Abstract
Claims have been made for associated degrees of impairment on both visual and auditory performance in unilateral neglect and extinction. Since this evidence is primarily based on different tests in each modality, it is difficult to properly quantify the degree of association between performance in vision and audition. The current study compares visual and auditory extinction and temporal order judgments (TOJs) in two cases with clinical visual neglect. Stimuli in both modalities were precisely matched in their temporal and spatial parameters. The results reveal a mixed pattern of association between different auditory tests and their visual counterparts. This suggests that associations between visual and auditory neglect can occur but these are neither obligatory nor pervasive. Instead, our data support models of spatial impairment in neglect and extinction that acknowledge differences in the contribution of spatial information to performance in each modality in responses to changing task demands.
INTRODUCTION
Unilateral neglect is a neurological syndrome associated with unilateral (predominantly right) cortical lesions. It affects up to two thirds of acute right hemisphere stroke patients (CitationParton, Malhotra, & Husain, 2004) and presents as a lack of awareness of objects and events on the side of space contralateral to the causal lesion (i.e., on the left side for a patient with right hemisphere damage). These symptoms exist despite the absence of basic sensory deficits (e.g., hemianopia) and are often attributed to an impairment of spatial representation and/or selective attention (e.g., E. CitationBisiach, 1993; CitationDi Pellegrino, Basso, & Frassinetti, 1997; CitationFarah, Wong, Monheit, & Morrow, 1989; CitationMesulam, 1999; CitationRorden, Mattingley, Karnath, & Driver, 1997). In some cases, contralesional neglect only occurs in the presence of a simultaneous ipsilesional object. This pattern of impairment, often called extinction, is usually considered part of the neglect syndrome (CitationParton et al., 2004). Despite the fact that neglect and extinction are most commonly described and diagnosed on the basis of visuospatial behavior, there is growing evidence that these deficits can occur in other sensory modalities (e.g., CitationCusack et al., 2000; CitationEramudugolla, Irvine, & Mattingley, 2007; CitationPavani, Husain, Ladavas, & Driver, 2004; CitationPavani, Ladavas, & Driver, 2003; CitationSpierer, Meuli, & Clarke, 2007). Associations between visual, auditory, and tactile neglect suggest that the syndrome reflects a perturbation of multimodal spatial selection and attention (E. CitationBisiach et al., 2004; L. CitationBisiach, Cornacchia, & Vallar, 1984; CitationBueti, Costantini, Forster, & Aglioti, 2004).
Models ascribing visuospatial deficits in neglect and extinction to changes in the distribution of neural activation representing stimuli presented in ipsilesional and contralesional space provide a theoretical account for spatiotemporal deficits in other modalities (e.g., CitationCate & Behrmann, 2002; CitationKinsbourne, 1993; CitationRizzolati, Scandolara, Matelli, & Gentilucci, 1981). This has led researchers to extend models of visuospatial neglect and extinction to explain deficits in audition and touch (e.g., CitationPavani et al., 2004; CitationPavani et al., 2003). This generalization is given further support by behavioral (e.g., CitationDriver & Spence, 1994; CitationSpence & Driver, 1997; CitationSpence, Lloyd, McGlone, Nicholls, & Driver, 2000), neuroanatomical (see CitationGhazanfar & Schroeder, 2006, for a recent review), and neuroimaging (CitationLewis, Beauchamp, & DeYoe, 2000; CitationMacaluso, Frith, & Driver, 2001) evidence and further indicates that spatial location and attention play an important part in multimodal integration.
Despite the above, multimodal accounts of neglect and extinction remain somewhat contentious (CitationGolay, Hauert, Greber, Schnider, & Ptak, 2005; CitationSinnett, Juncadella, Rafal, Azanon, & Soto‐Faraco, 2007; CitationSpierer et al., 2007). A number of studies have found dissociations between visual and auditory attentional biases in patients diagnosed with visual neglect (CitationDe Renzi, Gentilini, & Barbieri, 1989; CitationSoroker, Calamaro, Glicksohn, & Myslobodsky, 1997; CitationSpierer et al., 2007; CitationZimmer, Lewald, & Karnath, 2003). These have been interpreted as evidence for selective damage to modality-specific mechanisms of spatial representation (e.g., CitationDe Renzi, Gentilini, & Pattachini, 1984; CitationSoroker et al., 1997). This assertion emphasizes potential differences in the role of spatial selection in perceptual systems characterized by different coding principles (e.g., the spatiotopic visual and tonotopic auditory systems) and is consistent with evidence for modality-specific spatial cuing effects in vision and audition (CitationMcDonald & Ward, 1999; CitationRoberts, Summerfield, & Hall, 2006). An alternative explanation is that the reported dissociations between measures of attentional bias may, in part, reflect differences in the specificity and sensitivity of the visual and auditory tests employed rather than the nature of the underlying impairment itself. Given the potential remedial utility afforded by intact auditory-spatial representation in patients with visual neglect (CitationGolay et al., 2005), it is important to establish the degree of association between visual and auditory impairment when potential measurement artifacts are controlled.
Much of the evidence for an association between visual and auditory attentional bias is based on tests that place different demands on performance in each modality (e.g., CitationPavani et al., 2004; CitationPavani, Meneghello, & Ladavas, 2001; CitationPavani et al., 2003). Neglect assessment typically utilizes clinical “pen and paper” tests that may include line bisection, cancellation, and copying from sample tasks (e.g., CitationWilson, Cockburn, & Halligan, 1987). These are presented at the patient's bedside, with the patient being given unlimited time to view the material and to make a manual response. Performance on these tasks draws heavily on visuomotor control (CitationParton et al., 2004). In contrast, auditory neglect has typically been assessed using tasks in which the patient is asked to localize briefly presented sounds and to make either a verbal or a simple manual response that is orthogonal to left and right (e.g., CitationPavani et al., 2003; CitationPinek, Duhamel, Cave, & Brouchon, 1989).
Differences in task demands have also been confounded by differences in the location to which visual and auditory stimuli are presented. Most auditory tasks have used sounds presented over headphones (CitationBellmann, Meuli, & Clarke, 2001; CitationKarnath, Zimmer, & Lewald, 2002; CitationZimmer et al., 2003). These manipulate perceived location using binaural cues (interaural timing differences, ITDs, and interaural level differences, ILDs; e.g., L. CitationBisiach et al., 1984; CitationZimmer et al., 2003) or monaural cues. A sound presented to only one ear has an infinite ILD (e.g., CitationSinnett et al., 2007). This produces sounds that appear to arise from an intracranial position, a location for which there is no visual counterpart (CitationBlauert & Lindemann, 1986). Binaural cues also elicit a weak spatial percept to which even normal listeners, without training, can have difficulty orienting attention (CitationRoberts et al., 2006). The lack of a spatial coincidence between auditory stimuli presented over headphones and visual stimuli presented in extrapersonal space means that stimuli in either modality arise from different locations. Differences between binaural and monaural sounds can also make it difficult to distinguish between the involvement of spatial representation on one hand and the perceptive extinction of information from one ear on the other. Extinction for monaural sounds presented to the left ear and for binaural sounds lateralized to the left hemispace have also been shown to dissociate, suggesting distinct underlying neural processes in each case (CitationBellman et al., 2001; CitationSpierer et al., 2007).
An alternative method of sound presentation is to use head-related transfer functions. These generate virtual space by simulating the linear distortions in the magnitude and phase spectrum of the acoustic signal on its path from the sound source to the eardrums, which are caused by shadowing, reflection, and diffraction by the head and outer ear. These filter characteristics are known to disambiguate front–back confusions and improve localization accuracy (CitationWenzel, Arruda, Kistler, & Wightman, 1993). Although this method improves the perception of auditory spatial location for sounds presented over headphones, it is still not ideal as the sound source is rarely perceived as external unless personalized transfer functions are used (CitationWenzel et al., 1993; CitationWightman & Kistler, 1989). Even when these or free-field sounds have been used to assess auditory neglect in virtual or real space, comparative visual stimuli have either been absent (CitationBender & Diamond, 1965; CitationPavani, Lavadas, & Driver, 2002; CitationSoroker et al., 1997) or not matched for egocentric location. In CitationEramudugolla et al.'s (2007) study, for example, visual stimuli were presented at eccentricities of ±8.7˚ while auditory stimuli were presented at ±90˚ from fixation. This lack of explicit matching for location across visual and auditory tests has implications for the assessment of any multimodal spatial metric in neglect and makes quantitative comparisons across sensory modalities difficult (but see CitationFrassinetti et al., 2002, for a recent exception).
The current paper presents two case studies that examine the degree of correspondence between visual and auditory neglect and extinction using tasks carefully designed to match the extrapersonal spatial and temporal parameters of stimuli in each modality. Unlike group studies, the case-by-case approach allows direct comparisons of unimodal performance that are independent of variability introduced by differences in the site and extent of lesions among patients in the sample (CitationCaramazza & Badecker, 1989). Dissociations within a single case provide evidence for the absolute or relative separation of neural processes underlying individual performance under different test conditions (see CitationShallice, 1988, for a review of the logic underlying single-case methodology). In the first task, we report extinction indices that measure decrements in the detection of dual compared to single objects as a function of their egocentric location. To our knowledge, this is the first time that detection rates have been compared for visual and auditory stimuli presented at identical locations in the horizontal meridian (azimuth). In the second task, sensitivity to temporal asynchrony between two stimuli is measured using a temporal order judgment (TOJ) task. Previous TOJ results have revealed a temporal disadvantage for contralesional compared to ipsilesional stimuli (CitationDi Pellegrino et al., 1997; CitationEramudugolla et al., 2007; CitationKarnath et al., 2002; CitationRorden et al., 1997). Our experiment extends these studies by evaluating spatiotemporal judgments using visual and auditory stimuli that are matched precisely in terms of their spatial and temporal characteristics.
METHOD
Participants
Two patients (F.D. & J.B.) were recruited from the stroke rehabilitation ward at the Queen's Medical Centre, Nottingham. F.D. suffered a right middle cerebral artery (MCA) territory infarct resulting in low attenuation in the region of the superior right frontal lobe, caudate, and lentiform nucleus. J.B. had a hyperacute right MCA territory infarction with hyperdense right MCA branches. Hypodensity was also observed in the right insula and external capsules (radiographer's reports). Patients were assessed within one month of hospital admission (14 and 31 days) and undertook a battery of clinical tests, including a star cancellation task, line bisection, and figure drawing from sample. Each demonstrated a pattern of visuospatial performance consistent with a clinical diagnosis of neglect (see for a clinical description for each patient). Both patients had intact hemifields, and pure-tone audiometry revealed hearing within the range of age-matched controls (CitationDavis, 1995).
TABLE 1 Patient information and clinical characteristics
A total of 15 age-matched controls (mean = 66 years, SD = 2.5 years) were also recruited from the Nottingham area. Controls had no history of neurological impairment and showed no symptoms of neglect assessed using the same battery of tests as that administered to patients. All controls had intact hemifields and hearing performance within the normal range for their age (CitationDavis, 1995).
Stimuli and apparatus
Visual stimuli consisted of two geometric shapes: a red circle and a blue square. Each was closed and subtended 8.3˚ of visual angle. Auditory stimuli were two digitally created vowel sounds: “ah” and “ee.” Vowel pairs were separated in their fundamental frequency by four semitones so that they could be easily segregated from one another (CitationCulling, Summerfield, & Marshall, 1994). Vowels were presented at 69 dB(A) on single-target trials and at 66 dB(A) on dual-target trials to reduce the sound level cues that would otherwise distinguish single- from dual-target trials. Before testing, each participant was familiarized with the visual and auditory stimuli to ensure they could accurately identify and discriminate them.
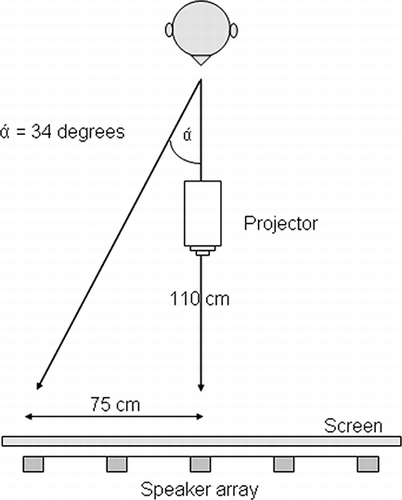
The apparatus (see ) consisted of five small Bose Cube loudspeakers arranged horizontally at ear level. Loudspeakers were mounted on a flat metal frame (height 150 cm, width 150 cm) supported off the floor by two metal legs. Each loudspeaker was located at an eccentricity of 0˚, 17˚, and 34˚ to either side of the fixation point, providing one position on the vertical midline, two in the left hemispace, and two in the right. A sheet of white fabric was stretched across the metal frame to cover the loudspeakers and to prevent any visual cues about their position. Participants sat in front of this screen at a viewing distance of 1,100 mm. Visual stimuli were projected from a ceiling-mounted projector (NEC WT610) onto the screen at precisely the same positions as the loudspeakers. Visual targets were the same size as the loudspeakers, and visual and auditory stimuli were presented at exactly the same location in extrapersonal space. A fixation cross was projected immediately below the horizontal midline and remained on the screen throughout each trial. Stimulus presentation was controlled using customized software running on a Viglen PC equipped with a standard graphics card and a Motu 24 I/O sound card. Verbal responses were coded and entered into the computer by the experimenter.
Figure 1. Schematic of the experimental apparatus. A projector, screen, and speaker array were used to present visual and auditory stimuli to the same egocentric position. The display had five locations equally distributed in the horizontal plane and centered on the participant's vertical midline. Visual and auditory stimuli were presented in separate blocks.
Procedure
Ethical approval for the study was granted by the Nottingham committee of the National Health Research Ethics Board, and written consent was obtained from all participants. Testing comprised the clinical assessment and the two experimental tasks. Clinical assessments were carried out on the ward while experimental tasks were conducted in a sound-attenuated booth at the Ear, Nose, and Throat department. For the patients, assessment and experimental tasks were carried out on consecutive days with cancellation and visual confrontation tasks repeated on the final day of testing. Controls completed all of their testing within a single session in the sound-attenuated booth.
Visual and auditory versions of the extinction and TOJ tasks were presented in separate blocks, with the order counterbalanced across participants. Once the participant was ready, trials were initiated by the experimenter and always started with fixation.
In the extinction task, stimuli were presented singly or in a pair for one second. Single-target trials comprised an equal number of each stimulus (e.g., “ah” and “ee”). Dual-target trials always contained one of each stimulus (e.g., a blue square and a red circle) and were always presented to locations separated by an angle of 34˚ (i.e., with one loudspeaker position between them). Single targets were presented to each of the five horizontal locations. Dual targets were presented to three spatial configurations in which the pair was (a) located in the right hemispace (RH), (b) across the central meridian (CM) or (c) located in the left hemispace (LH; ). Stimulus type, location, and number of single- and dual-target trials (12 within each spatial configuration) were sampled with equal probability in each task. Trials were presented in a random order. Participants were asked to report the number of targets (one or two), but were not required to identify them. Responses were, therefore, orthogonal to the location of the stimuli in order to negate any confounds associated with a spatially mediated response bias.
Figure 2. The left-hand panel illustrates the dual-target conditions in the extinction task. Dual targets were presented to the left hemispace (LH), to the right hemispace (RH), and across the central meridian (CM). Single targets were presented to each of the five locations. The visual angles correspond to the five locations at which stimuli were presented. The right-hand panel presents a schematic of the temporal order judgment (TOJ) task. Two stimuli were always presented at a fixed eccentricity of ±17˚. Stimulus onset asynchronies (SOAs) varied between 0 and 400 ms.
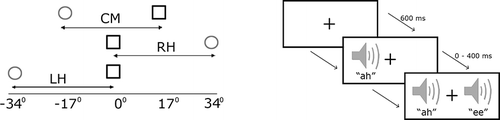
In the TOJ task, the pair of targets (e.g., blue square and red circle) was always presented at an eccentricity of 17˚ in the left and right hemifields (i.e., separated by a fixed angle of 34˚). The stimulus onset asynchrony (SOA) between the two targets could take the following values: 1, 50, 100, 150, 200, 250, 300, and 400 ms. The side and order in which each stimulus type was presented were counterbalanced. Six right- and six left-first target trials occurred at each location for all SOAs. This method of constant stimuli provided a spread of data that was sufficient to plot individual psychometric functions. The offset of the pair of targets was simultaneous, and the first stimulus was always presented for a total of one second. Both the SOA and the side on which the target first appeared were randomized. Participants indicated verbally which of the two targets appeared first by reporting its identity. Again, participants were required to report a stimulus attribute (identity) that was orthogonal to the location to which stimuli were presented (CitationShore & Spence, 2005).
Analyses: Extinction task
To assess the effect of eccentricity, the proportion of correctly detected single-target trials was entered into contingency tables for each participant. Trial outcome (0 = incorrect, 1 = correct) was entered as the dependent variable and eccentricity (−34˚, −17˚, 0˚, 17˚, 34˚) as the independent variable. Due to the binary nature of the response, the data were analyzed using Somers' d. This is a test of association between the outcome (frequency of correct responses) and the ordinal independent variable (eccentricity; see CitationSiegel & Castellan, 1988).
To evaluate the ability to detect dual targets compared to single targets, the data were transformed into an extinction index derived from that of CitationDuncan et al. (1999). An index was calculated separately for LH, CM, and RH configurations by dividing the proportion of correct responses on dual-target trials (PD) by the product of the proportion of correct responses for the single trials (PS) at the corresponding two locations: PD/(PSleft × PSright); see . An index value equal to unity represents equivalent performance on both single- and dual-target trials, whilst an index value of less than unity reflects relatively worse performance on dual-target trials. To measure the degree of association between each patient's visual and auditory extinction indices, stimulus modality only was permuted across all locations. Random permutations were repeated 1,000,000 times in order to estimate an empirical distribution for any differences in the index under the null hypothesis. Reported p-values estimate the two-tailed probability that there is no difference in the distribution of visual and auditory indices over the three spatial configurations. Randomization tests were also used to compare the patients' extinction indices with the observed indices for the control group. In this comparison the p-value reflects the two-tailed probability that the patient's performance is consistent with that of the controls (see ).
TABLE 2 Proportion of trials correctly identified as single- and dual-target displays and extinction indices
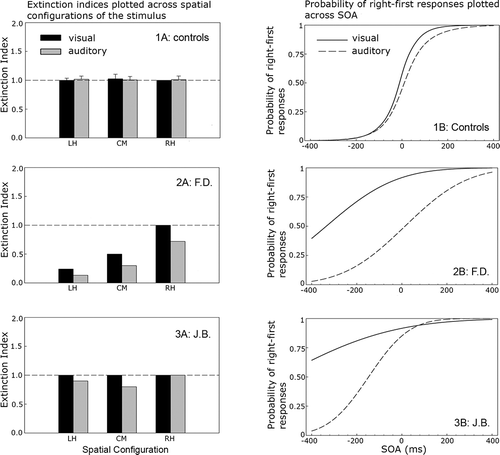
Figure 3. Left-hand column: mean extinction indices for the control group (Panel 1A, mean; error bars represent the 95% confidence interval, CI, around the mean) and individual indices for each patient (Panels 2A and 3A), by stimulus location and modality. Right-hand column: mean probit functions for the control group (Panel 1B, mean) and individual functions for patients (Panels 2B and 3B), by stimulus onset asynchrony (SOA) and modality.
Analyses: TOJ
The number of correctly named targets at each SOA was recoded to represent the proportion of “right-first” responses. These data were then analyzed using a probit analysis (CitationFinney, 1964), which produces a probability estimate for each participant of right-first responses as a linear function of SOA, transformed via a cumulative standard normal distribution (CitationMcCullagh & Nelder, 1983). The resulting probability functions were then used to estimate each participant's point of subjective simultaneity (PSS): the point at which right-first and left-first decisions are equally probable (i.e., .5). PSS values deviating from zero reflect slowed responses for the ipsilesional (PSS > 0) or contralesional stimulus (PSS < 0). Goodness-of-fit estimates (GoF) for the observed and estimated data for each individual were calculated using the deviance statistic (p-values < .05 suggest that the psychometric function is a poor fit). For one case (J.B. auditory test), a single incorrect response at SOA of 300 ms was omitted as an outlier on the basis that it was grossly inconsistent with performance at adjacent SOAs and reduced the GoF. The deviance statistic and the 95% confidence intervals around the PSS were calculated using Matlab's “glmfit” routine (Version 7.5.0; MathWorks Inc., 2007; see CitationCollett, 2002, for a description). For each individual, the sharpness of temporal acuity was also estimated using the just noticeable difference (JND). This is calculated as half of the SOA between the point on the probit function at which 25% and 75% of targets were reported as right-first trials (CitationShore & Spence, 2005). reports the statistics associated with the individual probit analyses for each patient.
TABLE 3 TOJ statistics by modality for the control group and individual patients
RESULTS
Controls
Controls performed near ceiling on both extinction tasks, and there was no significant effect of eccentricity (α = .05). For visual stimuli, mean target accuracy was 99% (SD = 0.9%) for single targets and 99% (SD = 0.6%) for dual targets. For auditory stimuli, mean accuracy was 99% (SD = 1.0%) for single targets and 99% (SD = 0.3%) for dual targets (see ). Mean extinction indices for stimuli at the LH, CM, and RH spatial configurations approached unity in the visual and auditory tasks (see , Panel 1A), demonstrating consistent performance between single- and dual-target trials.
Results for the TOJ task also demonstrated near veridical judgments of temporal asynchrony. Panel 1B in plots the mean probit function as a function of SOA for the visual and auditory stimuli. For comparison with the patients, the mean PSS values for the control group along with the 95% confidence intervals (CIs) are presented in . In the visual modality, the mean PSS was −13 ms (SD = 26 ms), and in the auditory modality it was 15 ms (SD = 44 ms). Neither PSS differed significantly from zero, t(14) = −1.97, p = .07, and t(14) = 1.33, p = .20, respectively. The steep slope of the probit function indicates good temporal acuity with a visual JND of 50 ms and an auditory JND of 59 ms.
Case: F.D
shows that F.D.'s single-target performance in the extinction task was close to ceiling at all three spatial configurations (mean = 97%). No effect of eccentricity upon performance was observed in either the visual or the auditory modality (p > .1). In contrast, extinction indices of < 1 revealed a significant reduction in accuracy on dual- compared to single-target trials on visual and auditory trials (, Panel 3A). This pattern is consistent with extinction in dual-target trials. With the exception of dual visual targets presented to the RH, F.D.'s performance was significantly worse than that of the controls at all locations (p < .001), decreasing monotonically as the stimuli moved from the RH to the LH spatial configurations. Statistical comparisons of visual and auditory extinction indices at each location revealed no difference (p = .15) indicating comparable rates of extinction in both modalities.
In the TOJ task, F.D. showed a general reduction of temporal acuity ( Panel 3B). Goodness-of-fit for the observed data and estimated function were acceptable (see ). The estimated mean fell outside the 95% CI around the control mean. F.D.'s visual PSS revealed a large bias away from the contralesional target. Visual targets presented on the left had to precede those on the right by an estimated 335 ms before their onsets were judged to be simultaneous. In contrast there was no evidence for an auditory spatiotemporal bias, with the auditory PSS close to that of the controls (14 ms). F.D.'s data, therefore, reveal a multimodal impairment in detecting simultaneous visual or auditory stimuli in the extinction task and a dissociation between visual and auditory judgments of temporal order. Importantly, this task by modality interaction occurs for stimuli presented to identical egocentric locations (i.e., ± 17˚).
Case: J.B
J.B.'s detection accuracy for single targets in the extinction task was 100% (see ). In the visual modality, extinction indices of unity demonstrated equally good performance for single- and dual-target trials at all locations (, Panel 4A). Auditory extinction indices for stimulus pairs presented to LH and CM revealed a slight decrement in dual- compared to single-target detection rates at these locations. Although responses to centrally presented auditory targets were reliably less accurate than those of controls (p < .05), visual and auditory performance was statistically consistent at this location (p > .2). This patient, therefore, demonstrates no reliable difference in performance across the two modalities.
In the TOJ task, JND estimates in the visual (266 ms) and auditory (96 ms) tasks showed impoverished temporal acuity compared to controls (, Panel 4B). The probit function on the visual task was particularly shallow and had to be extrapolated beyond the SOA range in order to estimate the PSS (−544 ms). In the auditory task, the deviation of the PSS from zero was much smaller (−148 ms), but in the same direction (95% CIs around the visual and auditory PSS did not overlap revealing a reliable reduction in auditory compared to visual spatiotemporal bias; see ). Despite this, both visual and auditory PSS values fall outside the 95% CI around the control mean, revealing a multimodal spatiotemporal bias towards the ipsilesional stimulus. J.B's data, therefore, reveal a multimodal impairment of temporal order judgments for asynchronous visual and auditory stimuli in the absence of multimodal extinction for the same objects when these are simultaneously presented.
DISCUSSION
Research investigating the degree of correspondence between visual and auditory attentional bias has produced mixed findings (e.g., CitationDe Renzi et al., 1989; CitationPavani et al., 2004; CitationPavani et al., 2003; CitationSinnett et al., 2007; CitationSoroker et al., 1997; CitationZimmer et al., 2003). Inconsistencies in estimates of the degree of association between visual and auditory neglect and extinction may in part be due to methodological factors: first, the use of tests that each afford different measurement sensitivity in each sensory modality, and, second, the grouping of data across individuals who vary in the site and extent of their causal lesion. The current study addresses these methodological considerations by comparing visual and auditory neglect symptoms separately for two patients using tests designed explicitly to match spatial and temporal stimulus parameters in each modality. Despite a common clinical diagnosis of visual neglect using pen and paper tests, both patients showed a unique pattern of impairment across two laboratory tasks commonly used to assess unimodal neglect and extinction. Our results reveal within-patient variability in the presence and degree of association between visual and auditory inattention as well as highlighting task-dependent variability in the way these are expressed.
In the extinction task, F.D. showed reduced awareness of contralesional visual and auditory stimuli when a competing stimulus was present. This bias increased across a leftward spatial gradient that is consistent with gradient models of neglect and extinction (CitationCate & Behrmann, 2002; CitationKinsbourne, 1993; CitationPouget & Driver, 2000). This suggests the balance of competition between stimuli with a fixed angle of separation is mediated by both their relative and egocentric locations. The lack of any significant difference between extinction indices in each modality suggests that performance in this patient was affected by a common gradient of inattention. This is consistent with a multimodal attentional impairment for visual and auditory information represented within a common spatial metric (e.g., CitationFrassinetti et al., 2002; CitationPavani et al., 2002, Citation2003). F.D.'s TOJ data, in contrast, reveal intact auditory performance in the presence of a visual deficit. In the auditory TOJ, F.D. performed within normal limits, although the shallow function indicated a general loss of temporal acuity. In the visual TOJ, F.D.'s data revealed a large spatiotemporal bias against the contralesional object. This is consistent with prior entry models of neglect and extinction, which attribute spatiotemporal deficits to a disruption in the competitive interactions between ipsi- and contralesional stimuli (CitationDi Pellegrino et al., 1997; CitationKarnath et al., 2002; CitationRorden et al., 1997). F.D.'s data, therefore, reveal an association between visual and auditory deficits on one task (extinction) and a clear dissociation between visual and auditory deficits on another (TOJ). Data for J.B. also reveal a dissociation between the extinction and TOJ tasks. In this patient, intact performance on the visual and auditory extinction tasks contrasts with a multimodal deficit in the TOJ for the same objects presented to the same location.
The pattern of dissociations above caution against the assumption that the same test presented in different modalities will tap identical attentional resources. Previous studies comparing associations between visual and auditory attentional bias using extinction and TOJ tasks have found conflicting results (e.g., CitationEramudugolla et al., 2007; CitationSinnett et al., 2007) and highlight the variability typically observed in comparisons of visual and auditory inattention using different measures. The use of visual and auditory stimuli precisely matched in their spatial and temporal parameters in this study suggests that this variation is attributable to the way individual patterns of impairment disrupt spatial and temporal selection in particular tasks rather than to confounds associated with the presentation of stimuli in each modality to different (intracranial and extrapersonal) locations (e.g., CitationSinnett et al., 2007). The strength of association between visual and auditory deficits in our patients depends upon whether the presentation of stimuli is simultaneous or asynchronous. Recent evidence from transcranial magnetic stimulation studies has revealed modality-specific substrates of visual and auditory time perception (Bueti, Bahrami, & Walsh, 2008). Spatial and temporal deficits in neglect and extinction often coincide (CitationBecchio & Berone, 2006; CitationDanckert et al., 2007), and it is likely that associations between visual and auditory deficits will reflect the contribution of these to task performance in each modality. In audition, temporal asynchronies have been shown to be a powerful determinant of scene segregation. Common onsets provide a cue for grouping while asynchronous onsets promote stimulus segregation (e.g., CitationHukin & Darwin, 1995). This temporal segregation is thought to occur prior to localization (CitationHill & Darwin, 1996; CitationWoods & Colburn, 1992) and would have provided a nonspatial mechanism for segregating sounds in the TOJ but not the extinction task. Asking respondents to report the identity rather than the position of the first target in the TOJ task might have accentuated this nonspatial effect, distinguishing our data from evidence of an ipsilesional bias in TOJ for auditory targets identified by their spatial location (CitationEramudugolla et al., 2007; CitationKarnath et al., 2002). Task-dependent variability in the effect of auditory spatial cues has previously been reported by CitationMcDonald and Ward (1999) and has been interpreted as evidence for mechanisms of auditory attention that operate independently of spatial location. This can be contrasted with the obligatory role of spatial information in delimiting object boundaries in the visual system (CitationBaylis & Driver, 1993; CitationKim & Cave, 2001; CitationKramer, Weber, & Watson, 1997; CitationLamy & Tsal, 2000) and provides a potential account for the (relative and absolute) dissociation of visual and auditory TOJs in our patients. An important implication of this account is that the assumption of equivalence that has informed much of the research investigating visual and auditory attentional biases inadequately describes modality-specific variation in the contribution of spatial information to performance on measures of neglect and extinction.
In conclusion, the current results suggest that associations between visual and auditory attentional deficits can occur but these are neither obligatory nor pervasive. Associations varied within patients and across tasks and were apparent despite the use of visual and auditory stimuli that were precisely matched in their spatial and temporal parameters. An important implication of this result is that measures of association between visual and auditory inattention will vary according to the type of task involved and the extent that performance in each modality is mediated by spatial information. This argues against the characterization of attention as either a unimodal or a supramodal resource and instead supports a more complex model of task-specific impairment that may vary across modalities in neglect. Research has revealed both unimodal and multimodal responses to spatial information across a distributed network of cortical areas (CitationBushara, Weeks, Ishii, Catalan, & Tian, 1999; CitationLewis et al., 2000; CitationMacaluso, Frith, & Driver, 2000), with neurons responding to spatially convergent input from different modalities found in the superior temporal and intraparietal sulci as well as the prefrontal cortices (CitationBeauchamp, 2005; CitationGifford & Cohen, 2004; CitationMaier, Neuhoff, Logothetis, & Ghazanfar, 2004). Neglect symptoms can arise from damage to many different cortical and subcortical regions (CitationCoulthard, Parton, & Husain, 2007; CitationHalligan, Fink, Marshall, & Vallar, 2003; CitationVallar, 2001), and it is likely that individual patterns of impairment will reflect the contribution of these to different levels of spatial description (CitationGhazanfar & Schroeder, 2006; CitationKarnath, 2001; CitationMarshall, Halligan, & Robertson, 1993; CitationMesulam, 1981). Accordingly the type of deficit observed when this distributed system is compromised will depend on the site and extent of the causal lesion as well as the nature of spatial representation required by the task. By using matched tasks to explore the manifestation of these deficits across different sensory modalities, confounds associated with different measures of performance can be reduced. This approach supplements evidence from existing investigations of neglect and extinction in order to specify more precisely the way spatial selection operates across different sensory modalities and tasks.
This work was supported by a Medical Research Council Career Development Grant awarded to the first author and formed part of an ongoing program of research at the Medical Research Council's Institute of Hearing Research, Nottingham, U.K. We would like to thank the consultants and staff at the Queen's Medical Centre, Nottingham, who provided support as well as to the patients themselves for their participation.
Related Research Data
REFERENCES
- Baylis , G. C. and Driver , J. 1993 . Visual attention and objects: Evidence for a hierarchical coding of location . Journal of Experimental Psychology: Human Perception and Performance , 19 : 451 – 470 .
- Beauchamp , M. S. 2005 . See me, hear me touch me: Multisensory integration in lateral occipital-temporal cortex . Current Opinion in Neurobiology , 15 : 145 – 153 .
- Becchio , C. and Berone , C. 2006 . Abnormal temporal dynamics in unilateral spatial neglect . Neuropsychologia , 44 : 2775 – 2782 .
- Bellman , A. , Meuli , R. and Clarke , S. 2001 . Two types of auditory neglect . Brain , 124 : 676 – 687 .
- Bender , M. B. and Diamond , S. P. 1965 . An analysis of auditory perceptual defects with observations on the localization of the dysfunction . Brain , 88 : 675 – 686 .
- Bisiach , E. 1993 . Mental representation in unilateral neglect and related disorders: The twentieth Bartlett Memorial Lecture . Quarterly Journal of Experimental Psychology , 46A : 435 – 461 .
- Bisiach , E. , McIntosh , R. D. , Dijkerman , C. , McClements , K. I. , Colombo , M. and Milner , D. 2004 . Visual and tactile length matching in spatial neglect . Cortex , 40 : 651 – 657 .
- Bisiach , L. , Cornacchia , R. and Vallar , G. 1984 . Disorders of perceived auditory lateralisation after lesions of the right hemisphere . Brain , 107 : 37 – 52 .
- Blauert , J. and Lindemann , W. 1986 . Spatial mapping of intracranial auditory events for various degrees of interaural coherence . Journal of the Acoustic Society of America , 79 : 806 – 813 .
- Bueti , D. , Costantini , M. , Forster , B. and Aglioti , S. M. 2004 . Uni- and cross-modal temporal modulation of tactile extinction in right brain damaged patients . Neuropsychologia , 42 : 1689 – 1696 .
- Bueti , D. , Bahrami , B. and Walsh , V. 2008a . Sensory and association cortex in time perception . Journal of Cognitive Neuroscience , 20 : 1045 – 1062 .
- Bushara , K. O. , Weeks , R. A. , Ishii , K. , Catalan , M.-J. and Tian , B. 1999 . Modality specific frontal and parietal areas for auditory and visual spatial localization in humans . Nature Neuroscience , 2 : 759 – 765 .
- Caramazza , A. and Badecker , W. 1989 . Patient classification in neuropsychological research . Brain and Cognition , 10 : 256 – 295 .
- Cate , A. and Behrmann , M. 2002 . Spatial and temporal influences on extinction . Neuropsychologia , 40 : 2206 – 2225 .
- Collett , D. 2002 . Modeling binary data , 2nd , Boca Raton, FL : Chapman & Hall/CRC Press .
- Coulthard , E. , Parton , A. and Husain , M. 2007 . The modular architecture of the neglect syndrome: Implications for action control in visual neglect . Neuropsychologia , 45 : 1982 – 1984 .
- Culling , J. F. , Summerfield , A. Q. and Marshall , D. H. 1994 . Effects of simulated reverberation on the use of binaural cues and fundamental-frequency differences for separating concurrent vowels . Speech Communication , 14 : 71 – 95 .
- Cusack , R. , Carlyon , R. P. and Robertson , I. H. 2000 . Neglect between but not within auditory objects . Journal of Cognitive Neuroscience , 12 : 1056 – 1065 .
- Danckert , J. , Ferber , S. , Pun , C. , Broderick , C. , Stiemer , C. Rock , S. 2007 . Neglected time: Impaired temporal perception of multisecond intervals in unilateral neglect . Journal of Cognitive Neuroscience , 19 : 1706 – 1720 .
- Davis , A. 1995 . Hearing in adults: The prevalence and distribution of hearing impairment and reporting hearing disability in the MRC Institute of Hearing Research's national study of hearing , London : Whurr Publishers .
- De Renzi , E. , Gentilini , M. and Barbieri , C. 1989 . Auditory neglect . Journal of Neurology, Neurosurgery and Psychiatry , 52 : 613 – 617 .
- De Renzi , E. , Gentilini , M. and Pattachini , F. 1984 . Auditory extinction following right hemisphere damage . Neuropsychologia , 22 : 733 – 744 .
- Di Pellegrino , G. , Basso , G. and Frassinetti , F. 1997 . Spatial extinction on double asynchronous stimulation . Neuropsychologia , 35 : 1215 – 1223 .
- Driver , J. and Spence , C. J. 1994 . “ Spatial synergies between auditory and visual attention ” . In Attention and Performance XVI: Information integration in perception and communication , Edited by: Inui , T. and McClelland , J. L. 209 – 235 . Cambridge, MA : MIT Press .
- Duncan , J. , Bundesen , C. , Olson , A. , Humphreys , G. , Chavda , S. and Shibuya , H. 1999 . Systematic analysis of deficits in visual attention . Journal of Experimental Psychology: General , 128 : 450 – 478 .
- Eramudugolla , R. , Irvine , D. R. F. and Mattingley , J. B. 2007 . Association of auditory and visual symptoms of unilateral neglect . Neuropsychologia , 45 : 2631 – 2637 .
- Farah , M. J. , Wong , A. B. , Monheit , M. A. and Morrow , L. A. 1989 . Parietal lobe mechanisms of spatial attention: Modality specific or supramodal? . Neuropsychologia , 27 : 461 – 470 .
- Finney , D. J. 1964 . Probit analysis: Statistical treatment of the sigmoid curve , London : Cambridge University Press .
- Frassinetti , F. , Pavani , F. and Ladavas , E. 2002 . Acoustical vision of neglected stimuli: Interaction among spatially converging audiovisual inputs in neglect patients . Journal of Cognitive Neuroscience , 14 : 62 – 69 .
- Ghazanfar , A. A. and Schroeder , C. E. 2006 . Is neocortex essentially multisensory? . Trends in Cognitive Sciences , 10 : 276 – 285 .
- Gifford , G. W. and Cohen , Y. E. 2004 . Effect of a central fixation light on auditory responses in area LIP . Journal of Neurophysiology , 91 : 2929 – 2933 .
- Golay , L. , Hauert , C. A. , Greber , C. , Schnider , A. and Ptak , R. 2005 . Dynamic modulation of visual detection by auditory cues in spatial neglect . Neuropsychologia , 43 : 1258 – 1265 .
- Halligan , P. W. , Fink , G. R. , Marshall , J. C. and Vallar , G. 2003 . Spatial cognition: Evidence from visual neglect . Trends in Cognitive Sciences , 7 : 125 – 133 .
- Hill , N. I. and Darwin , C. J. 1996 . Lateralisation of a perturbed harmonic: Effects of onset asynchrony and mistuning . Journal of the Acoustical Society of America , 100 : 2352 – 2364 .
- Hukin , R. W. and Darwin , C. J. 1995 . Comparison of the effect of onset asynchrony on auditory grouping in pitch matching and vowel identification . Perception and Psychophysics , 52 : 91 – 196 .
- Karnath , H.-O. 2001 . New insights into the functions of the superior temporal cortex . Nature Neuroscience , 2 : 568 – 576 .
- Karnath , H.-O. , Zimmer , U. and Lewald , J. 2002 . Impaired perception of temporal order in auditory extinction . Neuropsychologia , 40 : 1977 – 1982 .
- Kim , M.-S. and Cave , K. R. 2001 . Perceptual grouping via spatial selection in a focussed attention task . Vision Research , 41 : 611 – 624 .
- Kinsbourne , M. 1993 . “ Orientation bias model of unilateral neglect: Evidence from attentional gradients within hemispace ” . In Unilateral neglect, clinical and experimental studies , Edited by: Robertson , I. H. M. 63 – 86 . Hove, , UK : Psychology Press .
- Kramer , A. F. , Weber , T. A. and Watson , S. E. 1997 . Object-based attentional selection—grouped arrays or spatially invariant representations? Comment on Vecera and Farah (1994) . Journal of Experimental Psychology: General , 126 : 3 – 13 .
- Lamy , D. and Tsal , Y. 2000 . Object features, object locations and object files: Which does selective attention activate and when? . Journal of Experimental Psychology: Human Perception and Performance , 26 : 1387 – 1400 .
- Lewis , J. W. , Beauchamp , M. S. and DeYoe , E. A. 2000 . A comparison of visual and auditory motion processing in human cerebral cortex . Cerebral Cortex , 10 : 873 – 888 .
- Macaluso , E. , Frith , C. D. and Driver , J. 2000 . Selective spatial attention in vision and touch: Unimodal and multimodal mechanisms revealed by PET . Journal of Neurophysiology , 63 : 3062 – 3085 .
- Macaluso , E. , Frith , C. D. and Driver , J. 2001 . Multimodal mechanisms of attention related to rates of spatial shifting in vision and touch . Experimental Brain Research , 137 : 445 – 454 .
- Maier , J. X. , Neuhoff , J. G. , Logothetis , N. K. and Ghazanfar , A. A. 2004 . Multisensory integration of looming signals in rhesus monkeys . Neuron , 43 : 177 – 181 .
- Marshall , J. C. , Halligan , P. W. and Robertson , I. H. 1993 . “ Contemporary theories of unilateral neglect: A critical review ” . In Unilateral neglect: Clinical and experimental studies , Edited by: Robertson , I. H. and Marshall , J. C. 311 – 329 . Hove, , UK : Psychology Press .
- McCullagh , P. and Nelder , J. A. 1983 . Generalized linear models , London : Chapman & Hall .
- McDonald , J. B. and Ward , M. 1999 . Spatial relevance determines facilitatory and inhibitory effects of auditory covert spatial orienting . Journal of Experimental Psychology: Human Perception and Performance , 25 : 1234 – 1252 .
- Mesulam , M. M. 1981 . A cortical network for directed attention and unilateral neglect . Annals of Neurology , 10 : 309 – 325 .
- Mesulam , M. M. 1999 . Spatial attention and neglect: Parietal, frontal and cingulated contributions to mental representation and attentional targeting of salient extrapersonal events . Philosophical Transactions of the Royal Society of London. Series B, Biological Sciences , 354 : 1325 – 1346 .
- Parton , A. , Malhotra , P. and Husain , M. 2004 . Hemispatial neglect . Journal of Neurosurgery and Psychiatry , 75 : 13 – 21 .
- Pavani , F. , Husain , M. , Ladavas , E. and Driver , J. 2004 . Auditory deficits in visuospatial neglect patients . Cortex , 40 : 347 – 365 .
- Pavani , F. , Meneghello , F. and Ladavas , E. 2001 . Deficit of auditory space perception in patients with visuospatial neglect . Neuropsychologia , 39 : 1401 – 1409 .
- Pavani , F. , Lavadas , E. and Driver , J. 2002 . Selective deficit of auditory localisation in patients with visuospatial neglect . Neuropsychologia , 40 : 291 – 301 .
- Pavani , F. , Ladavas , E. and Driver , J. 2003 . Auditory and multisensory aspects of visuospatial neglect . Trends in Cognitive Sciences , 7 : 407 – 414 .
- Pinek , B. , Duhamel , J. , Cave , C. and Brouchon , M. 1989 . Audiospatial deficits in humans: Differential effects associated with left versus right hemisphere parietal damage . Cortex , 25 : 175 – 186 .
- Pouget , A. and Driver , J. 2000 . Object-centered visual neglect, or relative egocentric neglect? . Journal of Cognitive Neuroscience , 12 : 542 – 545 .
- Rizzolatti , G. , Scandolara , C. , Matelli , M. and Gentilucci , M. 1981 . Afferent properties of periarcuate neurons in macaque monkeys . Behavioural Brain Research , 2 : 147 – 163 .
- Roberts , K. L. , Summerfield , A. Q. and Hall , D. A. 2006 . Presentation modality influences behavioural measures of alerting, orienting, and executive control . Journal of the International Neuropsychological Society , 12 : 485 – 492 .
- Rorden , C. , Mattingley , J. B. , Karnath , O.-H. and Driver , J. 1997 . Visual extinction and prior entry: Impaired perception of temporal order with intact motor perception after unilateral parietal damage . Neuropsychologia , 35 : 421 – 433 .
- Shallice , T. 1988 . From neuropsychology to mental structure , Cambridge, , UK : Cambridge University Press .
- Shore , D. I. and Spence , C. S. 2005 . “ Prior entry ” . In Neurobiology of attention , Edited by: Itti , L. , Rees , G. and Tsotsos , J. K. 89 – 95 . Burlington, MA : Elsevier Academic Press .
- Siegel , S. and Castellan , N. J. 1988 . Nonparametric statistics for the behavioral sciences , New York : McGraw-Hill .
- Sinnett , S. , Juncadella , M. , Rafal , R. D. , Azanon , E. and Soto-Faraco , S. 2007 . A dissociation between visual and auditory hemi-inattention: Evidence from temporal order judgements . Neuropsychologia , 45 : 552 – 560 .
- Soroker , N. , Calamaro , N. , Glicksohn , J. and Myslobodsky , M. S. 1997 . Auditory inattention in right-hemisphere-damaged patients with and without visual neglect . Neuropsychologia , 35 : 249 – 256 .
- Spence , C. and Driver , J. 1997 . Audiovisual links in exogenous covert spatial orienting . Perception and Psychophysics , 59 : 1 – 22 .
- Spence , C. , Lloyd , D. , McGlone , F. , Nicholls , M. E. R. and Driver , J. 2000 . Inhibition of return is supramodal: A demonstration between all possible pairings of vision touch and audition . Experimental Brain Research , 134 : 42 – 48 .
- Spierer , L. , Meuli , R. and Clarke , S. 2007 . Extinction of auditory stimuli in hemineglect: Space versus ear . Neuropsychologia , 45 : 540 – 551 .
- Vallar , G. 2001 . Extrapersonal visual unilateral spatial neglect and its neuroanatomy . NeuroImage , 14 : S52 – S58 .
- Wenzel , E. M. , Arruda , M. , Kistler , D. J. and Wightman , F. L. 1993 . Localization using nonindividualized head-related transfer functions . Journal of the Acoustical Society of America , 94 : 111 – 123 .
- Wightman , F. L. and Kistler , D. J. 1989 . Headphone simulation of free-field listening. I: Stimulus synthesis . Journal of the Acoustical Society of America , 85 : 858 – 867 .
- Wilson , B. , Cockburn , J. and Halligan , P. W. 1987 . Behavioural Inattention Test , Bury St Edmonds, , UK : Thames Valley .
- Woods , W. A. and Colburn , S. 1992 . Test of a model of auditory object formation using intensity and interaural time difference discriminations . Journal of the Acoustical Society of America , 91 : 2894 – 2902 .
- Zimmer , U. , Lewald , J. and Karnath , H.-O. 2003 . Disturbed sound lateralization in patients with spatial neglect . Journal of Cognitive Neuroscience , 15 : 694 – 703 .