson Citation1984). Realising the species’ imminent threat of extinction from human interference, Clarke and his team decided to collect live specimens of the remaining twelve Partula species, bring them back to the UK and freeze tissue samples in order to preserve their DNA and allow for further study; they also established an international captive breeding programme at the Zoological Society of London (ZSL) in the hope of re-introducing the snail to their natural habitat. The fate of the Partula was playing out before their very eyes and the importance of collecting and preserving biological material from endangered animals eventually resulted in the creation of the Frozen Ark Project (Costa and Bruford Citation2018).
The founding partners of the UK registered charity include the London National History Museum and the Zoological Society of London, as well as the University of Nottingham, which provides laboratory and office space and serves as the seat of the Frozen Ark, while research is now mainly being carried out at Cardiff University. At the time of writing its consortium of zoos, aquaria and other conservation bodies counted 27 national and international partners from all over the world (Costa and Bruford Citation2018). The apocalyptic message conveyed by the project’s logo (), a ‘stylised ark on stormy seas’ (Chrulew Citation2017), is both clear and urgent: in the face of anthropogenic ecological loss, the collecting, storing and managing of biological material from endangered species might be the only chance for humanity and the species with which we co-habit the planet. Yet, unlike Noah’s Ark which, according to Genesis flood narrative carried a male and a female of all the world’s animals to save them from extinction by drowning, the Frozen Ark is a ‘cryogenic’ or ‘technoscientific ark’ (Parry Citation2004) that adheres to its website’s motto of ‘saving cells and DNA of endangered species’ – materials which act as ex-situ proxies of the living species they were taken from (see our discussion of proxies below).
Figure 1. The Frozen Ark logo (© The Frozen Ark).

The University of Nottingham currently provides two −80°C freezers storing just over 700 blood and tissue samples obtained from endangered non-human animals from UK-based zoos and aquaria. The charity’s collection in Nottingham consists of over 700 samples from a number of different animals including the scimitar horned oryx (extinct in the wild), the Colombian spider monkey, pileated gibbon, siamang gibbon, lar gibbon, snow leopard and Malayan tapir (all endangered). These are held as cultured cells, tissue and gametes stored in liquid nitrogen (see ). When our researchers visited the Nottingham laboratory we were shown how information on all the samples stored there is organised in physical file folders and includes, amongst other details, an internal identification number, a universal zoo number, the species, type and location of sample, what it is preserved in, sample quality, and, where applicable, a Whatman FTA card. The Frozen Ark’s interim Director, based in Cardiff, indicated that the ultimate objective is to form a confederated model that functions as both a physical and virtual infrastructure, storing and managing the genetic material from endangered species, sampled in the wild and in zoos and aquaria, from all over the world. At the time of writing, CryoArks, a Cardiff-based and BBSRC-funded (UK Biotechnology and Biosciences Research Council) initiative resulting from a collaboration between the Frozen Ark and some of its partner institutions as well as the UK node of the EAZA (European Association of Zoos and Aquaria) biobank, is in the process of being established. Due to limitations imposed by the Nagoya Protocol, which, according to the Convention on Biological Diversity website, ensures ‘the fair and equitable sharing of benefits arising from the utilisation of genetic resources, thereby contributing to the conservation and sustainable use of biodiversity’, CryoArks will be mainly focused on the UK and Ireland, whereas the Frozen Ark has a global remit – to this end it has already started cataloguing samples of extinct, endangered and threatenedFootnote3 species held by consortium members, and is aiming to increase the number of, and coordination between, consortium members. Unlike other biobanks around the world (e.g. San Diego Frozen Zoo, Smithsonian Biobank, Svalbard Global Seed Vault), which intend to form a single point on Earth where genetic material from all over the world is being stored inside a central biobank, the Frozen Ark aims to be a Nagoya-compliant back-up storage facility for institutions that, due to various reasons, cannot store their own samples or would like to have duplicates of existing collections and to hold centralised records relating to a distributed network of physical biobanks which store biosamples of endangered non-human animals.
Figure 2. Inside one of the −80°C freezers storing samples of biological material from endangered non-human animals at the Frozen Ark in Nottingham (Photograph by Esther Breithoff).

Based on interviews with the charity’s staff undertaken during the lab placement in Cardiff and subsequently with staff based in Nottingham, the Frozen Ark’s concern for preservation of genetic material for future generations initially outweighed active conservation efforts. With species going extinct all over the world and the dramatic anticipated loss of genetic information, the Frozen Ark eventually decided to change from acting purely as a repository to become an active collection. This decision seems to have been influenced partially by the emergence of new experimental genetic work, but also reflected a change in philosophy about the Ark’s role. ‘The focus was always for the future’ reflects Jude Smith, who has been the charity’s administrator from the beginning, ‘but as we’ve got on, it has become really obvious that the future is here now, you know, it’s now’ (interview with Esther Breithoff, 31 August 2017).
This new approach, described as more ‘pragmatic’ by current Interim Director, Professor of Biodiversity and Conservation geneticist Mike Bruford (interviews with Esther Breithoff, 18 and 21 July 2017), recognises the need to boost the profile of the charity in order to deliver on its promises for the future: the collection, safeguarding and managing of biological and genetic material from endangered species for both anticipated and unanticipated future uses. The vision is for the Frozen Ark to become an active and ethical facility for genomic resource management that helps identify and prioritise which animal species are at risk of extinction and thus in need of sampling, and develop the most effective techniques of collecting, storing and managing biological material. In its educational role, the Frozen Ark supports institutions both in the UK and abroad with setting up their own biobank facilities and/or successfully managing already existing repositories. According to the charity, its main goals are:
i) coordinating global efforts in animal biobanking; (ii) sharing expertise; (iii) offering help to organisations and governments that wish to set up biobanks in their own countries; (iv) providing the physical and informatics infrastructure that will allow conservationists and researchers to search for, locate and use this material wherever possible without having to resample from wild populations (Costa and Bruford Citation2018).
In the current absence of coordination and lack of shared protocols and databases between different biobanks nationally and internationally, the Frozen Ark plans on setting up a virtual stand-alone open-access database connecting existing biobanks on a global level. This would facilitate increased access to research material for researchers and conservationists internationally. The Frozen Ark sees its role in safeguarding and managing genetic diversity as part of a joint effort between ex-situ and in-situ conservation practices. Cryostoring biomaterial of endangered species in freezers and liquid nitrogen tanks – although space effective – does however come with a high carbon footprint, which one could suggest ultimately increases the threat of extinction to the animals it was designed to protect. The Frozen Ark website emphasises that establishing and maintaining a global biobank at present is also a costly undertaking that has been suffering from a lack of funding since its inception.
Time is running out for many species. Conservation efforts will undoubtedly save some but we must preserve the genetic record of all endangered species for our future. Time is also running out for the Frozen Ark, which has been running with volunteers on a shoestring budget for several years. Help us save Nature’s genetic heritage so that future generations can enjoy the natural world as we have all done. (The Frozen Ark Citation2018)
Like the endangered species whose biological material it aims to secure in the race against irretrievable loss of biodiversity, the Frozen Ark itself also senses a risk of its own endangerment in articulating these difficulties of establishing long term funding to secure its future operations. These issues of uncertainty relating to the securing of ongoing financial resources for the organisation’s research and collections were a regular topic of discussion; in the laboratory, in conferences, and in more formal interview contexts; and form another of the various ways in which the urgency of the work of the organisation, and biodiversity conservation more generally, is expressed.
Biodiversity and its proxies
We have already noted that the concept of biodiversity is an ambiguous and often plastic one, capable of describing multiple phenomena at different scales, mobilising a range of different activities in and on individuals and collectives of both plants and (both human and non-human) animals, and adapted to a range of different political circumstances. It encodes complicated relationships between parts and wholes in the sense in which the diversity in biodiversity is so normative that it becomes impossible to imagine a scenario in which an ecosystem, landscape, national park or other unit of analysis could ever be biodiverse enough – it represents a yearning for completeness and infinitude which, like that of other universal categorical systems (Bowker and Star Citation2000), constitutes an impossible goal. It is realised by way of indices, measurements, and catalogues of species, designated at various degrees of risk (Turner Citation2007; Heise Citation2016), and is often represented metonymically as certain charismatic species come to stand in for the concept of biodiversity more generally. A familiar example is the individual starving polar bear as a symbol of the environmental impacts of anthropogenic climate change. Captivatingly staged studio photography of individual animals and apparent family groups from endangered species – taken by renowned animal photographer Tim Flach and often assuming anthropomorphised expressions with the animal looking direct into the camera – forms an important part of the Frozen Ark’s promotional materials. Such metonymic relationships can be achieved in other ways too. For example, Carrithers et al. (Citation2011), explore how the ‘species as person’ trope in conservation discourse allows similar anthropomorphised relations to be realised for less charismatic species. This is also typical of the normative nature of arguments regarding the value of diversity in which diversity is understood to represent a value which is in itself inherent (Heyd Citation2010).
This is one way of understanding how biodiversity as a concept is realised by way of proxies which represent units – individual animals, species, protected areas, ecosystems – on which certain forms of conservation processes might be enacted. The term ‘conservation proxy’ has a specific meaning in conservation biology, and describes how certain units of analysis (species, landscapes, soils, etc) act as indicators of the relative ‘health’ or diversity of a particular system or larger unit of analysis – a national park, an ecosystem, a waterway, etc. These surrogates or flagship species become a shorthand measurement to guide the distribution of resource or to measure the intensity of action which is required on a particular system (e.g. Caro Citation2010). The term has a slightly different meaning in rewilding discourse, where it refers to the introduction of contemporary species which stand in for extinct ones, mimicking an extinct animal’s role in the ecosystem. This is sometimes also referred to as ‘taxon substitution’ (e.g. Josh et al. Citation2006).
There is another, equally important way of understanding the concept of proxies in relation to ex-situ biodiversity conservation practices. This is in the way in which certain kinds of biological materials, in combination with different forms of data pertaining to those materials, come to stand in for the species from which those materials have been taken (Parry Citation2004; Van Dooren Citation2009). The transformation of such objects into the targets of conservation activity themselves can be understood in relation to Latour’s concept of ‘immutable mobiles’ in the history of the development of modern sciences. These are transportable textual or visual fragments, observations and representations of objects, places and phenomena which are collected from ‘the field’ and returned to ‘centres of collection and calculation’ where they are combined with other such objects and subjected to translation to produce forms of scientific knowledge which predicate action, including action back upon the fields from which they were collected (Latour Citation1987, Citation1999). In the case of the Frozen Ark, biological materials (including cells and DNA samples) act as ex-situ proxies, and these biomaterials, their containers, and the data pertaining to them increasingly become themselves the focus of conservation activity (see –). But the application of these concepts to the Frozen Ark also suggests certain ways in which Latour’s model might be modified to accommodate more complicated sets of relations which have arisen in the late modern bioeconomies in which such biobanks have come to proliferate. First, the case of the Frozen Ark suggests that Latour’s conception of ‘the field’ might be broadened to accommodate collections made from existing captive collections. The genetic material kept at the Frozen Ark is not only harvested from living and dead animals in the ‘wild’ (often national parks and other protected areas), but also from zoos, aquaria and other ex-situ living collections. Accepting that ‘nature’ is both social construction (e.g. Wilson Citation1992) and itself physically managed and humanly produced (Lewis and Maslin Citation2018), we suggest that late modern ‘fields’ might not always be remote, as in Latour’s model of the history of the modern field sciences, but might instead constitute proximal spaces from which certain resources might be harvested and set into motion for other uses through the application of technosciences (Heatherington Citation2012), the ‘distant’ microscopic fields of which can only be reached with the intervention of specific technical imaging devices which require complex computational interpretation to visualise. What is significant here is not the geographical distance of the field, as in La Perouse’s journey which forms the basis for Latour’s arguments in Science in Action (1987), but the application of certain forms of highly specialised technoscience to visualise, isolate, extract, store and manipulate biomaterials which would not otherwise be able to be collected from these living collections. Second, because the Frozen Ark is composed of a consortium of zoos, aquaria and other conservation bodies, it initially seems problematic to use the term ‘centre of calculation’ to describe it. But like other collections (e.g. see the discussion of the relations of French metropolitan and colonial museums in Bennett et al. Citation2017), centres of calculation may be distributed and operate across multiple centres as part of a confederated model. Importantly, these are held together by virtual networks, in this case databases containing information relating to samples of biomaterials from endangered non-human animals, which manage and constrain the ways in which data is collected and which render them able to be combined and manipulated in specific ways.
Figure 3. The FTA® is a paper-based system that allows the collection and storage of DNA (mostly from blood and buccal swabs) by dabbing a bit of the fluid into one of the pink circles inside the card. It allows a quick and safe storage of DNA without the need of liquid nitrogen and freezers, making them a practical alternative when sampling in the field. Unfortunately, the cards are expensive and only allow 4 samples per card. Such technologies make the collection, transportation and subsequent storage of DNA/biomaterials as conservation proxies possible (Photograph by Esther Breithoff).

Figure 4. Lab book entry showing PCR data (amplifications of a known section of DNA often used to either identify the presence or absence of a genetic trait) and agarose gel images (the florescent rectangles visible in the images identify the presence or absence of a genetic trait), which act as proxies for the flycatcher bird that the DNA was taken from for a study at the School of Biosciences, University of Cardiff (Photograph by Esther Breithoff).

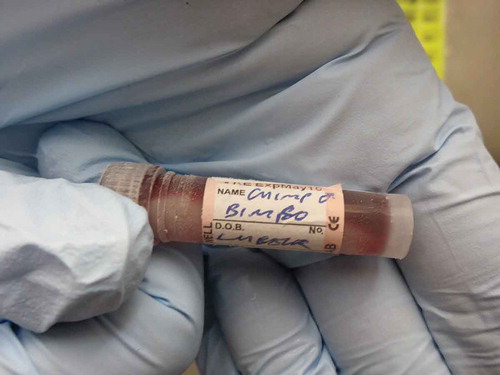
Figure 5. Frozen blood sample of a chimp, stored in one of the Frozen Ark’s −80°C freezers at the University of Nottingham, as a further example of a biomaterial proxy of the animal it was extracted from (Photograph by Esther Breithoff).
These more complicated sets of relations are similarly realised through the Ark’s relations with other institutions, where it acts as a ‘back up’ or copy of a copy held in other biobanks. Even though these are ‘copies’, they are also perceived as authentic or ‘immutable’ at the level of the DNA sample, which could be conceived as divided, rather than replicated (see further discussion of ‘copies of copies’ below). The conservation of such biomaterials is a form of ‘latent’ or ‘deferred’ conservation in the sense in which these cells and data are frozen for subsequent scientific research which may aid conservation efforts in some, as yet undefined manner. In this way, such biomaterials exemplify a process which could be understood to be common to ex-situ natural and cultural heritage conservation more generally.
Playing god in the Anthropocene: biodiversity, cryopreservation and future-making
In 1993, the Stephen Spielberg film Jurassic Park seemed to offer an improbable view of an alternative future in which long extinct species could be regenerated from ancient DNA. We have shown that initiatives to collect and store the raw materials for such a process in the form of frozen blood, tissue and other human and nonhuman animal organic materials have a much longer genealogy. However, recent developments in genetic rescue programmes which aim to revive extinct and threatened animal species suggest such genomic engineering is scientifically possible. Several projects which sound equally implausible – including work currently being undertaken by Revive and Restore (www.reviveandrestore.org) to clone extinct Passenger Pigeons and Woolly Mammoths – are likely to realise results within the next decade (e.g. see Jørgensen Citation2013; Shapiro Citation2015). Sherkow and Greely (Citation2013) explain that the three approaches which appear most likely to yield results are back-breeding, in which selective breeding is used to produce the phenotypes of extinct species; cloning using cryopreserved tissue; and genetic engineering using whole genome sequencing and the editing of DNA in cells from genetically similar extant animals. In many ways these projects constitute a realisation of the latent futures which are resourced by frozen zoos and cryopreservation technologies. The move within the Frozen Ark away from perceiving its role primarily as a passive collecting institution for the future, to one of active experimental conservation in the present, exemplifies the ways in which such collections resource the development of new realities in which the possibilities of reviving extinct species through hybridisation with extant ones is increasingly becoming fulfilled. But in their enabling of certain forms of what Vidal and Dias helpfully terms ‘restitution fantasies’ (Citation2016, 1) they also re-enforce dominant (although not uncomplicated – see Dibley Citation2012, Citation2015) forms of anthropocentrism which remain barely hidden within the Anthropocene chronotope (c.f. Pratt Citation2017) in the fulfilment of humanity’s ultimate mastery over nature: the ability to resurrect the species that we have ourselves rendered extinct. The quest for such a reality is embodied in the Frozen Ark’s own creation narrative in which the founders’ attempts to save the Partula land snail through more conventional methods of captive breeding are unsuccessful and force them to turn to cryopreservation for future hybridisation and de-extinction programmes as the last hope for this totemic species.
The complicated ethical questions raised by such projects, and their reception by different publics, are only now beginning to be explored (e.g. see National Academies of Sciences, Engineering, and Medicine Citation2016). There are significant opportunities for important conversations across natural and cultural heritage practice here around persistent questions relating to authenticity, values, and the ways in which such natural and cultural ‘restoration’ projects actually constitute active forms of future making practices at both the micro (genetic) and macro (landscape) scale. Genetic biobanking does so in the sense in which it provides literal raw materials for the development of scientific experimentation and new technologies which might realise de-extinction. These debates similarly touch on a range of discussions within the field of nature conservation, especially those relating to re-wilding (e.g. Lorimer Citation2015; DeSilvey and Bartolini Citation2018), and more radically, to ‘terraforming’ (e.g. Sagan (Citation1961, Citation1973); Oberg Citation1981; Fogg Citation1995). Significantly, many of these actual and hypothetical developments have been prefigured within speculative fictions of different genres (for example in the work of Margaret Attwood) in a process which we see as analogous to the relationship between the rhetorics of cryopreservation (Chrulew Citation2017) and the new realities they resource which we have discussed in this paper.
From ark to bank: biodiversity and biocapital
It is in the transformation of these latent possibilities into new economic (as well as ecological) realities that we are able to determine shifts in the nature of biobanking facilities and the forms of value they both generate and are caught up within (Shukin Citation2009). A significant literature in science studies, which develops and expands upon Foucault’s ‘late’ work on biopower/biopolitics, has traced the development of what Cooper (Citation2008) terms the ‘bioeconomy’ since the 1970s in the specific relations of biotechnology, neoliberal politics and economic policy (e.g. see Doyle Citation1997; Thacker Citation2006; Rose Citation2006; Waldby and Mitchell Citation2006; Shukin Citation2009; Franklin Citation2013; Cooper and Waldby Citation2014). Central to the bioeconomy has been the emergence and evolution of a range of new forms of ‘biocapital’. We draw on Helmreich’s (Citation2008; see also Rajan Citation2006) definition of biocapital as the surplus values generated by the commodification and circulation of forms of biological life within economic systems. Helmreich points out, however, that biocapital is understood and deployed in a number of different ways by scholars across science studies and itself may manifest in a range of different forms, as parts of different sociomaterial assemblages. It is the ways in which biocapital emerges flexibly and replicates itself across these different sociomaterial assemblages which concerns us here. Given the significance of the study of concepts of value to critical heritage studies, we might ask how cryobanks such as the Frozen Ark have contributed to the development of new forms of value? And in what ways are those new values accumulated and distributed within the bioeconomy?
We return here to Benirschke’s (Citation1984) discussion of the Frozen Zoo concept cited earlier in the paper in his observation of the relationship between the growth of cryopreservational technologies and the dwindling biological resources these are produced to conserve. As biodiversity (bearing in mind that this concept is itself plastic and subject to shifts in meaning) diminishes, the value of these banked biomaterials increases both individually and collectively. As Harrison (Citation2017) has argued in relation to the work of crop seed biobanks, these processes are forms of speculative biocapital accumulation, banking on, yet simultaneously imaginatively resourcing (c.f. Turner Citation2007; Thacker Citation2006, Citation2010), the development of the biotechnologies which will realise these future values. Thus extinction, biobanking, biocapital and biodiversity come to be linked in a complicated network of values within the emerging bioeconomy.
In its speculation on, and investment in the anticipation of loss, the work of the Frozen Ark (and the field of biodiversity cryopreservation more generally) can also be understood to represent a response to neoliberal economics in the ways in which it constitutes an optimisation of the use of space and resources. Cryobanking ‘represents a technically viable method for helping to conserve species biodiversity, without having to maintain large captive populations of each organism’ (Hosey, Malfi, and Pankhurst Citation2009, 319 as quoted by Chrulew Citation2017), nor, indeed, the designated landscapes in which these organisms might conventionally be preserved (as national parks, for example). As Chrulew goes on to surmise from these comments in his own discussion of the Frozen Ark, ‘the forms of preservation and exchange made possible by the frozen zoo transform the relationships between humans, animals, and technologies, reorganising space and time beyond familiar constraints in the interests of optimal efficiency and diversity’ (Chrulew Citation2017, 297). The ability of biodiversity conservation to designate conservation proxies which are immutable, combinable mobiles (in the Latourian sense) is thus central to the ways in which biobanks function within a bioeconomy to accumulate biocapital. As Harrison (Citation2017) has observed of the seeds in ex-situ seed banks, while these are conceptualised as copies of biomaterials held in other collections, or as we qualify here, not so much as copies as fragments of the original sample which remain authentic at the level of the DNA – indeed, as Chrulew (Citation2017) notes, doubles of doubles held in captivity which are themselves doubles of wild animals – they are not, in fact, duplicates, as their presence within these particular biosocial archives allows them to accumulate new forms of value, and indeed, possible new genetic characteristics which do not directly replicate those from which they were originally copied. This is again reflected in the change of perception of the function of the Frozen Ark from repository – where frozen biomaterials would be collected untouched for the future, to speculative reinvestment – where such biomaterials would be part of active and ongoing genetic experimentation with saving threatened species and potentially reversing extinction, in particular through the generation of hybrids which combine genetic materials from both living and extinct species.
Finally, cryobanking reconfigures relationships between life and death. Talking in the context of frozen genetic material from humans, Lemke observes that:
“human material” transcends the living person. The person who dies today is not really dead. He or she lives on, at least potentially. Or more precisely, parts of a human being – his or her cells or organs, blood, bone marrow, and so on – can continue to exist in the bodies of other people, whose quality of life they improve or who are spared death through their incorporation. The organic materials of life are not subordinate to the same biological rhythms as the body is. These materials can be stored as information in biobanks or cultivated in stem cell lines. Death can be part of a productive circuit and used to improve and extend life. The death of one person may guarantee the life and survival of another. Death has also become flexible and compartmentalised (Lemke Citation2012, 95).
Similarly, biotechnologies employed by the Frozen Ark allow for the breaking down of species into a range of components at the biomolecular level which allow for almost endless recombination (Doyle Citation1997; Chrulew Citation2017), further complicating the question of the relevant units by which biodiversity might be measured, and the relative values of such units and their proxies. The importance of the late capitalist context of these developments cannot be overstated. This extension of life and expansion of what constitutes biological reproduction is a function of what Cooper refers to as the bioeconomy’s transformation of biological life into surplus value (Cooper Citation2008; see also Shukin Citation2009; Thacker Citation2010). As in the case of Svalbard Global Seed Vault (see Harrison Citation2017), the operations of the Frozen Ark can be understood to accumulate and generate surplus value through reversing what are perceived to be ‘natural’ as well as humanly produced entropic processes of biodiversity decay (Sepkoski Citation2016); but importantly, the new forms of value which it produces are not simply inherent to its proxies themselves, but also derive from the latent (c.f. Radin Citation2017) potential for new and experimental forms of life they may be used to produce. In this sense, the Frozen Ark contributes to what Radin terms a form of ‘planned hindsight’ (Radin Citation2015) – it realises its own technofutures through its collecting policies in the present. Its latent generation of future value in the form of biocapital requires direct speculation upon the extinction and biodiversity loss it is created to secure the present against. The Frozen Ark counterintuitively depends upon the future biodiversity loss which it works against, but simultaneously anticipates, in its present operations.
Conclusion
Our aim in this paper has been to critically explore the field of ex-situ biodiversity cryopreservation and its place in the late capitalist global economy by exploring how biobanks speculate upon and help to realise new futures as a response to an anticipated global loss of biodiversity. Here we draw on a strong tradition of such critical analyses within the study of cultural heritage (e.g. Harrison Citation2013) and apply it to the study of natural heritage. We have used our work with the Frozen Ark as a starting point to begin to think about the reorganisation of biodiversity values which the rise of such facilities reflects, and the largely unexplored relationship between biodiversity conservation and the bioeconomy more generally. We want to be clear that this is not intended as a criticism of the work of the organisation, the individuals who work for the organisation, nor of biodiversity conservation more generally, but is ‘critical’ in a broader sense of trying to begin to understand how biodiversity conservation is caught up in systemic networks of power relations within the context of late capitalism. If biodiversity conservation is viewed through the lens of a critical exploration of the forms of value it generates and their interactions with one another, ex-situ biobanks are no longer dormant genetic ‘arks’ but rather ‘investment banks’ which accumulate and produce values through speculation upon the forms of extinction which they themselves seek to resist through their reconfiguration of post-genomic life.
We have also sought to introduce some new critical concepts to the comparative study of natural heritage preservation practices, oriented towards the understanding of such practices as forms of future making in which each set of practices is understood to produce its own distinctive future worlds. We see these concepts as potentially also helpful in rethinking cultural heritage preservation practices, especially given the arbitrary distinction between them (see Harrison Citation2015). One might argue that the newly emergent bioeconomy discussed here constitutes the logical product of a recognition of our current epoch – the Anthropocene – as one in which humans have become the primary force of global geological and climatological change (e.g. Lewis and Maslin Citation2018). These biotechnologies and their resultant post-extinction imaginaries are the ghosts which haunt the landscapes touched by the violence and conflicts of modernity, and the monsters which emerge out of the resulting hybrid human/non-human social relations which Tsing et al. (Citation2017) and their contributors see as defining the post-Anthropocene planetary poetics. Of course, the notion implied within the idea of the Anthropocene is precisely that we live in what Marris (Citation2013) terms a ‘post-wild world’. In presenting the distinctive future-making practices of this particular field of ex-situ biodiversity crypreservation, we also aim to demonstrate the value of understanding and engaging critically and comparatively with the whole range of heritage practices, and the distinctive futures they enact and resource. Given the significant transformations of both cultural and natural heritage and their broadening of scope to include an ever-increasing range of objects, species, places, ecosystem, practices and materials (and indeed, the complicated and blended relations between them) which has occurred over the past decades (c.f. Harrison Citation2013), understanding the implications of such practices seems increasingly important, and the role of a critical heritage studies in doing so, increasingly significant.
Acknowledgments
The research presented in this article draws on field visits, interviews and ongoing collaborations with Frozen Ark undertaken by the authors and Sefryn Penrose as part of a broader comparative study of natural and cultural diversity conservation practices, one of four major areas of thematic foci for the Heritage Futures research program. We thank Mike Bruford, Mafalda Costa, Jude Smith and the researchers at the School of Biosciences, Cardiff University, for their assistance and contributions to the fieldwork on which this paper draws. Heritage Futures is funded by an Arts and Humanities Research Council (AHRC) ‘Care for the Future: Thinking Forward through the Past’ Theme Large Grant (AH/M004376/1), awarded to Rodney Harrison (principal investigator), Caitlin DeSilvey, Cornelius Holtorf, Sharon Macdonald (co-investigators), Antony Lyons (senior creative fellow), and Nadia Bartolini, Sarah May, Jennie Morgan, and Sefryn Penrose (postdoctoral researchers), and assisted by Esther Breithoff, Harald Fredheim (postdoctoral researchers), Hannah Williams and Kyle Lee-Crossett. It receives generous additional support from its host universities and partner organisations. See www.heritage-futures.org for further information. RH’s work on this project is also supported by his AHRC Heritage Priority Area Leadership Fellowship Grant (AH/P009719/1). See www.heritage-research.org for further information.
Disclosure statement
No potential conflict of interest was reported by the authors.
Additional information
Funding
Notes on contributors
Esther Breithoff
Esther Breithoff is a Postdoctoral Fellow on the Norwegian Research Council funded Unruly Heritage research programme based at the Department of Archaeology, History, Religious Studies and Theology, UiT The Arctic University of Norway. Prior to this, she was a postdoctoral research associate on the Arts and Humanities Research Council (AHRC)-funded Heritage Futures research programme and the AHRC/GCRF-funded Restricted Access Pilot Project on nuclear heritage landscapes in South America, both based at the UCL Institute of Archaeology. She has a background in conflict archaeology and anthropology, with a special focus on the heritage of armed violence within a 20th century South American context. Her PhD focused on the conflict landscapes and material culture of the Chaco War. Her research interests include conflict heritage, archaeologies of the contemporary world, human-thing relationships, posthumanism and future-making practices. Her monograph War at the End of World: Conflict Heritage and the Chaco will be published by Routledge in 2019.
Rodney Harrison
Rodney Harrison is Professor of Heritage Studies at the UCL Institute of Archaeology and Arts and Humanities Research Council (AHRC) Heritage Priority Area Leadership Fellow. He is Principal Investigator of the AHRC-funded Heritage Futures Research Programme; Director of the Heritage Futures Laboratory at UCL; and leads the Work Package on ‘Theorizing heritage futures in Europe: heritage scenarios’ as part of the European Commission-funded Marie Sklodowska-Curie action [MSCA] Doctoral Training Network CHEurope: Critical Heritage Studies and the Future of Europe. He is the founding editor and editor-in-chief of the Journal of Contemporary Archaeology and was a founding executive committee member of the Association of Critical Heritage Studies. He is the (co)author or (co)editor of more than a dozen books and guest edited journal volumes and over 70 peer reviewed journal articles and book chapters. In addition to the AHRC his research has been funded by the Global Challenges Research Fund, British Academy, Wenner-Gren Foundation, Australian Research Council, Australian Institute of Aboriginal and Torres Strait Islander Studies and the European Commission.
Notes
1. The work presented here draws on a 6-week intensive placement during which Esther Breithoff worked with Frozen Ark team members and conservation biologists based in the research lab in Biomedical Sciences at Cardiff University, and a number of additional interviews and laboratory visits to Frozen Ark team members based at the University of Nottingham and with affiliated researchers at the London Zoo and Natural History Museum undertaken by both Esther Breithoff and Sefryn Penrose. During the Cardiff placement, Breithoff shadowed staff and students in the laboratory, interviewed them about their work, and completed desktop research tasks in support of the production of a report on ethics of non-human biobanking. Further, the Frozen Ark are one of twenty-four partner organisations of the Heritage Futures research programme, whose staff have participated in a series of intensive, week long collaborative knowledge exchange events throughout the project which have aimed at understanding the work of each of the partner organisations and co-designing research to address common issues for the sector. The paper also draws on these more experimental ‘para-ethnographic’ (Holmes and Marcus Citation2005, Citation2006, Citation2008; see Harrison et al. Citation2016) engagements with the organisation as part of the research programme over a longer 3-year period.
2. The zoo also owns a patent on Frozen Zoo®.
3. These terms have specific technical definitions which relate to the categories established by the IUCN Red List of Endangered Species.
References
- Adam, B., and C. Groves. 2007. Future Matters: Action, Knowledge, Ethics. Leiden: Brill.
- Bargheer, S. 2016. “Conserving the Future: UNESCO Biosphere Reserves as Laboratories for Sustainable Development.” In Endangerment, Biodiversity, and Culture, edited by F. Vidal and N. Dias, 115–133. Abingdon: Routledge.
- Barnosky, A. D., N. Matzke, S. Tomiya, G. O. U. Wogan, B. Swartz, T. B. Quental, C. Marshall, et al. 2011. “Has the Earth’s Sixth Mass Extinction Already Arrived?” Nature 471: 51–57. doi:10.1038/nature09678.
- Benirschke, K. 1984. “The Frozen Zoo Concept.” Zoo Biology 3: 325–328. doi:10.1002/zoo.1430030405.
- Bennett, T., F. Cameron, N. Dias, B. Dibley, I. Jacknis, R. Harrison, and C. McCarthy. 2017. Collecting, Ordering, Governing: Anthropology, Museums and Liberal Government. Durham: Duke University Press.
- Bird Rose, D. 2011. Wild Dog Dreaming: Love and Extinction. Charlottesville, VA: University of Virginia Press.
- Bird Rose, D., T. van Dooren, and M. Chrulew, eds. 2017. Extinction Studies: Stories of Time, Death, and Generations. New York, NY: Columbia University Press.
- Bowker, G. C. 2000. “Biodiversity Datadiversity.” Social Studies of Science 30: 643–683. doi:10.1177/030631200030005001.
- Bowker, G. C. 2005. Memory Practices in the Sciences. Cambridge, MA: MIT Press.
- Bowker, G. C., and S. L. Star. 2000. Sorting Things Out: Classification and Its Consequences. Cambridge, MA: MIT Press.
- Bowkett, A. E. 2009. “Recent Captive‐Breeding Proposals and the Return of the Ark Concept to Global Species Conservation.” Conservation Biology 23: 773–776. doi:10.1111/j.1523-1739.2008.01157.x.
- Cahill, A. E., M. E. Aiello-Lammens, M. C. Fisher-Reid, X. Hua, C. J. Karanewsky, H. Yeong Ryu, G. C. Sbeglia, et al. 2012. “How Does Climate Change Cause Extinction?” Proceedings of the Royal Society. B. Biological Sciences. doi:10.1098/rspb.2012.1890.
- Caro, T. 2010. Conservation by Proxy: Indicator, Umbrella, Keystone, Flagship, and Other Surrogate Species. Washington, DC: Island Press.
- Carrithers, M., L. J. Bracken, S. Emery, M. Candea, V. Das, C. Degnen, I. Gershon, F. Girke, V. Strang, and M. Strathern. 2011. “Can A Species Be A Person? A Trope and Its Entanglements in the Anthropocene Era.” Current Anthropology 52: 661–685. doi:10.1086/661287.
- CBD, 1992. United Nations Convention on Biological Diversity. Online at https://www.cbd.int/kb/Results?q=1992 Accessed 10 August 2018.
- CBD, 2000. Secretariat of the Convention on Biological Diversity. Sustaining Life on Earth: How the Convention on Biological Diversity Promotes Nature and Human Well-Being. Online at https://www.cbd.int/convention/guide/default.shtml?id=web Accessed 10 August 2018
- Chemnick, L. G., M. L. Houck, and O. A. Ryder. 2009. “Banking of Genetic Resources.” In Conservation Genetics in the Age of Genomics, edited by R. DeSalle, 124–130. New York, NY: Columbia University Press.
- Chrulew, M. 2017. “Freezing the Ark: The Cryopolitics of Endangered Species Preservation.” In Cryopolitics: Frozen Life in a Melting World, edited by J. Radin and E. Kowal, 283–306. Cambridge, MA: MIT Press.
- Clarke, A. G. 2009. “The Frozen Ark Project: The Role of Zoos and Aquariums in Preserving the Genetic Material of Threatened Animals.” International Zoo Yearbook 43: 222–230. doi:10.1111/j.1748-1090.2008.00074.x.
- Clarke, B., J. Murray, and M. S. Johnson. 1984. ““The Extinction of Endemic Species by a Program of Biological Control.” Pacific Science 38 (2): 97–104.
- Cooper, M., and C. Waldby. 2014. Clinical Labor: Tissue Donors and Research Subjects in the Global Bioeconomy. Durham, NC: Duke University Press.
- Cooper, M. E. 2008. Life as Surplus: Biotechnology and Capitalism in the Neoliberal Era. Seattle, WA: University of Washington Press.
- Corley‐Smith, G. E., and B. P. Brandhorst. 1999. “Preservation of Endangered Species and Populations: A Role for Genome Banking, Somatic Cell Cloning, and Androgenesis?” Molecular Reproduction and Development 53: 363–367. doi:10.1002/(SICI)1098-2795(199907)53:3<363::AID-MRD12>3.0.CO;2-0.
- Costa, M., and M. Bruford. 2018. The Frozen Ark Project: Biobanking and Endangered Animal Samples for Conservation and Research. https://insideecology.com/2018/01/12/the-frozen-ark-project-biobanking-endangered-animal-samples-for-conservation-and-research/
- DeSilvey, C., and N. Bartolini. 2018. “Where Horses Run Free? Autonomy, Temporality and Rewilding in the Côa Valley, Portugal.” Transactions of the Institute of British Geographers 1–16. doi:10.1111/tran.12251.
- Dibley, B. 2012. “‘The Shape of Things to Come’: Seven Theses on the Anthropocene and Attachment.” Australian Humanities Review 52: 139–153.
- Dibley, B. 2015. “Anthropocene: The Enigma of ‘The Geomorphic Fold.” In Animals in the Anthropocene: Critical Perspectives on Non-Human Futures, edited by Human Animal Research, 19–32. Sydney: Sydney University Press.
- Doyle, R. 1997. On Beyond Living: Rhetorical Transformations of the Life Sciences. Stanford, CA: Stanford University Press.
- Fletcher, A. L. 2008. “Mendel’s Ark: Conservation Genetics and the Future of Extinction.” Review of Policy Research 25: 598–607. doi:10.1111/j.1541-1338.2008.00367_1.x.
- Fogg, M. J. 1995. Terraforming: Engineering Planetary Environments. Warrendale, PA: SAE International.
- Franklin, S. 2013. Biological Relatives: IVF, Stem Cells, and the Future of Kinship. Durham, NC: Duke University Press.
- Friese, C. 2013. Cloning Wild Life: Zoos, Captivity, and the Future of Endangered Animals. New York, NY: New York University Press.
- Frozen Ark. 2018. The Frozen Ark. Online at www.frozenark.org accessed 10 August 2018.
- Gemeinholzer, B., G. Dröge, H. Zetzsche, G. Haszprunar, H.-P. Klenk, A. Güntsch, W. G. Berendsohn, and J.-W. Wägele. 2011. “The DNA Bank Network: The Start from a German Initiative.” Biopreservation and Biobanking 9: 51–55. doi:10.1089/bio.2010.0029.
- Harrison, R. 2013. Heritage: Critical Approaches. Abingdon: Routledge.
- Harrison, R. 2015. “Beyond Natural and Cultural Heritage: Towards an Ontological Politics of Heritage in the Age of the Anthropocene.” Heritage and Society 8 (1): 24–42. doi:10.1179/2159032X15Z.00000000036.
- Harrison, R. 2016. “World Heritage Listing and the Globalization of the Endangerment Sensibility.” In Endangerment, Biodiversity and Culture, edited by F. Vidal and N. Dias, 195–217. Abingdon: Routledge.
- Harrison, R. 2017. “Freezing Seeds and Making Futures: Endangerment, Hope, Security, and Time in Agrobiodiversity Conservation Practices.” Culture, Agriculture, Food and Environment 39: 80–89. doi:10.1111/cuag.12096.
- Harrison, R., N. Bartolini, C. DeSilvey, C. Holtorf, A. Lyons, S. Macdonald, S. May, J. Morgan, and S. Penrose. 2016. “Heritage Futures.” Archaeology International 19: 68–72. doi:10.5334/ai.1912.
- Heatherington, T. 2012. “From Ecocide to Genetic Rescue: Can Technoscience Save the Wild?” In The Anthropology of Extinction: Essays on Culture and Species Death, edited by G. M. Sodikoff, 39–66. Bloomington, IN: Indiana University Press.
- Heise, U. K. 2016. Imagining Extinction: The Cultural Meanings of Endangered Species. Chicago, IL: University of Chicago Press.
- Helmreich, S. 2008. “Species of Biocapital.” Science as Culture 17: 463–478. doi:10.1080/0950543080251925.6.
- Heyd, D. 2010. “Cultural Diversity and Biodiversity: A Tempting Analogy.” Critical Review of International Social and Political Philosophy 13: 159–179. doi:10.1080/13698230903326315.
- Holdgate, M. 1999. The Green Web: A Union for World Conservation. London: Routledge.
- Holmes, D. R., and G. E. Marcus. 2005. “Cultures of Expertise and the Management of Globalization: Toward the Re-Functioning of Ethnography.” In Global Assemblages: Technology, Politics, and Ethics as Anthropological Problems, edited by A. Ong and S. J. Collier, 235–252. Oxford: Blackwell Publishing.
- Holmes, D. R., and G. E. Marcus. 2006. “Fast Capitalism: Para-Ethnography and the Rise of the Symbolic Analyst.” In Frontiers of Capital. Ethnographic Reflections on the New Economy, edited by M. S. Fisher and G. Downey, 33–57. Durham, NC: Duke University Press.
- Holmes, D. R., and G. E. Marcus. 2008. “Collaboration Today and the Re-Imagination of the Classic Scene of Fieldwork Encounter.” Collaborative Anthropologies 1: 81–101. doi:10.1353/cla.0.0003.
- Hosey, G., V. Malfi, and S. Pankhurst. 2009. Zoo Animals: Behaviour, Management, and Welfare. Oxford: Oxford University Press.
- Howard, J., A. M. Donoghue, M. A. Barone, K. L. Goodrowe, E. S. Blumer, K. Snodgrass, D. Starnes, M. Tucker, M. Bush, and D. E. Wildt. 1992. “Successful Induction of Ovarian Activity and Laparoscopic Intrauterine Artificial Insemination in the Cheetah (Acinonyx Jubatus).” Journal of Zoo and Wildlife Medicine 23 (3): 288–300.
- Howard, J. G., C. Lynch, R. M. Santymire, P. E. Marinari, and D. E. Wildt. 2016. “Recovery of Gene Diversity Using Long-Term Cryopreserved Spermatozoa and Artificial Insemination in the Endangered Black-Footed Ferret: Black-Footed Ferret Gene Restoration.” Animal Conservation 19: 102–111. doi:10.1111/acv.12229.
- Jørgensen, D. 2013. “Reintroduction and De-Extinction.” BioScience 63: 719–720. doi:10.1525/bio.2013.63.9.6.
- Josh, D. C., J. Berger, C. E. Bock, J. H. Bock, A. David, J. A. Burney, D. Foreman, et al. 2006. “Pleistocene Rewilding: An Optimistic Agenda for Twenty‐First Century Conservation.” The American Naturalist 168 (5): 660–681. doi:10.1086/508027.
- Kirchner, J. W., and A. Weil. 2000. “Delayed Biological Recovery from Extinctions Throughout the Fossil Record.” Nature 404: 177. doi:10.1038/35004564.
- Latour, B. 1987. Science in Action: How to Follow Scientists and Engineers through Society. Cambridge, MA: Harvard University Press.
- Latour, B. 1999. Pandora’s Hope: Essays on the Reality of Science Studies. Cambridge, MA: Harvard University Press.
- Lemke, T. 2012. “Second Nature: In the Age of Biobanks.” The Yearbook of Comparative Literature 58: 188–192.
- Lewis, S. L., and M. A. Maslin. 2018. The Human Planet: How We Created the Anthropocene. London: Pelican Books.
- Lorimer, J. 2015. Wildlife in the Anthropocene: Conservation after Nature. Minneapolis, MN: University of Minnesota Press.
- Lowenthal, D. 1985. The Past is a Foreign Country. Cambridge: Cambridge University Press.
- Marris, E. 2013. Rambunctious Garden: Saving Nature in a Post-Wild World. New York, NY: Bloomsbury Publishing.
- Meskell, L. 2012. The Nature of Heritage: The New South Africa. Chichester: Wiley Blackwell.
- National Academies of Sciences, Engineering, and Medicine. 2016. Gene Drives on the Horizon: Advancing Science, Navigating Uncertainty, and Aligning Research with Public Values. Washington, DC: National Academies Press. https://doi.org/10.17226/23405.
- O’Connor, M. R. 2015. Resurrection Science: Conservation, De-Extinction and the Precarious Future of Wild Things. New York, NY: St. Martin’s Press.
- Oberg, J. E. 1981. New Earths: Restructuring Earth and Other Planets. Harrisburg, PA: Stackpole Books.
- Parry, B. 2004. Trading the Genome: Investigating the Commodification of Bio-Information. New York, NY: Columbia University Press.
- Perrings, C., C. Folke, and K.-G. Mäler. 1992. “The Ecology and Economics of Biodiversity Loss: The Research Agenda.” Ambio 21 (3): 201–211.
- Pilcher, H. 2016. Bring Back the King: The New Science of De-Extinction. London: Bloomsbury Press.
- Pratt, M.-L. 2017. “Coda: Concept and Chronotope.” In Arts of Living on a Damaged Planet, edited by A. L. Tsing, N. Bubandt, E. Gan, and H. A. Swanson, 169–174. Minneapolis, MN: University of Minnesota Press.
- Radin, J. 2013. “Latent Life: Concepts and Practices of Human Tissue Preservation in the International Biological Program.” Social Studies of Science 43: 484–508. doi:10.1177/0306312713476131.
- Radin, J. 2015. “Planned Hindsight: The Vital Valuations of Frozen Tissue at the Zoo and the Natural History Museum.” Journal of Cultural Economy 8: 361–378. doi:10.1080/17530350.2015.1039458.
- Radin, J. 2017. Life on Ice: A History of New Uses for Cold Blood. Chicago, IL: University of Chicago Press.
- Radin, J., and E. Kowal. 2017. Cryopolitics: Frozen Life in a Melting World. Cambridge, MA: MIT Press.
- Rajan, K. S. 2006. Biocapital: The Constitution of Postgenomic Life. Durham, NC: Duke University Press.
- Rico, T. 2015. “Heritage at Risk: The Authority and Autonomy of a Dominant Preservation Framework.” In Heritage Keywords: Rhetoric and Redescription in Cultural Heritage, edited by K. Lafrenz Samuels and T. Rico, 147–162. Boulder, CO: University Press of Colorado.
- Rico, T. 2016. Constructing Destruction: Heritage Narratives in the Tsunami City. Abingdon: Routledge.
- Rose, N. 2006. The Politics of Life Itself: Biomedicine, Power, and Subjectivity in the Twenty-First Century. Princeton, NJ: Princeton University Press.
- Sagan, C. 1961. “The Planet Venus.” Science 133: 849–858. doi:10.1126/science.133.3456.849.
- Sagan, C. 1973. “Planetary Engineering on Mars.” Icarus 20 (4): 513–514. doi:10.1016/0019-1035(73)90026-2.
- Sarkar, S. 1999. “Defining ‘Biodiversity’; Assessing Biodiversity.” The Monist 85: 131–155. doi:10.5840/monist20028515.
- Sepkoski, D. 2016. “Extinction, Diversity, and Endangerment.” In Endangerment, Biodiversity and Culture, edited by F. Vidal N. and N. Dias, 62–86. Abingdon: Routledge.
- Shapiro, B. 2015. How to Clone a Mammoth: The Science of De-Extinction. Princeton, NJ: Princeton University Press.
- Sherkow, J. S., and H. T. Greely. 2013. “What If Extinction Is Not Forever?” Science 340: 32–33. doi:10.1126/science.1236965.
- Shukin, N. 2009. Animal Capital: Rendering Life in Biopolitical Times. Minneapolis, MN: Univeristy of Minnesota Press.
- Singh, J. S. 2002. “The Biodiversity Crisis: A Multifaceted Review.” Current Science 82: 627–638.
- Sodikoff, G. M., ed. 2012. The Anthropology of Extinction: Essays on Culture and Species Death. Bloomington, IN: Indiana University Press.
- Swingland, I. R. 2001. “Biodiversity, Definition Of.” Encyclopedia of Biodiversity 1: 377–391.
- Thacker, E. 2006. The Global Genome: Biotechnology, Politics, and Culture. Cambridge, MA: MIT Press.
- Thacker, E. 2010. After Life. Chicago, IL: University of Chicago Press.
- Tsing, A. L., N. Bubandt, E. Gan, and H. A. Swanson. 2017. Arts of Living on a Damaged Planet: Ghosts and Monsters of the Anthropocene. Minneapolis, MN: University of Minnesota Press.
- Turner, S. S. 2007. “Open-Ended Stories: Extinction Narratives in Genome Time.” Literature and Medicine 26: 55–82. doi:10.1353/lm.2008.0007.
- Van Dooren, T. 2009. “Banking Seed: Use and Value in the Conservation of Agricultural Diversity.” Science as Culture 18: 373–395. doi:10.1080/09505430902873975.
- Van Dooren, T. 2014. Flight Ways: Life and Loss at the Edge of Extinction. New York, NY: Columbia University Press.
- Vidal, F., and N. Dias. 2016. “Introduction: The Endangerment Sensibility.” In Endangerment, Biodiversity and Culture, edited by F. Vidal and N. Dias, 1–38. Abingdon: Routledge.
- Waldby, C. and R. Mitchell. 2006. Tissue Economies: Blood, Organs, And Cell Lines In Late Capitalism. Durham, NC: Duke University Press.
- Watson, P., and W. V. Holt. 2001. Cryobanking the Genetic Resource: Wildlife Conservation for the Future? London: Taylor and Francis.
- Westley, F. 1997. “‘Not on Our Watch’: The Biodiversity Crisis and Global Collaboration Response.” Organization & Environment 10: 342–360. doi:10.1177/192181069701000402.
- Wildt, D. E., W. F. Rall, J. K. Critser, S. L. Monfort, and U. S. Seal. 1997. “Genome Resource Banks.” BioScience 47: 689–698. doi:10.2307/1313209.
- Wilson, A. 1992. The Culture of Nature. Cambridge, MA: Blackwell Publishing.
- Wilson, E. O., ed. 1988. Biodiversity. Washington, DC: National Academy Press.
- Wisely, S. M., O. A. Ryder, R. M. Santymire, J. F. Engelhardt, and B. J. Novak. 2015. “A Road Map for 21st Century Genetic Restoration: Gene Pool Enrichment of the Black-Footed Ferret.” Journal of Heredity 106: 581–592. doi:10.1093/jhered/esv041.
- WWF. 2016. “Living Planet Report: Risk and Resilience in a New Era.” http://wwf.panda.org/about_our_earth/all_publications/lpr_2016/.